Abstract
This study evaluated the microbial quality, safety, and ecology of a retail delicatessen Galotyri-like fresh acid-curd cheese traditionally produced by mixing fresh natural Greek yogurt with ‘Myzithrenio’, a naturally fermented and ripened whey cheese variety. Five retail cheese batches (mean pH 4.1) were analyzed for total and selective microbial counts, and 150 presumptive isolates of lactic acid bacteria (LAB) were characterized biochemically. Additionally, the most and the least diversified batches were subjected to a culture-independent 16S rRNA gene sequencing analysis. LAB prevailed in all cheeses followed by yeasts. Enterobacteria, pseudomonads, and staphylococci were present as <100 viable cells/g of cheese. The yogurt starters Streptococcus thermophilus and Lactobacillus delbrueckii were the most abundant LAB isolates, followed by nonstarter strains of Lactiplantibacillus, Lacticaseibacillus, Enterococcus faecium, E. faecalis, and Leuconostoc mesenteroides, whose isolation frequency was batch-dependent. Lactococcus lactis isolates were sporadic, except for one cheese batch. However, Lactococcus lactis, Enterobacteriaceae, Vibrionaceae, Salinivibrio, and Shewanellaceae were detected at fairly high relative abundances culture-independently, despite the fact that their viable counts in the cheeses were low or undetectable. Metagenomics confirmed the prevalence of S. thermophilus and Lb. delbrueckii. Overall, this delicatessen Galotyri-like cheese product was shown to be a rich pool of indigenous nonstarter LAB strains, which deserve further biotechnological investigation.
1. Introduction
Acid-curd cheeses form a distinct group of soft, spreadable, low-pH (pH 3.5 to 5.0), but high-moisture (60–75%; water activity (aw) >0.98 to 0.99) dairy foods produced at industrial or artisan scale and consumed fresh or ripened [1,2]. Cottage cheese is the most popular and most often investigated acid-curd cheese variety worldwide [3,4]. Several traditional acid-curd cheese varieties are also produced in Greece from raw, thermized, pasteurized, or boiled ewe’s and goat’s milks, including five Protected Designation of Origin (PDO) cheese varieties: Anevato, Galotyri, Katiki, Kopanisti, and Pichtogalo Chanion [5,6,7,8,9]. Galotyri is considered the oldest and most popular Greek acid-curd cheese traditionally made using ‘boiled’ (>80 to 90 °C), naturally acidified milk in the regions of Epirus and Thessaly [5,10]. Samelis and Kakouri [11] pointed out major technological and microecological differences between an industrial-type and five artisanal Galotyri PDO cheese products marketed before or after the explosion of the Greek financial crisis, which has caused major reformations in the national food sector from 2010 onward. Specifically, the Greek regulatory authorities currently require all small and medium-size enterprises producing PDO (dairy) foodstuffs or specialties to be certified by AGROCERT and their products to be packaged and labeled accordingly. Statistical production data of each Greek PDO food product should also be kept [11].
Particularly with regard to traditional PDO cheeses, the General Directorate of Food Quality Control and Assurance of the Hellenic Agricultural Organization ‘DIMITRA’ has increased inspections of products and plants in an attempt to preserve and protect PDO cheese authenticity. It has become pivotal to ensure that all certified brand-named PDO cheese products comply with the respective specifications described in article 83D of the Hellenic Code of Food and Beverages regarding the origin of the milk and the compositional and technological attributes of the final cheese [5,11]. Thus, according to its PDO description, the technology of authentic Galotyri cheese aims at the production of a uniform fresh curd following acidification of the ‘boiled’ milk with lactic acid bacteria (LAB) [5,12]. LAB can be added to ferment the ‘boiled’ or industrially pasteurized Galotyri cheese milk in the form of natural undefined starter cultures, including back slope techniques [10,13,14], or commercially defined starter cultures [11,15,16]. Rennet is often added to enhance curdling of the milk and to stabilize the curd [5,12], while draining, salting, and mainly ripening [17,18] further alter the microbial ecology, sensory characteristics, and total quality attributes of the artisanal Galotyri PDO cheese products [11,12].
It must be clarified that the manufacturing technology of Galotyri PDO cheese does not include any mixing steps of the fresh curd with other dairy products for ‘external aromatization’ purposes [5,12]. However, addition of fresh feta cheese trimmings at a 1:5 proportion in basic yogurt curds has been reviewed as a common empirical practice for traditional Galotyri cheese processors [19]. Samelis and Kakouri [11] opinioned that the ‘arbitrary’ dispersion in fresh Galotyri curds of feta trimmings should be neither permitted nor encouraged because it is in contradiction of the PDO certification of this authentic cheese [5]. In fact, following the recent increases in national food inspections, several new delicatessen Galotyri-like acid-curd cheese products have appeared on the Greek market with new trade names. Most of them are fresh yogurt spreads with feta cheese [11]. This more simplified and profitable acid-curd cheese making process also relies on traditional practices. In most Greek rural areas, excess ewe’s/goat’s milks, mainly in the summer, are preserved by making yogurt at home, which is strained and often mixed with trimmed white brined cheeses to consume as an appetizer. In addition to naturally fermented yogurt, fresh acid-curd cheeses and white brine cheeses produced in Greece, Bulgaria, and other Balkan or East Mediterranean countries since antiquity have been associated with beneficial effects on human health, attributed to the probiotic properties of their indigenous LAB [6,19,20]. Therefore, it is important to characterize the microbiota of Greek yogurt or of traditional Greek acid-curd cheese products prepared with yogurt in order to detect novel LAB strains with high biotechnological or probiotic potential for the development of novel functional foods.
An alternate traditional practice is to mix fresh naturally fermented Greek yogurt with naturally fermented and ripened whey cheese trimmings to prepare fresh acid-curd spread homogenates with pleasant, refreshing taste and aroma. The above type of fresh Galotyri-like cheese specialty, which may also have human health-promoting properties, has been commercialized locally, but its microbial quality, safety, and ecology have yet to be investigated. Therefore, the aim of this study was to characterize the natural microbiota of this retail delicatessen cheese product using classical microbiological methods and a culture-independent 16S rRNA sequencing metagenomic approach. The isolation and characterization of the natural LAB biota of this traditional acid-curd cheese of Epirus represent the first research tasks of BIO TRUST, an ongoing collaborative research project, which targets the development of novel indigenous starter, protective, or probiotic cultures for application in different traditional Greek cheese technologies [21].
2. Materials and Methods
2.1. Preparation and Sampling of Retail Galotyri-Like Cheese Products
Five sample portions (500 g each) of five individual batches of bulk ready-to-eat Galotyri-like acid-curd cheeses, distributed as “Galatyri” by the producer, were obtained from the Ioannina retail outlet shop of the Pappas Bros. traditional dairy (Skarfi E.P.E., Filippiada, Epirus, Greece). Samples were placed in plastic flexible containers with a lid and were transported to the microbiology laboratory of the Dairy Research Department in insulated ice boxes. Each 500 g sample represented one retail cheese batch. The five batches analyzed during this study were produced on independent and chronologically distant days, from December to March, corresponding to two annual ewe/goat milking periods. Each cheese batch was prepared by mixing 50 kg of a fresh uniform ‘basal’ curd of traditional Greek yogurt with 25 kg of a naturally fermented and ripened granular whey cheese, named ‘Myzithrenio’, traditionally produced as described below.
Briefly, the whey collected after processing of Graviera cooked hard cheese in the Pappas dairy plant, according to the standard manufacturing protocol described by Noutsopoulos et al. [22], was enriched with 5% raw ewe’s milk and heated at 85–90 °C in a stainless-steel vessel over a gas burner to obtain a fresh whey cheese mass via the traditional manufacturing method. The whey–protein coagulum was transferred into perforated plastic cheese molds placed in a cooling room at 12 °C, drained for 48 h, dry-salted with ca. 3% edible sea salt, and filled into empty wooden cheese barrels. The barrels were closed firmly and turned upside down, while the cheese mass was left to acidify and ripen naturally at 18 °C for 30 days. Afterward, the mature ‘Myzithrenio’ cheese was distributed for bulk retail selling, directly from the barrel. Otherwise, relative to this study, the barrels were opened in the plant, and the ripened whey cheese mass was distributed in 5 kg portions into cryovac bags, which were vacuum-packaged (VP) to prevent growth of aerobic spoilage yeasts and molds and stored at 4 °C for 3–4 months. Periodically, the desired VP quantities of naturally acidified (pH 4.0–4.5) and ripened ‘Myzithrenio’ cheese were mixed proportionally with fresh yogurt, as described above, to obtain the fresh Galotyri-like acid-curd cheese product illustrated in Figure 1.

Figure 1.
Traditional ready-to-eat (RTE) fresh Galotyri-like acid-curd cheese produced and retail-distributed by the Skarfi EPE-Pappas Bros. traditional dairy.
The basic yogurt curd was fresh and nondrained. It was traditionally made by seeding full-fat ewe’s milk with a natural yogurt starter routinely prepared by batch circulation in the Pappas plant. The milk was heated at 90–92 °C and cooled at 48 °C to add the yogurt starter [23] diluted with water. The yogurt fermentation process lasted 3 h upon transferring the inoculated milk in a controlled chamber at 42 °C. We previously analyzed this type of natural yogurt starter and confirmed it to contain typical strains of Streptococcus thermophilus and Lactobacillus delbrueckii subsp. bulgaricus [24], in agreement with relevant studies by others on the microbiology of yogurt and yogurt starters [20,23].
After the fresh yogurt curd and the ripened ‘Myzithrenio’ cheese were mixed, no further ripening was applied to the final RTE delicatessen cheese product (Figure 1). In general, this Galotyri-like acid-curd cheese specialty is distributed for selling after preparation, is stored aerobically under refrigeration during retail distribution, and has an approximate retail shelf-life of 2 weeks before the manifestation of spoilage yeasts on the soft cheese surface. All present retail cheese samples were less than 1 week old at the time of analysis.
2.2. Cheese Analyses
All cheese samples were analyzed microbiologically and for pH within 2 h of their transportation to our laboratory. The pH was measured using a digital pH meter (Jenway 3510, Dunmow, Essex, UK). The glass electrode was immersed in the soft cheese mass after the microbiological sampling done as described below.
Each plastic container was opened near a Bunsen burner, and the cheese mass was stirred thoroughly with a sterile spoon spatula. Afterward, 25 g of cheese was homogenized with 225 mL of 0.1% w/v buffered peptone water (BPW) in a stomacher (Lab Blender 400, Seward, London, UK) for 60 s at room temperature. Serial decimal dilutions in 0.1% BPW were prepared, and duplicate 1 mL or 0.1 mL samples of appropriate dilutions were poured or spread on the total or selective agar plates. Unless otherwise stated, all diluents, enumeration agar media, and supplements were purchased from Neogen Culture Media (Heywood, Bury, UK).
All samples were analyzed for total bacteria, total mesophilic and thermophilic LAB, total mesophilic and thermophilic dairy (lactose-fermenting) LAB, enterococci, coliform bacteria, Pseudomonas-like bacteria, total and pathogenic (coagulase-positive) staphylococci, and yeasts. The agar media and incubation conditions were according to the analytical procedures described in previous studies [11,24,25]; they are summarized in Table 1. The lowest detection limit was set at 100 CFU/g. For coliforms and RFP+ staphylococci, the lowest detection limit was set at 10 CFU/g of cheese. The presence of natural Listeria spp. and Salmonella spp. contaminants was assessed by one-step culture enrichment of 25 g cheese samples [13,24]. Presumptive Listeria spp. or Salmonella spp. colonies on selective Palcam or Rambach plates were respectively identified using the API Listeria or the API 20E identification kits (BioMerieux, Marcy l’ Etoile, France).

Table 1.
Microbial populations (log CFU/g) and pH values of five retail individual batches of RTE Galotyri-like acid-curd cheese.
2.3. Isolation and Biochemical Characterization of the Cheese LAB Biota
The LAB types prevailing in each of the five cheese batches were determined by isolating five colonies from the highest dilution plate of each of the first six LAB-selective enumeration agar media listed in Table 1, namely, Milk Plate Count agar (MPCA)/37 °C (total cheese bacteria), MRS/30 °C and M17/22 °C (total mesophilic LAB), MRS/45 °C and M17/42 °C (total thermophilic LAB), and Slanetz and Bartley (SB) agar 37 °C (enterococci). Attention was paid to collect representative isolates of all macroscopically different colonies on each plate. In this manner, 150 presumptive LAB isolates (30/cheese batch) were collected. Colonies isolated from MPCA and M17 agar plates were transferred for growth to 10 mL of M17 broth. Colonies isolated from MRS and SB agar plates were transferred for growth to 10 mL of MRS broth. The tubes were incubated at 30 °C or 37 °C, depending on whether the isolates were from agar media incubated at 22 to 30 °C or 37 to 45 °C, respectively (Table 1). Following growth, all isolates were checked for purity on streaked MRS or M17 agar plates, and the purified isolates were stored in 5 mL of MRS broth with 20% glycerol at −30 °C. The stock LAB isolates were resuscitated in 10 mL of MRS or M17 broth at 30 °C or 37 °C for 24 to 72 h, and they were subcultured twice before testing.
The isolates were characterized biochemically according to established phenotypic criteria [26,27,28]. Test methods were according to our previous studies [11,24]. Briefly, all isolates were first tested rapidly for Gram stain and catalase reactions. Gram-positive and catalase-negative isolates were tested further for cell morphology by phase contrast microscopy, gas (CO2) production from glucose, ammonia production from arginine, growth at 15 °C and 45 °C, growth in 6.5% salt, ability for typical growth on Kanamycin Aesclulin Azide (KAA) agar, and the fermentation of 13 sugars (2% v/w) in 96-well presterilized miniplates: L-arabinose, cellobiose, galactose, lactose, maltose, mannitol, melibiose, raffinose, ribose, sorbitol, sucrose, trehalose, and xylose (Sigma-Aldrich Chemie GmbH, Steinheim, Germany). Filter-sterilized sugar solutions (10% w/v; 40 μL) were placed into the wells first. Sugar-free basal MRS broth (pH 6.9–7.0), prepared from its basic constituents with 0.17 g/L of bromocresol purple (Sigma) added as a pH indicator, was used to prepare washed cell suspensions of the LAB isolates for inoculation of the miniplates (160 μL/well). All tests were done in duplicate, and the isolates were grouped at the genus or species level, according to key differentiating criteria [26,27,28].
2.4. DNA Extraction from Cheese Samples and 16S rRNA Sequencing of Microbiota
The cheese batches A and B were selected and subjected to characterization of the total bacterial biota culture independently, with the aim of comparing the two methods and gaining more information on the acid-curd cheese biodiversity. The Nucleospin Food kit (Macherey-Nagel, Düren, Germany) was used to extract the DNA (20 ng/μL) directly from the cheese samples (one sample per batch). Following DNA extraction, the V2–4–8 and V3–7–9 hypervariable regions of 16S rRNA gene were amplified using an Ion 16S Metagenomics kit (Thermo Fisher, Waltham, Massachusetts, USA). The sequencing of 400 bp amplicons was performed using the Ion Torrent PGM by CeMIA SA (https://cemia.eu/) (Larissa, Greece) (accessed on 17 May 2019). After removing the chimeras and noises, the Ion Reporter software was used to analyze the sequences, and the Nucleotide Basic Local Alignment Search Tool (BLASTn) against the NCBI database (www.ncbi.nlm.nih.gov) (accessed on 9 August 2019) was used to taxonomically classify the operational taxonomic units (OTUs) at >97% similarity level. The GenBank Bioproject ID is PRJNA722121. A t-test was applied to estimate the significant differences (95% confidence level) among OTU abundances. In addition, Venn diagrams were prepared using the online platform Venn 2.1 [29] to estimate the bacterial relationships at the family, genus, and species level shared by the two cheese batches.
3. Results and Discussion
3.1. Microbiological Attributes and pH Variation of Retail Galotyri-Like Acid-Curd Cheeses
The results of the microbial quantification analyses along with the pH values of the five cheese batches are shown in Table 1. Results are presented separately for each batch because, in many cases, populations grown on the same media varied greatly (Table 1). This indicated a high microbiological variability between the cheese batches, which prevented their treatment as replicate samples for statistical analyses. Nevertheless, to specify general trends, the mean and standard deviation values are also presented in Table 1.
Starting from the pH data, mixing of fresh natural Greek yogurt curds (pH 3.7–4.0; according to the routine plant measurements) with ripened ‘Myzithrenio’ whey cheese granules (pH 4.0–4.5; as above) at an approximate 2:1 ratio resulted in acidic (pH 4.11 ± 0.25) RTE Galotyri-like acid-curd cheese products, as desired technologically. However, the high variation in the resultant pH between batches (Table 1) indicated that this acid-curd cheese is a nonstandardized delicatessen dairy product.
Microbiologically, LAB prevailed in all batches followed by yeasts (Table 1). This finding was in general accordance with previous quantification data of the total LAB and yeast population levels in Galotyri PDO [10,11,12] and other traditional (Greek) acid-curd cheese products [6,7,8,9]. Particularly acid-tolerant spoilage yeasts are favored for growth under the low-pH conditions prevailing in fresh acid-curd cheeses after completion of the LAB fermentation process, and they most often become the terminal spoilers of traditional, fresh or ripened, acid-curd cheeses stored under refrigeration [1,10,30].
Major batch-to-batch variations existed in the populations of total thermophilic and/or mesophilic LAB grown on MPCA, M17, and MRS agars at incubation temperatures from 22 to 45 °C. Specifically, batches A, B, and E had higher LAB populations on M17 agar at 42 °C than batches C and D. Furthermore, batches A and B had the highest total viable counts on MPCA at 37 °C, as opposed to batch D which had the lowest total viable counts.
Overall, the LAB quantification results suggested that the natural yogurt starter strain(s) of S. thermophilus prevailed in batches A, B, and E, since S. thermophilus and other streptococci are favored for growth on M17 and MPCA agars at 37 to 45 °C. Conversely, the populations of mesophilic dairy LAB, whose growth is favored on M17 agar at 22 °C and MRS agar at 30 °C, were more stable within 7 to 8 log CFU/g in all cheese batches (Table 1). Mesophilic LAB in Galotyri-like acid-curd cheese likely originated from the ripened ‘Myzithrenio’ whey cheese rather than from the fresh yogurt curd, which should be a synergic natural culture of S. thermophilus and thermophilic lactobacilli [20,23,24].
However, major and quite randomized fluctuations also existed in the level of thermophilic Lactobacillus selectively enumerated on MRS agar at 45 °C. Their levels ranged from 7 to 8 log CFU/g in batches B, D, and mainly A, whereas batches E and C showed equal to lower thermophilic LAB levels, respectively, than those of enterococci on SB agar at 37 °C (Table 1). In previous studies, enterococci were the only LAB group isolated from MRS agar at 45 °C when thermophilic dairy lactobacilli in milk or cheese were few or absent [31]. Compared to the other LAB genera, enterococcal populations were subdominant by 1–2 log units in all cheese batches (Table 1). However, their counts (6.0–6.8 log CFU/g) were still high for a Galotyri-like acid-curd (pH < 4.5) cheese [11].
Total staphylococci and aerobic Gram-negative, Pseudomonas-like bacteria were below 2 log CFU/g. Coliforms and pathogenic (RFP+) staphylococci were below 1 log CFU/g in all cheese batches. Moreover, neither Salmonella nor Listeria spp. were detected in 25 g samples of cheese after culture enrichment. Hence, the hygienic quality and safety of all RTE cheese products was fairly good. The potent presence of hemolytic and/or antibiotic-resistant strains within the Enterococcus biota was the only concern [32,33]. Indeed, several previous E. faecalis isolates from PDO Galotyri cheese harbored the cytolysin gene, while one E. faecalis strain genotype was strongly β-hemolytic [33]. Thus, regardless of their strong antilisterial activity, all previous E. faecalis isolates from Galotyri cheese were a priori excluded from dairy (food) applications.
3.2. Biochemical Characterization and Distribution of the LAB Biota in Galotyri-Like Cheeses
In total, 145 out of the 150 isolates recovered from the five cheese batches were Gram-positive, catalase-negative, nonsporogenic bacteria, presumably LAB (Table 2). The remaining five colonies were catalase-positive bacteria or yeasts isolated from the cheese batches C and D (Table 2 and Table 3).
Table 2.
Biochemical characterization and grouping of the 145 LAB isolates from RTE Galotyri-like acid-curd cheese at the genus/subgenus level in accordance to their numerical distribution in each of the five cheese batches analyzed, and their total percent isolation frequency.
Table 3.
Numerical distribution of the 145 LAB isolates from RTE Galotyri-like acid-curd cheese biochemically characterized at the genus/subgenus level and the remaining five non-LAB or yeast isolates in association with the selectivity of their enumeration/isolation agar media.
The 145 LAB isolates were assigned to eight distinct biochemical groups at the genus level on the basis of the microscopic appearance and ability of each isolate to produce CO2 from glucose and NH3 from arginine, as well as to grow at 15 and/or 45 °C, in 6.5% NaCl, and on KAA agar (Table 2). According to their reactions, most (30.3%) of the isolates were characterized as Enterococcus followed by mesophilic, facultative heterofermentative Lactobacillus (23.4%), thermophilic Streptococcus (15.9%), and thermophilic homoferementative Lactobacillus (10.3%). Fewer isolates belonged to the genera Lactococcus, Leuconostoc, or Weissella and/or the obligatory heterofermentative Lactobacillus group (Table 2). However, at this point, it needs to be emphasized that the percentage isolation frequency of each LAB group in Table 2, calculated on the basis of the net number of five isolates from each of the first six LAB-selective enumeration agar media in Table 1, does not represent the actual population density of each same LAB group in each of the five cheese batches. Indeed, enterococci had the highest isolation frequency overall (Table 2) despite the fact that their populations were subdominant of the populations of most other LAB genera by 1–2 log units in all batches (Table 1). However, half of the Enterococcus isolates were collected from their selective SB agar plates, while the other half were random isolates from the MPCA/37 °C, M17/22 °C, MRS/30 °C, and mainly the MRS/45 °C agar plates. It is well documented that dairy (lactose-fermenting) enterococci promote excellent growth on all the above agar media at any incubation temperature in the range from 15 to 45 °C [31].
Therefore, to realize the importance of the media selectivity effects, the isolates of each of the cheese LAB genera or groups in Table 2 were tabulated in accordance with their growth agar medium (Table 3). Clearly, Enterococcus isolates were collected from all six isolation agar media, in full contrast to thermophilic lactobacilli isolated from MRS/45 °C agar only. Thermophilic streptococci, inclusive of S. thermophilus, were isolated from M17/42 °C and less often from the MPCA/37 °C agar plates only, whereas lactococci were isolated from all media, except for MRS/45 °C and SB/37 °C (Table 3). Notably, three mesophilic Lactobacillus isolates were recovered from SB/37 °C agar. This was not surprising because several mesophilic nonstarter lactobacilli, particularly from the Lb. plantarum group, have been shown to grow with a 24 to 48 h delay on SB (as whitish colonies; discriminated from the red-brown Enterococcus colonies) and KAA (as black colonies, like enterococci) at 37 °C, thus compromising the selectivity of the enterococcal media [11,31].
Enterococci and mesophilic lactobacilli, i.e., Lb. plantarum and Lb. casei groups, are among the most resistant nonstarter LAB of cheese, particularly able to survive long ripening periods [24,34,35,36] and grow on all LAB-selective growth media (Table 3) at or below 45 °C [31]. Leuconostoc-like bacteria, gas-forming lactobacilli, and the non-LAB and yeast isolates were recovered from MPCA, M17, and MRS agar plates incubated at 37 °C or below. In summary, MRS/45 °C and MPCA/37 °C were the most and the least LAB-selective agar media, respectively, under the conditions of this study (Table 3).
Based on these considerations, the major batch-to-batch variations noted among the five cheeses with regard to the size of thermophilic and mesophilic LAB populations (Table 1) were reflected in the type and numerical distribution of the LAB genera isolated from each batch (Table 2). Specifically, thermophilic streptococci were isolated from all batches. However, they were dominant in batch A, followed by batches B and E. In contrast, mesophilic lactobacilli were dominant in batch D, whereas batch C was the only cheese without detectable Lactobacillus spp., instead dominated by lactococci and enterococci (Table 2). Enterococci were isolated from all cheese batches. However, their isolations were more numerous from batches C, E, and B for reasons explained above regarding the selectivity of the isolation agar media (Table 3). Leuconostoc-like bacteria prevailed in batch E only. Lastly, in full accordance with their population sizes grown on the selective MRS/45 °C agar plates (Table 1 and Table 3), cultivable thermophilic dairy lactobacilli were only recovered from the batches A, B, and D (Table 2).
On the basis of their sugar fermentation profiles, the 145 cheese LAB isolates were further characterized to the species level and, when possible, they were discriminated at the subspecies or the strain biotype level. The biochemical characterization results, accompanied by the numerical distribution of each LAB species, subspecies, or biotype in the cheese batches A to E, are summarized in Table 4 for Enterococcus, Table 5 for mesophilic LAB (i.e., mesophilic Lactobacillus, Lactococcus, Leuconostoc, and gas-forming Lactobacillus) and Table 6 for thermophilic dairy Streptococcus and Lactobacillus. For table simplification, the biochemical LAB species characterization of each group is listed in the footnote of Table 5 and Table 6.
Table 4.
Biochemical characterization of 44 Enterococcus spp. isolates from five retail RTE Galotyri-like acid-curd cheese batches and their numerical distribution in each cheese batch.
Table 5.
Biochemical characterization of 66 mesophilic LAB isolates from five retail RTE Galotyri-like acid-curd cheese batches and their numerical distribution in each cheese batch.
Table 6.
Biochemical characterization of 38 thermophilic LAB isolates from five retail RTE Galotyri-like acid-curd cheese batches and their numerical distribution in each cheese batch.
Characterization of the 44 Enterococcus isolates at the species level was based on the biochemical identification key of Manero and Blanch [28] and the methodology used by Vandera et al. [24]. Accordingly, the present isolates were assigned to two species only: E. faecium (24 isolates; biotypes A1–A4) and E. faecalis (20 isolates; biotypes B1–B5) (Table 4). The key differentiating characteristic between them was the ability of all E. faecium isolates to ferment l-arabinose, unlike all E. faecalis isolates [28]. Beyond this key differentiation, all typical E. faecium isolates were included in the biotypes A1 and A2, which differed in the ability of the former isolates to ferment sucrose. The most atypical phenotypic feature of the E. faecium biotype-A3 isolates was their inability to ferment trehalose, whereas the single biotype-A4 GLP-217 isolate showed the richest sugar fermentation profile (data not shown), including sorbitol and raffinose (Table 4). Although biochemically identical to E. faecium KE102 genotyped by Vandera et al. [24], this particular isolate from batch E might be an E. gallinarum or E. raffinosus strain; additional biochemical tests and genotyping are required to elucidate this. Regarding E. faecalis, typical mannitol-, sucrose-, and sorbitol-positive isolates of this species were included in biotypes B1 and B3, while biotype-B2 included atypical sucrose-negative isolates. The most atypical sorbitol-negative and/or mannitol-negative E. faecalis isolates were grouped in biotypes B4 and B5, respectively (Table 4). they also require additional biochemical and molecular characterization because they possibly represent very atypical strain biotypes of E. durans or closely related species of the E. faecium/durans genomic group [24,28]. Regarding the numerical distribution of the isolates, E. faecium was more manifest in batch E and E. faecalis was more manifest in batch C. Batches A and B were the least and most diversified cheeses, respectively, in Enterococcus biotypes (Table 4).
Tn total, 26 of the 34 (76.5%) mesophilic Lactobacillus isolates were characterized as rather atypical nonstarter dairy strain biotypes (C1, C3, and C4) of the Lb. plantarum group (Table 5). All fermented melibiose, mannitol, and of course lactose. However, only the single biotype C4 isolate fermented l-arabinose, while it failed to ferment sorbitol. Similarly, several of the l-arabinose-negative isolates of C1 and C3 biotypes failed to ferment sorbitol. Moreover, only the most prevalent l-arabinose-negative C1 biotype fermented raffinose (Table 5). The observed high degree of heterogeneity in the sugar fermentation pattern is uncommon for typical Lb. plantarum strains, which ferment raffinose, sorbitol, and most often l-arabinose [24,26,31]. Hence, biotypes C1, C3, and C4 are presently classified as members of the Lb. plantarum/paraplantarum cluster in Table 5. None of them fermented d-xylose, as Lb. pentosus typically does [26]. Differentiation of Lb. plantarum, Lb. paraplantarum, and Lb. pentosus by phenotypic criteria is difficult; actually, it requires a multiplex PCR assay with specific primers targeting the heterogeneity of the recA gene [24,37]. Lastly, the C2 biotype included eight melibiose-negative isolates which matched the Lb. paracasei strains previously found to prevail in mature Graviera cheeses produced in Pappas plant. Seven isolates did not ferment sorbitol (Table 5), like the indigenous, least prevalent biotype II of Lb. paracasei isolates from mature Graviera cheeses [24,31]. Batch B also was the most diversified batch with regard to the number of mesophilic Lactobacillus biotypes isolated (Table 5).
As mentioned, lactococci were found to be predominant in the cheese batch C only, while they were sporadically isolated from the same growth culture media used in batches A and E. The majority (85.7%) of Lactococcus isolates were arginine-negative, divided in two distinct biotypes, D1 and D2 (Table 5), representing typical industrial CSC starter strains of Lc. lactis subsp. cremoris (D1) and Lc. lactis subsp. lactis (D2), respectively. Conversely, biotype D3 included typical arginine-positive and quite oligo-fermenting Lc. lactis subsp. lactis, which also matched industrial CSC starter strains imported in Greece. Specifically, none of them fermented d-xylose (Table 5), as most wild plant-derived Lc. lactis isolates do [38], including the nisin A-producing Lc. lactis subsp. cremoris genotype (strains M78 and M104) from Greek raw milk [39], currently applied in commercial Graviera cheese production in the Pappas plant [22].
The remaining 15 mesophilic heterofermentative LAB isolates, preliminarily characterized as Leuconostoc-like bacteria, arginine-positive Weissella, or gas-forming Lactobacillus (Table 2), fermented d-xylose strongly (Table 5). Twelve of them (80%) formed a very heterogeneous biochemical group E1 which was assignable to Leuconostoc mesenteroides despite the variability of the isolates in several sugar fermentation reactions [27]. The remaining three were single gas-forming isolates. GL34 (biotype E3) and GLP205 (biotype E4) seemed to be Lb. brevis strain variants. GLP212 (biotype E2) resembled strain GLP205 in most sugar fermentation reactions, including the key negative reactions with trehalose and sucrose; however, it did not show an arginine-positive reaction, which is required for assignment to the Lb. brevis group [26]. Therefore, at present, GLP212 should be considered an unidentified Leuconostoc sp. (Table 5). Molecular taxonomic approaches are required to accurately identify the gas-forming E1–E4 LAB groups at the species level.
All Galotyri-like acid-curd cheese isolates of thermophilic dairy lactobacilli (Table 6) were assigned to Lb. delbrueckii, specifically to the subspecies bulgaricus (Z1) and lactis (Z2), respectively [26]. Probably all were strain constituents of the natural yogurt starter [23,24]. None of the isolates were lactose-negative (Table 6) and, thus, the isolates were unable to be assigned to Lb. delbrueckii subsp. delbrueckii [26]. Moreover, most (87%) streptococci from all batches were typical isolates of S. thermophilus starter strain/s [11] since all fermented lactose and sucrose strongly, while few gave a very weak, practically negative, fermentation reaction with ribose, maltose, and most importantly galactose (biotype H1 in Table 6). The latter finding indicated that the natural yogurt starter used to make the basic yogurt curd in the plant did not contain ‘new-generation’ galactose-positive S. thermophilus starter strains, which were increasingly detected in industrial and small-size traditional Greek dairy plants and products during the last decade [11]. Most probably, these are also imported CSC S. thermophilus strains that gained the ability to ferment galactose via genetic manipulation [40,41,42]. The remaining three Streptococcus (biotypes H2, H3 and H4), also isolated from the most diversified batch B, had more complex sugar fermentation profiles than S. thermophilus biotype H1 (Table 6). They were single isolates of either autochthonous streptococci from animals or indigenous strains of the S. salivarius/thermophilus group derived from the basal yogurt curd or, less likely, the ripened ‘Myzithrenio’ cheese. Additional polyphasic identification tests are in progress to elucidate the species identity and evaluate the hygienic safety of these sporadic Streptococcus-like isolates from the delicatessen Galotyri-like cheese.
3.3. Bacterial Communities Estimated by 16S rRNA Amplicon Sequencing
DNA was directly extracted from two cheese sample portions, the respective Galotyri-like acid-curd cheese batches A and B (Table 1, Table 2, Table 3, Table 4, Table 5 and Table 6), to estimate the bacterial communities present in cheese and figure out potential differences between the two batches. Sequencing analysis revealed the presence of 19 different families. Specifically, 13 of the 19 families had a relative abundance >0.2% in at least one of the two batches (Figure 2), while 18 and 14 of them were detected in batches A and B, respectively (Figure 3a). However, statistical analysis based on the relative abundances of detected OTUs, showed that no significant differences were observed between the two batches (p > 0.05).
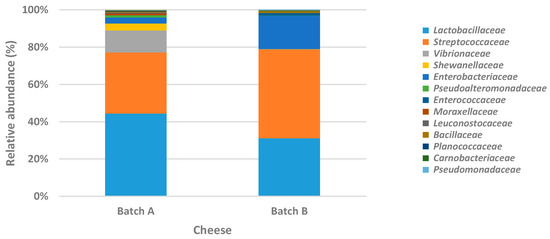
Figure 2.
Abundances of bacterial families (relative abundances, %) detected on the two batches of Galotyri-like acid-curd cheese by next-generation sequencing.
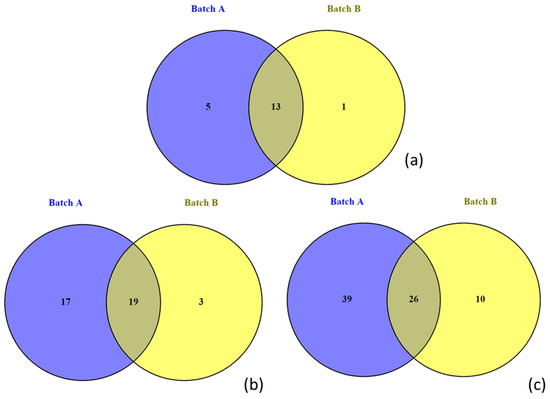
Figure 3.
Venn diagrams showing the OTUs shared between batch A and batch B of cheese at the family (a), genus (b), and species level (c).
Bacterial communities were dominated by Lactobacillaceae and Streptococcaceae in both batches. In brief, these families were detected at 77.21% and 78.76% total relative abundance in batch A and batch B, respectively. However, the relative abundance of Lactobacillaceae was higher in batch A (44.48%) compared to Streptococcaceae (32.73%), which in turn was detected in higher percentage (47.57%) in batch B compared to Lactobacillaceae (31.19%). Moreover, the Gram-negative bacterial families Vibrionaceae (11.77%), Shewanellaceae (3.73%), Enterobacteriaceae (3.29%), and Pseudoalteromonadaceae (1.08%) were also detected in batch A with relative abundance >1%. Conversely, in batch B, the only families apart from Lactobacillaceae and Streptococcaceae with a relative abundance >1% were Enterobacteriaceae (17.85%) and Enterococcaceae (1.31%). In Figure 3b,c, the number of shared OTUs between the two batches is illustrated at the genus and species level, respectively. The relative abundances of all detected genera and species are shown in Table S1 (Supplementary Materials). From the Venn diagrams, it seems that the bacterial diversity of batch A was higher than that of batch B; however, the percentage of the most of these genera or species was low (< 1%) (Figure 4). In brief, the number of OTUs shared between batch A and batch B marked on the overlapped area in Venn diagrams represents the number of the most common detected families, genera, or species on both batches of cheese. Here, it should be noted that the assigned family or genus names are mentioned throughout the text instead of the genus or species names in the case that sequencing analysis succeeded only up to family or genus level, respectively.
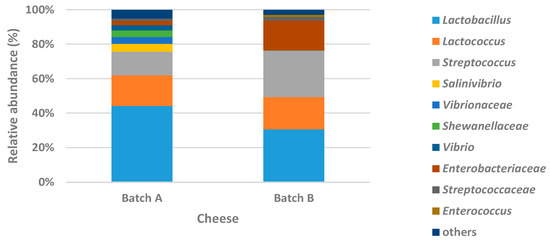
Figure 4.
Abundances of bacterial genera detected on the two batches of cheese (relative abundance >1%) by next-generation sequencing. Family names are shown in the case that sequencing analysis succeeded only up to family level. The genera with relative abundances <1% are presented in the figure as “others”.
Considering the prevalence of Lactobacillaceae and Streptococcaceae families, the genera Lactobacillus, Streptococcus, Lactococcus, and marginally Pediococcus (0.03% in batch A only) were detected (Figure 4). In the case of the Lactobacillaceae family, Lb. delbrueckii was the most dominant species (43.62% and 29.4%) in both batches A and B, respectively (Table S1, Supplementary Materials). In addition, Lb. helveticus was detected marginally (0.07%) in batch B only. The remaining percentage of Lactobacillaceae failed to be identified at the genus (0.25% and 0.5%) or species (0.58% and 1.22%) level in batch A and B, respectively. Moreover, high abundances of Streptococcus (13.69% and 26.81%) and Lactococcus (17.81% and 18.72%) were observed in batches A and B, respectively (Figure 4). S. thermophilus (11.03% and 17.75% in batches A and B, respectively) was the most abundant Streptococcus species detected in both batches (Table S1, Supplementary Materials). Beyond the unidentified percentages of 2.57% and 8.91% in batch A and B, respectively, assigned to the Streptococcus genus, the species Streptococcus parauberis, Streptococcus uberis, and Streptococcus gallolyticus were detected in low abundances (<0.1%). Regarding the genus Lactococcus, the primary species Lactococcus lactis was identified in batches A (5.05%) and B (7.82%). In addition, the species Lactococcus chungangensis, Lactococcus raffinolactis, and unidentified Lactococcus sp. were also detected in low abundances (<0.1%). However, a high percentage of the Lactococcus genus, ranging from 10.43% to 12.41%, failed to be identified at the species level.
Regarding the rest detected bacteria in relative high abundances (>1%) (Figure 4), Salinivibrio, unidentified members of Vibrionaceae, mainly Vibrio sp., Shewanellaceae, and Enterobacteriaceae constituted most of the bacterial community of batch A, together with Lactobacillaceae and Streptococcaceae. However, all former Gram-negative bacteria, except for Enterobacteriaceae, were detected in relatively low abundances (total <1%) in batch B. Additionally, the Enterococcus genus was marginally detected (0.03%) in batch A. In contrast, Enterococcus (1.28%) and mainly Enterobacteriaceae (17.30%), along with Lactobacillaceae and Streptococcaceae, constituted 96.85% of total bacterial community detected in batch B (Figure 4).
To compare the results of the 16S rRNA amplicon sequencing approach based on total bacterial DNA extraction directly from cheese, with the classical microbiological approach based on total and selective bacterial enumeration and characterization, the main findings of both approaches are summarized in Table 7. The relative abundances of the genera and species detected at >0.1% in at least one of the cheese batches (A or B) are compared with the respective relative population density (RPD, %) of each genus or main species found in the two cheese batches (Table 7); the RPD values were calculated as described previously [43]. Overall, the greatest difference between the two methods was the sole detection of LAB genera and species using a culture-dependent method, while additional non-LAB bacterial families/genera were detected by the culture-independent approach in both cheese batches (Table 7). This state could be attributed to (a) the lowest detection limit of 100 cells/g of the remaining bacterial populations being theoretically equal to an RPD < 0.000005% compared to the maximum total viable (LAB) population of 8.6 log CFU/g in both batches (Table 1), and (b) the primary observation that most of the above culture-independent detections of non-LAB (mainly of the Gram-negative families or genera) (Figure 2, Figure 3 and Figure 4; Table S1, Supplementary Materials) were probably associated with bacterial DNA extracted or released from bacterial cells that had already died off. Otherwise, members of the Enterobacteriaceae family, found to be amongst the most prevalent (17.30%) constituents of the bacterial community in batch B on the basis of OTUs (Table 7), should have been detected at levels above 6 log units on VRB/37 °C agar (Table 1). Their detection as viable counts should have been high even in the case that they were not coliforms but lactose-negative enterobacteria. However, the VRB/37 °C and CFC/25 °C agar plates of all batches were free (<100 cells/g) of Gram-negative bacterial growth, including batch A (pH 3.84) and batch B (pH 4.09) (Table 1) subjected to metagenomic analysis. Evidently, the fresh low-pH yogurt curds made using boiled ewe’s milk with natural yogurt starter soon before Galotyri-like acid-curd cheese preparation did not contain Gram-negative bacteria. However, based on the whey cheese literature [44,45], natural contamination and growth levels of Enterobacteriaceae and other Gram-negative bacteria during natural fermentation and ripening of the traditional ‘Myzithrenio’ in wooden cheese barrels might have been quite high. In particular, the growth of salt-tolerant Vibrio and Salinivibrio might have been favored after the fresh whey cheese was dry-salted with 3% salt before transfer to the barrels. Furthermore, wooden cheese barrels have pores and are difficult to clean and disinfect; hence, in addition to various starter or nonstarter LAB, a great diversity of natural Gram-negative and Gram-positive non-LAB contaminants could be transferred to and grow early in fresh ripening cheese [6,19]. Acid-sensitive Gram-negative bacteria decline progressively during feta cheese ripening and may die off later [19,46], as they probably did in the acidified ‘Myzithrenio’ cheese after 1 month of ripening followed by 3–4 months of refrigerated VP storage. In general agreement, a recent microbiological and metagenomic study of feta cheese PDO matured in plastic or stainless-steel containers showed that the abundances of Enterobacteriaceae (which were high in the fresh cheese), Moraxellaceae, and Pseudomonadaceae were numerically reduced in the mature cheese after 120 days of storage [47]. Lactococcus was the most abundant genus in both fresh and mature industrial feta cheeses [47], produced with CSCs that probably contained L. lactis in addition to S. thermophilus and Lb. delbrueckii subsp. bulgaricus. In this study, CSC-like L. lactis strains were dominant in batch C only (Table 2), not selected for metagenomic analysis. Nonetheless, those L. lactis strains were probably acquired from the feta cheese barrels or other fresh whey cheese contamination sources in the plant. Moreover, the ‘Myzithrenio’ whey cheese used to make batch C might have been less ripened to harbor more Lc. lactis survivors compared to the other batches (Table 2 and Table 7). During ripening of traditional Greek cheeses, the levels of Lc. lactis (starter) strains decline proportionally to the mesophilic nonstarter Lb. casei/paracasei and Lb. plantarum/paraplantarum groups [11,19,31], recently reclassified as two separate genera, Lacticaseibacillus and Lactiplantibacillus, following splitting of the large diverse genus Lactobacillus [48]. Additionally, many industrial CSC Lc. lactis strains have been genetically modified for autolysis in cheese to enhance flavor development via the release of intracellular enzymes during ripening [49,50]. The latter property might correlate with the low RPD (1.58%) or the absence (no isolates) of Lc. lactis in batches A and B, respectively, despite the relative abundance of Lactococcus, Lc. lactis in particular, being fairly high in both batches (Table 7). Probably most Lactococcus had also died off during ‘Myzithrenio’ ripening, as well as during the previous heating of the Pappas Graviera whey, obviously enriched in CSC Lc. lactis strains [22,31].
Table 7.
Prevalence of the main bacterial genera and species in the two retail RTE Galotyri-like acid-curd cheese batches based on their relative abundances of detected OTUs versus their relative population density (RPD) values calculated on the basis of microbial enumeration data.
Regarding the dominant thermophilic yogurt-starter LAB in Galotyri-like acid-curd cheese, the metagenomic analysis and classical microbiological results followed similar trends, despite certain inconsistencies of the two methods between the two batches (Table 7). According to the classical method, S. thermophilus was predominantly isolated as the sole Streptococcus species from batch A (66.56%), while, in batch B, S. thermophilus occurred together with other thermophilic Streptococcus (total RPD 58.47%), which probably belong to the species listed in Table S1 (Supplementary Materials). Furthermore, Lb. delbrueckii was more prevalent in batch A (RPD 16.6%) than in batch B (RPD 3.4%) culture-dependently. The above differences between the two cheese batches were also detected culture-independently. However, particularly for Lb. delbrueckii, the related RPD percentages were always much lower than the relative abundances in both cheese batches, probably because the populations of Lb. delbrueckii solely isolated from MRS/45 °C agar anaerobically (Table 1, Table 2 and Table 3) were lower than the actual prevalence of this species in batches A and B. Lb. delbrueckii may occur at highly viable, nonculturable levels on MPCA and MRS agars incubated at 37 °C or lower growth temperatures, under aerobic conditions. Conversely, detection of Enterococcus was higher in batch B than batch A, and vice versa for Leuconostocs, using both methods (Table 7). However, the percentage RPD values of the above two genera were higher than the corresponding percentage OTUs in both batches (Table 7). The death of the non-LAB groups discussed above and the fact that growth of leuconostocs and mainly enterococci was enhanced by the MRS and SB agar selectivity (Table 3) increased their RPD values. Likewise, the percentage RPD values of mesophilic Lactobacillus were always higher than the abundance of the remaining Lactobacillus species, excluding Lb. delbrueckii, in both batches. Surprisingly, neither Lacticaseibacillus (Lb.) paracasei nor Lactiplantibacillus (Lb.) plantarum/paraplantarum were identified culture-independently, despite their joint RPD values being 9.71% and 20.50% in batches A and B, respectively (Table 7).
In summary, the biochemical identification and metagenome analyses of the LAB species present in this retail delicatessen Galotyri-like cheese product based on yogurt corroborate the findings of Ivanov et al. [20], who reported different (batch-dependent) high abundances of S. thermophilus and Lb. delbrueckii subsp. bulgaricus in Bulgarian home-made yogurts, accompanied by batch-dependent low abundances of several subdominant LAB species, including Lc. lactis, Lc. garvieae, Pediococcus acidilactici, Lb. paracasei, Leuc. mesenteroides, Lb. helveticus, Lb. rhamsosus, and wild Streptococcus. Commercial starter strains of S. thermophilus and Lb. delbrueckii subsp. bulgaricus were isolated nearly as a pure synergic culture from an industrial Galotyri PDO product labeled as a fresh cheese combining the nutritional properties of yogurt [11]. Conversely, the artisan-type Galotyri PDO cheese varieties were far more diversified in terms of LAB ecology, containing (as percentage of total LAB isolates) strains of Lc. lactis (19.8%), Lb. plantarum (16.9%), S. thermophilus (14.7%), Leuc. mesenteroides group (11.3%), Pediococcus inopinatus (7.3%), E. faecalis (8.5%), E. faecium (6.2%), E. durans (5.1%), Lb. rhamnosus (3.4%), Lb. gasseri (3.4%), Lb. bulgaricus (2.8%), and Lb. acidipiscis (0.6%). The prevalence of each species in artisan Galotyri cheeses was also product- and batch-dependent, while it was difficult to discriminate which of the strains were commercial starters or cultures in addition to the primary starter species S. thermophilus and Lc. lactis [11]. Limited identification data exist regarding the LAB species diversity in traditional Greek PDO acid-curd cheeses other than Galotyri [6,19]. Specifically, strains of Lc. lactis subsp. lactis, Leuc. paramesenteroides, and Lb. plantarum were most frequently isolated from Anevato, while Lb. plantarum and Lb. casei dominated in Kopanisti, followed by Pediococcus pentosaceous and enterococci [6]. Conversely, E. durans was the most frequently found species (27.5%) in Katiki, followed by Lb. plantarum (17.6%), E. pseudoavium (13.7%) and many other Enterococcus, Leuconostoc, Weissella, and Lactobacillus spp. at low isolation frequencies [19]. Altogether, our present and previous data and the preceding discussion justify that the microbial (LAB) ecology of the retail delicatessen acid-curd cheese analyzed during this study is very similar to that of PDO Galotyri produced with thermophilic natural or commercial yogurt starters.
4. Conclusions
In conclusion, the classical microbiological methods and the metagenomic analyses were in general agreement with regard to the dominant LAB families, genera, or species that remained viable during the quantification and isolation of the Galotyri-like acid-curd cheese biota. However, results were variable with regard to many other bacterial (LAB) genera or species that had died off in the cheeses; their DNA interfered with the DNA of the viable bacterial cells. Therefore, the total bacterial DNA-based metagenomic method used in this study seems a very powerful approach for mapping the microbial ecology and diversity of fresh foods, such as raw milk and raw milk cheese [51] and fresh meat [52], but it has obvious limitations when processed foods, which include preceding bacterial inactivation steps, are to be analyzed. In the case of Galotyri-like acid-curd cheese, the basal yogurt curd was fresh. However, the ‘Myzithrenio’ whey cheese had previously undergone several bacterial inactivation treatments, namely, salting, fermentation, and ripening, as well as boiling of the Graviera whey to obtain the fresh whey cheese mass, boiling of the 5% raw ewe’s milk supplemented in the whey plus pasteurization or thermization of the original raw milk, and cooking of the whey during Graviera cheese production. Nevertheless, this delicatessen Galotyri-like acid-curd cheese product appears to be a rich pool of indigenous nonstarter strain biotypes of the new genera Lactiplantibacillus and Lacticaseibacillus and of the species E. faecium, E. faecalis, and Leuconostoc mesenteroides derived from the naturally ripened whey cheese used for its manufacture. Additional studies are in progress to evaluate the technological, safety, and antipathogenic and potential probiotic properties of selected Galotyri-like acid-curd cheese LAB isolates.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/fermentation7020067/s1: Table S1. Abundances of bacterial genera (A) (relative abundances, %) and species (B) detected on the two batches of cheese by next-generation sequencing. Family names are shown in the case that sequencing analysis succeeded only up to the family level.
Author Contributions
Conceptualization, J.S.; cheese preparation, D.P.; methodology, V.B., A.I.D. and J.S; formal analysis, A.K., V.B., A.I.D. and J.S.; resources, J.S.; data curation, J.S. and A.I.D.; writing—original draft preparation, A.I.D. and J.S.; writing—review and editing, A.I.D. and J.S.; Project Administration, J.S. All authors have read and agreed to the published version of the manuscript.
Funding
Funding of this study was provided by the European Union and Greek national funds through the EPAnEK 2014-2020 Operational Program Competitiveness, Entrepreneurship, and Innovation under RESEARCH-CREATE-INNOVATE (project T1EDK-00968; project acronym BIO TRUST). The metagenomic analysis and V.B.’s support were funded by ProMedFoods (ARIMNet 2, 2nd Transnational Call, 2016; proposal ID 9028; Grant No. 618127).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study, as well as the bacterial strains identified, are available on request by from the corresponding authors. The GenBank 16S metagenomic Bioproject ID is PRJNA722121.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Guinee, T.P.; Pudja, P.D.; Farkye, N.Y. Fresh acid-curd cheese varieties. In Cheese Chemistry, Physics and Microbiology, 1st ed.; Springer: Boston, MA, USA, 1993; Volume 2, pp. 363–419. [Google Scholar]
- Gérard, A.; El-Hajjaji, S.; Niyonzima, E.; Daube, G.; Sindic, M. Prevalence and survival of Listeria monocytogenes in various types of cheese-A review. Int. J. Dairy Technol. 2018, 71, 825–843. [Google Scholar] [CrossRef]
- Ho, T.M.; Howes, T.; Bhandari, B.R. Methods to extend the shelf-life of cottage cheese—A review. Int. J. Dairy Technol. 2016, 69, 313–327. [Google Scholar] [CrossRef]
- Bello, B.D.; Cocolin, L.; Zeppa, G.; Field, D.; Cotter, P.D.; Hill, C. Technological characterization of bacteriocin producing Lactococcus lactis strains employed to control Listeria monocytogenes in Cottage cheese. Int. J. Food Microbiol. 2012, 153, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Anonymous. Cheeses of protected denomination of origin. In Hellenic Code of Food and Beverages, 3rd ed.; Ministry of Finance, National Publishing Office: Athens, Greece, 2014; Article 83D; pp. 14–59. [Google Scholar]
- Litopoulou-Tzanetaki, E.; Tzanetakis, N. Microbiological characteristics of Greek traditional cheeses. Small Rum. Res. 2011, 101, 17–32. [Google Scholar] [CrossRef]
- Xanthopoulos, V.; Polychroniadou, A.; Litopoulou-Tzanetaki, E.; Tzanetakis, N. Characteristics of Anevato cheese made from raw or heat-treated goat milk inoculated with a lactic starter. Leben. Wissen. Technol. 2000, 33, 483–488. [Google Scholar] [CrossRef]
- Karali, F.; Georgala, A.; Massouras, T.; Kaminarides, S. Volatile compounds and lipolysis levels of Kopanisti, a traditional Greek raw milk cheese. J. Sci. Food Agric. 2013, 93, 1845–1851. [Google Scholar] [CrossRef]
- Papageorgiou, D.K.; Abrahim, A.; Bori, M.; Doundounakis, S. Chemical and Bacteriological Characteristics of Pichtogalo Chanion Cheese and Mesophilic Starter Cultures for Its Production. J. Food Prot. 1998, 61, 688–692. [Google Scholar] [CrossRef] [PubMed]
- Samelis, J.; Kakouri, A. Microbial and safety qualities of PDO Galotyri cheese manufactured at the industrial or artisan scale in Epirus, Greece. Ital. J. Food Sci. 2007, 19, 91–99. [Google Scholar]
- Samelis, J.; Kakouri, A. Major technological differences between an industrial-type and five artisan-type Greek PDO Galotyri market cheeses as revealed by great variations in their lactic acid microbiota. AIMS Agric. Food 2019, 4, 685–710. [Google Scholar] [CrossRef]
- Sameli, N.; Skandamis, P.N.; Samelis, J. Application of Enterococcus faecium KE82, an enterocin A-B-P-producing strain, as an adjunct culture enhances inactivation of Listeria monocytogenes during traditional protected designation of origin Galotyri processing. J. Food Prot. 2021, 84, 87–98. [Google Scholar] [CrossRef]
- Rogga, K.; Samelis, J.; Kakouri, A.; Katsiari, M.; Savvaidis, I.; Kontominas, M. Survival of Listeria monocytogenes in Galotyri, a traditional Greek soft acid-curd cheese, stored aerobically at 4 °C and 12 °C. Int. Dairy J. 2005, 15, 59–67. [Google Scholar] [CrossRef]
- Joishy, T.K.; Dehingia, M.; Khan, M.R. Bacterial diversity and metabolite profiles of curd prepared by natural fermentation of raw milk and back sloping of boiled milk. World J. Microbiol. Biotechnol. 2019, 35, 102. [Google Scholar] [CrossRef]
- Katsiari, M.; Kondyli, E.; Voutsinas, L. Τhe quality of Galotyri-type cheese made with different starter cultures. Food Control 2009, 20, 113–118. [Google Scholar] [CrossRef]
- Kondyli, E.; Katsiari, M.; Voutsinas, L. Chemical and sensory characteristics of Galotyri-type cheese made using different procedures. Food Control. 2008, 19, 301–307. [Google Scholar] [CrossRef]
- McSweeney, P.L.H.; Sousa, M.J. Biochemical pathways for the production of flavor compounds in cheeses during ripening: A review. Lait 2000, 80, 293–324. [Google Scholar] [CrossRef]
- Kondyli, E.; Massouras, T.; Katsiari, M.; Voutsinas, L. Lipolysis and volatile compounds of Galotyri-type cheese made using different procedures. Small Rumin. Res. 2013, 113, 432–436. [Google Scholar] [CrossRef]
- Litopoulou-Tzanetaki, E.; Tzanetakis, N. The Microfloras of Traditional Greek Cheeses. Microbiol. Spectr. 2014, 2, CM-0009-2012. [Google Scholar]
- Ivanov, I.; Petrov, K.; Lozanov, V.; Hristov, I.; Wu, Z.; Liu, Z.; Petrova, P. Bioactive compounds produced by the accompanying microflora in Bulgarian yogurt. Processes 2021, 9, 114. [Google Scholar] [CrossRef]
- Samelis, J.; Argyri, A.; Doulgeraki, A.; Koukkou, A.I.; Skandamis, P.; Athanasoulas, A.; Pappas, D.; Chorianopoulos, N. De-velopment of novel biofunctional foods and total quality enhancement of traditional dairy products by suitable management of their microbial ecology—BIO TRUST. In Proceedings of the 26th International ICFMH Conference FoodMicro, Freie Universität, Berlin, Germany, 3–6 September 2018; Volume P1.62, p. 152. [Google Scholar]
- Noutsopoulos, D.; Kakouri, A.; Kartezini, E.; Pappas, D.; Hatziloukas, E.; Samelis, J. Growth, nisA gene expression and in situ activity of novel Lactococcus lactis subsp. cremoris costarter culture in commercial hard cheese production. J. Food Prot. 2017, 80, 2137–2146. [Google Scholar] [CrossRef]
- Tamine, A.Y.; Robinson, R.K. Microbiology of yoghurt starter cultures. In Yoghurt Science and Technology, 1st ed.; Tamine, A.Y., Robinson, R.K., Eds.; Pergamon Press: Oxford, UK, 1985; pp. 276–294. [Google Scholar]
- Vandera, E.; Kakouri, A.; Koukkou, A.I.; Samelis, J. Major ecological shifts within the dominant nonstarter lactic acid bacteria in mature Greek Graviera cheese as affected by the starter culture type. Int. J. Food Microbiol. 2019, 290, 15–26. [Google Scholar] [CrossRef]
- Samelis, J.; Lianou, A.; Kakouri, A.; Delbès, C.; Rogelj, I.; Bogovič-Matijašić, B.; Montel, M.-C. Changes in the Microbial Composition of Raw Milk Induced by Thermization Treatments Applied Prior to Traditional Greek Hard Cheese Processing. J. Food Prot. 2009, 72, 783–790. [Google Scholar] [CrossRef]
- Hammes, W.P.; Hertel, C. Genus I Lactobacillus Beijernick 1901, 212AL. In Bergey’s Manual of Systematic Bacteriology, the Fir-micutes, 2nd ed.; Whitman, W.B., Ed.; Springer: New York, NY, USA, 2009; Volume 3, pp. 465–511. [Google Scholar]
- Holzapfel, W.H.; Bjorkroth, J.A.; Dicks, L.M.T. Genus I Leuconostoc van Tieghem 1878, 198AL. In Bergey’s Manual of Systematic Bacteriology, the Firmicutes, 2nd ed.; Whitman, W.B., Ed.; Springer: New York, NY, USA, 2009; Volume 3, pp. 624–635. [Google Scholar]
- Manero, A.; Blanch, A.R. Identification of Enterococcus spp. with a biochemical key. Appl. Environ. Microbiol. 1999, 65, 4425–4430. [Google Scholar] [CrossRef] [PubMed]
- Oliveros, J.C. Venny. An Interactive Tool for Comparing Lists with Venn Diagrams. 2007. Available online: http://bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 20 November 2013).
- Tofalo, R.; Fusco, V.; Böhnlein, C.; Kabisch, J.; Logrieco, A.F.; Habermann, D.; Cho, G.-S.; Benomar, N.; Abriouel, H.; Schmidt-Heydt, M.; et al. The life and times of yeasts in traditional food fermentations. Crit. Rev. Food Sci. Nutr. 2019, 60, 3103–3132. [Google Scholar] [CrossRef] [PubMed]
- Samelis, J.; Kakouri, A.; Pappa, E.C.; Matijašić, B.B.; Georgalaki, M.D.; Tsakalidou, E.; Rogelj, A. Microbial Stability and Safety of Traditional Greek Graviera Cheese: Characterization of the Lactic Acid Bacterial Flora and Culture-Independent Detection of Bacteriocin Genes in the Ripened Cheeses and Their Microbial Consortia. J. Food Prot. 2010, 73, 1294–1303. [Google Scholar] [CrossRef] [PubMed]
- Hanchi, H.; Mottawea, W.; Sebei, K.; Hammami, R. The Genus Enterococcus: Between Probiotic Potential and Safety Concerns—An Update. Front. Microbiol. 2018, 9, 1791. [Google Scholar] [CrossRef] [PubMed]
- Vandera, E.; Parapouli, M.; Kakouri, A.; Koukkou, A.-I.; Hatziloukas, E.; Samelis, J. Structural enterocin gene profiles and mode of antilisterial activity in synthetic liquid media and skim milk of autochthonous Enterococcus spp. isolates from artisan Greek Graviera and Galotyri cheeses. Food Microbiol. 2020, 86, 103335. [Google Scholar] [CrossRef] [PubMed]
- Bottari, B.; Levante, A.; Neviani, E.; Gatti, M. How the Fewest Become the Greatest. L. casei’s Impact on Long Ripened Cheeses. Front. Microbiol. 2018, 9, 2866. [Google Scholar] [CrossRef]
- Settanni, L.; Moschetti, G. Non-starter lactic acid bacteria used to improve cheese quality and provide health benefits. Food Microbiol. 2010, 27, 691–697. [Google Scholar] [CrossRef]
- Gobbetti, M.; de Angelis, M.; di Cagno, R.; Mancini, L.; Fox, P.F. Pros and cons for using non-starter lactic acid bacteria (NSLAB) as secondary/adjunct starters for cheese ripening. Trends Food Sci. Technol. 2015, 45, 167–178. [Google Scholar] [CrossRef]
- Torriani, S.; Felis, G.E.; Dellaglio, F. Differentiation of Lactobacillus plantarum, L. pentosus, and L. paraplantarum by recA Gene Sequence Analysis and Multiplex PCR Assay with recA Gene-Derived Primers. Appl. Environ. Microbiol. 2001, 67, 3450–3454. [Google Scholar] [CrossRef]
- Cavanagh, D.; Fitzgerald, G.F.; McAuliffe, O. From field to fermentation: The origins of Lactococcus lactis and its domestication to the dairy environment. Food Microbiol. 2015, 47, 45–61. [Google Scholar] [CrossRef] [PubMed]
- Parapouli, M.; Delbès-Paus, C.; Kakouri, A.; Koukkou, A.-I.; Montel, M.-C.; Samelis, J. Characterization of a Wild, Novel Nisin A-Producing Lactococcus Strain with an L. lactis subsp. Cremoris Genotype and an L. lactis subsp. Lactis Phenotype, Isolated from Greek Raw Milk. Appl. Environ. Microbiol. 2013, 79, 3476–3484. [Google Scholar] [CrossRef]
- de Vos, W.M. Metabolic engineering of sugar catabolism in lactic acid bacteria. Ant. Leeuwen. 1996, 70, 223–242. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, E.E.; van den Bogaard, P.T.C.; Catzeddu, P.; Kuipers, O.P.; de Vos, W.M. Activation of silent gal genes in the lac-gal regulon of Streptococcus thermophilus. J. Bacteriol. 2001, 183, 1184–1194. [Google Scholar] [CrossRef]
- Giaretta, S.; Treu, L.; Vendramin, V.; Duarte, V.D.S.; Tarrah, A.; Campanaro, S.; Corich, V.; Giacomini, A. Comparative Transcriptomic Analysis of Streptococcus thermophilus TH1436 and TH1477 Showing Different Capability in the Use of Galactose. Front. Microbiol. 2018, 9, 1765. [Google Scholar] [CrossRef]
- Samelis, J.; Kakouri, A. Cell Growth Density and Nisin a Activity of the Indigenous Lactococcus lactis subsp. cremoris M78 Costarter Depend Strongly on Inoculation Levels of a Commercial Streptococcus thermophilus Starter in Milk: Practical Aspects for Traditional Greek Cheese Processors. J. Food Prot. 2020, 83, 542–551. [Google Scholar] [CrossRef]
- Pintado, M.E.; Macedo, A.C.; Malcata, F.X. Review: Technology, chemistry and microbiology of whey cheeses. Food Sci. Technol. Int. 2001, 7, 105–116. [Google Scholar] [CrossRef]
- Perin, L.M.; Sardano, M.L.S.; Nero, L.A.; Neviani, E.; Gatti, M. Bacterial ecology of artisanal Minas cheeses assessed by cul-ture-dependent and independent methods. Food Microbiol. 2017, 65, 160–169. [Google Scholar] [CrossRef]
- Manolopoulou, E.; Sarantinopoulos, P.; Zoidou, E.; Aktypis, A.; Moschopoulou, E.; Kandarakis, I.G.; Anifantakis, M.E. Evo-lution of microbial populations during traditional Feta cheese manufacture and ripening. Int. J. Food Microbiol. 2003, 82, 153–161. [Google Scholar] [CrossRef]
- Spyrelli, E.D.; Stamatiou, A.; Tassou, C.C.; Nychas, G.-J.E.; Doulgeraki, A.I. Microbiological and Metagenomic Analysis to Assess the Effect of Container Material on the Microbiota of Feta Cheese during Ripening. Fermentation 2020, 6, 12. [Google Scholar] [CrossRef]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.P.A.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended dscritption of the genus Lactobacillus Beijerinck1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, L.; Morgan, S.M.; Ross, R.P.; Hill, C. Elevated Enzyme Release from Lactococcal Starter Cultures on Exposure to the Lantibiotic Lacticin 481, Produced by Lactococcus lactis DPC5552. J. Dairy Sci. 2002, 85, 2130–2140. [Google Scholar] [CrossRef]
- Lortal, S.; Chapot-Chartier, M.-P. Role, mechanisms and control of lactic acid bacteria lysis in cheese. Int. Dairy J. 2005, 15, 857–871. [Google Scholar] [CrossRef]
- Quigley, L.; O’ Sullivan, O.; Beresford, T.P.; Ross, P.R.; Fitzgerald, G.F.; Cotter, P.D. A comparison of methods used to extract bacterial DNA from raw milk and raw milk cheese. J. Appl. Microbiol. 2012, 113, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Doulgeraki, A.I.; Ercolini, D.; Villani, F.; Nychas, G.-J.E. Spoilage microbiota associated to the storage of raw meat in different conditions. Int. J. Food Microbiol. 2012, 157, 130–141. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).