Abstract
The advanced version of the author’s inversion concept of the origin of terrestrial life and its application for life in the Universe has been substantiated. A key step in the transition to life consists in the thermodynamic inversion of non-living prebiotic microsystems when the contributions of free energy (F) and information (I) become prevalent over the contribution of entropy (S). It is based the thermodynamic corridor that is mandatory for all chemical scenarios for the origin of life: F + I < S (prebiotic microsystem) → F + I ≈ S (intermediate stage, inversion moment) → F + I > S (primary living unit). A prebiotic organic microsystem can reach the intermediate state between non-life and life only under high-frequency and multilevel oscillations of physic-chemical parameters in hydrothermal environments. The oscillations are considered the fourth required condition for the origin of life, in addition to the three well-known ones: the availability of organic matter, an aqueous medium, and a source of energy. The emergence of initial life sparks in nonequilibrium prebiotic microsystems (being at the intermediate state) proceeds through the continuous response (counteraction) of prebiotic microsystems to incessant physic-chemical oscillations (stress). The next step of laboratory simulations on the origin of life directed to the exploration of the microsystems’ response to high-frequency oscillations (>10−10 s–<30 min) is proposed. Finally, some fragments of the general scenario of the origin of life in the Universe based on the whole four required conditions have been outlined.
1. Introduction
The discovery of extrasolar planets in 1995 opened the way for a search of habitable worlds beyond Earth. Thousands of discovered exoplanets gave us great opportunities for this direction of research. Being the only known example of life in the Universe, the Earth is considered the natural “model” of a habitable world that could guide the search others. A lot of interdisciplinary and special data that have been obtained in the course of our planetary exploration represents a good basis for this study.
One of the most intriguing questions that refers to habitable worlds is their rise, i.e., the emergence of the simplest living units on planets or satellites. The current knowledge of physics, chemistry, geology and biology, which are relevant to the origin of life on Earth, can be expanded and interpreted in the context of the general scenario of the origin of life in the Universe. A part of this knowledge is related to distinguishing the main conditions that are required for the transformation of prebiotic chemical systems into primary life forms. How can these conditions be substantiated and how universal they are? The aim of this article is to consider recent achievements in this area and advance the author’s inversion concept of the origin of life, including the substantiation of its experimental verification.
2. Three Accepted Required Conditions for the Origin of Life
According to almost all present-day concepts, the origin of life on early Earth demanded three required conditions: the availability of organic matter, a liquid medium, and a source of energy.
Organic matter is the substantial basis for all known forms of life. Carbon takes a unique position in the Periodic Table. Its ties with hydrogen and some other chemical elements are extraordinarily flexible. Due to this property, organic matter is capable of the boundless complication that is a necessary factor of biological evolution. On the early Earth, organic compounds could be synthesized of inorganic components in the hydrosphere and wet atmosphere by means of thermal energy and lightning, which was experimentally proved by Miller [1]. High-temperature synthesis of organic compounds might take place in the course of volcanic eruptions [2] and hydrothermal process [3]. Besides, organics could be delivered to the primordial Earth from space and then be involved in the planetary endogenous and exogenous processes. Organic compounds are also widely spread in space, including gas-dust clouds (more than 160 simple compounds), planets (Jupiter, Saturn, Mars), satellites (Titan, Enceladus), and comets. Many organic components were detected in meteorites, including some amino acids that are important for terrestrial biochemistry. In fact, organic matter is everywhere in the Universe, representing wide potential opportunities for the emergence of habitable planets.
Water is a universal medium for the existence and evolution of the simplest prokaryotic cells. It served as a host environment for original forms of life. The oldest thermophilic species of Archaea and Bacteria, which take a position at the root of the Phylogenetic Tree, inhabit hot aquifers in upper zones of hydrothermal systems. Water is also one of the main constituents of living beings: the substantial basis of terrestrial life consists of a specific combination of water and organic material. Besides, water is detected everywhere in the Solar system (ice layers in the polar regions on Mars and Moon, ice crust of the satellites Europa and Enceladus, vapor in the atmosphere of Venus, etc.). Potentially habitable zones around stars are estimated within the temperature interval 0–100 °C, where liquid water may exist.
The third required condition for the origin of life—a source of energy—is necessary to launch the biological processes that follow the laws of thermodynamics. Moreover, the evolution of the Earth’s biosphere proceeds with the continuous accumulation and concentration of free energy, which demands an unceasing energy inflow. At the present time, sunlight, thermal and chemical energy are considered possible sources for the origin of life on early Earth. Sunlight is the usual source for concepts that propose the ocean as the Cradle of Life, while thermal and chemical energy are attributed to hydrothermal scenarios. One of the versions in this way proposes high thermal, redox and concentration gradients at the interface of contrasting seawater and vent fluids in submarine hot vents as a permanent source of electrochemical energy for the origin of life [4]. Lightning in the atmosphere and above erupting volcanoes as the form of non-thermal energy sources is also important for abiotic organic synthesis (including the fixation of nitrogen in amino acids and sulfur). This thesis is supported by Miller’s experiments [1] and the exploration of volcanic eruptions by Markhinin [2]. Alternative non-thermal energy sources are gamma-radiation and high-energy particles from the young Sun discussed recently [5].
Basing on the listed three conditions, a lot of experiments on the origin of life have been carried out. Several types of prebiotic models with multiform variations were involved in these experiments: coacervates [6], RNA World [7], Protein World [8], Lipid World [9], and Aromatic hydrocarbons World [10]; besides, some researchers have developed specific ‘biomineral’ versions [11]. The appearance of some signs of the living state in the models has been actually proven during the experiments: catalytic activity, the ability for division, the selective extraction of a substance, and even self-replication. However, the revealed signs were not self-maintained. The formed prebiotic models were not converted into a kind of the simplest living units capable of the initial biological evolution. Nevertheless, the conducted laboratory simulations designate the potential capability of the used organic components to serve as a substantial basis for life.
Summarizing, it can be concluded that the listed three required conditions are insufficient to convert any kind of non-living prebiotic microsystems into a primary form of life. To understand the way of such transition clearer, it is necessary to define a principal distinction between living systems and non-living ones (including prebiotic models). The relevant comparison between them has been made into the framework of the inversion concept of the origin of life [12,13,14].
3. Thermodynamic Corridor for Chemical Scenarios of the Origin of Life
A core of the inversion concept, along with the consequent fourth required condition for the emergence of life, is given below. Its starting point is the comparison of living and the main types of non-living systems based on the universal thermodynamic criteria. The following three notions were used in this study:
- Free energy: a measure of high-energy value (a part of inner energy that can be converted into any kind of work).
- Information: its entering reduces uncertainty in a system’s state.
- Entropy: a measure of both the low-energy value and a measure of disorganization
A key thermodynamic balance in natural systems that characterizes both non-living and living ones is the “contribution of free energy (F) and information (I)/contribution of entropy (S)”, shortly, F + I/S. The main types of active non-biological systems (stars, geologically active planets, magmatic and hydrothermal systems) dissipate free energy outside, while biological systems, on the contrary, extract free energy (as well as information) from the environment and concentrate it. So, the balance F + I/S is negative in non-living systems and positive in living ones (in organisms and the whole biosphere). This balance cannot be quantitatively estimated because free energy and information are measured in different units. However, the prevalence of the contributions (either free energy with information, or entropy) can be evaluated basing on the thermodynamic trend of natural systems’ evolution. Thus, the obvious mega-evolution of the biosphere consists of the continuous rise of free energy and (bio)information surplus. This trend indicates their actual prevalence over the contribution of entropy, whose values become negative. Another interpretation of the biosphere thermodynamic mega-evolution would mean the disturbance of the 2nd law of thermodynamics. However, the existence of biological systems is in congruence with the 2nd law. The entropy in them increases too, but the contributions of free energy and (bio)information rise faster due to the specific method of biological organization.
Following the carried out comparison, the rise of primary communities of microorganisms on any planet means the appearance of natural systems characterizing with the negentropy (negative entropy) method of organization, i.e., the biological one. From the thermodynamic point of view, their existence and evolution are maintained by the excessive over-entropy free energy and (bio)information that cannot be suppressed by entropy [13,14,15]. Correspondingly, the transition of prebiotic organic assembles (a kind of non-biological (micro)system) into the simplest forms of life may occur only through thermodynamic inversion, i.e., the radical turn of the balance “contribution of free energy and information/contribution of entropy”. The inversion can be expressed as follows:
F + I < S (prebiotic microsystem) → F + I ≈ S (its intermediate state, inversion moment) → F + I > S (primary living unit).
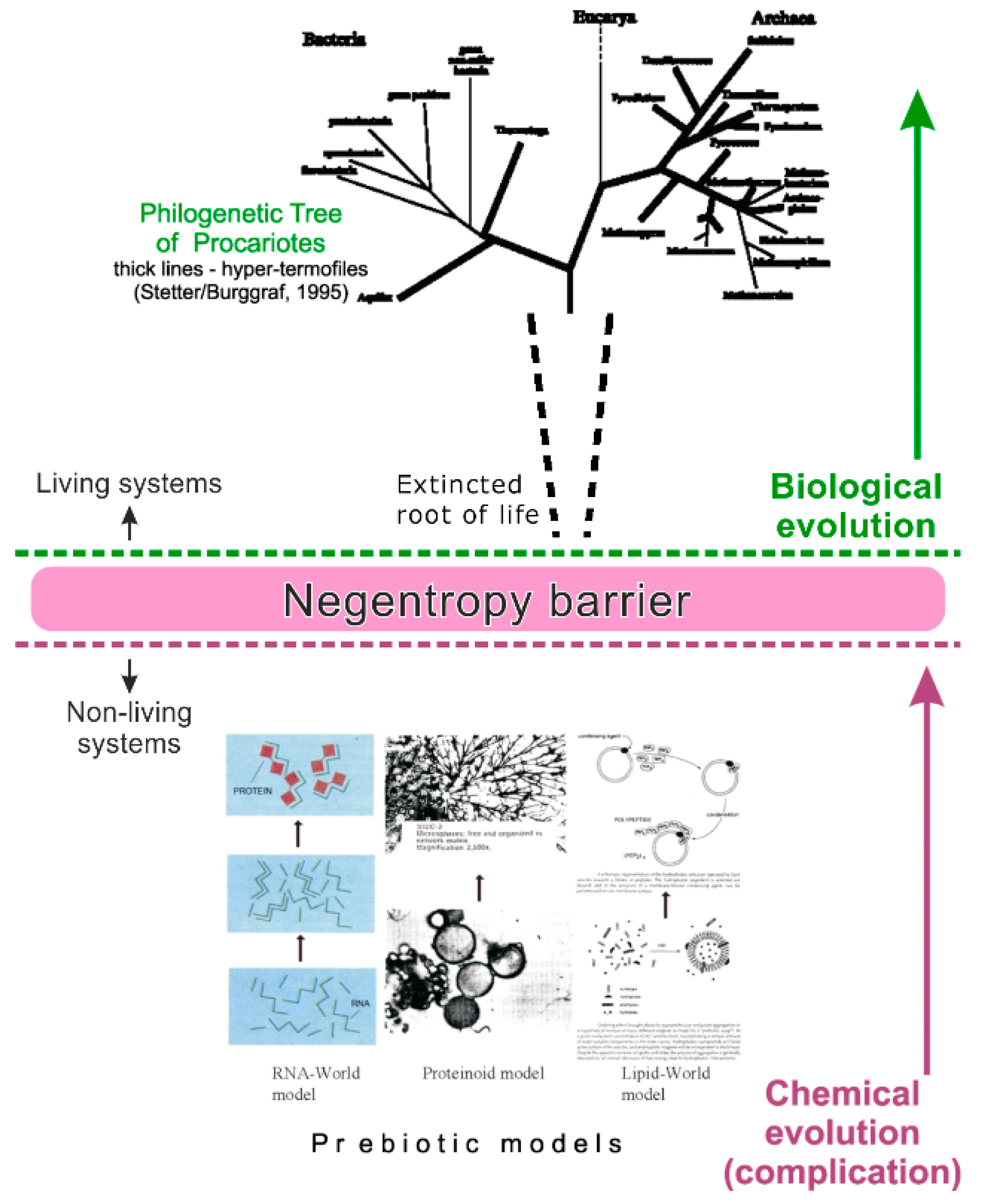
In fact, this transition represents the thermodynamic corridor for the chemical scenarios of the origin of life. The corridor is fair for life origin everywhere, because the used notions—entropy, free energy and information—are fundamental attributes of the Universe. Due to this reason, any chemical scenario of the origin of life, including the type of the prebiotic model and kinetic description of the chemical reactions, must be corresponded with limitations of the thermodynamic corridor. On the way to life, a prebiotic microsystem must overcome the negentropy barrier that exists between the living and non-living natural systems (as well as between the chemical and biological evolution) (Figure 1). The existence of the barrier follows the 2nd law of thermodynamics, which postulates an increase of entropy in non-living natural systems. That means that entropy rises in them due to the general tendency of decrease of various energy gradients. Overcoming the barrier is possible only by means of strong periodic impulses of free energy (and information), which temporarily prevails over the rise of entropy. Such impulses can be generated by high-amplitude oscillations of physic-chemical parameters (both regular and irregular) in the maternal medium. The negentropy barrier cannot be overcome through the gradual complication of organic microsystems in an environment with stable conditions; this case would disturb the 2nd law of thermodynamics. In particular, a huge rise of energy flux into a prebiotic system under rather stable conditions would lead to a huge dissipation of free energy outside, but not to its conversion into a living unit [14].
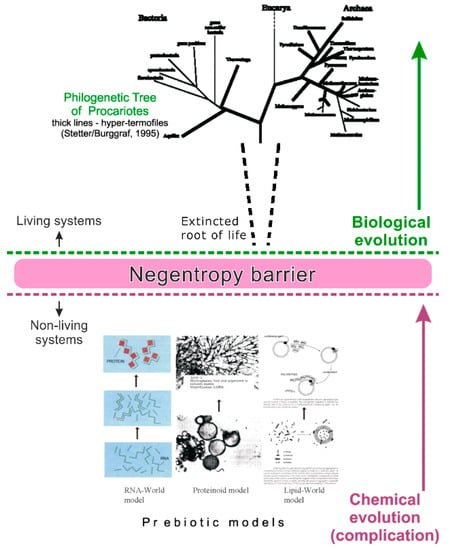
Figure 1.
The thermodynamic (‘negentropy’) barrier between chemical and biological evolution.
4. Additional Fourth Required Condition: Its Role in the Origin of Life
According to the inversion concept, the conversion of organic microsystems into initial living units demands specific far-from-equilibrium conditions, including both the temporal (oscillations) and spatial (gradients) heterogeneity of the origin-of-life medium, i.e., the changeability of the physic-chemical parameters [13,16]. Some fundamental notions of nonequilibrium thermodynamics formulated by I. Prigogine and his followers [17,18] were used as a basis for the elaboration of the inversion approach. The oscillations of temperature, pressure, concentrations, pH, Eh, and other parameters in the medium should proceed in the regime that is optimal for thermodynamic inversion in prebiotic microsystems. Such a regime should be multilevel and include regular micro-oscillation that are interrupted from time to time by irregular macro-fluctuations. The last ones maintain far-from-equilibrium conditions in the Cradle of Life, allowing for prebiotic microsystems to be continuously on the way of transition from the initial stable state into a new stable state and back. In other words, the microsystems oscillate around the unstable point of bifurcation between two attractors—the initial and new states. Regular external micro-oscillations balance the internal oscillations of the microsystem around the bifurcation point, keeping its intermediate position between the attractors. In this way, the micro-oscillations relatively stabilize the bifurcate instability in the microsystems. On Earth, such a combination of high-frequency micro-oscillations and irregular macrooscillations of pressure, temperature, and chemical parameters have been revealed in hydrothermal systems, in particular in the Kamchatka peninsula and Slovenia [19,20,21]. Just fluctuating hydrothermal environments seem to be the most appropriate for the origin of life.
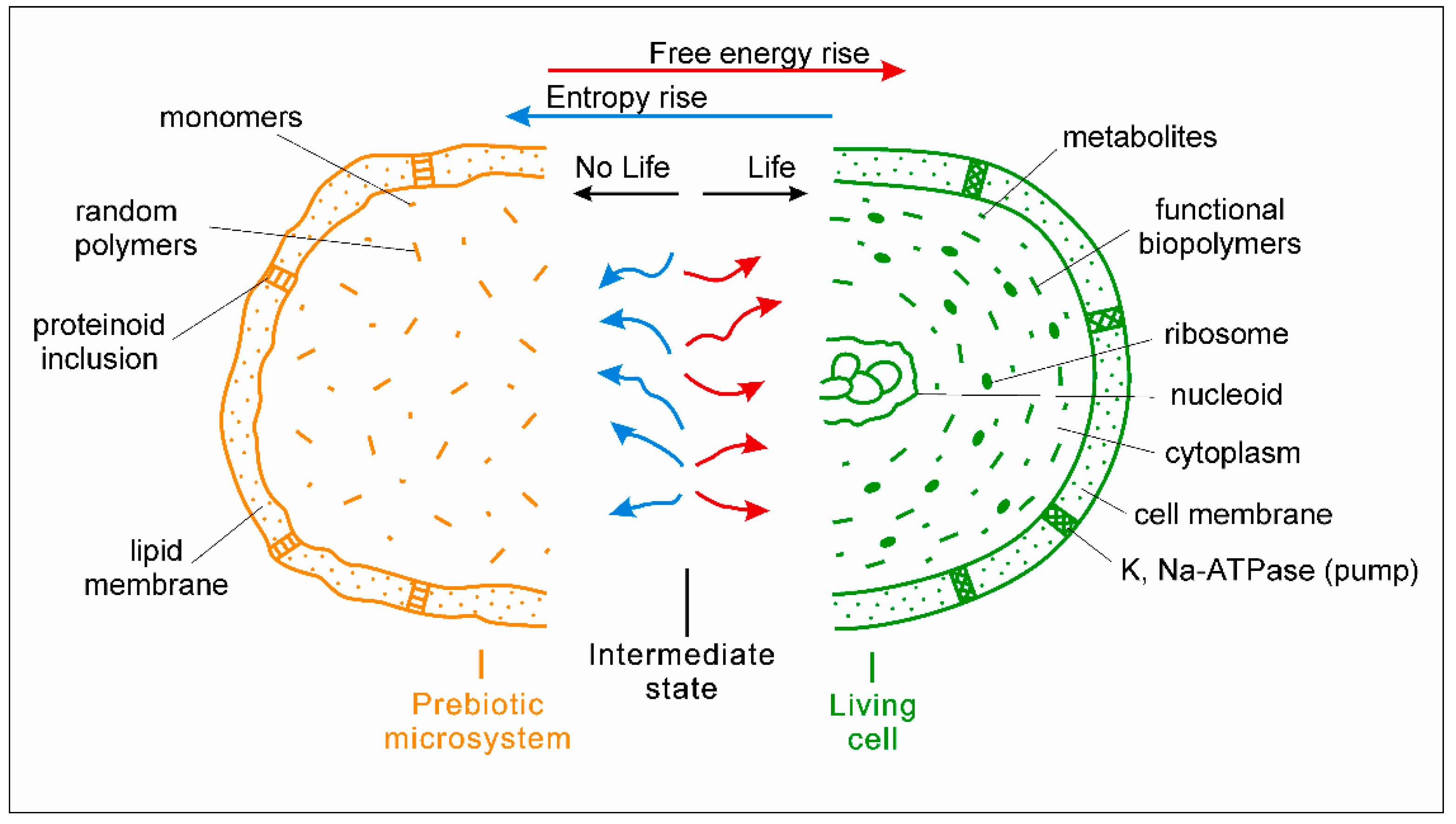
Multilevel physic-chemical oscillations in the medium are considered by the author as the 4th required condition for the origin of life [14]. Under fluctuating conditions in hydrothermal environments, chemical reactions in prebiotic microsystems proceed in an oscillating regime; correspondingly, the balance F + I/S in them oscillates too. A key idea is that the relevant high-amplitude external oscillations might sharply shift the balance up to positive values (F + I > S). In this case, an excessive over-entropy free energy (and information) that is not suppressed by entropy already temporarily appears. This is a crucial moment of the origin of life: a certain surplus of the excessive free energy launches the processes against entropy in the microsystems, including the rise of energy gradients. From this moment, the balance is turned over from positive to negative and back due to the optimal physic-chemical oscillations both in the medium and inside the microsystem. From a thermodynamic point of view, this case corresponds with the intermediate stage between “non-life and life”, characterized by the relative equality of the contributions of free energy with information and entropy in the microsystem (F + I ≈ S). The spontaneous chemical reactions (decreasing energy gradients and producing entropy) and the non-spontaneous ones (restoring energy gradients and decreasing entropy) proceed simultaneously in an oscillating manner and compensate each other (Figure 2). Further, the prebiotic microsystem can either be converted into the simplest living unit through the efficient reorganization of the chemical reactions network or lose the intermediate state due to the irreversible increase of entropy.
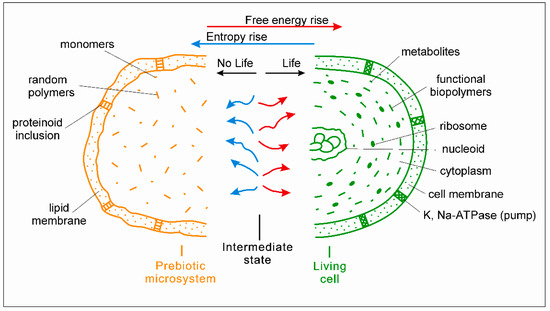
Figure 2.
The intermediate state between ‘non-life’ and life. Left, yellow—non-living prebiotic microsystem (half) composed of random polymers, monomers, and simple molecules. Center: thermodynamically intermediate state between non-life and life, relative equality of the contributions of chemical reactions producing free energy (red) and entropy (blue). Right, green—primary living microorganism (half). Its advanced version is shown—a modern prokaryotic cell with the main cellular structures: nucleoid (ring DNA), ribosomes, and cell membrane.
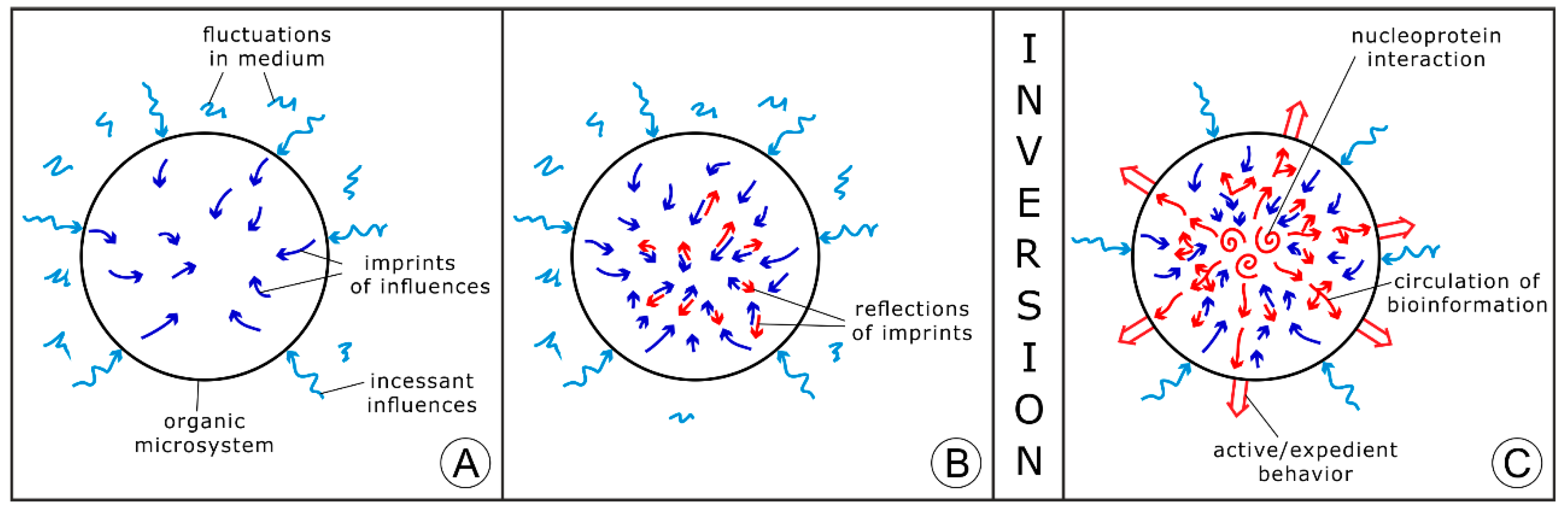
Specificity of a chemical nonequilibrium system, being under external influences, consists in its immediate response to significant changes in the surroundings. The rise of extremely fast chemical reactions induced in response to very short pulses of energy was explored, in particular, by Eigen [22,23]; he was awarded for this work by a Nobel Prize in Chemistry, along with Wreyford Norrish and Porter, in 1967. In accordance with this research, a single or periodic impact of temperature, pressure, and an electric field or another physical factor initiates the equilibrium shift in a chemical system following its transition into a new equilibrium state through the point of bifurcation. This research can be applied to the exploration of prebiotic organic microsystems under oscillating conditions in the framework of the inversion approach. The effect of a single physical impact on a nonequilibrium prebiotic system can be subdivided into two periods. At first, the impact strongly influences (stresses) the network of chemical reactions (Figure 3A), then the system partially compensates the effect of the executed impact, in accordance with the Le Chatelier principle (Figure 3B). Under highly fluctuating conditions, new and new chemical reactions are generated in a prebiotic microsystem as the counteraction to continuous stress. Real chemical equilibrium is never achieved in those nonequilibrium systems until incessant oscillations in the surroundings are maintained. Under a favorable regime of oscillations and a suitable composition of the microsystem, the counteraction may expand and become intensified in comparison to the effect of external stress (Figure 3C). Such a microsystem converses into a primary living unit, in accordance with the author’s concept [13,14]. External stress and an adequate response to it are within the foundation of life processes that are postulated in the theory of stress [24].
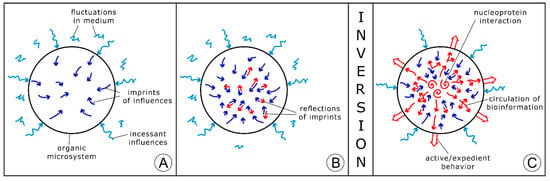
Figure 3.
The scheme of conversion of a nonequilibrium prebiotic microsystem into the primary form of life under oscillating conditions. (A) The rise (self-assembly) of an organic prebiotic microsystem within the oscillating medium; the penetration of continuous external influences into the microsystem and the transformation of the network of chemical reactions (blue arrows); (B) the initiation of the system’s response to continuous external stress, arising of the chemical reactions counteracting the stress and partially compensating it; (C) the conversion of the prebiotic microsystem in the initial form of life when its activity (counteraction) exceeds the pressing from the oscillating environment. The conversion corresponds to the thermodynamic inversion.
The described general scenario of the origin of life is based on the four required conditions: aqueous medium (1st), the availability of organic matter (2nd), the presence of an energy source (3rd), and multilevel physic-chemical oscillations in the medium (4th). Accepting all of them, the author’s inversion concept proposes a highly fluctuating hydrothermal medium as the Cradle of Life on early Earth. This approach does not admit to the emergence of primary life forms in a stable (near equilibrium) environment, like the ocean (beyond submarine hot vents) or ice layers. The chemical composition of prebiotic microsystems suitable for thermodynamic inversion is outlined in chapter 7 of Reference [14]. Shortly, the availability of the following three components seems desirable for thermodynamic inversion (Figure 2, left part):
- A lipid membrane that can prevent the microsystem from substantially dissipating free energy;
- proteinoid inclusions in the membrane that could be transformed into pumps, allowing the microsystem to actively regulate exchange with the environment;
- the combination of polynucleotide and polyamino acid chains, which can be involved into the initial circulation of bioinformation.
However, the suitability of other prebiotic chemical models for the inversion event can be examined too. They should correspond with the four required conditions for the origin of life (at least for life as we know it). In particular, all chemical scenarios (including laboratory experiments and kinetic models) must follow the thermodynamic corridor for the origin of life.
The main condition for the self-assembly of microsystems consists of a rather high concentration of organic molecules in a solution, in case other physic-chemical parameters are appropriate. A suitable mechanism of organics concentration in the local parts of hydrothermal systems is proposed in the author’s book ([14], Chapter 7). The mechanism is related to the pulsating migration of fluid to the surface when organic molecules periodically accumulate due to the closing of the cracks. In particular, this means a huge number of simultaneously assembling and interacting microsystems that form a heterogeneous cluster.
5. The Next Step of Laboratory Simulations on the Origin of Life Based on the Four Conditions
By now a general goal of the experiments in prebiotic chemistry consists in the exploration of various pathways of chemical evolution based on different starting organic components or models. The addition of the multilevel oscillations into a set of required conditions for the origin of life demands a change in the experimental direction. A key idea in this way is that self-maintained life processes were launched through continuous responses of prebiotic microsystems to incessant physic-chemical oscillations in the outside world. This corresponds with the Le Chatelier principle and the data on chemical reactions induced in response to very short pulses of energy: no substantial multi-oscillations in an aqueous medium (for instance, in the calm ocean), no incessant recombination within containing organic microsystems and no perspective for their transformation into living units.
By now almost all experiments on prebiotic chemistry have been carried out under stable conditions. However, some rare simulations were conducted under simple oscillating environments [25,26,27,28]. Although a response of organic molecules/microsystems to external influences were not explored during these experiments, they have helped demonstrate the importance of oscillating conditions for the advancement of prebiotic evolution. Their general results consist of the increased yield of polycondensation products, in comparison to the simulations under stable (non-oscillating) conditions. For instance, the synthesis and oligomerization of glycine take place in rather stable hydrothermal conditions, but the synthesized molecules display the tendency to decompose [29]. However, under oscillations of temperature, these molecules can be reorganized into increasingly complex structures. Some laboratory experiments have demonstrated advancements of prebiotic chemical evolution under fluctuations of temperature. For instance, the amplification of DNA under temperature oscillations between 64 °C and 92 °C accelerated the division of giant vesicles [27]. Oscillations of wet-dry conditions (the model of a periodically evaporating and filling hydrothermal pond) promoted the synthesis of RNA-like molecules from mononucleotides [28]. The regime of thermocycling around the water boiling point strengthens the condensation of amino acids into peptides [26]. Moreover, the computational modeling of thermal cycling also demonstrates the acceleration of the pace of chemical prebiotic evolution [30].
Substantiation of the required thermodynamic corridor for all chemical scenarios for the origin of life offers the next step of laboratory simulations in this field. The proposed experiments would give us a chance to check the theoretically elaborated inversion concept. A goal of such modified laboratory experiments would consist of the exploration of the intermediate state between non-life and life in various prebiotic models (microsystems) that are of highly fluctuating environments. The appearance of the response to external changes and its development within the organic matrix can be verified. The intermediate area between organic chemistry and primary biochemistry (i.e., “transitional” chemistry) is very vast. For instance, it is possible to detect the strengthening of active transport which is responsible for molecules/atoms transferring against the energy (concentration) gradients. The rise of homochirality degree during the proposed experiments could also indicate the initial sparks of biochemical processes. One more criterion follows of the experiments by Fox and co-authors [8,31]: although abiotically synthesized proteinoid chains are similar to biological protein ones, they cannot be arranged into tertiary structures; the appearance of the last ones could also be traced in the course of the modified experiments. It is expected that the continuous self-assembly of prebiotic microsystems under oscillating conditions (close to the critical point: assembly and re-assembly) facilitates the incessant recombination of (macro)molecules. It would be exceptionally interesting to explore the oscillating interaction of prebiotic microsystems within the clusters.
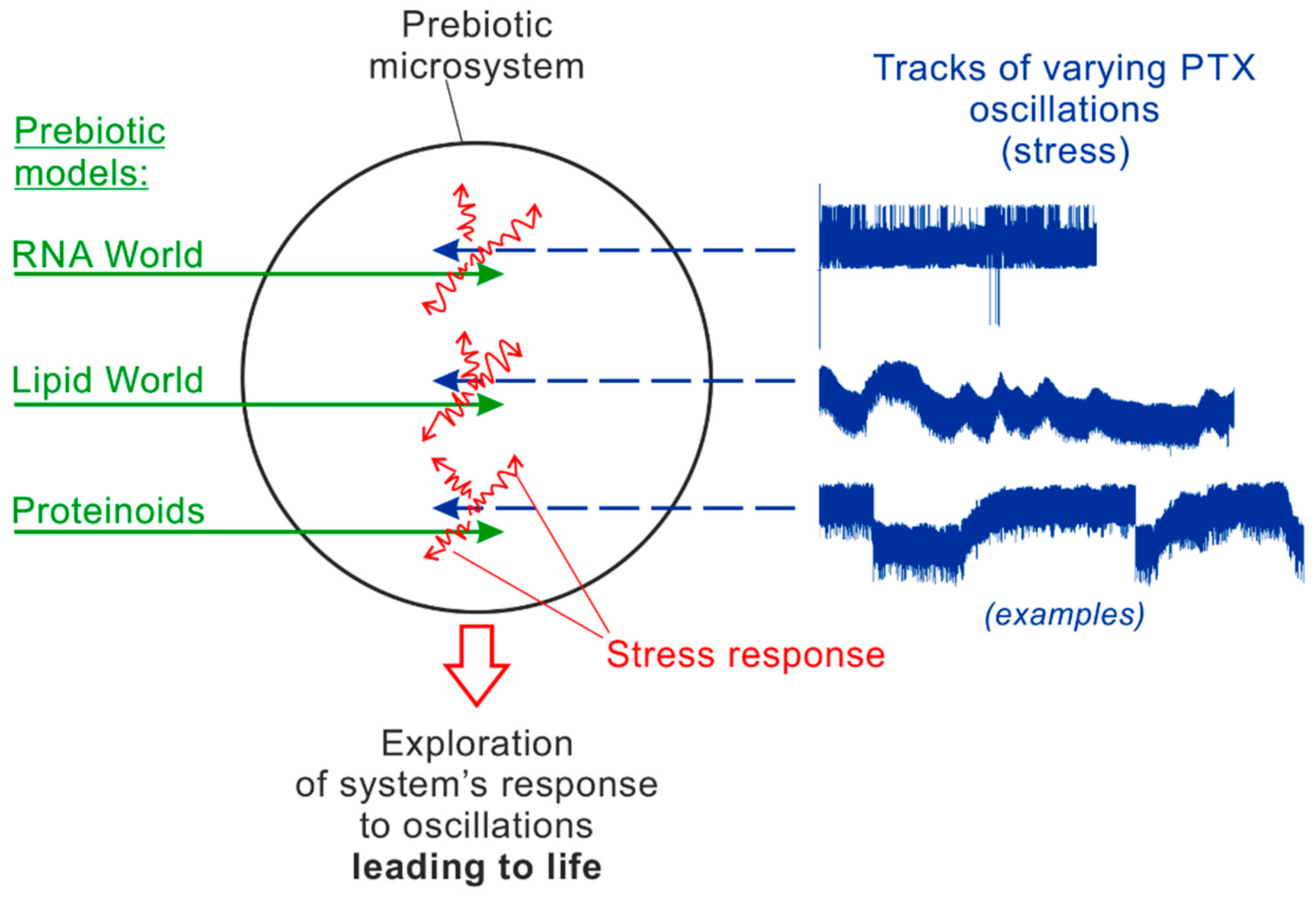
A general scheme of the suggesting laboratory simulations is given in Figure 4. They involve two constituents: various kinds of prebiotic models (left, green) and varying tracks of physic-chemical oscillations in the experimental chamber (right, blue). The shown examples of the tracks represent real fragments of pressure oscillations measured in the Mutnovsky hydrothermal system in Kamchatka [19,20]. It is supposed that the long “pumping” of the models by external oscillations generate a continuous response in them, including induced chemical reactions (red) directed to compensation for the external stress. The “pumping” tracks can be different in duration, the regime of oscillations (amplitudes, periods/frequencies, etc.) and the oscillating parameters (pressure, temperature, electric potential, pH, Eh, etc.). The first step in this experimental method would be finding the most promising combination of prebiotic models (1) and characteristic tracks of oscillations (2) which provides obvious signs of an intermediate state between non-life and life.
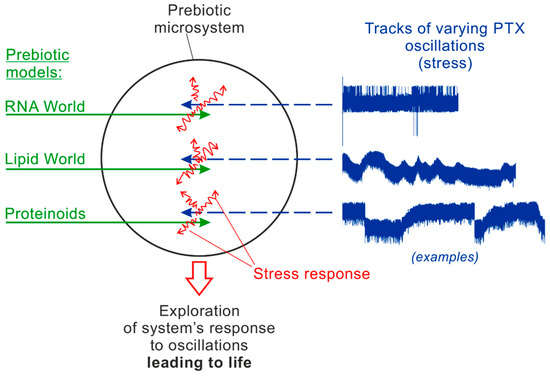
Figure 4.
The scheme of the new direction of laboratory simulations on the origin of life based on the four required conditions.
According to the preliminary evaluation, the regular constituent of the oscillations should be within the interval between >10−10 s and <30 min. On the one hand, the exploration of extremely fast chemical reactions demonstrates the return of the chemical system to an equilibrium state after the temperature impacts for a period of around 10−10 s. However, this period should be much longer in natural fluctuating hydrothermal environments because many physic-chemical parameters oscillate and influence each other. On the other hand, the lifetime (from initial division to final one) of the oldest thermophilic microorganisms can be as short as 30 min. Obviously, periods of the external oscillations which stimulated the (bio)chemical processes within initial living units on the early Earth should be much shorter in terms of their entire lifetime. Summarizing, just the high-frequency diapason of physic-chemical oscillations is optimal for laboratory simulations on prebiotic chemistry aimed at achieving the intermediate stage between non-life and life. Such a modeling environment has still not been created during the experiments on the origin of life.
6. Conclusions: Fragments of the General Scenario of Habitable Planets Arising in the Universe
This paper represents an advanced version of the inversion concept. Its new points can be narrowed down to the following items.
1. The interrelation between thermodynamics and chemistry with respect to life origin has been clearly substantiated. Thermodynamics determines the general corridor that could lead to the conversion of non-living organic microsystems into primary life forms. Various chemical scenarios of conversion (the both theoretical and experimental) must follow within the corridor’s limitations in order to reach the intermediate state between non-life and life. First of all, the thermodynamic limitations concern the necessity of optimal high-frequency oscillations of physic-chemical parameters in the maternal medium.
2. An experimental way to verify the theoretical notions of the inversion concept is offered. The experiments are considered as the next step of laboratory simulations on the origin of life that should be conducted under high-frequency oscillations. A general diapason of their periods is defined between >10−10 s and <30 min.
3. The universality of thermodynamic and chemical regularities for life origin in the Universe is considered. The positive balance “contribution of free energy and information > contribution of entropy” (thermodynamic criterion) is the most universal statement and should be peculiar to life everywhere because entropy, free energy and information are attributes of the whole Universe. We cannot imagine the mega-evolution of a biosphere on any planet without the continuous accumulation and concentration of free energy and information that is possible only through their persistent prevalence over entropy. The chemical properties of the terrestrial life may not be precisely peculiar to life on other planets. It is supposed that some specific (geo)biochemical pathways of alien life can be different because the exploration of the solar planets and satellites demonstrates their significant difference in terms of geochemical composition.
The author offers that the four considered conditions should be included into the general scenario of the origin of life in the Universe because all of them seem to be required (at least for life as we know it). The 4th condition implies the availability of multilevel physic-chemical oscillations in an aqueous high-temperature medium that demands intensive volcanic-tectonic processes for life to appear. Therefore, a habitable world can arise only on geologically active planets or satellites (with a surface temperature below 100 °C). Such a planet should be inevitably characterized with the vertical migration of fluid within the crust, forming hydrothermal systems similar to those in the Earth’s crust. The genesis of organic compounds involved in hydrothermal circulation can be different (hydrothermal, volcanic, atmospheric, or extraterrestrial). However, the major factor of the origin of life is the method of their arrangement into primary living units under high-frequency oscillations. The substantiated set of the four required conditions was present on early Mars about 3–4 billion years ago. Besides, it might take place at some satellites in the Solar system, where geysers (Enceladus) and signs of submarine vents beneath ice crusts (Europa) have been detected.
Thermodynamic inversion itself designates only the formation of a kind of natural systems in which free energy and information rise faster than entropy (i.e., biological ones). It does not directly concern the chemical composition of living systems. Therefore, at the present level of our knowledge, we cannot exclude the possible existence of another version of natural systems that also possesses the ability to concentrate over-entropy free energy and information, on the one hand, and a discrepancy from terrestrial life, on the other hand (possible life as we do not know). Such a version should be related to organic matter and water too, but can be organized in a different way. If such life exists, it should probably be simpler than terrestrial life. One of the possible locations for the search of such a theoretically admissible version of life is in the Saturn’s satellite Titan.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Miller, S.L. A production of amino acids under possible primitive earth conditions. Science 1953, 117, 528–529. [Google Scholar] [CrossRef] [PubMed]
- Markhinin, E.K.; Podkletnov, N.E. The phenomenon of formation of prebiological compounds in volcanic processes. Orig. Life Evol. Biosph. 1977, 8, 225–235. [Google Scholar] [CrossRef]
- Holm, N.G.; Andersson, E. Hydrothermal simulation experiments as a tool for studies of the origin of life on earth and other terrestrial planets: A review. Astrobiology 2005, 5, 444–460. [Google Scholar] [CrossRef] [PubMed]
- Barge, L.M.; Branscomb, E.; Brucato, J.R.; Cardoso, S.S.S.; Cartwright, J.H.E.; Danielache, S.O.; Galante, D.; Kee, T.P.; Miguel, Y.; Mojzsis, S.; et al. Thermodynamics, disequilibrium, evolution: Far-from-equilibrium geological and chemical considerations for origin-of-life research. Orig. Life Evol. Biosph. 2016, 47, 39–56. [Google Scholar] [CrossRef]
- Airapetian, V.S.; Glocer, A.; Gronoff, G.; Hébrard, E.; Danchi, W. Prebiotic chemistry and atmospheric warming of early Earth by an active young Sun. Nature Geosci. 2016, 9, 409. [Google Scholar] [CrossRef]
- Oparin, A.I. Origin of Life on the Earth; Nauka: Moscow, Russia, 1957. [Google Scholar]
- Joice, G.F.; Orgel, L.E. Prospects for understanding the origin of the RNA world. In The RNA World; Gold Spring Harbor laboratory Press: New York, NY, USA, 1993; pp. 1–25. [Google Scholar]
- Fox, S.W.; Bahn, P.R.; Pappelis, A.; Yu, B. Experimental retracement of terrestrial origin of an excitable cell: Was it predictable? In Chemical Evolution: Physics of the Origin and Evolution of Life; Springer Nature: Basingstoke, UK, 1996; pp. 21–32. [Google Scholar]
- Deamer, D.W. Prebiotic amphiphilic compounds. In Origins. Cellular Origin, Life in Extreme Habitats and Astrobiology; Seckbach, J., Ed.; Springer: Dordrecht, The Netherlands, 2004; pp. 75–89. [Google Scholar]
- Ehrenfreund, P.; Rasmussen, S.; Cleaves, J.; Chen, L. experimentally tracing the key steps in the origin of life: The aromatic world. Astrobiology 2006, 6, 490–520. [Google Scholar] [CrossRef] [PubMed]
- Ferris, J.P. Montmorillonite catalysis of 30–50 mer oligonucleotides: Laboratory demonstration of potential steps in the origin of the RNA world. Orig. Life Evol. Biosph. 2002, 32, 311–332. [Google Scholar] [CrossRef]
- Kompanichenko, V. Three stages of the origin of life process: Bifurcation, stabilization and inversion. Int. J. Astrobiol. 2008, 7, 27–46. [Google Scholar] [CrossRef]
- Kompanichenko, V.N. Inversion concept of the origin of life. Orig. Life Evol. Biosph. 2012, 42, 153–178. [Google Scholar] [CrossRef]
- Kompanichenko, V.N. Thermodynamic Inversion: Origin of Living Systems; Springer International Publishing: Cham, Switzerland, 2017. [Google Scholar]
- Kompanichenko, V.N. Thermodynamic inversion and self-reproduction with variations: Integrated view on the life-nonlife border. J. Biomol. Struct. Dyn. 2012, 29, 637–639. [Google Scholar] [CrossRef]
- Kompanichenko, V. Changeable hydrothermal media as potential cradle of life on a planet. Planet. Space Sci. 2009, 57, 468–476. [Google Scholar] [CrossRef]
- Prigogine, I.; Stengers, I. Order out of Chaos; Bantam: New York, NY, USA, 1984. [Google Scholar]
- Ebeling, W.; Engel, A.; Feistel, R. Physik der Evolutionsprozesse (Physics of Evolution); Akademie-Verlag: Berlin, Germany, 1990. (In German) [Google Scholar]
- Kompanichenko, V.N.; Shlyufman, K.V. The amplitude-frequency function of pressure variations: Steam-water mixture in the Verkhne-Mutnovskii hydrothermal system. J. Volcanol. Seismol. 2013, 7, 338–344. [Google Scholar] [CrossRef]
- Kompanichenko, V.N.; Poturay, V.A.; Shlufman, K.V. Hydrothermal systems of kamchatka are models of the prebiotic environment. Orig. Life Evol. Biosph. 2015, 45, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Kralj, P.; Kralj, P. Thermal and mineral waters in north-eastern Slovenia. Environ. Geol. 2000, 39, 488–500. [Google Scholar] [CrossRef]
- List of Publications by Manfred Eigen. Biophys. Chem. 1987, 26, 103–108. [CrossRef]
- Eigen, M. Selforganization of matter and the evolution of biological macromolecules. Naturwissenschaften 1971, 58, 465–523. [Google Scholar] [CrossRef] [PubMed]
- Selye, H. Stress without Distress; JB Lippincott Company: Philadelfia, PA, USA, 1974. [Google Scholar]
- Yokoyama, S.; Koyama, A.; Nemoto, A.; Honda, H.; Imai, E.-I.; Hatori, K.; Matsuno, K. Amplification of diverse catalytic properties of evolving molecules in a simulated hydrothermal environment. Orig. Life Evol. Biosph. 2003, 33, 589–595. [Google Scholar] [CrossRef]
- Varfolomeev, S.D. Kinetic models of the prebiological evolution of macromolecules. Mendeleev Commun. 2007, 17, 7–9. [Google Scholar] [CrossRef]
- Mikhailov, A.S.; Ertl, G.; Sugawara, T.; Kurihara, K.; Suzuki, K. Constructive approach towards protocells. Anal. Control Complex Nonlinear Process. Phys. Chem. Biol. 2012, 359–374. [Google Scholar]
- Ross, D.S.; Deamer, D. Dry/wet cycling and the thermodynamics and kinetics of prebiotic polymer synthesis. Life 2016, 6, 28. [Google Scholar] [CrossRef]
- Cleaves, H.; Aubrey, A.D.; Bada, J.L. An evaluation of the critical parameters for abiotic peptide synthesis in submarine hydrothermal systems. Orig. Life Evol. Biosph. 2008, 39, 109–126. [Google Scholar] [CrossRef] [PubMed]
- Osipovitch, D.C.; Barratt, C.; Schwartz, P.M. Systems chemistry and Parrondo’s paradox: Computational models of thermal cycling. New J. Chem. 2009, 33, 2022–2027. [Google Scholar] [CrossRef]
- Fox, S.; Dose, K. Molecular Evolution and the Origin of Life; Dekker: New York, NY, USA, 1975. [Google Scholar]
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).