Metabolic Engineering of Escherichia coli for Producing Astaxanthin as the Predominant Carotenoid


Abstract
1. Introduction
2. Results and Discussion
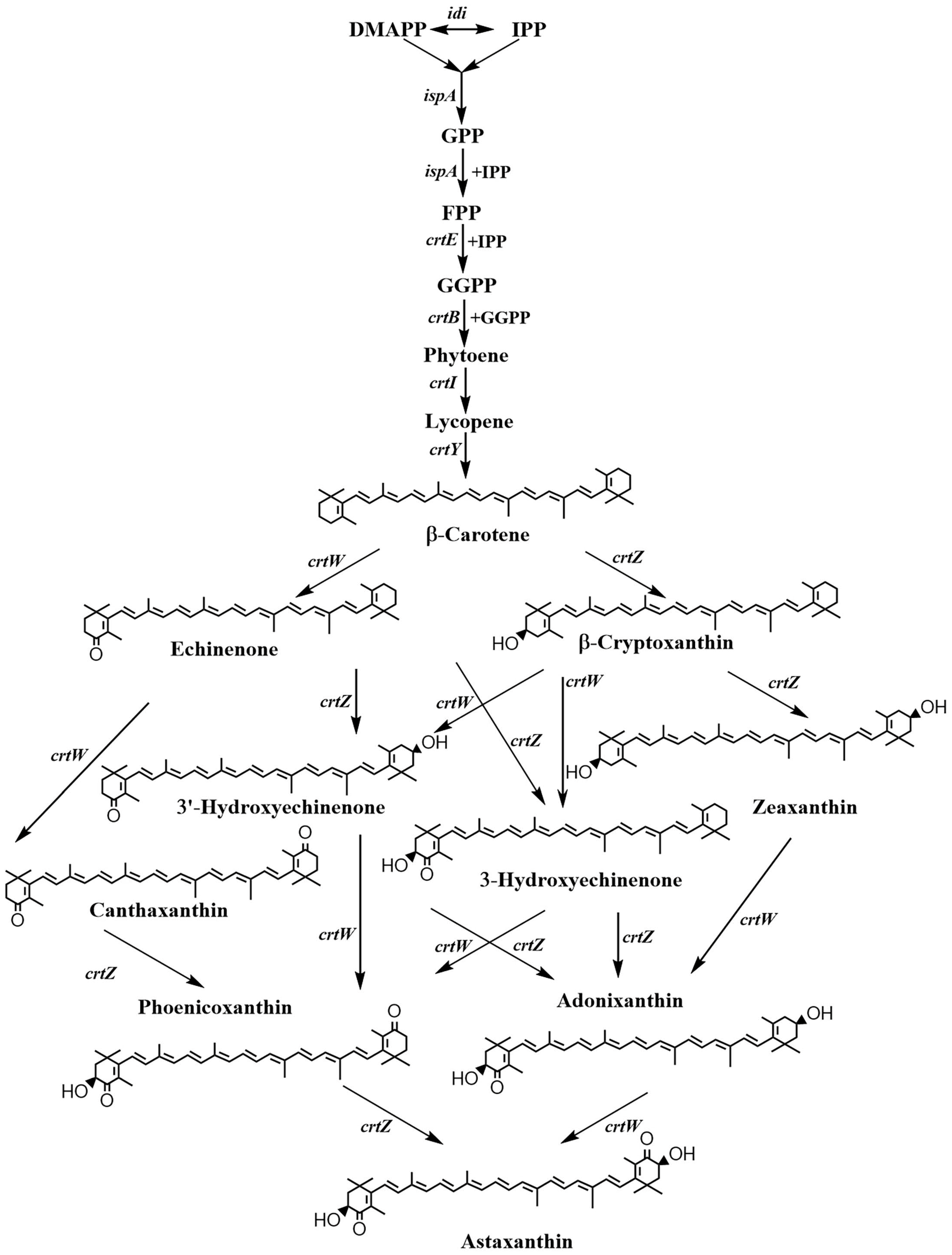
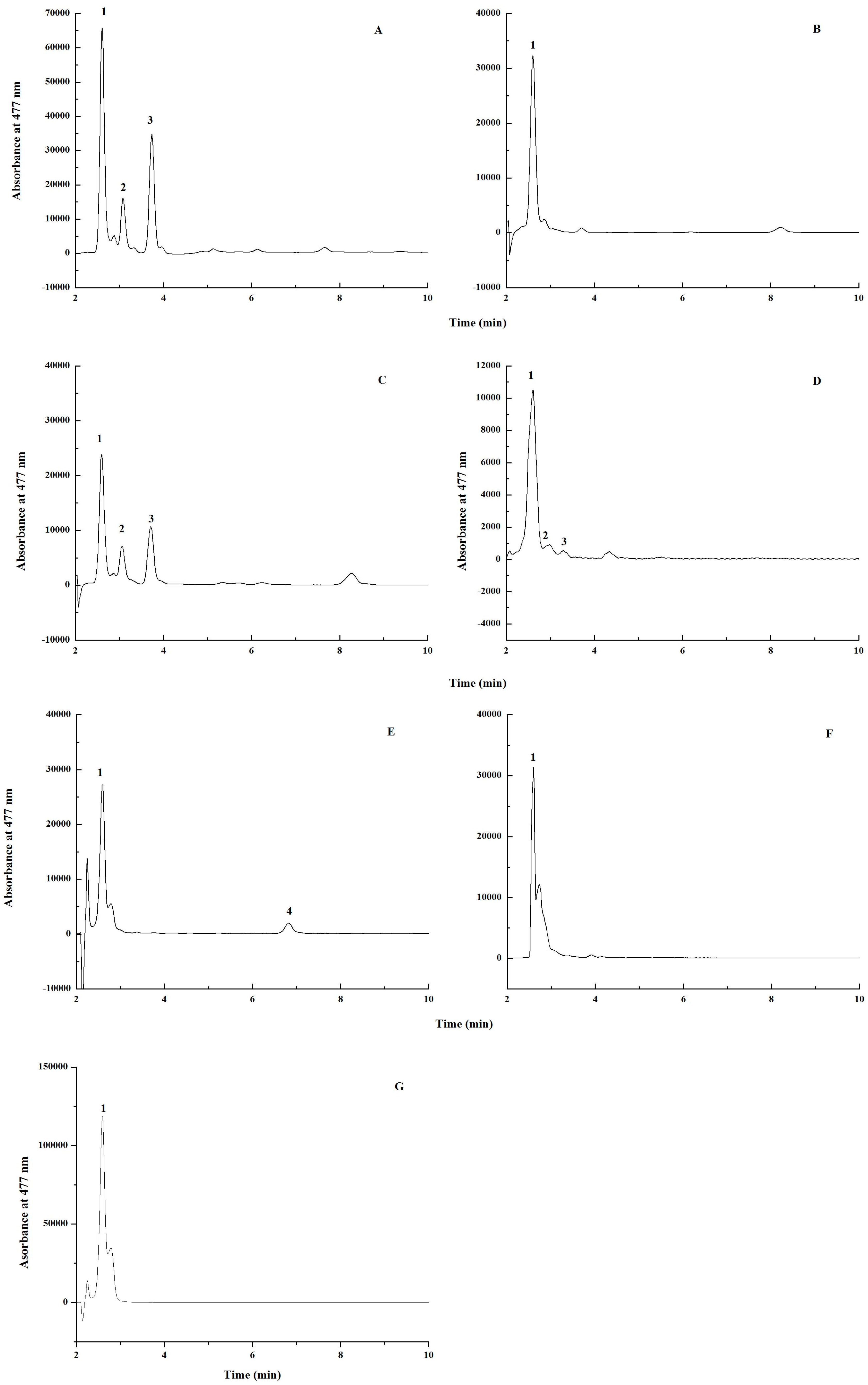
2.1. Screening of β-Carotene Ketolase
2.2. Balancing the Activities of β-Carotene Ketolase and Hydroxylase
3. Materials and Methods
3.1. Strains, Plasmids and Primers
3.2. Genetic Methods
3.3. Astaxanthin Production in Shake Flasks
3.4. Extraction and Quantification of Carotenoids
3.5. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| B. sp. | Brevundimonas sp. |
| S. sp. | Sphingomonas sp. |
| P. sp. | Paracoccus sp. |
| P. ananatis | Pantoea ananatis |
| H. pluvialis | Haematococcus pluvialis |
| P. agglomerans | Pantoea agglomerans |
| C. reinhardtii | Chlamydomonas reinhardtii |
| A. sp. | Anabaena sp. |
| N. punctiforme | Nostoc punctiforme |
| MEV | mevalonate |
References
- Hussein, G.; Sankawa, U.; Goto, H.; Matsumoto, K.; Watanabe, H. Astaxanthin, a carotenoid with potential in human health and nutrition. J. Nat. Prod. 2006, 69, 443–449. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.C.; Schmidt-Dannert, C. Metabolic engineering towards biotechnological production of carotenoids in microorganisms. Appl. Microbiol. Biotechnol. 2002, 60, 1–11. [Google Scholar] [PubMed]
- Ye, V.M.; Bhatia, S.K. Pathway engineering strategies for production of beneficial carotenoids in microbial hosts. Biotechnol. Lett. 2012, 34, 1405–1414. [Google Scholar] [CrossRef] [PubMed]
- Lemuth, K.; Steuer, K.; Albermann, C. Engineering of a plasmid-free Escherichia coli strain for improved in vivo biosynthesis of astaxanthin. Microb. Cell Fact. 2011, 10, 29. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.P.; Ye, L.D.; Xie, W.P.; Lv, X.M.; Yu, H.W. Highly efficient biosynthesis of astaxanthin in Saccharomyces cerevisiae by integration and tuning of algal crtZ and bkt. Appl. Microbiol. Biotechnol. 2015, 99, 8419–8428. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.P.; Xie, W.P.; Li, A.P.; Wang, F.; Yao, Z.; Bian, Q.; Zhu, Y.Q.; Yu, H.W.; Ye, L.D. Alleviation of metabolic bottleneck by combinatorial engineering enhanced astaxanthin synthesis in Saccharomyces cerevisiae. Enzyme Microb. Technol. 2017, 100, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Henke, N.A.; Heider, S.A.E.; Peters-Wendisch, P.; Wendisch, V.F. Production of the marine carotenoid astaxanthin by metabolically engineered Corynebacterium glutamicum. Mar. Drugs 2016, 14, 124. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.A.; Wilczek, J.; Odom, J.M.; Cheng, Q.O. Engineering a β-carotene ketolase for astaxanthin production. Metab. Eng. 2006, 8, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.F.; Gudina, E.; Barredo, J.L. Conversion of β-carotene into astaxanthin: Two separate enzymes or a bifunctional hydroxylase-ketolase protein? Microb. Cell Fact. 2008, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Misawa, N. Carotenoid beta-ring hydroxylase and ketolase from marine bacteria-promiscuous enzymes for synthesizing functional xanthophylls. Mar. Drugs 2011, 9, 757–771. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.K.; Nishida, Y.; Matsuda, S.; Adachi, K.; Kasai, H.; Peng, X.; Komemushi, S.; Miki, W.; Misawa, N. Characterization of beta-carotene ketolases, CrtW, from marine bacteria by complementation analysis in Escherichia coli. Mar. Biotechnol. 2005, 7, 515–522. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.K.; Harada, H.; Matsuda, S.; Misawa, N. Characterization of two beta-carotene ketolases, CrtO and CrtW, by complementation analysis in Escherichia coli. Appl. Microbiol. Biotechnol. 2007, 75, 1335–1341. [Google Scholar] [CrossRef] [PubMed]
- Makino, T.; Harada, H.; Ikenaga, H.; Matsuda, S.; Takaichi, S.; Shindo, K.; Sandmann, G.; Ogata, T.; Misawa, N. Characterization of cyanobacterial carotenoid ketolase CrtW and hydroxylase CrtR by complementation analysis in Escherichia coli. Plant Cell Physiol. 2008, 49, 1867–1878. [Google Scholar] [CrossRef] [PubMed]
- Scaife, M.A.; Burja, A.M.; Wright, P.C. Characterization of cyanobacterial β-carotene ketolase and hydroxylase Genes in Escherichia coli, and their application for astaxanthin biosynthesis. Biotechnol. Bioeng. 2009, 103, 944–955. [Google Scholar] [CrossRef] [PubMed]
- Scaife, M.A.; Ma, C.A.; Ninlayarn, T.; Wright, P.C.; Armenta, R.E. Comparative analysis of beta-carotene hydroxylase genes for astaxanthin biosynthesis. J. Nat. Prod. 2012, 75, 1117–1124. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.W.; Stead, K.J.; Yao, H.; He, H.X. Mutational and functional analysis of the beta-carotene ketolase involved in the production of canthaxanthin and astaxanthin. Appl. Environ. Microb. 2006, 72, 5829–5837. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.J.; Huang, J.C.; Liu, J.; Li, Y.; Jiang, Y.; Xu, Z.F.; Sandmann, G.; Chen, F. Functional characterization of various algal carotenoid ketolases reveals that ketolating zeaxanthin efficiently is essential for high production of astaxanthin in transgenic Arabidopsis. J. Exp. Bot. 2011, 62, 3659–3669. [Google Scholar] [CrossRef] [PubMed]
- Li, X.R.; Tian, G.Q.; Shen, H.J.; Liu, J.Z. Metabolic engineering of Escherichia coli to produce zeaxanthin. J. Ind. Microbiol. Biotechnol. 2015, 42, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.J.; Cheng, B.Y.; Zhang, Y.M.; Tang, L.; Li, Z.; Bu, Y.F.; Li, X.R.; Tian, G.Q.; Liu, J.Z. Dynamic control of the mevalonate pathway expression for improved zeaxanthin production in Escherichia coli and comparative proteome analysis. Metab. Eng. 2016, 38, 180–190. [Google Scholar] [CrossRef] [PubMed]
- Fraser, P.D.; Miura, Y.; Misawa, N. In vitro characterization of astaxanthin biosynthetic enzymes. J. Biol. Chem. 1997, 272, 6128–6135. [Google Scholar] [CrossRef] [PubMed]
- Fraser, P.D.; Shimada, H.; Misawa, N. Enzymic confirmation of reactions involved in routes to astaxanthin formation, elucidated using a direct substrate in vitro assay. Eur. J. Biochem. 1998, 252, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Zhou, Y.J.; Li, X.W.; Zhu, F.Y.; Cheng, Y.B.; Liu, Y.; Deng, Z.X.; Liu, T.G. Genome mining of astaxanthin biosynthetic genes from Sphingomonas sp. ATCC 55669 for heterologous overproduction in Escherichia coli. Biotechnol. J. 2016, 11, 228–237. [Google Scholar] [CrossRef] [PubMed]
- Zelcbuch, L.; Antonovsky, N.; Bar-Even, A.; Levin-Karp, A.; Barenholz, U.; Dayagi, M.; Liebermeister, W.; Flamholz, A.; Noor, E.; Amram, S.; et al. Spanning high-dimensional expression space using ribosome-binding site combinatorics. Nucleic Acids Res. 2013, 41. [Google Scholar] [CrossRef] [PubMed]
- Guzman, L.M.; Belin, D.; Carson, M.J.; Beckwith, J. Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter. J. Bacteriol. 1995, 177, 4121–4130. [Google Scholar] [CrossRef] [PubMed]
- Boel, G.; Letso, R.; Neely, H.; Price, W.N.; Wong, K.H.; Su, M.; Luff, J.D.; Valecha, M.; Everett, J.K.; Acton, T.B.; et al. Codon influence on protein expression in E. coli correlates with mRNA levels. Nature 2016, 529, 358–363. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Plasmid | OD600 * | Astaxanthin Concentration, mg/L | Astaxanthin Content, mg/gDCW |
|---|---|---|---|
| Single gene | |||
| pZS-crtWBsp | 9.55 ± 0.16 | 8.1 ± 0.1 | 2.7 ± 0.1 |
| pZS-crtWPsp | 8.05 ± 0.47 | 1.3 ± 0.1 | 0.5 ± 0.1 |
| pZS-crtWSsp | 8.61 ± 0.21 | 0.8 ± 0.1 | 0.3 ± 0.1 |
| pZS-bkt | 11.07 ± 0.20 | 5.0 ± 0.2 | 1.4 ± 0.1 |
| Double genes | |||
| pZS-2crtWBsp | 19.67 ± 0.33 | 28.8 ± 0.2 | 4.6 ± 0.1 |
| pZS-2bkt | 22.15 ± 0.33 | 20.9 ± 1.6 | 3.5 ± 0.1 |
| pZS-2crtWPsp | 16.82 ± 0.56 | 7.0 ± 0.1 | 1.3 ± 0.1 |
| pZS-2crtWSsp | 14.91 ± 0.31 | 1.1 ± 0.6 | 0.2 ± 0.1 |
| Mixed genes | |||
| pZS-crtWBspcrtWPsp | 19.93 ± 0.38 | 12.6 ± 0.6 | 2.0 ± 0.1 |
| pZS-crtWBspcrtWSsp | 21.73 ± 0.19 | 24.6 ± 0.4 | 3.5 ± 0.1 |
| pZS-crtWBspbkt | 19.9 ± 1.27 | 20.9 ± 1.6 | 3.3 ± 0.1 |
| pZS-crtWPspcrtWSsp | 16.6 ± 0.17 | 8.5 ± 0.2 | 1.6 ± 0.2 |
| pZS-crtWPspbkt | 20.01 ± 0.12 | 10.5 ± 1.6 | 1.6 ± 0.1 |
| pZS-crtWSspbkt | 21.57 ± 0.38 | 11.2 ± 0.7 | 1.6 ± 0.1 |
| Strain | Astaxanthin Yield | Astaxanthin Ratio (%) | Reference |
|---|---|---|---|
| E. coli | 5.8 mg/g DCW | N.D. * | [23] |
| E. coli | 8.64 mg/g DCW | N.D. | [22] |
| E. coli | 1.4 mg/g DCW | 95 | [4] |
| E. coli | 1.99 mg/g DCW | 90 | [14] |
| E. coli | 7.4 ± 0.3 mg/g DCW | 96.6 | This study |
| S. cerevisiae | 4.7 mg/g DCW | N.D. | [5] |
| S. cerevisiae | 8.10 mg/g DCW | N.D. | [6] |
| C. glutamicum | 0.4 mg/L/h | N.D. | [7] |
| Name | Description | Reference/Sources |
|---|---|---|
| Strain | ||
| E. coli BETA-1 | β-Carotene producing strain | [18] |
| E. coli ZEAX | Zeaxanthin producing strain, one copy of Pantoea ananatis crtZ under the control of the P37 promoter was integrated into E. coli BETA-1 chromosome | [19] |
| E. coli ZEAX-4 | Zeaxanthin producing strain, two copies of P. ananatis crtZ under the control of the P37 promoter was integrated into E. coli BETA-1 chromosome | This study |
| E. coli ASTA | Astaxanthin producer, one of B. sp. SD212 crtW under the control of the P37 promoter was integrated into E. coli ZEAX-4 chromosome | This study |
| E. coli ASTA-1 | Astaxanthin producer, another copy of P. ananatis crtY under the control of the P37 promoter was integrated into E. coli ASTA-1 chromosome | This study |
| Plasmid | ||
| pZSABP | Constitute expression vector, pSC101 ori, P37 promoter, Ampr, BglBrick, ePathBrick containing four isocaudamer (AvrII, NheI, SpeI and XbaI) | [18] |
| pBAD33 | Expression vector, PBAD, p15A ori, Cmr | [24] |
| pZS-crtWBsp | pZSABP containing Brevundimonas sp. SD212 crtW under the control of the P37 promoter | This study |
| pZS-crtWPsp | pZSABP containing Paracoccus sp. N81106 crtWL175W under the control of the P37 promoter | This study |
| pZS-crtWSsp | pZSABP containing Sphingomonas sp. DC18 crtWF213L/R203W under the control of the P37 promoter | This study |
| pZS-bkt | pZSABP containing Chlamydomonas reinhardtii bkt under the control of the P37 promoter | This study |
| pZS-2crtWBsp | pZSABP containing two copies of B.s sp. SD212 crtW under the control of the P37 promoter | This study |
| pZS-2bkt | pZSABP containing two copies of C. reinhardtii bkt under the control of the P37 promoter | This study |
| pZS-2crtWPsp | pZSABP containing two copies of P. sp. N81106 crtWL175W under the control of the P37 promoter | This study |
| pZS-2crtWSsp | pZSABP containing two copies of S. sp. DC18 crtWF213L/R203W under the control of the P37 promoter | This study |
| pZS-crtWBspcrtWPsp | pZSABP containing B.s sp. SD212 crtW under the control of the P37 promoter and P. sp. N81106 crtWL175W under the control of the P37 promoter | This study |
| pZS-crtWBspcrtWSsp | pZSABP containing B.s sp. SD212 crtW under the control of the P37 promoter and S. sp. DC18 crtWF213L/R203W under the control of the P37 promoter | This study |
| pZS-crtWBspbkt | pZSABP containing B.s sp. SD212 crtW under the control of the P37 promoter and C. reinhardtii bkt under the control of the P37 promoter | This study |
| pZS-crtWPspcrtWSsp | pZSABP containing P. sp. N81106 crtWL175W under the control of the P37 promoter and S. sp. DC18 crtWF213L/R203W under the control of the P37 promoter | This study |
| pZS-crtWPspbkt | pZSABP containing P. sp. N81106 crtWL175W under the control of the P37 promoter and C. reinhardtii bkt under the control of the P37 promoter | This study |
| pZS-crtWSspbkt | pZSABP containing S. sp. DC18 crtWF213L/R203W under the control of the P37 promoter and C. reinhardtii bkt under the control of the P37 promoter | This study |
| pBAD-crtZ | pBAD33 containing P. ananatis crtZ | This study |
| pBAD-crtWBsp | pBAD33 containing B.s sp. SD212 crtW | This study |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, Q.; Bu, Y.-F.; Liu, J.-Z. Metabolic Engineering of Escherichia coli for Producing Astaxanthin as the Predominant Carotenoid. Mar. Drugs 2017, 15, 296. https://doi.org/10.3390/md15100296
Lu Q, Bu Y-F, Liu J-Z. Metabolic Engineering of Escherichia coli for Producing Astaxanthin as the Predominant Carotenoid. Marine Drugs. 2017; 15(10):296. https://doi.org/10.3390/md15100296
Chicago/Turabian StyleLu, Qian, Yi-Fan Bu, and Jian-Zhong Liu. 2017. "Metabolic Engineering of Escherichia coli for Producing Astaxanthin as the Predominant Carotenoid" Marine Drugs 15, no. 10: 296. https://doi.org/10.3390/md15100296
APA StyleLu, Q., Bu, Y.-F., & Liu, J.-Z. (2017). Metabolic Engineering of Escherichia coli for Producing Astaxanthin as the Predominant Carotenoid. Marine Drugs, 15(10), 296. https://doi.org/10.3390/md15100296