Abstract
The present study describes how flour and phenolic enriched extracts (PEE) are obtained from seed (food waste) of 10 different P. alba (algarrobo blanco) clones and their characterization to be used as non-conventional sources of potential functional ingredients. Seed flour and PEE obtained from Argentinian P. alba cultivars were chemically characterized. The antioxidant capacity was also determined. The results showed variability in macronutrient composition of seed flour obtained from different clones. Among them, seed flour obtained from P4, P5, P6, P10, P12, and P13 clones showed a higher protein and fiber content than the other clones. On the other hand, PEE obtained from P6, P7, and P10 clones showed the highest content of phenolic component (7.32–8.58 mg GAE/g flour). The extracts obtained from them also showed high antioxidant activity (scavenging activity on ABTS•+, HO•, and H2O2). C-glycosyl flavones, including vicenin II, isoschaftoside, schaftoside, vitexin, and isovitexin were the major components extracted in all clones. These compounds have remarkable properties for disease prevention linked to oxidative stress. Therefore, the P. alba seed could be considered as functional food waste with a great potential to be used as a novel renewable and sustainable material for the production of bioactive food formulations.
1. Introduction
Prosopis includes 45 species that belong to the Mimosoideae subfamily within the Fabaceae family, distributed in America, Asia, and Africa [1]. Prosopis grows easily in tropical, subtropical, and semi-arid soils, enduring high temperatures, drought, alkalinity, and soil salinity [2]. In Argentina, there are 27 biotypes of Prosopis spp. [3]. In our country, 100,000 tons of Prosopis wood is harvested annually, to be used in the making of doors, floors, blinds, and windows [4,5]. On the other hand, Prosopis pods have been used since pre-Hispanic times to feed both livestock and human. They are used to prepare drinks (añapa, aloja, and chicha), and numerous foods such as syrups, flour, and sweets (jam, arrope, patay) [6]. In fact, P. alba whole pod flour (Article 681 tris) and P. alba seed flour (Article 681) are included in the Argentine Food Code [7]. Until now, Prosopis flours were obtained from whole pods, although in this process the seed were discarded as waste. The pods flours were evaluated functionally and chemically and its properties, i.e., antioxidant, hypoglycemic, antilipase, and anti-inflammatory activities were demonstrated, but the P. alba seed flours were not yet fully characterized [1,5,8,9,10,11,12,13,14]. The production of P. alba whole pod flour is growing, thus increasing the generation of biomass that should be used. In this sense, an important aspect regarding economic benefit is to recover the high-added-valued constituents present in fruit processing wastes. The availability of this vegetable raw material, together with the problem represented by the accumulation of waste and its unexploited potential, has encouraged researchers to develop projects on the value-addition potential of fruit processing waste, such as enrichment in food formulation.
One of the main difficulties faced by the industrialization of P. alba flour is its high variability, for both biomass and pod production [15]. Regarding progenies, this happens because Prosopis are pollinated by insects and are self-incompatible. Therefore the seeds of the same tree are highly likely to have multiple crosses. In this way, clonal propagation is an essential tool for the genetic improvement of P. alba. Fortunately, it can be propagated clonally by grafting, rooting the cuttings, and air layering [15]. Clonal selections of P. alba have been made for its economically valuable characteristics, such as rapid growth rate, high pod production, and improved flavor (sweeter pods without bitter characteristics) [15,16]. Progeny trials of elite trees have been conducted in Santiago del Estero (27°45′ S, 64°15′ W) with seeds of 57 individual trees from 8 regions of Argentina. The initial selection was based on productivity (biomass) and pod taste (sweet or very sweet, astringent or very astringent). The plants that produced sweet or very sweet pods were cloned by grafting stems from mature trees. The rejuvenated clones (10) were multiplied by rooting cuttings [15,16,17].
The present study describes for the first time the chemical characterization and antioxidant capacity of flour and phenolic enriched extracts (PEE) obtained from seeds of previously selected P. alba clones.
2. Materials and Methods
2.1. Plant Material
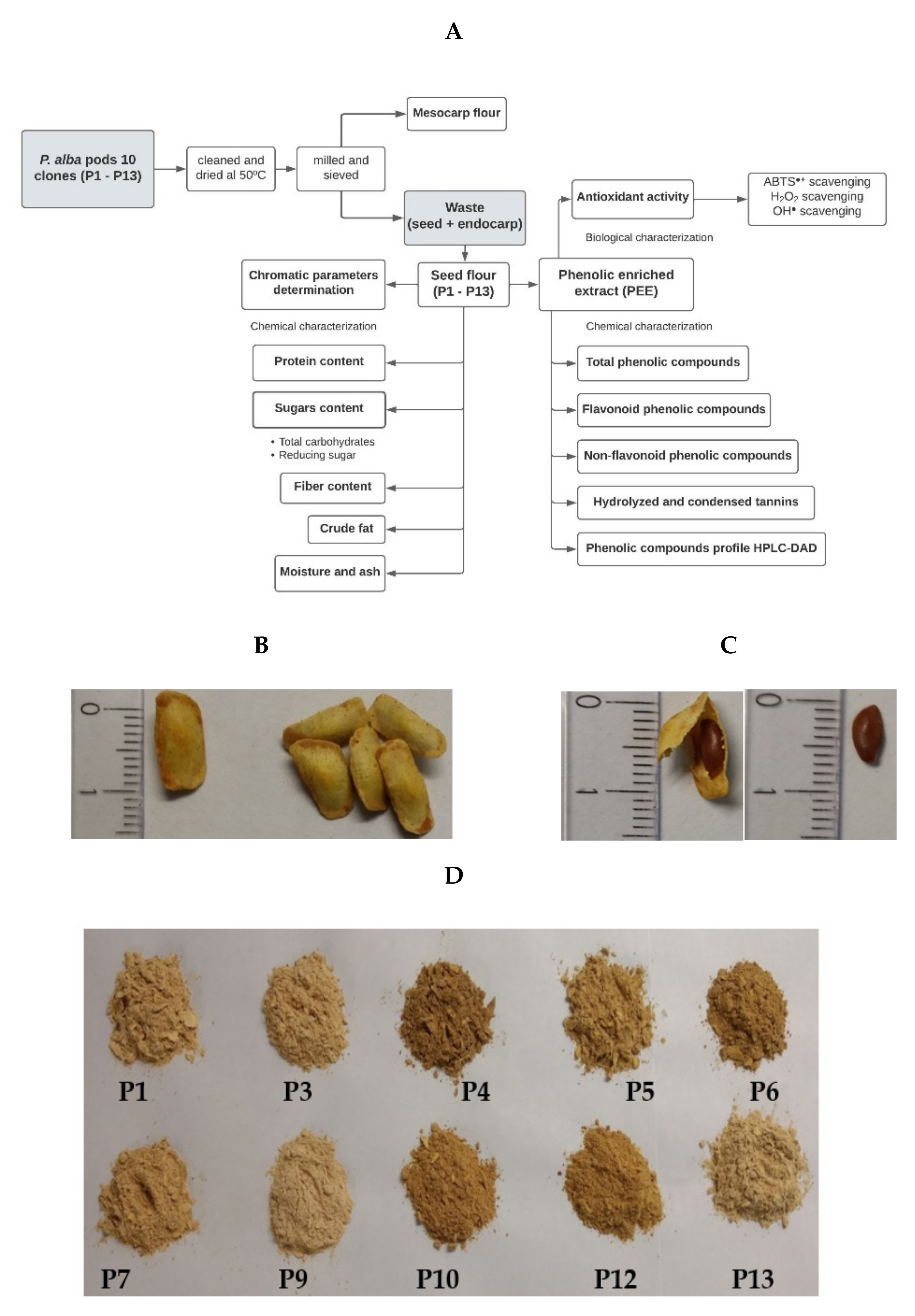
The mature pods of 10 clones of Prosopis alba were collected in an experimental field of the Universidad Católica de Santiago del Estero in a town called Fernández, in the province of Santiago del Estero, Argentina. Figure 1A shows a summary of the methodology.
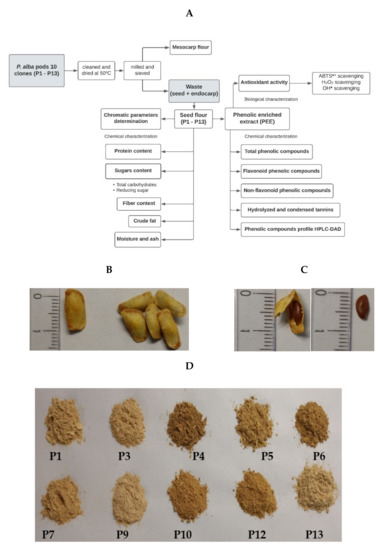
Figure 1.
(A) Flowchart of the methodology. (B) Seeds with endocarp. (C) Seeds and (D) flour of seeds from 10 clones of Prosopis alba (P1–P13).
2.2. P. alba Seed Flour Preparation and Chromatic Parameter Determination
The pods with the same ripening stage were cleaned so as to remove any strange material and dried in a forced-air oven at 50 °C until reaching constant weight. The pods were named P1 to P13. The pods P2, P8, and P11 were discarded because they had abundant polluting plant material. The dried pods were milled and then sieved to separate both the mesocarp flour and whole seeds with endocarp (considered as waste). The seeds (Figure 1B,C) were processed with a knife mill to obtain the fine flour (≤149 μm) of each sample (seed flours P1–P13), as shown in Figure 1D. The seed flour was stored under vacuum at −20 °C until use. The chromatic parameters of the flour were measured with a Chroma meter CR-400 (Konica Minolta, Tokyo, Japan) colorimeter using the CIELab system, the results being expressed as chromaticity coordinates L*, a*, and b* (objective parameter). The coordinated L* represents lightness (contribution of black or white varying between 0 and 100); a* represents the contribution of green or red (negative or positive), and b* represents the contribution of yellow or blue (negative or positive) [18].
2.3. Determination of Chemical Composition
2.3.1. Macronutrient Determination
A differential extraction from P. alba seed flour was carried out to obtain proteins and sugars [19]. Crude protein content was determined by using the Kjeldahl method [20]. Total carbohydrates and reducing sugar were measured using the method of Orqueda et al., 2017 [21]. Crude fat was determined with a Soxhlet apparatus, extracted with petroleum ether (40–60 °C) for 4 h according to AOCS, 1989 [22]. Results were expressed as mg of macronutrient/g of flour. The moisture content was determined by gravimetric method between the fresh and the dry samples [23]. The latter were incinerated in a muffle (500 °C) and, then, the ashes were weighed on an analytical scale to determine the ash content [24].
2.3.2. Fiber Analysis
Fiber content was determined by using acid and alkaline digestion and heating [19]. Crude fibers were calculated as the difference in weight before and after incineration in muffle. Results were expressed as g of fiber/100 g of flour.
2.3.3. Phenolic Enriched Extracts: Characterization
Five g of P. alba seed flour was mixed with 50 mL of 70° ethanol in an ultrasonic bath for 10 min at 25 °C. Then, the samples were placed in a shaker for 24 h at 25 °C and 120 rpm. Then, each sample was vacuum filtered to obtain the phenolic enriched extract (PEE). Then, the PEE was dried under reduced pressure at 40 °C and freeze-dried. The powder obtained was stored at −20 °C until use. Total phenolic compounds content was determined by using Folin-Ciocalteu reagent (Merck, Darmstadt, HE, Germany) [25] and was expressed as mg of gallic acid equivalent (GAE)/g flour.
Total non-flavonoid phenolic compounds were determined by using the method described by Isla et al. (2014) [26]. First, flavonoids were precipitated with formaldehyde (8 g/L). Then, the samples were centrifuged at 9000× g for 5 min (Rolco, Buenos Aires, Argentina), and non-flavonoid phenolic compounds were quantified in the supernatant with the Folin–Ciocalteau reagent. Flavonoid phenolic compounds were calculated by using the difference between the total phenolic and non-flavonoid phenolic content. Flavone and flavonol content were determined by using the aluminum chloride colorimetric method [21]. Hydrolyzed and condensed tannins were extracted and quantified as described by Costamagna et al. (2013) [19].
2.3.4. Profile of Phenolic Compounds Using HPLC-DAD
The chromatographic profiles of PEE were obtained by HPLC coupled to a diode array detector (HPLC-DAD) (Waters Corporation, Milford, MA, USA) in an analytical C18 column (XBridge) by using a linear gradient solvent system consisting of 0.1% acetic acid in water (A) and 0.1% acetic acid in methanol as follows: 90% to 43% A for 45 min, 43% to 0% A for 45 to 60 min, holding 100% B for 5 min. The injected sample volume was 20 µL with a flow rate of 0.5 mL/min. The identification of phenolic compounds was carried out by comparing the retention times and spectral data (220–600 nm) of each peak with those of standards (HPLC quality) from Sigma-Aldrich (St. Louis, MO, USA) and Fluka Chemical Corp. (Ronkonkoma, NY, USA).
2.4. Antioxidant Activity
2.4.1. ABTS●+ Scavenging Activity
ABTS●+ scavenging activity was measured [27] on a microplate reader by using 200 µL of ABTS●+ solution and 100 µL of sample to final concentrations between 2 and 10 µg GAE/mL of phenolic compounds. Absorbance was measured at 1 and 6 min at 734 nm. The sample concentration required to scavenge 50% ABTS●+ was determined as 50% ABTS●+ scavenging capacity (SC50). Results are expressed in µg gallic acid equivalent (GAE)/mL of PEE.
2.4.2. Hydrogen Peroxide Scavenging Activity
Hydrogen peroxide (H2O2) scavenging activity of PEE obtained from seed flours was carried out through the optimized enzymatic colorimetric assay [28] adapted to a microplate assay. Briefly, different sample concentrations diluted in 84 mM sodium phosphate buffer (pH7) reacted with H2O2, phenol, and 4-aminoantipyrine reaction system in the presence of horseradish peroxidase (HRP), at 37 °C. This reaction was measured at 504 nm. SC25 values denote the sample concentration required to scavenge 25% H2O2. Results are expressed in µg gallic acid equivalent (GAE)/mL.
2.4.3. Hydroxyl Radical Scavenging Activity
The hydroxyl radical scavenging activity of PEE was determined according to Cattaneo et al. (2016) [10]. The reaction mixture contained 2-deoxy-D-ribose/FeCl3 and different extract concentrations (up to 20 µg GAE/mL) with and without EDTA. The reaction was started with H2O2 and ascorbic acid at 37 °C for 60 min. Then, 2-thiobarbituric acid was added and incubated for 20 min at 100 °C. Absorbance was recorded at 532 nm. Results are presented as SC50 values in µg GAE/mL required to inhibit the degradation of 50% 2-deoxy-D-ribose. Quercetin (5–50 µg/mL) was used as a reference compound.
2.5. Statistical and Chemometric Analysis
The comparison of mean values was made by ANOVA, followed by a Tukey post-test. A value of p ≤ 0.05 was considered significant.
The chemical components concentration and the SC50 values of free radicals by several methodologies for each flour sample were exported in a matrix of 14 columns and 30 rows, where each column represents a variable and each row represents a clone. Principal component analysis (PCA) and hierarchical clusters (HCA) using the Infostat software version 2015 [29] (Universidad Nacional de Córdoba, Córdoba, Argentina) were applied. PCA scaling and centering were used. Mean values were used for graphs, where each point on the graph corresponded to the mean value for each clone. HCA was performed by calculating the Euclidean distance and considering average link clustering as a method for calculating the distance between clusters. In addition, data was standardized. The number of groups was determined considering the cut-off line half the Euclidean distance.
3. Results and Discussion
Clones of P. alba were identified in a progeny trial established in 1990 in Santiago del Estero (Argentina) [15]. The trees showed rapid growth, high pod production, and sweet pods. All clones had good sensory properties for use in human food [15,16]. The pods of 10 clones were milled to obtain mesocarp flour. The discarded seeds in the pod milling process were submitted to grinding to obtain seed flour. Seed flour chromatic values were determined to compare all clones. The values were L* = 58.41 ± 10 to 60.40 ± 10; a* = 3.42 ± 1 to 8.38 ± 1 and b* = 33.43 ± 2 to 35 ± 2. Similar results were obtained to P. alba mesocarp flour [11]. The flours P4, P5, P6, P10, and P12 showed the highest values of a* (8.38; 7.20; 7.32; 7.30; 7.10, respectively), as shown in Figure 1D. This parameter showed the highest chromatic variability between clones.
3.1. Macronutrients
The carbohydrate content of seed flour exhibits the total carbohydrate content between 5.16 g/100 g and 23.15 g/100 g. P6, P7, P9, and P10 showed the highest sugar content. The total sugar content of seed flour was lower than that of mesocarp flour (73.95 g/100 g), data reported by Cardozo et al., 2010 [1] (Table 1). The mesocarp of pods belonging to Prosopis species is rich in sugars [5,13,30] and the seed is rich in protein [9,10,30]. Besides, P10 showed the highest content of reducing sugar.

Table 1.
Proximal composition of the seed flour from 10 clones of Prosopis alba.
The protein content (Table 1) of P. alba seed flour varied between 7.98 and 13.34 g/100 g. P4 and P5 clone seed flours showed even greater total protein content than P. alba mesocarp flour [1]. Protein values were similar to amaranth seed, with values between 13.2 and 18.2% [31]. According to guidelines for the use of nutrition and health claims of the Codex Alimentarius of FAO (Food and Agricultural Organization, 2011) [32], when a solid sample has a protein content ≥10% it could be considered a source of protein. All P. alba seed flours showed this condition; only P3 and P9 clones exhibited less than 10% protein values. Fat values were lower in seed flour (1.70 to 3.98 g/100 g) than in mesocarp flour (11.3 g/100 g) [1]. P4–P6 clones showed the highest fat content (Table 1).
Crude dietary fiber content varied between 13.90 and 27.69 g/100 g. P1, P4, and P5 showed the highest crude fiber content. The deficiency of dietary fiber in the human diet has been associated with constipation, diverticulosis, cardiovascular diseases, and cancer, so its study is important [33]. In addition, some P. alba seed flours had a higher fiber content than that reported for mesocarp and cotyledon flour of P. alba [1,10]. Ash determines the nutritional quality of food, affecting the rheological properties and cooking quality [34]. P4 flour showed the highest ash content (4.25%).
The low moisture content (less than 10%) would improve its storage stability by preventing mold growth and other biochemical reactions and extending the final product shelf life. All seed flours have this condition, except for P10 and P12 (11.77% and 12.50%, respectively).
3.2. Secondary Metabolites
The secondary metabolites play an important role in the defense systems of plants against environmental stress and pathogenic attacks. In addition, these compounds have various biological activities, so they are used as medicinal, food, and cosmetic ingredients [35]. There are some studies regarding polyphenols in P. alba cotyledons, seeds, and mesocarp flour [8,10,12,13,14]. Table 2 shows the phytochemical composition of 10 clones of P. alba. The total phenolic compound content extracted from P. alba seed flour was between 5.05 and 8.58 mg GAE/g of flour. These values are higher than that obtained by Sciammaro et al. (2016) [13] (3 mg GAE/g DW) for P. alba seeds collected in Ingeniero Juárez, Formosa in the northeast of Argentina, but were lower than the total phenolic content of P. alba cotyledon flour (11 mg GAE/g of flour) reported by Cattaneo et al., (2016) [10] for P. alba collected in Amaicha del Valle (Tucumán, Argentina). Flours of P6, P7, and P10 clones showed the highest phenolic content, higher than P. alba mesocarp flour (4.6 mg GAE/g flour), [8,14] and Ceratonia siliqua pulp flour [36]. Non-flavonoid phenolic compounds were the main phenolic components of P. alba seed flour (2.68 to 4.01 mg GAE/g of flour). P10 flour showed the highest flavones and flavonol content. The flavonoid content was between 0.098 and 0.308 mg of quercetin equivalent (QE)/g flour (mg) and was lower than those found in P. alba cotyledons flour (3.9 mg EQ/g flour) and mesocarp flour (33–87.5 mg EQ/g flour) [1,8,10,14].
Table 2.
Phenolic components of the seed flour from 10 clones of Prosopis alba.
Regarding the content of condensed tannins, the values varied between 44.75 and 223.73 mg of procyanidin B2 equivalent (PB2E)/100 g flour. The highest content was observed in P10, P12, and P13 clones. P7 presented a similar content of these compounds to that reported to P. alba cotyledon flours (175 mg PB2E/100 g flour) [10].
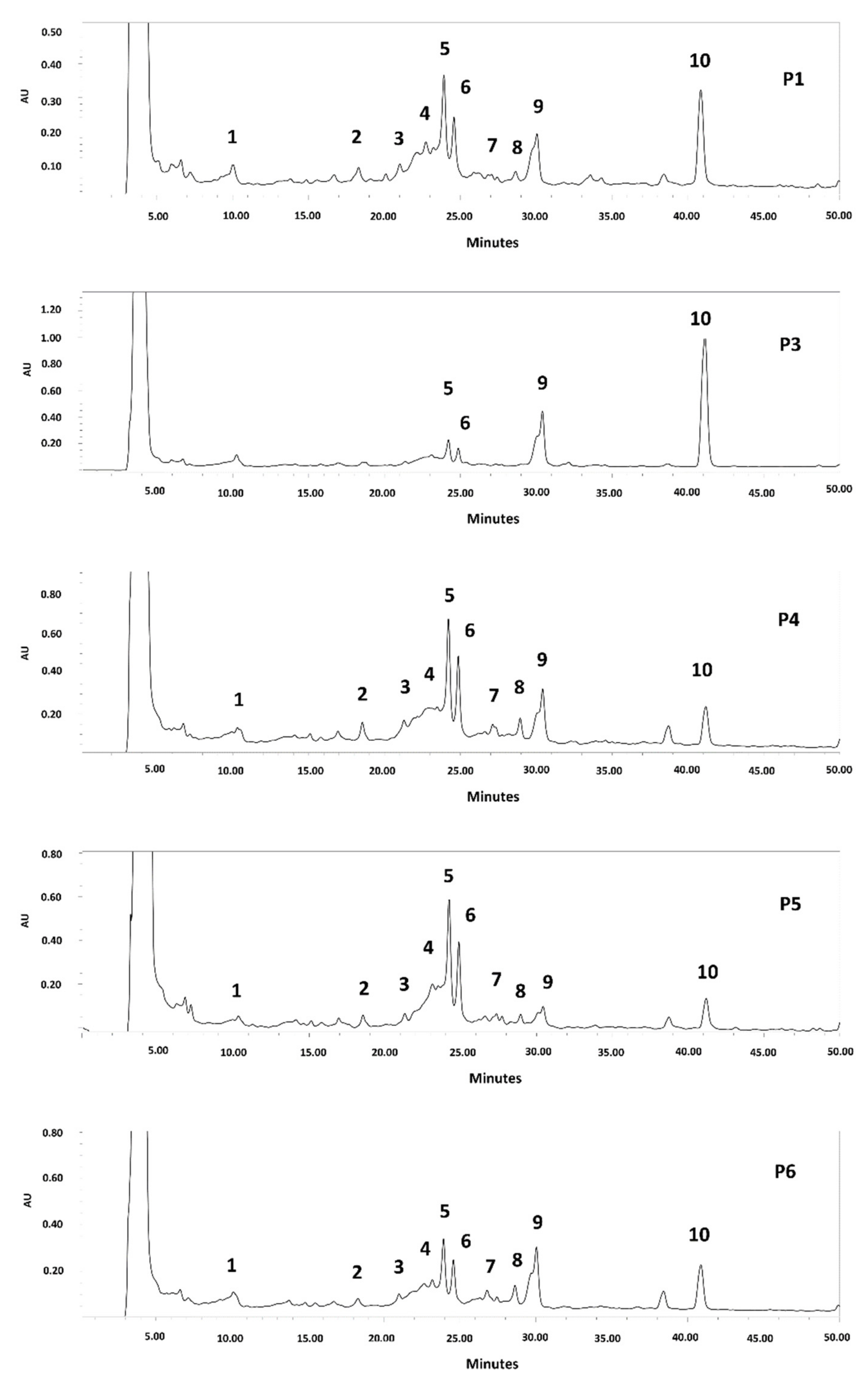
3.3. HPLC Analysis
Since an extract rich in polyphenols was obtained from the seeds of all the clones, it was decided to determine their phenolic compound profile. The HPLC chromatogram is shown in Figure 2 and Figure S1 (Supplementary Materials). C-glycosyl flavones, including vicenin II, isoschaftoside, schaftoside, vitexin, and isovitexin, were identified in the seed extracts of all analyzed clones. The same results were reported for P. alba and P. nigra mesocarp flour [8,11,12,13,14,37,38] and P. nigra seed [39]. Other authors have reported several biological activities for apigenin-C glycosides, such as antioxidant, anti-inflammatory, anti-cancer, anti-platelet activity, angiotensin converting enzyme inhibition, protective effects against neurological and psychiatric diseases, protective activity in the cardiovascular system, protective effects against diseases of the endocrine system and metabolic, anti-microbial, and antiviral effects, among others [38,39,40]. In addition, they are rapidly absorbed after oral administration and are distributed by plasma in different tissues [40].
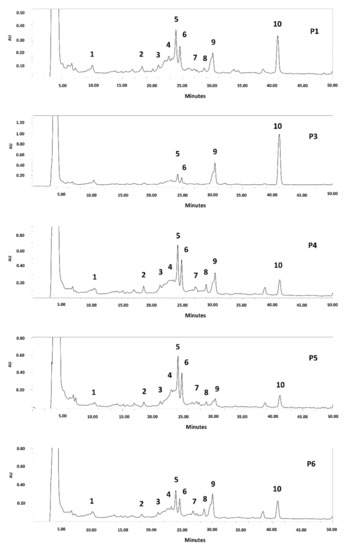
Figure 2.
HPLC chromatogram of polyphenols from P. alba seed flour (Clones P1–P6). Detection: UV254 nm. Compounds: 1: isoschaftoside hexoside; 2: schaftoside hexoside; 3: Vicenin II (apigenin-di-C-hexoside)/isomer; 4: vicenin II/isomer; 5: isoschaftoside (apigenin-C-hexoside-C-pentoside); 6: schaftoside; 7: unknown; 8: vitexin; 9: isovitexin; 10: unknown.
3.4. Biological Activities
This study analyzed the antioxidant activity of PEE obtained from seed flour (Table 3). The samples exhibited ABTS•+ reducing capacity with SC50 values between 4 and 6.7 µg GAE/mL. The antioxidant capacity of PEE obtained from seed flour was higher than P. alba mesocarp flour PEE (13 µg GAE/mL) [11]. The ABTS•+ scavenging activity by the PEE was lower than that of a commercial natural antioxidant such as quercetin (SC50 = 1.4 µg/mL) but similar to BHT, a synthetic antioxidant used in the food industry (SC50 = 3.52 µg/mL).
Table 3.
Antioxidant activity of the seed flour from 10 clones of Prosopis alba.
The PEE of P. alba seed flour also showed scavenging activity of H2O2 with SC25 values between 5.9 and 13.6 for the most active extracts and between 20.5 and 23.3 µg GAE/mL for the less active extracts. Scavenging activity of HO• was also found with SC50 values lower than those found for PEE from P. alba mesocarp flour [10]. Phenolic extract antioxidant activity could be attributed to C-glycosyl flavonoids, compounds with demonstrated antioxidant capacity [8,11,37,38,39,40].
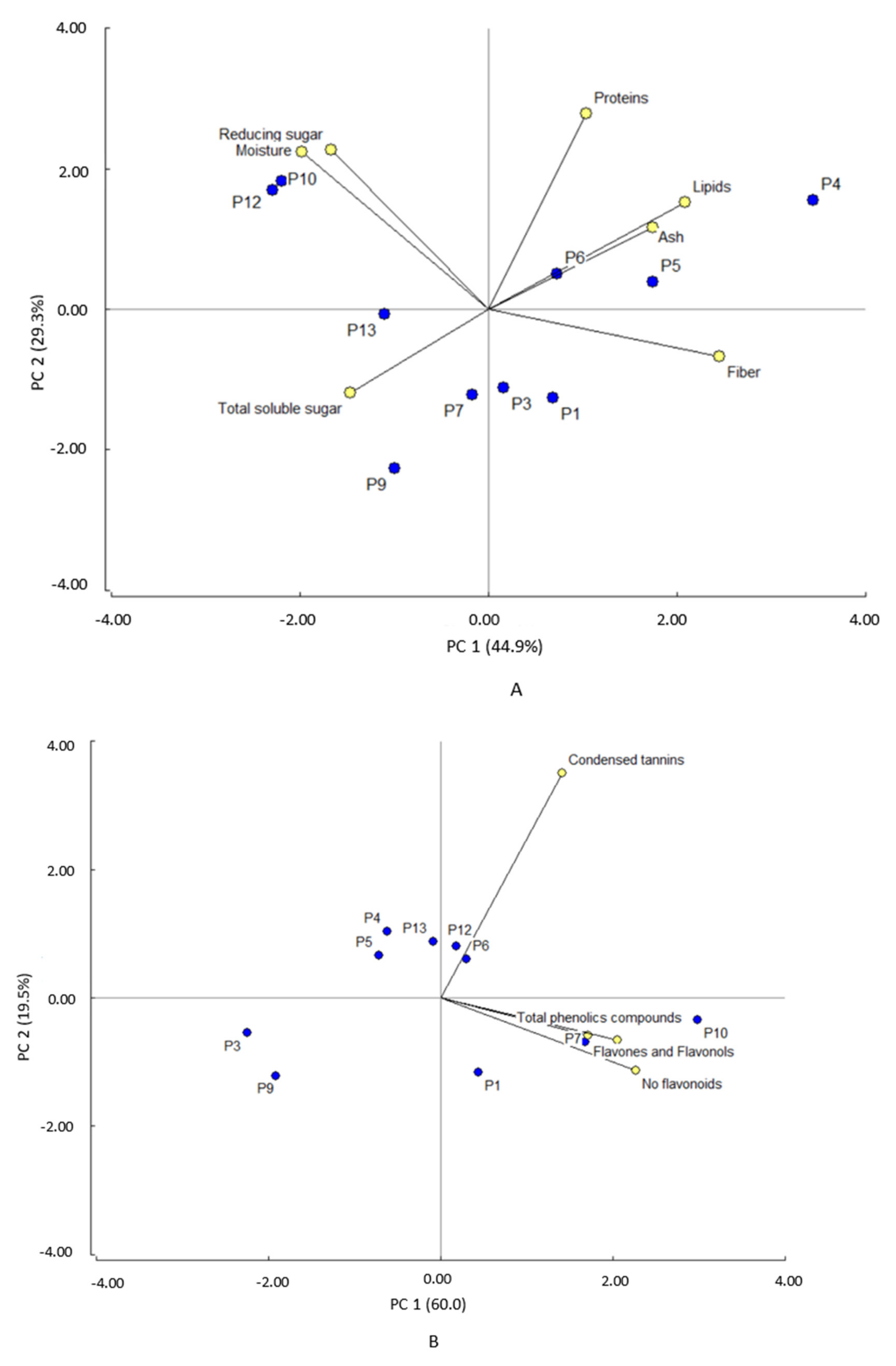
3.5. Principal Component Analysis
Considering the macronutrient components as variables, the principal component analysis (PCA) showed two components that explain 74.2% of the variability (Figure 3A). Principal component 1 (PC1) explains 44.9% of the variability and is positively influenced by fibers and lipids and negatively by moisture. This component clearly separates P4, P5, P6, and P1 clones (+) from P12, P10, P13, and P9 clones (−). The first group showed high fiber and lipid content and low moisture content (Table 1), whereas the second group presented low lipid and fiber level and high moisture content. Principal component 2 (PC2) explains 29.3% of the variability and is influenced by protein and reducing sugars. This component clearly separates P12, P10, and P4 clones from P9, P7, P3, and P1 clones. The former group has higher protein content, reducing sugars and moisture than the latter group. Considering phytochemicals composition as a variable for the PCA analysis, the principal component analysis (Figure 3B) showed that the two components explain 79% of the variability. PC1 explains 60% and is influenced by the content of both non-flavonoid phenolic compounds and total phenolic compounds. This component separates P10 and P7 clones (+) from P9 and P3 clones (−). PC2 explains 19.5% of the variability and is influenced by condensed tannin content. This component separates P1, P9, P3, P7, and P10 clones from P4, P5, P6, P12, and P13 clones. The latter group showed the highest content of condensed tannins.
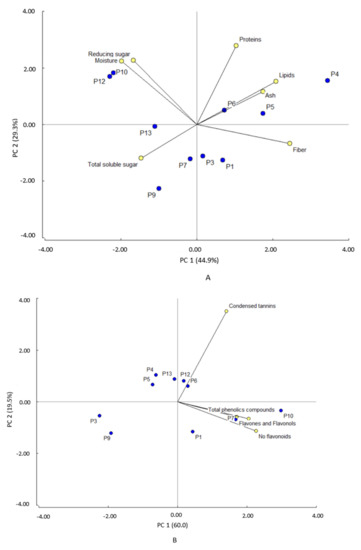
Figure 3.
Principal component analysis for (A) proximal components and (B) phytochemicals.
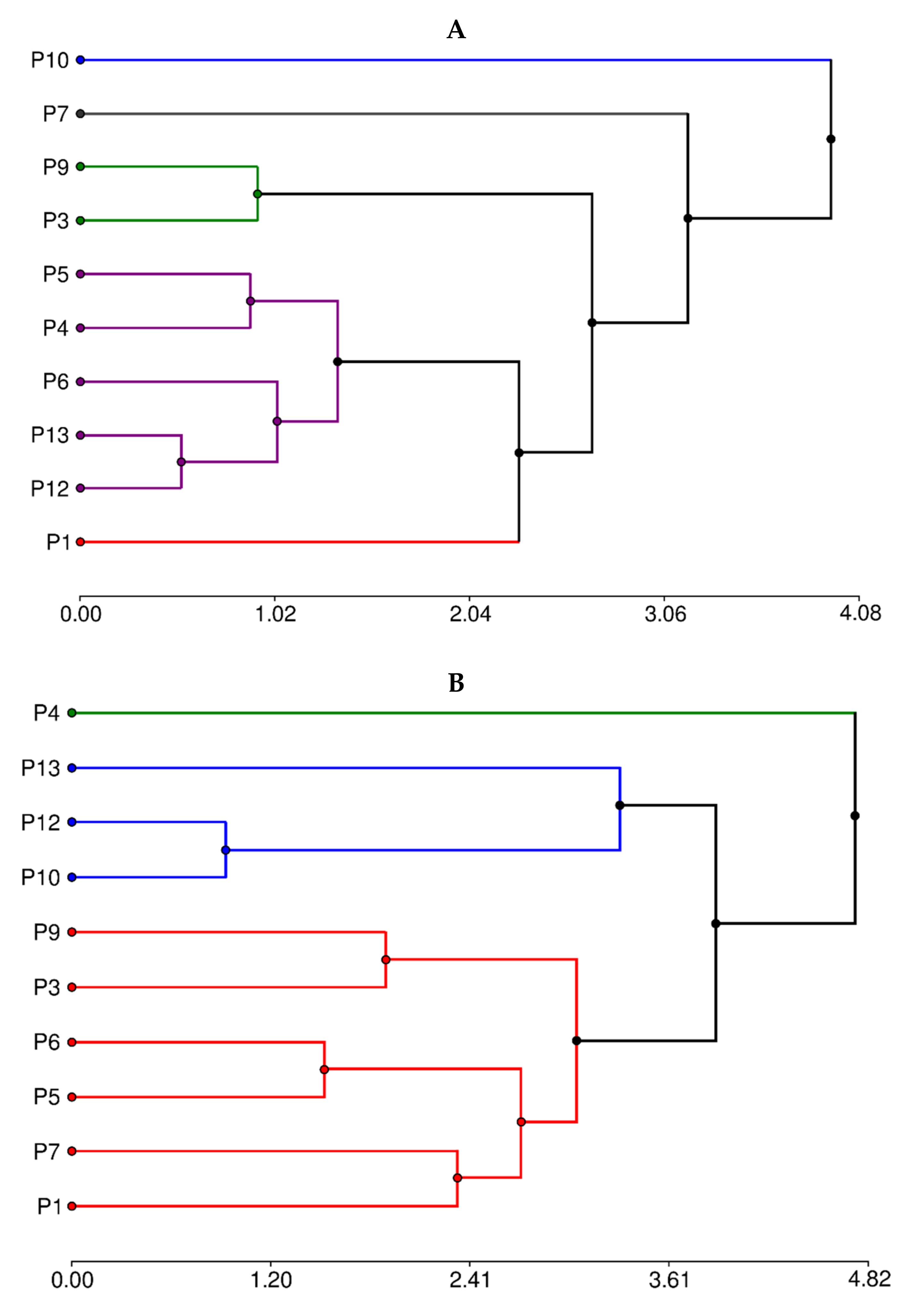
3.6. Hierarchical Cluster Analysis
When attempting to determine the groups of secondary metabolites, two well-marked—besides three individual ones—appear (Figure 4A).
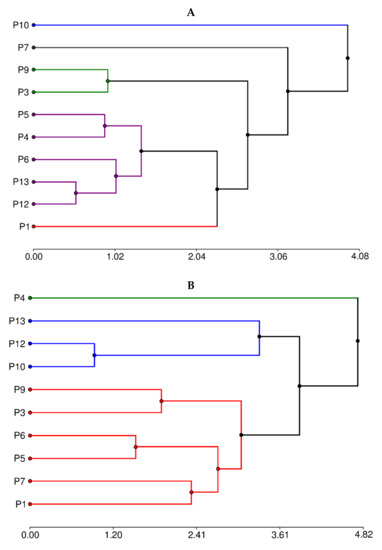
Figure 4.
Hierarchical clusters analysis by (A) secondary metabolites composition and (B) proximal composition.
The group with the highest number of individuals (P4, P5, P6, P12, and P13) is determined by the highest condensed tannin concentration. The second group, P9 and P3, is characterized by the lowest content of all phytochemicals. On the other hand, clones P1, P7, and P10 are each considered to correspond to a different group. P10 showed the highest content in flavones and flavonols, total phenolic, and non-flavonoids. P1 and P7 showed lower values of these compounds concerning P10 but higher than the other groups.
Hierarchical cluster analysis by proximal composition (Figure 4B) suggests three groups. The group formed by P10, P12, and P13 is determined by the low concentration of fiber and lipids. The P4 clone showed the highest lipid, protein, and ash concentration. The third covered the rest of the clones and represented the samples with intermediate values of all components.
4. Conclusions
The results obtained in this study established that principal component analysis and hierarchical cluster analysis were efficient tools for clone selection according to the chemical composition and functional properties of P. alba fruit waste.
Seed flour obtained from P4, P5, P6, P10, P12, and P13 clones showed a higher protein and fiber content than the other clones. On the other hand, PEE obtained from P6, P7, and P10 showed the highest content of phenolic component, mainly C-glycosyl flavones and high antioxidant activity.
Considering the potential use of seed flour for nutritional properties or phytochemical extraction, the selection of adequate clones is possible. Therefore, both the flour and the PEE obtained from P. alba fruit wastes have a great potential to be used as a novel renewable and sustainable material for the production of bioactive food formulations.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/foods11182857/s1, Figure S1: HPLC chromatogram of polyphenols from P. alba seed flour (Clones P7–P13). Detection: UV254 nm. Compounds: 1: isoschaftoside hexoside; 2: schaftoside hexoside; 3: Vicenin II (Apigenin-di-C-hexoside)/Isomer; 4: Vicenin II/Isomer; 5: Isoschaftoside (Apigenin-C-hexoside-C-pentoside); 6: Schaftoside; 7: unknown 8: Vitexin; 9: Isovitexin; 10: unknown.
Author Contributions
Experiments conceived and designed: F.M.C.U., F.C., M.R.A., I.C.Z. and M.I.I. Experiments performed: F.M.C.U., F.C., M.R.A., I.C.Z., L.M.M. and M.I.I. Data analysis: F.M.C.U., F.C., M.R.A., L.M.M., I.C.Z. and M.I.I. Paper written and editing: F.M.C.U., I.C.Z. and M.I.I. Project conceived and initiated: M.I.I. and I.C.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by Universidad Nacional de Tucumán (PIUNT 2018-G637 Project), Agencia Nacional de Promoción Científica y Técnica (ANPCyT PICT 2017-4416 Project: PICT 2020-3619 Project), Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET-PUE 2018-0011 Project) and Biolates network “P320RT0186—Sustainable use of Ibero-American vegetable biomass resources in cosmetics” (CYTED).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is contained within the article or Supplementary Materials.
Acknowledgments
The authors acknowledge the collaboration of Felker P. and Ewens M. for the provision of pods and the financial support from Secretaría de Ciencia, Arte e Innovación Tecnológica (SCAIT-UNT), Argentina, Agencia Nacional de Promoción Científica y Técnica (ANPCyT), Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina, and Programa Iberoamericano de Ciencia y Tecnología para el desarrollo, (CYTED).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
PEE: phenolic enriched extracts, GAE: gallic acid equivalent, QE: quercetin equivalent, PB2E: procyanidin B2 equivalent, HPLC-DAD: HPLC coupled to a diode array detector, SC50: 50% scavenging capacity, HRP: horseradish peroxidase, PCA: principal component analysis, HCA: hierarchical clusters (HCA).
References
- Cardozo, M.L.; Ordoñez, R.M.; Zampini, I.C.; Cuello, A.S.; Di Benedetto, G.; Isla, M.I. Evaluation of antioxidant capacity, genotoxicity and polyphenol content of non-conventional foods: Prosopis flour. Food Res. Int. 2010, 43, 1505–1510. [Google Scholar] [CrossRef]
- Villagra, P.E.; Cony, M.A.; Mantován, N.G.; Rossi, B.E.; González Loyarte, M.M.; Villalba, R.; Marone, L. Ecología y Manejo de los Algarrobales de la Provincia Fitogeográfica del Monte. Ecología y Manejo de Bosques Nativos de Argentina; Editorial Universidad Nacional de La Plata: La Plata, Argentina, 2004. [Google Scholar]
- Burkart, A. A monograph of the genus Prosopis (Leguminosae subfam. Mimosoideae). J. Arnold Arbor. 1976, 57, 219–249, 450–425. [Google Scholar] [CrossRef]
- Felker, P.; Guevara, J.C. Potential of commercial hardwood forestry plantations in arid lands—An economic analyses of Prosopis lumber production in Argentina and the United States. For. Ecol. Manag. 2003, 18, 271–286. [Google Scholar] [CrossRef]
- Felker, P.; Takeoka, G.; Dao, L. Pod mesocarp flour of north and south American species of Leguminous tree Prosopis (Mesquite): Composition and food applications. Food Rev. Int. 2013, 29, 49–66. [Google Scholar] [CrossRef]
- Felker, P.; Grados, N.; Cruz, G.; Prokopiuk, D. Economic assessment of production of flour from Prosopis alba and P. pallida pods for human food applications. J. Arid Environ. 2003, 53, 17–528. [Google Scholar] [CrossRef]
- Código Alimentario Argentino. Capítulo IX: Alimentos Farináceos, Cereales, Harinas y Derivados (681 y 681 Tris). 2021. Available online: https://www.argentina.gob.ar/sites/default/files/2018/05/capitulo_ix_harinasactualiz_2021-08.pdf (accessed on 1 August 2021).
- Pérez, M.J.; Cuello, A.S.; Zampini, I.C.; Ordoñez, R.M.; Alberto, M.R.; Quispe, C.; Schmeda-Hirschmann, G.; Isla, M.I. Polyphenolic compounds and anthocyanin content of Prosopis nigra and Prosopis alba pods flour and their antioxidant and anti-inflammatory capacity. Food Res. Int. 2014, 64, 762–771. [Google Scholar] [CrossRef]
- Cattaneo, F.; Sayago, J.E.; Alberto, M.R.; Zampini, I.C.; Ordoñez, R.M.; Chamorro, V.; Pazos, A.; Isla, M.I. Anti-inflammatory and antioxidant activities, functional properties and mutagenicity studies of protein and protein hydrolysate obtained from Prosopis alba seed flour. Food Chem. 2014, 161, 391–399. [Google Scholar] [CrossRef]
- Cattaneo, F.; Costamagna, M.S.; Zampini, I.C.; Sayago, J.E.; Alberto, M.R.; Chamorro, V.; Pazos, A.; Thomas-Valdez, S.; Schmeda-Hirschmann, G.; Isla, M.I. Flour from Prosopis alba cotyledons: A natural source of nutrient and bioactive phytochemicals. Food Chem. 2016, 208, 89–96. [Google Scholar] [CrossRef]
- Rodriguez, I.F.; Pérez, M.J.; Cattaneo, F.; Zampini, I.C.; Cuello, A.S.; Mercado, M.I.; Ponessa, G.; Isla, M.I. Botanical, chemical and functional characterization of Prosopis alba pericarp flours of different particle size. Food Chem. 2018, 274, 583–591. [Google Scholar] [CrossRef]
- Peréz, M.J.; Zampini, I.C.; Alberto, M.R.; Isla, M.I. Prosopis nigra mesocarp fine flour, a source of phytochemicals with potential effect on enzymes linked to metabolic syndrome, oxidative stress, and inflammatory process. J. Food Sci. 2018, 83, 1454–1462. [Google Scholar] [CrossRef]
- Sciammaro, L.P.; Ferrero, C.; Puppo, M.C. Chemical and nutritional properties of different fractions of Prosopis alba pods and seeds. J. Food Meas. Charact. 2016, 10, 103–112. [Google Scholar] [CrossRef]
- Puppo, M.C.; Felker, P. Prosopis as a heat-tolerant nitrogen-fixing desert food legume. In Prospects for Economic Development in Arid Lands; Edited by Elsevier; Academic Press: Cambridge, MA, USA, 2022; Chapters 19–21. [Google Scholar]
- Felker, P.; Lopez, C.; Soulier, C.; Ochoa, J.; Abdala, R.; Ewens, M. Genetic evaluation of Prosopis alba (algarrobo) in Argentina for cloning elite trees. Agrofor. Syst. 2001, 53, 65–76. [Google Scholar] [CrossRef]
- Ewens, M.; Felker, P. A comparison of pod production and insect ratings of 12 elite Prosopis alba clones in a 5 year semi arid argentine field trial. For. Ecol. Manag. 2010, 260, 378–383. [Google Scholar] [CrossRef]
- Ewens, M.; Gezan, S.; Felker, P. Five year field evaluation of Prosopis alba clones on pH 9–10 soils in Argentina selected for growth in the greenhouse at seawater salinities (45 dS m−1). Forests 2012, 3, 95–113. [Google Scholar] [CrossRef]
- Afyounizadeh Esfahani, D.; Goli, M. The effect of Stevioside-Isomalt, whey protein concentrate, and Bacillus coagulans on the physicochemical and sensory properties of Iranian probiotic Masghati sweet. J. Food Process. Preserv. 2021, e16028. [Google Scholar] [CrossRef]
- Costamagna, M.S.; Ordóñez, R.M.; Zampini, I.C.; Sayago, J.E.; Isla, M.I. Nutritional and antioxidant properties of Geoffroea decorticans, an Argentinean fruit, and derived products (flour, arrope, decoction and hydroalcoholic beverage). Food Res. Int. 2013, 54, 160–168. [Google Scholar] [CrossRef]
- American Association of Cereal Chemists (AACC). Methods 46-10. In Approved Methods of the AACC, 10th ed.; American Association of Cereal Chemists: St Paul, MN, USA, 2000. [Google Scholar]
- Orqueda, M.E.; Rivas, M.; Zampini, I.C.; Alberto, M.R.; Torres, S.; Cuello, A.S.; Sayago, J.E.; Thomas-Valdes, S.; Jimenez-Aspee, F.; Schmeda-Hirschman, G.; et al. Chemical and functional characterization of seed, pulp and skin powder from chilto (Solanum betaceum), an Argentine native fruit. Phenolic fractions affect key enzymes involved in metabolic syndrome and oxidative stress. Food Chem. 2017, 216, 70–79. [Google Scholar] [CrossRef] [PubMed]
- AOCS. Official methods and recommended practices of the American Oil Chemists’ Society. In Official Methods and Recommended Practices of the American Oil Chemists’ Society; American Oil Chemists’ Society: Champaign, IL, USA, 1989. [Google Scholar]
- AOAC. Method 925.10 Solids (Total) and Moisture in Flour. In Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemists: Rockville, MD, USA, 1998. [Google Scholar]
- American Association of Cereal Chemists (AACC). Methods 08-01. In Approved Methods of the AACC, 10th ed.; American Association of Cereal Chemists: St Paul, MN, USA, 2000. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventos, R.M. Analysis of total phenols and other oxidation substrates and antioxidants. Meth. Enzymol. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Isla, M.I.; Salas, A.L.; Danert, F.C.; Zampini, I.C.; Ordoñez, R.M. Analytical methodology optimization to estimate the content of non-flavonoid phenolic compounds in Argentine propolis extracts. Pharm. Biol. 2014, 52, 835–840. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Fernando, C.D.; Soysa, P. Optimized enzymatic colorimetric assay for determination of hydrogen peroxide (H2O2) scavenging activity of plant extracts. MethodsX 2015, 2, 283–291. [Google Scholar] [CrossRef] [PubMed]
- Di Rienzo, J.A. Grupo InfoStat, FCA, Universidad; InfoStat Version 2015; Unversidad Nacional de Córdoba: Cordoba, Argentina, 2015. [Google Scholar]
- Díaz-Batalla, L.; Hernández-Uribe, J.P.; Román-Gutiérrez, A.D.; Cariño-Cortés, R.; Castro-Rosas, J.; Téllez-Jurado, A.; Gómez-Aldapa, C.A. Chemical and nutritional characterization of raw and thermal-treated flours of Mesquite (Prosopis laevigata) pods and their residual brans. CYTA—J. Food 2018, 16, 444–451. [Google Scholar] [CrossRef]
- Gorinstein, S.; Pawelzik, E.; Delgado-Licon, E.; Haruenkit, R.; Weisz, M.; Trakhtenberg, S. Characterization of pseudocereal and cereal proteins by protein and amino acid analyses. J. Sci. Food Agric. 2002, 82, 886–891. [Google Scholar] [CrossRef]
- Food and Agricultural Organization. Findings and recommendations of the 2011 FAO Expert Consultation on Protein. In Dietary Protein Quality Evaluation in Human Nutrition; Report of a FAO Expert Consultation; FAO: Rome, Italy, 2011; Chapter 4. [Google Scholar]
- O’ Keefe, S.J. The association between dietary fibre deficiency and high-income lifestyle-associated diseases: Burkitt’s hypothesis revisited. Lancet Gastroenterol. Hepatol. 2019, 4, 984–986. [Google Scholar] [CrossRef]
- Bilge, G.; Sezer, B.; Efe, K.; Berberoglu, H.; Koksel, H.; Hakki, I. Análisis de cenizas de una muestra de harina mediante el uso de espectroscopia de descomposición inducida por láser. Spectrochim. Acta Part B At. Spectrosc. 2016, 124, 74–78. [Google Scholar] [CrossRef]
- Yang, L.; Wen, K.; Ruan, X.; Zhao, Y.; Wei, F.; Wang, Q. Response of plant secondary metabolites to environmental factors. Molecules 2018, 23, 762. [Google Scholar] [CrossRef] [PubMed]
- Rico, D.; Martín-Diana, A.B.; Martínez-Villaluanga, C.; Aguirre, L.; Silván, J.M.; Dueñas, M.; De Luis, D.A.; Lasa, A. In vitro approach for evaluation of carob byproducts as source bioactive ingredients with potential to attenuate metabolic syndrome (MetS). Heliyon 2019, 5, e01175. [Google Scholar] [CrossRef]
- Quispe, C.; Petroll, K.; Theoduloz, C.; Schmeda-Hirschmann, G. Antioxidant effect and characterization of South American Prosopis pods syrup. Food Res. Int. 2014, 56, 174–181. [Google Scholar] [CrossRef]
- Schmeda-Hirschmann, G.; Quispe, C.; Soriano Mdel, P.C.; Theoduloz, C.; Jiménez-Aspee, F.; Pérez, M.J.; Cuello, A.S.; Isla, M.I. Chilean Prosopis mesocarp flour: Phenolic profiling and antioxidant activity. Molecules 2015, 20, 7017–7033. [Google Scholar] [CrossRef]
- Pérez, M.J.; Rodríguez, I.F.; Zampini, I.C.; Cattaneo, F.; Mercado, M.I.; Ponessa, G.; Isla, M.I. Prosopis nigra fruits waste characterization, a potential source of functional ingredients for food formulations. LWT 2020, 132, 109828. [Google Scholar] [CrossRef]
- Courts, F.L.; Williamson, G. The occurrence, fate and biological activities of C-glycosyl flavonoids in the human diet. Crit. Rev. Food Sci. Nutr. 2015, 55, 1352–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).