
N-Glycosylation Site in the Middle Region Is Involved in the Sperm-Binding Activity of Bovine Zona Pellucida Glycoproteins ZP3 and ZP4

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Construction of Recombinant Baculovirus Transfer Plasmids for ZP Proteins
2.2. Expression of Recombinant ZP Proteins
2.3. Purification of Recombinant ZP Proteins from Culture Supernatants
2.4. SDS-PAGE
2.5. Adsorption of Recombinant ZP Proteins to Plastic Wells
2.6. Sperm Binding to Recombinant ZP Proteins Adsorbed to Plastic Wells [13,19]
2.7. Statistical Analysis
2.8. Modeling of bZP Proteins
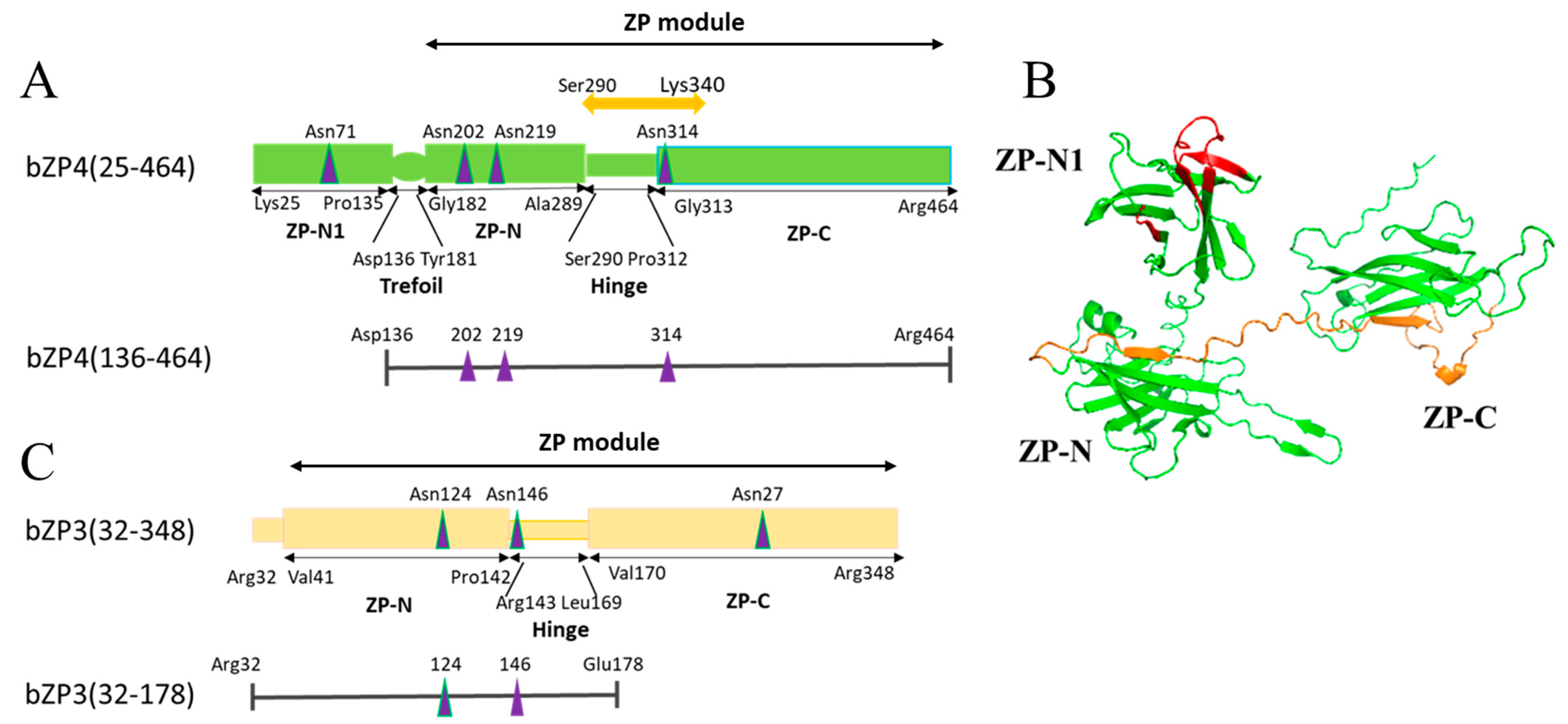
3. Results
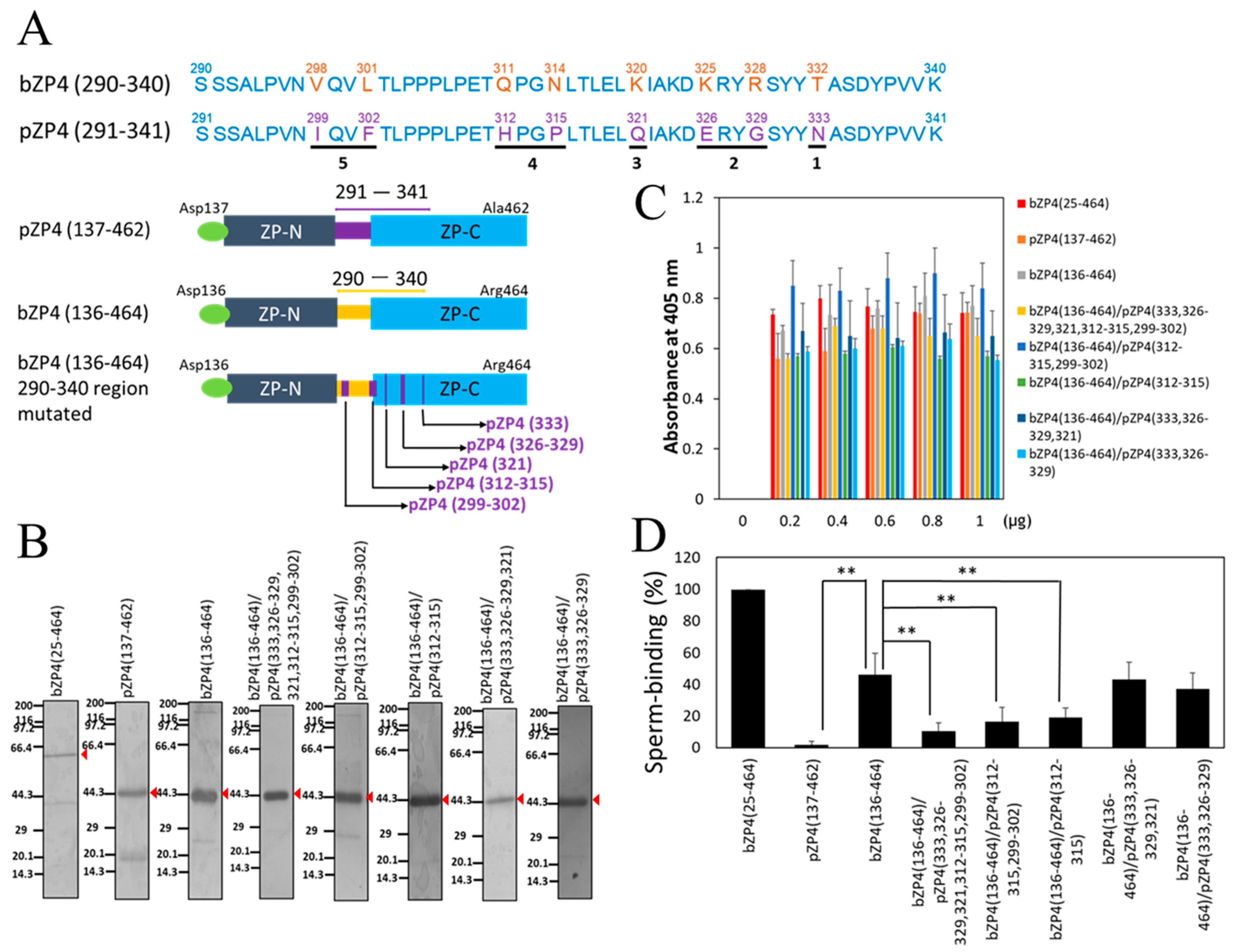
3.1. Identification of Sperm-Binding Sites on bZP4 (290–340)
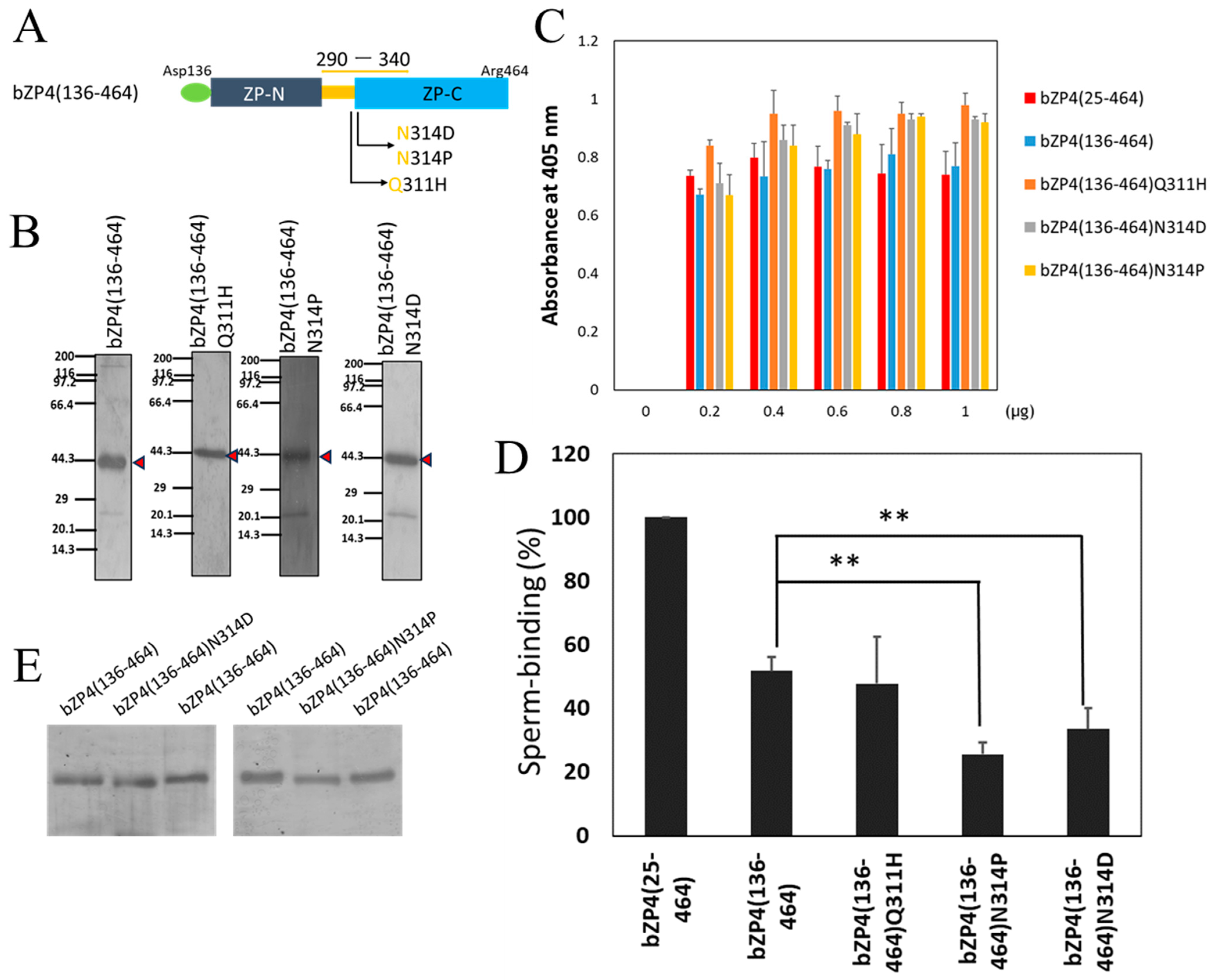
3.2. N-Glycosylation Site on bZP4 (290–340) Is Required for Sperm-Binding Activity
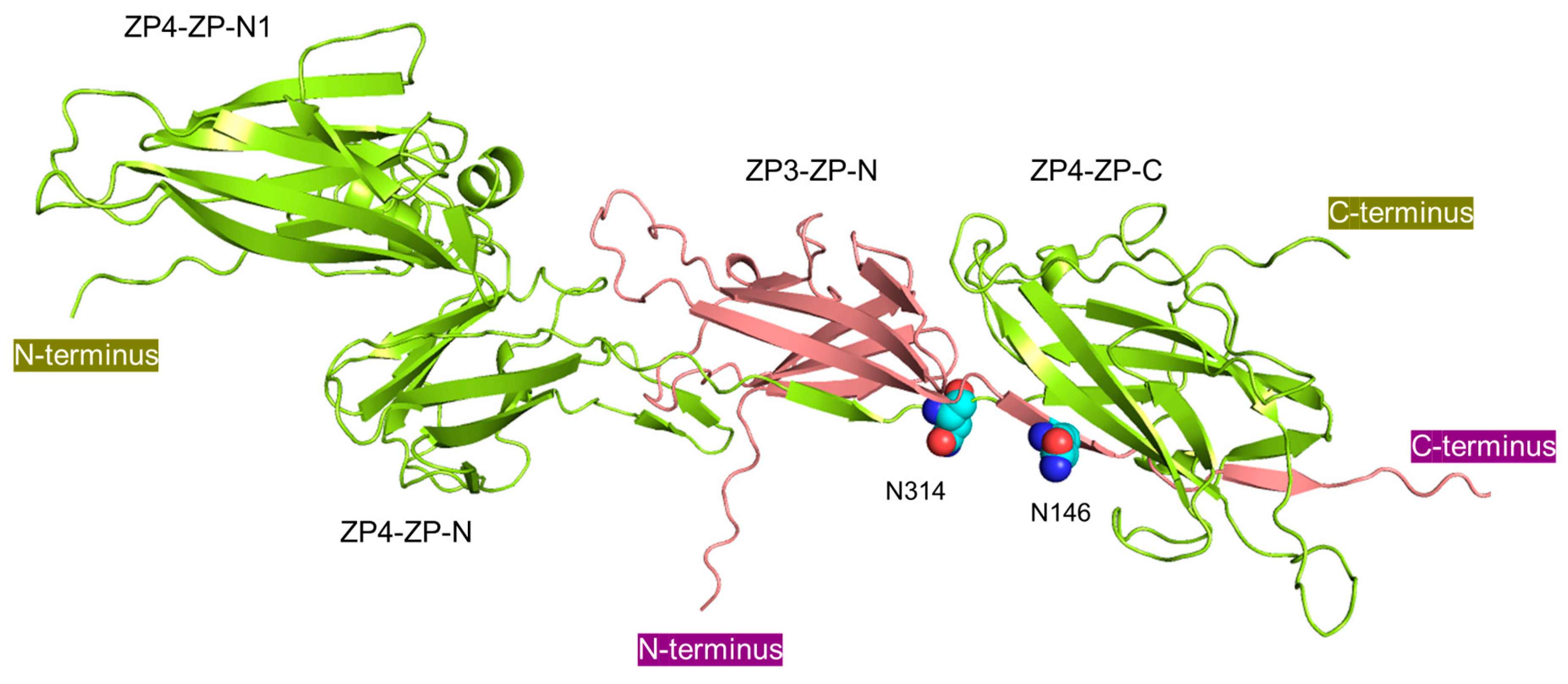
3.3. N-Glycosylation Sites on Both bZP3 and bZP4 Are Involved in Sperm-Binding Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Avella, M.A.; Xiong, B.; Dean, J. The molecular basis of gamete recognition in mice and humans. Mol. Hum. Reprod. 2013, 19, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Wassarman, P.M.; Litscher, E.S. Zona Pellucida Genes and Proteins: Essential Players in Mammalian Oogenesis and Fertility. Genes 2021, 12, 1266. [Google Scholar] [CrossRef] [PubMed]
- Yonezawa, N. Posttranslational modifications of zona pellucida proteins. Adv. Exp. Med. Biol. 2014, 759, 111–140. [Google Scholar] [PubMed]
- Lefièvre, L.; Conner, S.J.; Salpekar, A.; Olufowobi, O.; Ashton, P.; Pavlovic, B.; Lenton, W.; Afnan, M.; Brewis, I.A.; Monk, M.; et al. Four zona pellucida glycoproteins are expressed in the human. Hum. Reprod. 2004, 19, 1580–1586. [Google Scholar] [CrossRef]
- Bleil, J.D.; Wassarman, P.M. Structure and function of the zona pellucida: Identification and characterizaion of the proteins of the mouse oocyte’s zona pellucida. Dev. Biol. 1980, 76, 185–202. [Google Scholar] [CrossRef]
- Noguchi, S.; Yonezawa, N.; Katsumata, T.; Hashizume, K.; Kuwayama, M.; Hamano, S.; Watanabe, S.; Nakano, M. Characterization of the zona pellucida glycoproteins from bovine ovarian and fertilized eggs. Biochim. Biophys. Acta 1994, 1201, 7–14. [Google Scholar] [CrossRef]
- Jovine, L.; Qi, H.; Williams, Z.; Litscher, E.S.; Wassarman, P.M. The ZP domain is a conserved module for polymerization of extracellular proteins. Nat. Cell Biol. 2002, 4, 457–461. [Google Scholar] [CrossRef]
- Jovine, L.; Qi, H.; Williams, Z.; Litscher, E.S.; Wassarman, P.M. A duplicated motif controls assembly of zona pellucida domain proteins. Proc. Natl. Acad. Sci. USA 2004, 101, 5922–5927. [Google Scholar] [CrossRef]
- Han, L.; Monné, M.; Okumura, H.; Schwend, T.; Cherry, A.L.; Flot, D.; Matsuda, T.; Jovine, L. Insights into egg coat assembly and egg-sperm interaction from the X-ray structure of full-length ZP3. Cell 2010, 143, 404–415. [Google Scholar] [CrossRef]
- Avella, M.A.; Baibakov, B.; Dean, J. A single domain of the ZP2 zona pellucida protein mediates gamete recognition in mice and humans. J. Cell Biol. 2014, 205, 801–809. [Google Scholar] [CrossRef]
- Tokuhiro, K.; Dean, J. Glycan-Independent Gamete Recognition Triggers Egg Zinc Sparks and ZP2 Cleavage to Prevent Polyspermy. Dev. Cell 2018, 46, 627–640. [Google Scholar] [CrossRef] [PubMed]
- Yonezawa, N.; Kanai-Kitayama, S.; Kitayama, T.; Hamano, A.; Nakano, M. Porcine zona pellucida glycoprotein ZP4 is responsible for the sperm-binding activity of the ZP3/ZP4 complex. Zygote 2012, 20, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Dilimulati, K.; Orita, M.; Undram, G.; Yonezawa, N. Sperm-binding regions on bovine egg zona pellucida glycoprotein ZP4 studied in a solid supported form on plastic plate. PLoS ONE 2021, 16, e0254234. [Google Scholar] [CrossRef] [PubMed]
- Yonezawa, N.; Kanai, S.; Nakano, M. Structural significance of N-glycans of the zona pellucida on species-selective recognition of spermatozoa between pig and cattle. Soc. Reprod. Fertil. Suppl. 2007, 63, 217–228. [Google Scholar]
- Yonezawa, N.; Amari, S.; Takahashi, K.; Ikeda, K.; Imai, F.L.; Kanai, S.; Kikuchi, K.; Nakano, M. Participation of the nonreducing terminal beta-galactosyl residues of the neutral N-linked carbohydrate chains of porcine zona pellucida glycoproteins in sperm-egg binding. Mol. Reprod. Dev. 2005, 70, 222–227. [Google Scholar] [CrossRef]
- Amari, S.; Yonezawa, N.; Mitsui, S.; Katsumata, T.; Hamano, S.; Kuwayama, M.; Hashimoto, Y.; Suzuki, A.; Takeda, Y.; Nakano, M. Essential role of the nonreducing terminal alpha-mannosyl residues of the N-linked carbohydrate chain of bovine zona pellucida glycoproteins in sperm-egg binding. Mol. Reprod. Dev. 2001, 59, 221–226. [Google Scholar] [CrossRef]
- Kanai, S.; Yonezawa, N.; Ishii, Y.; Tanokura, M.; Nakano, M. Recombinant bovine zona pellucida glycoproteins ZP3 and ZP4 co expressed inSf9 cells form a sperm-binding active heterocomplex. FEBS J. 2007, 274, 5390–5405. [Google Scholar] [CrossRef]
- Bokhove, M.; Nishimura, K.; Brunati, M.; Han, L.; de Sanctis, D.; Rampoldi, L.; Jovine, L. A structured interdomain linker directs self-polymerization of human uromodulin. Proc. Natl. Acad. Sci. USA 2016, 113, 1552–1557. [Google Scholar] [CrossRef]
- Dilimulati, K.; Orita, M.; Yonahara, Y.; Imai, F.L.; Yonezawa, N. Identification of Sperm-Binding Sites in the N-Terminal Domain of Bovine Egg Coat Glycoprotein ZP4. Int. J. Mol. Sci. 2022, 23, 762. [Google Scholar] [CrossRef]
- Suzuki, K.; Tatebe, N.; Kojima, S.; Hamano, A.; Orita, M.; Yonezawa, N. The Hinge Region of Bovine Zona Pellucida Glycoprotein ZP3 Is Involved in the Formation of the Sperm-Binding Active ZP3/ZP4 Complex. Biomolecules 2015, 5, 3339–3353. [Google Scholar] [CrossRef]
- Yonezawa, N.; Kudo, K.; Terauchi, H.; Kanai, S.; Yoda, N.; Tanokura, M.; Ito, K.; Miura, K.; Katsumata, T.; Nakano, M. Recombinant porcine zona pellucida glycoproteins expressed in Sf9 cells bind to bovine sperm but not to porcine sperm. J. Biol. Chem. 2005, 280, 20189–20196. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Brackett, B.G.; Oliphant, G. Capacitation of rabbit spermatozoa in vitro. Biol. Reprod. 1975, 12, 260–274. [Google Scholar] [CrossRef] [PubMed]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold: Making protein folding accessible to all. Nat. Methods 2022, 19, 679–682. [Google Scholar] [CrossRef] [PubMed]
- Yonezawa, N.; Fukui, N.; Kuno, M.; Shinoda, M.; Goko, S.; Mitsui, S.; Nakano, M. Molecular cloning of bovine zona pellucida glycoproteins ZPA and ZPB and analysis for sperm-binding component of the zona. Eur. J. Biochem. 2001, 268, 3587–3594. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Kikuchi, K.; Uchida, Y.; Kanai-Kitayama, S.; Suzuki, R.; Sato, R.; Toma, K.; Geshi, M.; Akagi, S.; Nakano, M.; et al. Binding of sperm to the zona pellucida mediated by sperm carbohydrate-binding proteins is not species-specific in vitro between pigs and cattle. Biomolecules 2013, 3, 85–107. [Google Scholar] [CrossRef]
- Yurewicz, E.C.; Sacco, A.G.; Subramanian, M.G. Structural characterization of the Mr = 55,000 antigen (ZP3) of porcine oocyte zona pellucida. Purification and characterization of alpha- and beta-glycoproteins following digestion of lactosaminoglycan with endo-beta-galactosidase. J. Biol. Chem. 1987, 262, 564–571. [Google Scholar] [CrossRef]
- Yurewicz, E.C.; Hibler, D.; Fontenot, G.K.; Sacco, A.G.; Harris, J. Nucleotide sequence of cDNA encoding ZP3 alpha, a sperm-binding glycoprotein from zona pellucida of pig oocyte. Biochim. Biophys. Acta 1993, 1174, 211–214. [Google Scholar] [CrossRef]
- Stsiapanava, A.; Xu, C.; Brunati, M.; Zamora-Caballero, S.; Schaeffer, C.; Bokhove, M.; Han, L.; Hebert, H.; Carroni, M.; Yasumasu, S.; et al. Cryo-EM structure of native human uromodulin, a zona pellucida module polymer. EMBO J. 2020, 39, e106807. [Google Scholar] [CrossRef]
- Jovine, L. Using machine learning to study protein-protein interactions: From the uromodulin polymer to egg zona pellucida filaments. Mol. Reprod. Dev. 2021, 88, 686–693. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dilimulati, K.; Yulin, Z.; Imai, F.L.; Yonezawa, N. N-Glycosylation Site in the Middle Region Is Involved in the Sperm-Binding Activity of Bovine Zona Pellucida Glycoproteins ZP3 and ZP4. Biomolecules 2023, 13, 1636. https://doi.org/10.3390/biom13111636
Dilimulati K, Yulin Z, Imai FL, Yonezawa N. N-Glycosylation Site in the Middle Region Is Involved in the Sperm-Binding Activity of Bovine Zona Pellucida Glycoproteins ZP3 and ZP4. Biomolecules. 2023; 13(11):1636. https://doi.org/10.3390/biom13111636
Chicago/Turabian StyleDilimulati, Kamila, Zhang Yulin, Fabiana Lica Imai, and Naoto Yonezawa. 2023. "N-Glycosylation Site in the Middle Region Is Involved in the Sperm-Binding Activity of Bovine Zona Pellucida Glycoproteins ZP3 and ZP4" Biomolecules 13, no. 11: 1636. https://doi.org/10.3390/biom13111636
APA StyleDilimulati, K., Yulin, Z., Imai, F. L., & Yonezawa, N. (2023). N-Glycosylation Site in the Middle Region Is Involved in the Sperm-Binding Activity of Bovine Zona Pellucida Glycoproteins ZP3 and ZP4. Biomolecules, 13(11), 1636. https://doi.org/10.3390/biom13111636