
Downregulation of de Novo Fatty Acid Synthesis in Subcutaneous Adipose Tissue of Moderately Obese Women


Abstract
:
1. Introduction
2. Results
2.1. Baseline Characteristics of the Cohort Studied

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Controls (n = 35) | Moderately Obese Patients (n = 55) | p-Value |
|---|---|---|---|
| Mean ± SD | Mean ± SD | ||
| AGE (years) | 49.57 ± 14.17 | 52.94 ± 14.24 | 0.289 |
| WEIGHT (kg) | 57.53 ± 7.17 | 84.49 ± 11.65 | <0.001 |
| WC (cm) | 76.43 ± 11.42 | 109.85 ± 11.15 | <0.001 |
| BMI (kg/m2) | 22.28 ± 1.63 | 33.67 ± 2.70 | <0.001 |
| GLUCOSE (mg/dL) | 86.51 ± 22.96 | 110.61 ± 45.32 | 0.002 |
| INSULIN (mU/L) | 6.84 ± 5.43 | 13.88 ± 9.65 | <0.001 |
| HbA1c (%) | 4.92 ± 0.64 | 5.49 ± 1.18 | 0.015 |
| HOMA2-IR | 0.88 ± 0.75 | 1.90 ± 1.38 | <0.001 |
| SBP (mmHg) | 124.63 ± 18.20 | 130.37 ± 18.11 | 0.157 |
| DBP (mmHg) | 69.8 ± 9.52 | 74.27 ± 11.45 | 0.063 |
| TOTAL CHOLESTEROL (mg/dL) | 181.33 ± 37.57 | 182.92 ± 51.38 | 0.867 |
| HDL-C (mg/dL) | 55.34 ± 14.93 | 43.29 ± 10.86 | <0.001 |
| LDL-C (mg/dL) | 107.33 ± 31.02 | 113.6 ± 43.37 | 0.482 |
| TRIGLYCERIDES (mg/dL) | 93.57 ± 58.13 | 149.15 ± 82.57 | 0.001 |
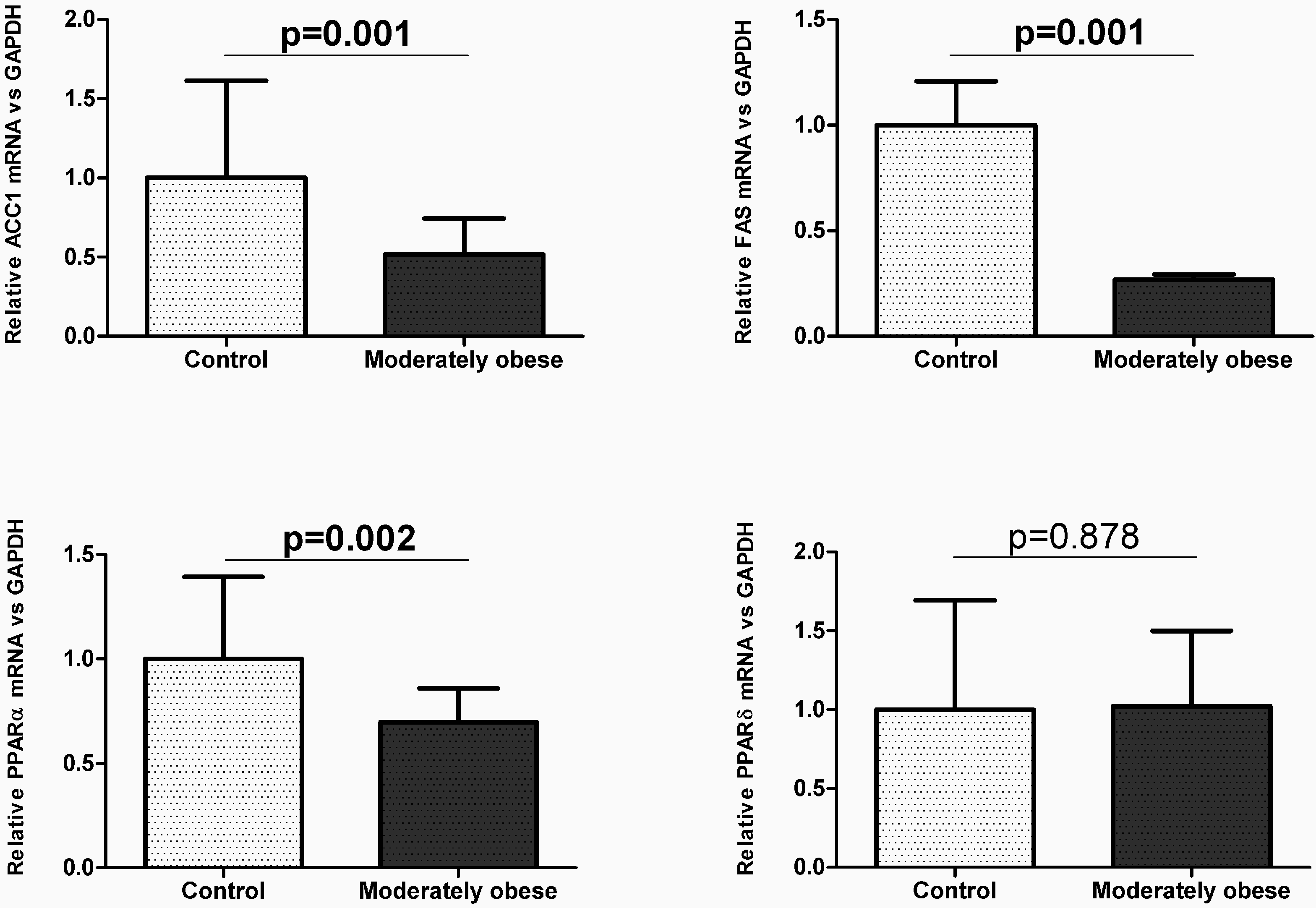
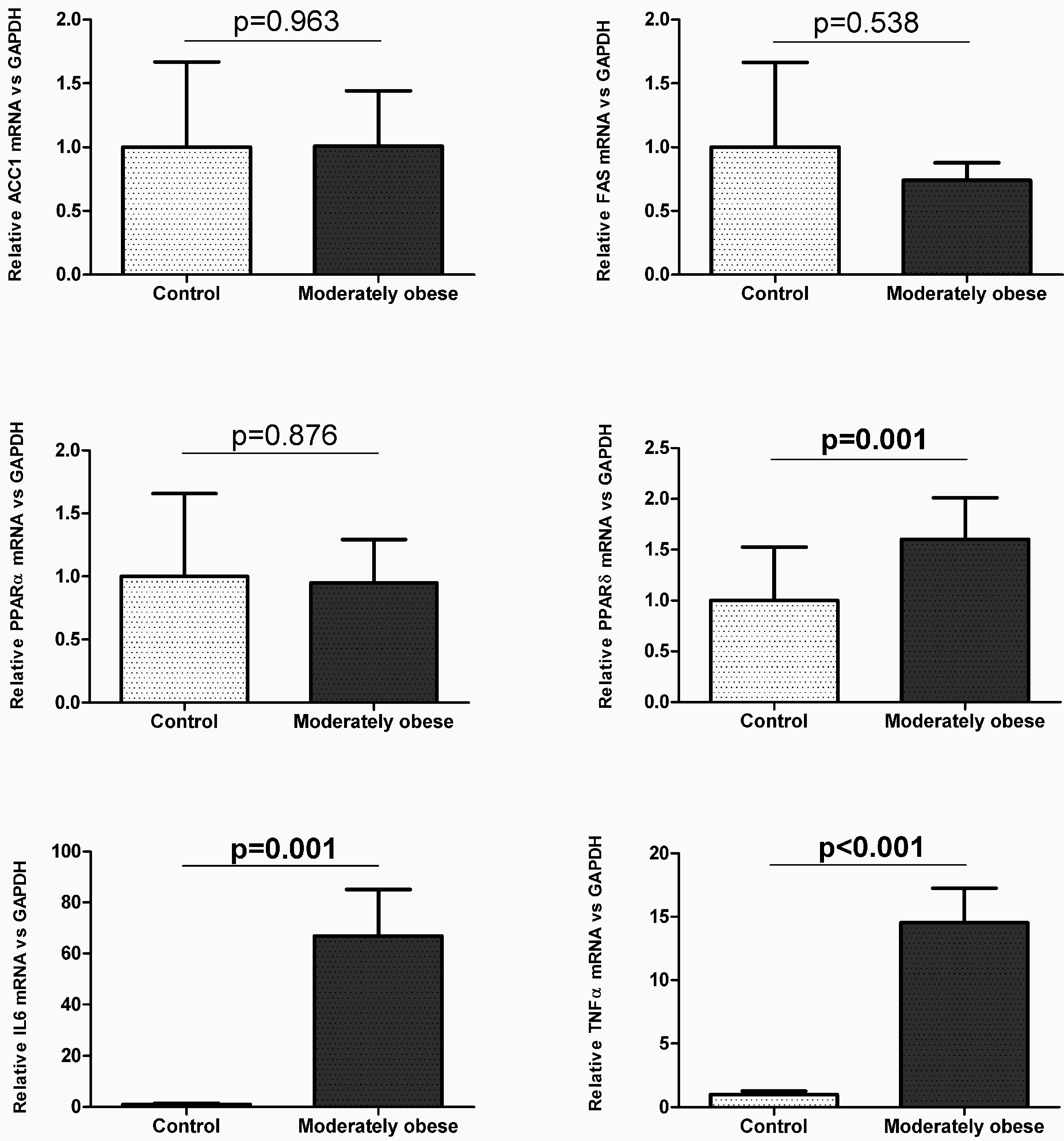
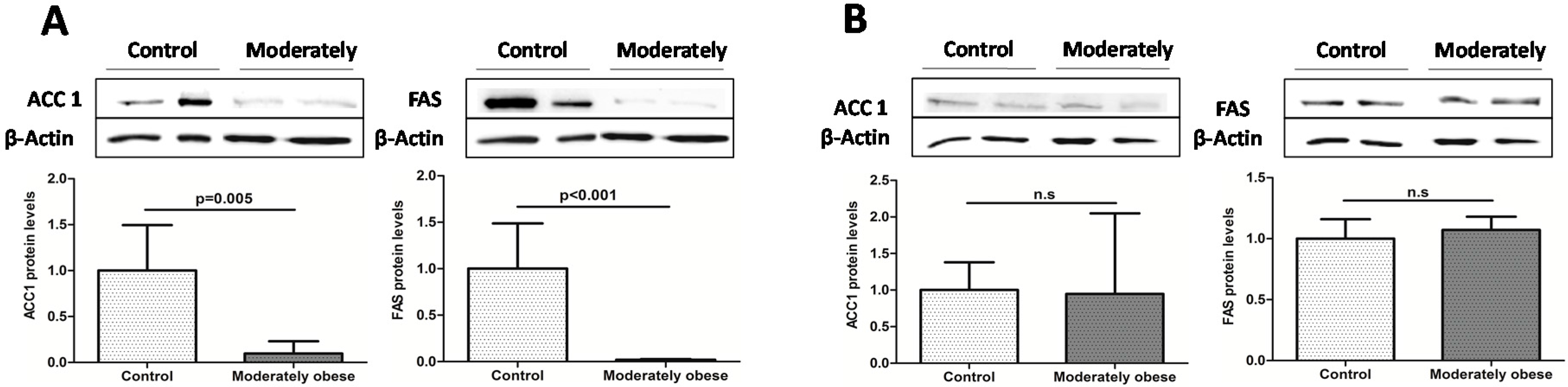
2.2. Evaluation of the Expression of FA Metabolism Genes and Their Protein Levels in SAT and VAT
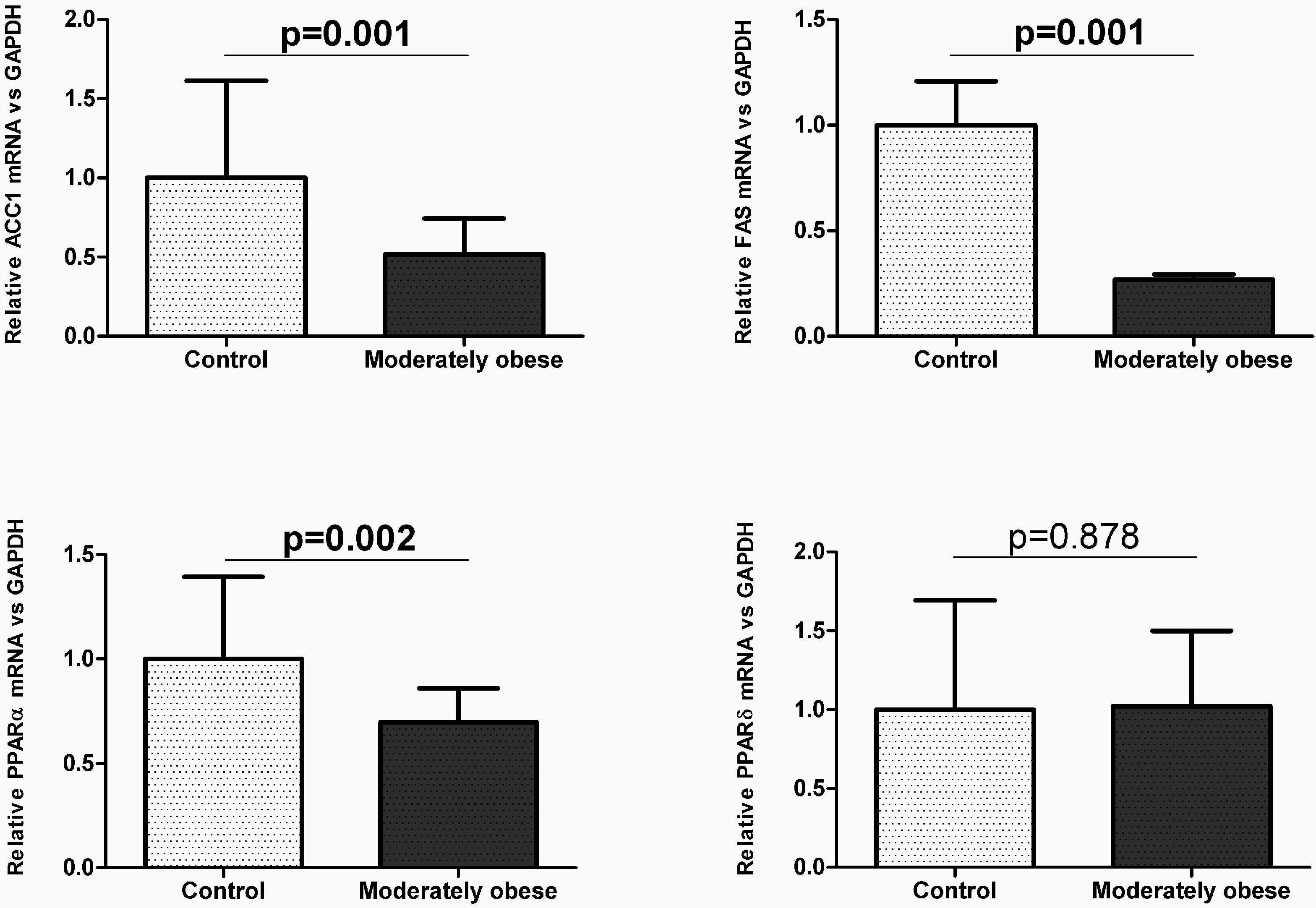

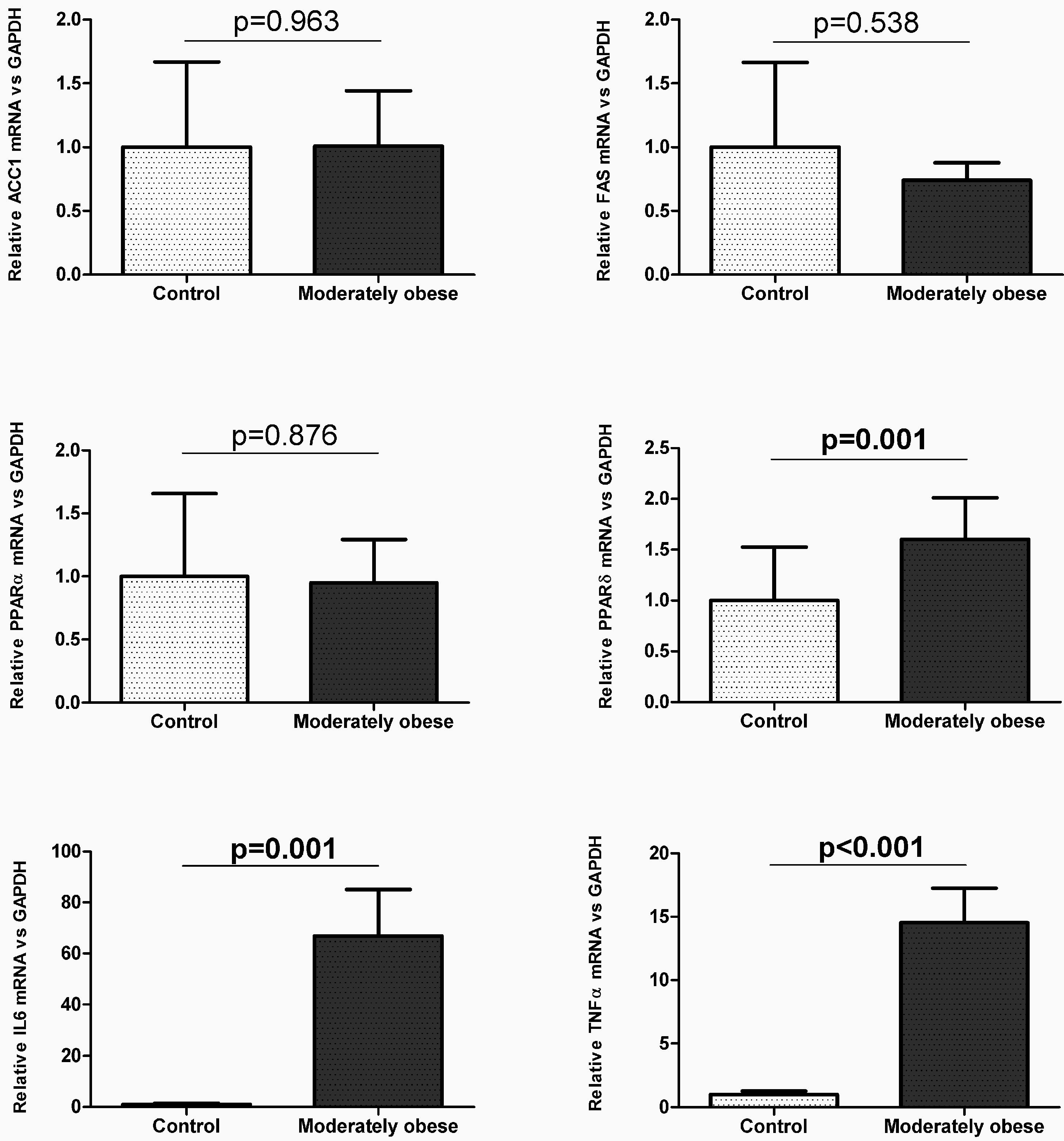
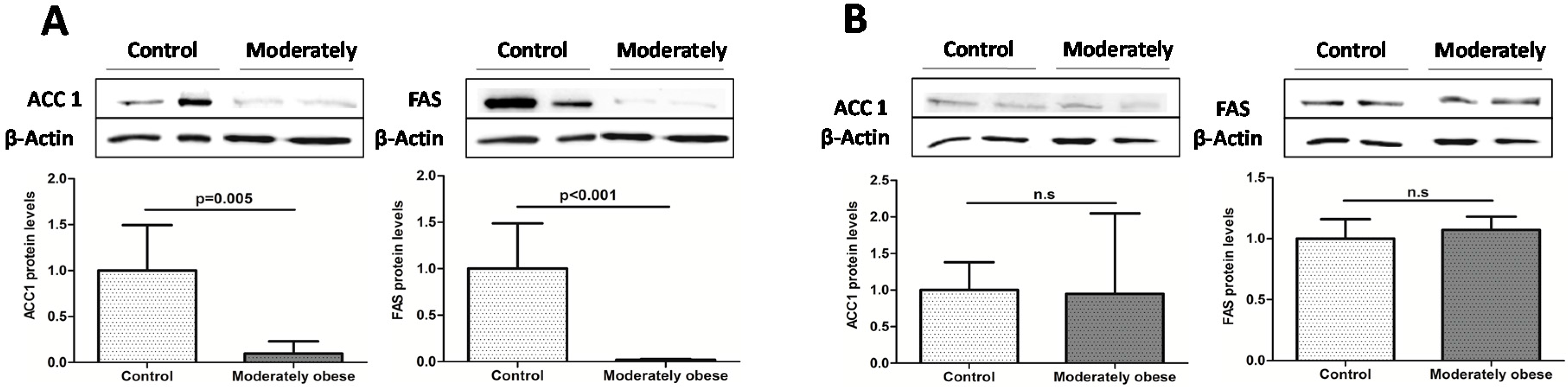
2.3. Correlation of VAT and SAT mRNA Expression with Parameters of Obesity and Glucose Metabolism
| Variables | ACC1 | FAS | PPARα | PPARδ | IL6 | TNFα | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| r | p-Value | r | p-Value | r | p-Value | r | p-Value | r | p-Value | r | p-Value | |
| WEIGHT (kg) | −0.044 | 0.797 | −0.010 | 0.957 | −0.141 | 0.464 | 0.387 | 0.018 | 0.430 | 0.020 | 0.214 | 0.275 |
| WC (cm) | 0.196 | 0.521 | 0.291 | 0.335 | −0.314 | 0.320 | 0.527 | 0.044 | 0.243 | 0.447 | 0.179 | 0.597 |
| BMI (kg/m2) | −0.027 | 0.874 | −0.044 | 0.812 | −0.146 | 0.450 | 0.464 | 0.004 | 0.459 | 0.012 | 0.351 | 0.067 |
| GLUCOSE (mg/dL) | −0.122 | 0.487 | −0.153 | 0.411 | −0.261 | 0.180 | 0.136 | 0.423 | 0.573 | 0.001 | 0.533 | 0.004 |
| INSULIN (mU/L) | −0.045 | 0.808 | −0.251 | 0.198 | 0.375 | 0.065 | 0.041 | 0.816 | 0.374 | 0.046 | 0.526 | 0.007 |
| HbA1c (%) | 0.105 | 0.587 | −0.212 | 0.298 | −0.031 | 0.887 | 0.221 | 0.233 | 0.496 | 0.002 | 0.427 | 0.042 |
| HOMA2-IR | −0.087 | 0.642 | −0.279 | 0.158 | −0.418 | 0.042 | 0.062 | 0.730 | 0.479 | 0.009 | 0.558 | 0.004 |
| Variables | ACC1 | FAS | PPARα | PPARδ | IL6 | TNFα | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| r | p-Value | r | p-Value | r | p-Value | r | p-Value | r | p-Value | r | p-Value | |
| WEIGHT (kg) | −0.483 | <0.001 | −0.429 | <0.001 | −0.369 | 0.003 | −0.002 | 0.990 | 0.488 | <0.001 | 0.247 | 0.030 |
| WC (cm) | −0.366 | 0.072 | −0.388 | 0.031 | −0.475 | 0.026 | 0.162 | 0.400 | 0.451 | 0.018 | 0.493 | 0.005 |
| BMI (kg/m2) | −0.526 | <0.001 | −0.496 | <0.001 | −0.483 | <0.001 | 0.000 | 0.998 | 0.449 | <0.001 | 0.269 | 0.018 |
| GLUCOSE (mg/dL) | −0.192 | 0.108 | −0.240 | 0.022 | −0.205 | 0.116 | 0.074 | 0.527 | 0.244 | 0.040 | 0.048 | 0.684 |
| INSULIN (mU/L) | −0.216 | 0.081 | −0.298 | 0.013 | −0.180 | 0.185 | 0.182 | 0.132 | 0.275 | 0.028 | 0.264 | 0.028 |
| HbA1c (%) | −0.042 | 0.750 | −0.076 | 0.562 | −0.127 | 0.367 | 0.264 | 0.034 | 0.107 | 0.428 | 0.147 | 0.257 |
| HOMA2-IR | −0.216 | 0.082 | −0.297 | 0.013 | −0.183 | 0.177 | 0.188 | 0.120 | 0.341 | 0.006 | 0.238 | 0.049 |
2.4. Relationship between the mRNA Expression of Genes Involved in Lipogenesis, FA Oxidation and Inflammation in VAT and SAT
3. Discussion
4. Material and Methods
4.1. Subjects
4.2. Biochemical Analyses
4.3. RNA Isolation and Gene Expression
4.4. Western Analysis
4.5. Statistical Analyses
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
References
- Lean, M.E. Pathophysiology of obesity. Proc. Nutr. Soc. 2000, 59, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Reaven, G.M. Perspective importance of identifying the overweight patient who will benefit the most by losing weight. Ann. Intern. Med. 2003, 138, 420–423. [Google Scholar] [CrossRef] [PubMed]
- Klöting, N.; Fasshauer, M.; Dietrich, A.; Kovacs, P.; Schön, M.R.; Kern, M.; Stumvoll, M.; Blüher, M. Insulin-sensitive obesity. Am. J. Physiol. Endocrinol. Metab. 2010, 299, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Stefan, N.; Kantartzis, K.; Machann, J.; Schick, F.; Thamer, C.; Ritting, K.; Balletshofer, B.; Machicao, F.; Fritsche, A.; Häring, H.U. Identification and characterization of metabolically benign obesity in humans. Arch. Intern. Med. 2008, 168, 1609–1616. [Google Scholar] [CrossRef] [PubMed]
- Blüher, M. Adipose tissue dysfunction contributes to obesity related metabolic diseases. Best Pract. Res. Clin. Endocrinol. Metab. 2013, 27, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Manolopoulos, K.N.; Karpe, F.; Frayn, K.N. Gluteofemoral body fat as a determinant of metabolic health. Int. J. Obes. 2010, 34, 949–959. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.; Fontana, L.; Young, L.; Coggan, A.R.; Kilo, C.; Patterson, B.W.; Mohammed, S. Absence of an effect of liposuction on insulin action and risk factors for coronary heart disease. N. Engl. J. Med. 2004, 350, 2549–2557. [Google Scholar] [CrossRef] [PubMed]
- Thörne, A.; Lönnqvist, F.; Apelman, J.; Hellers, G.; Arner, P. A pilot study of long-term effects of a novel obesity treatment: Omentectomy in connection with adjustable gastric banding. Int. J. Obes. 2002, 26, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Ameer, F.; Scandiuzzi, L.; Hasnain, S.; Kalbacher, H.; Zaidi, N. De novo lipogenesis in health and disease. Metabolism 2014, 63, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Diraison, F.; Dusserre, E.; Vidal, H.; Sothier, M.; Beylot, M. Increased hepatic lipogenesis but decreased expression of lipogenic gene in adipose tissue in human obesity. Am. J. Physiol. Endocrinol. Metab. 2002, 282, 46–51. [Google Scholar]
- Letexier, D.; Pinteur, C.; Large, V.; Fréring, V.; Beylot, M. Comparison of the expression and activity of the lipogenic pathway in human and rat adipose tissue. J. Lipid Res. 2003, 44, 2127–2134. [Google Scholar] [CrossRef] [PubMed]
- Minehira, K.; Vega, N.; Vidal, H.; Acheson, K.; Tappy, L. Effect of carbohydrate overfeeding on whole body macronutrient metabolism and expression of lipogenic enzymes in adipose tissue of lean and overweight humans. Int. J. Obes. Relat. Metab. Disord. 2004, 28, 1291–1298. [Google Scholar] [CrossRef] [PubMed]
- Hudgins, L.C.; Baday, A.; Hellerstein, M.K.; Parker, T.S.; Levine, D.M.; Seidman, C.E.; Neese, R.A.; Tremaroli, J.D.; Hirsch, J. The effect of dietary carbohydrate on the genes for fatty acid synthase and inflammatory cytokines in adipose tissue from lean and obese subjects. J. Nutr. Biochem. 2008, 19, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Ranganathan, G.; Unal, R.; Pokrovskaya, I.; Yao-Borengasser, A.; Phanavanh, B.; Lecka-Czernik, B.; Rasouli, N.; Kern, P.A. The lipogenic enzymes DGAT1, FAS, and LPL in adipose tissue: Effects of obesity, insulin resistance, and TZD treatment. J. Lipid Res. 2007, 47, 2444–2450. [Google Scholar] [CrossRef] [PubMed]
- Tinahones, F.J.; Garrido-Sanchez, L.; Miranda, M.; García-Almeida, J.M.; Macias-Gonzalez, M.; Ceperuelo, V.; Gluckmann, E.; Rivas-Marin, J.; Vendrell, J.; García-Fuentes, E. Obesity and insulin resistance-related changes in the expression of lipogenic and lipolytic genes in morbidly obese subjects. Obes. Surg. 2010, 20, 1559–1567. [Google Scholar] [CrossRef] [PubMed]
- Goto, T.; Lee, J.Y.; Teraminami, A.; Kim, Y.I.; Hirai, S.; Uemura, T.; Inoue, H.; Takahashi, N.; Kawada, T. Activation of peroxisome proliferator-activated receptor-alpha stimulates both differentiation and fatty acid oxidation in adipocytes. J. Lipid Res. 2011, 52, 873–884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.Y.; Hashizaki, H.; Goto, T.; Sakamoto, T.; Takahashi, N.; Kawada, T. Activation of peroxisome proliferator-activated receptor-α enhances fatty acid oxidation in human adipocytes. Biochem. Biophys. Res. Commun. 2011, 407, 818–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Lee, C.; Tiep, S.; Yu, R.T.; Ham, J.; Kang, H.; Evans, R.M.; Jolla, L. Peroxisome-proliferator-activated receptor d activates fat metabolism to prevent obesity. Cell 2003, 113, 159–170. [Google Scholar] [CrossRef]
- Auguet, T.; Guiu-Jurado, E.; Berlanga, A.; Terra, X.; Martinez, S.; Porras, J.A.; Ceausu, A.; Sabench, F.; Hernandez, M.; Aguilar, C.; et al. Downregulation of lipogenesis and fatty acid oxidation in the subcutaneous adipose tissue of morbidly obese women. Obesity (Silver Spring) 2014, 22, 2032–2038. [Google Scholar] [CrossRef] [PubMed]
- Dahlman, I.; Linder, K.; Nordstrom, E.A.; Andersson, I.; Liden, J.; Verdich, C.; Sorensen, T.; Arner, P. Changes in adipose tissue gene expression with energy restricted diets in obese women. Am. J. Clin. Nutr. 2005, 81, 1275–1285. [Google Scholar] [PubMed]
- Dolinková, M.; Dostálová, I.; Lacinová, Z.; Michalský, D.; Haluzíková, D.; Mráz, M.; Kasalický, M.; Haluzík, M. The endocrine profile of subcutaneous and visceral adipose tissue of obese patients. Mol. Cell. Endocrinol. 2008, 291, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Ortega, F.J.; Mayas, D.; Moreno-Navarrete, J.M.; Catalán, V.; Gómez-Ambrosi, J.; Esteve, E.; Rodriguez-Hermosa, J.I.; Ruiz, B.; Ricart, W.; Peral, B.; et al. The gene expression of the main lipogenic enzymes is downregulated in visceral adipose tissue of obese subjects. Obesity (Silver Spring) 2009, 18, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Galilea, M.; Pérez-Matute, P.; Prieto-Hontoria, P.L.; Sáinz, N.; López-Yoldi, M.; Houssier, M.; Martínez, J.A.; Langin, D.; Moreno-Aliaga, M.J. A lipoic acid reduces fatty acid esterification and lipogenesis in adipocytes from overweight/obese subjects. Obesity 2014, 22, 2210–2215. [Google Scholar] [CrossRef] [PubMed]
- Lodhi, I.J.; Yin, L.; Jensen-urstad, A.P.L.; Funai, K.; Coleman, T.; Baird, H.; Ramahi, M.K.; Razani, B.; Song, H.; Fu-hsu, F.; et al. Inhibiting adipose tissue lipogenesis reprograms thermogenesis and PPARγ activation to decrease diet-induced obesity. Cell Metab. 2013, 16, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Mardinoglu, A.; Kampf, C.; Asplund, A.; Fagerberg, L.; Hallstro, M.; Edlund, K.; Blu, M.; Ponte, F.; Uhlen, M.; Nielsen, J. Defining the human adipose tissue proteome to reveal metabolic alterations in obesity. J. Proteome Res. 2014, 13, 5106–5119. [Google Scholar] [CrossRef] [PubMed]
- MacLaren, R.E.; Cui, W.; Lu, H.; Simard, S.; Cianflone, K. Association of adipocyte genes with ASP expression: A microarray analysis of subcutaneous and omental adipose tissue in morbidly obese subjects. BMC Med. Genom. 2010, 3, 3. [Google Scholar] [CrossRef] [PubMed]
- Ebbert, J.; Jensen, M. Fat depots, free fatty acids, and dyslipidemia. Nutrients 2013, 5, 498–508. [Google Scholar] [CrossRef] [PubMed]
- Klöting, N.; Blüher, M. Adipocyte dysfunction, inflammation and metabolic syndrome. Rev. Endocr. Metab. Disord. 2014, 15, 277–287. [Google Scholar] [CrossRef] [PubMed]
- Després, J.P.; Lemieux, I. Abdominal obesity and metabolic syndrome. Nature 2006, 444, 881–887. [Google Scholar] [CrossRef] [PubMed]
- Kotronen, A.; Yki-Järvinen, H.; Sevastianova, K.; Bergholm, R.; Hakkarainen, A.; Pietiläinen, K.H.; Juurinen, L.; Lundbom, N.; Sørensen, T.I. Comparison of the relative contributions of intra-abdominal and liver fat to components of the metabolic syndrome. Obesity (Silver Spring) 2011, 19, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Savage, D.B.; Tan, G.D.; Acerini, C.L.; Jebb, S.A.; Agostini, M.; Gurnell, M.; Williams, R.L.; Umpleby, A.M.; Thomas, E.L.; Bell, J.D.; et al. Human metabolic syndrome resulting from dominant-negative mutations in the nuclear receptor peroxisome proliferator-activated receptor-γ. Diabetes 2003, 52, 910–917. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, D.P.; Forbes, S.; Walker, B.R. Glucocorticoids and fatty acid metabolism in humans: Fuelling fat redistribution in the metabolic syndrome. J. Endocrinol. 2008, 197, 189–204. [Google Scholar] [CrossRef] [PubMed]
- Yim, J.E.; Heshka, S.; Albu, J.B.; Heymsfield, S.; Gallagher, D. Femoral-gluteal subcutaneous and intermuscular adipose tissues have independent and opposing relationships with CVD risk. J. Appl. Physiol. 2008, 104, 700–707. [Google Scholar] [CrossRef] [PubMed]
- Snijder, M.B.; Dekker, J.M.; Visser, M.; Yudkin, J.S.; Stehouwer, C.D.A.; Bouter, L.M.; Heine, R.J.; Nijpels, G.; Seidell, J.C.; Coen, D.A.; et al. Larger thigh and hip circumferences are associated with better glucose tolerance: The hoorn study. Obes. Res. 2003, 11, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Snijder, M.B.; Dekker, J.M.; Visser, M.; Bouter, L.M.; Stehouwer, C.D.A.; Yudkin, J.S.; Heine, R.J.; Nijpels, G.; Seidell, J.C. Trunk fat and leg fat have independent and opposite associations with fasting and postload glucose levels: The Hoorn study. Diabetes Care 2004, 27, 372–377. [Google Scholar] [CrossRef] [PubMed]
- Pelt, R.E.V.A.N.; Evans, E.M.; Schechtman, K.B.; Ehsani, A.A.; Kohrt, W.M.; Contributions, W.M.K. Contributions of total and regional fat mass to risk for cardiovascular disease in older women. Am. J. Physiol. Endocrinol. Metab. 2002, 282, 1023–1028. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.K.; Gavrilova, O.; Chen, Y.; Reitman, M.L.; Shulman, G.I. Mechanism of insulin resistance in A-ZIP/F-1 fatless mice. J. Biol. Chem. 2000, 275, 8456–8460. [Google Scholar] [CrossRef] [PubMed]
- Huang-Doran, I.; Sleigh, A.; Rochford, J.J.; O’Rahilly, S.; Savage, D.B. Lipodystrophy: Metabolic insights from a rare disorder. J. Endocrinol. 2010, 207, 245–255. [Google Scholar] [CrossRef] [PubMed]
- Foster, M.T.; Softic, S.; Caldwell, J.; Kohli, R.; DeKloet, A.D.; Seeley, R.J. Subcutaneous adipose tissue transplantation in diet-induced obese mice attenuates metabolic dysregulation while removal exacerbates it. Physiol. Rep. 2013, 1, e00015. [Google Scholar] [CrossRef] [PubMed]
- Virtue, S.; Vidal-Puig, A. It’s not how fat you are, it's what you do with it that counts. PLoS Biol. 2008, 6, e237. [Google Scholar] [CrossRef] [PubMed]
- Geer, E.B.; Shen, W. Gender differences in insulin resistance, body composition, and energy balance. Gend. Med. 2009, 6, 60–75. [Google Scholar] [CrossRef] [PubMed]
- Karastergiou, K.; Smith, S.R.; Greenberg, A.S.; Fried, S.K. Sex differences in human adipose tissues—The biology of pear shape. Biol. Sex Differ. 2012, 3, 13. [Google Scholar] [CrossRef] [PubMed]
- Varlamov, O.; Bethea, C.L.; Roberts, C.T. Sex-specific differences in lipid and glucose metabolism. Front. Endocrinol. (Lausanne) 2015, 5, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Association, A.D. Classification and diagnosis of diabetes. Diabetes Care 2015, 38, S8–S16. [Google Scholar] [CrossRef] [PubMed]
- Terra, X.; Auguet, T.; Broch, M.; Sabench, F.; Hernández, M.; Pastor, R.M.; Quesada, I.M.; Luna, A.; Aguilar, C.; del Castillo, D.; et al. Retinol binding protein-4 circulating levels were higher in nonalcoholic fatty liver disease vs. histologically normal liver from morbidly obese women. Obesity 2013, 21, 170–177. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guiu-Jurado, E.; Auguet, T.; Berlanga, A.; Aragonès, G.; Aguilar, C.; Sabench, F.; Armengol, S.; Porras, J.A.; Martí, A.; Jorba, R.; et al. Downregulation of de Novo Fatty Acid Synthesis in Subcutaneous Adipose Tissue of Moderately Obese Women. Int. J. Mol. Sci. 2015, 16, 29911-29922. https://doi.org/10.3390/ijms161226206
Guiu-Jurado E, Auguet T, Berlanga A, Aragonès G, Aguilar C, Sabench F, Armengol S, Porras JA, Martí A, Jorba R, et al. Downregulation of de Novo Fatty Acid Synthesis in Subcutaneous Adipose Tissue of Moderately Obese Women. International Journal of Molecular Sciences. 2015; 16(12):29911-29922. https://doi.org/10.3390/ijms161226206
Chicago/Turabian StyleGuiu-Jurado, Esther, Teresa Auguet, Alba Berlanga, Gemma Aragonès, Carmen Aguilar, Fàtima Sabench, Sandra Armengol, José Antonio Porras, Andreu Martí, Rosa Jorba, and et al. 2015. "Downregulation of de Novo Fatty Acid Synthesis in Subcutaneous Adipose Tissue of Moderately Obese Women" International Journal of Molecular Sciences 16, no. 12: 29911-29922. https://doi.org/10.3390/ijms161226206