Abstract
Strawberry (Fragaria × ananassa Duch.) propagation has evolved significantly over the past 20 years, transitioning from traditional field nursery systems to advanced, controlled, environment production. This review synthesizes recent advances in propagation methods, environmental regulation, and disease management strategies. Traditional field systems face mounting challenges from soilborne pathogens (Neopestalotiopsis species, Phytophthora cactorum, Verticillium dahliae) and regulatory restrictions on methyl bromide fumigation. Plug plant technology offers 80–95% disease reduction and 3–7-week production cycles versus 12–16-weeks traditional cycles, although at higher unit costs. Advanced tray plant systems developed in the Netherlands enable 10–11 months cold storage and programmed year-round production schedules. Elevated bench propagation systems have emerged as dominant commercial technology in East Asian regions, particularly Korea and Japan, where disease pressure necessitated alternatives to conventional nurseries. Micropropagation via temporary immersion bioreactors achieves 50–100% higher multiplication rates, while ensuring virus-free status. Environmental control research reveals complex photoperiod–temperature-chilling interactions regulating dormancy and flowering. Emerging technologies include F1 hybrid seed propagation and AI-driven automation, achieving 15–25% energy efficiency gains. Despite progress, challenges remain in cost optimization, climate adaptation, and region-specific protocols. This review provides a comparative framework for nursery system selection under evolving climatic and regulatory constraints, identifying critical knowledge gaps and future research priorities for sustainable strawberry propagation.
1. Introduction
Strawberry (Fragaria × ananassa Duch.) ranks among the most economically important small fruit crops worldwide, with global production exceeding 9 million tons annually [1]. Unlike most fruit crops that are seed-propagated, commercial strawberry cultivation relies primarily on vegetative propagation to maintain cultivar identity and desired fruit-quality characteristics. This dependence on clonal propagation presents both the advantages and challenges that have shaped the evolution of nursery production systems over the past two decades.
Traditional strawberry propagation involves establishing mother plants in field nurseries, allowing runners (stolons) to develop, and harvesting daughter plants for commercial production. While this system has served the industry for generations, it faces mounting pressures from pathogen accumulation, particularly Colletotrichum gloeosporioides, Phytophthora cactorum and Verticillium dahliae, regulatory restrictions on soil fumigants like methyl bromide, and the need for more sustainable production practices [2,3,4]. The phase-out of methyl bromide under the Montreal Protocol has accelerated the search for alternative propagation strategies that minimize disease pressure while maintaining production efficiency.
The past 20 years have witnessed remarkable innovations in strawberry propagation technology. The 1998 symposium on strawberry plug transplant technology at the XXV International Horticultural Congress in Brussels marked a turning point, bringing global attention to soilless propagation methods and plug tray-based systems [5]. Building on these innovations, elevated bench tray propagation systems emerged as a significant advancement, particularly in East Asian countries, where space optimization and improved sanitation through raised growing platforms became normal nursery practice. Concurrently, advances in tissue culture protocols enabled commercial-scale micropropagation, offering pathogen-free starting material and accelerated multiplication rates [6,7]. More recently, controlled environment agriculture (CEA) technologies have revolutionized nursery production, providing unprecedented control over the environmental factors that regulate plant development.
The environmental regulation of strawberry development is complex, involving intricate interactions between photoperiod, temperature, and chilling requirements that control dormancy induction, flowering, and runnering [8,9]. Understanding these relationships has been crucial for developing propagation systems that can produce high-quality transplants year-round. Short-day (SD) cultivars require specific low-temperature and short-photoperiod conditions for flower bud initiation, while day-neutral and everbearing types respond differently, enabling production in diverse climatic conditions [10].
This review synthesizes advances in strawberry propagation over the past two decades, examining the evolution from traditional field systems to modern controlled environment production. We analyze developments in plug and tray plant technology, micropropagation systems, environmental control strategies, disease management approaches, and emerging technologies including F1 hybrid seed propagation. By identifying current knowledge gaps and future research priorities, this review aims to guide the continued development of sustainable, efficient propagation systems for the global strawberry industry.
This review also addresses critical knowledge gaps including: (1) economic viability comparisons across propagation systems at different production scales; (2) environmental sustainability assessments incorporating carbon footprint and resource use; (3) region-specific adaptation strategies for diverse climatic zones; and (4) integration pathways for emerging biotechnologies. The structure progresses from established technologies to emerging innovations, concluding with comparative analyses and future research priorities.
2. Traditional Runner-Based Propagation Systems
2.1. Field Nursery Production
Traditional strawberry propagation in field nurseries has remained the predominant method globally, particularly in major production regions such as California, Spain, and Italy [4]. The system involves establishing certified mother plants at densities of 15,000–25,000 plants per hectare in fumigated soil, typically on raised beds covered with plastic mulch. Mother plants produce runners (stolons) that develop into daughter plants throughout the growing season, with multiplication ratios typically ranging from 1:8 to 1:15, depending on the cultivar, climate, and management practices [11].
California’s nursery system exemplifies large-scale traditional propagation, where certified mother plants from foundation stock are established in April–May; runners are harvested from September through October. The harvested plants are graded, typically as fresh plants, and shipped to production fields. Fresh plants are used immediately after harvest, while frigo plants are cold-stored at −2 to 0 °C for extended periods, providing production flexibility [4,12,13]. The frigo plant system enables year-round planting by accumulating chilling hours during storage, essential for subsequent flowering in many cultivars.
In Europe, frigo plants for strawberry production in cold temperate areas are produced in the northern cold areas, planting mother plants in April and harvesting and cleaning them from January to March, then cold-stored until plantation late July–early August.
For strawberry cultivation in warm temperate areas, of high interest is the production of fresh plants, produced by mother plants planted always in colder areas, planted in March–April and harvested in August–September, for direct planting in southern areas in October [2,14,15].
2.2. Challenges and Limitations
The sustainability of traditional field propagation faces several critical challenges for both frigo and fresh plants produced in soil conditions. Soil-borne pathogens, particularly Colletotrichum gloeosporioides and Phytophthora cactorum, which causes crown rot, and Verticillium dahlia, which causes verticillium wilt, accumulate over successive production cycles, necessitating intensive soil fumigation [16,17]. Phytophthora infections can reduce plant survival by 30–60% in untreated fields, with economic losses estimated at USD 40–60 million annually in the United States alone [18]. The pathogen’s zoospores disperse through splash water and contaminated soil, making control challenging in field conditions.
Regulatory restrictions on methyl bromide and chloropicrin have complicated disease management. The Montreal Protocol mandated methyl bromide phase-out by 2005 in developed countries, with critical use exemptions granted for strawberry nurseries due to the lack of effective alternatives [19]. However, these exemptions are progressively being eliminated, driving the search for alternative fumigants and propagation systems. Chemical alternatives, like 1,3-dichloropropene and metam sodium, provide incomplete control and environmental concerns remain [20]. Non-chemical approaches, including soil solarization, anaerobic soil disinfestation, and resistant cultivars, show promise but require further development for commercial-scale implementation.
2.3. Strawberry Propagation in China: The World’s Largest Producer
China accounts for approximately 40% of total world production, with output exceeding 3.3 million tons in 2022 and reaching 3.4 million tons in 2023 [21], making the integrity of its propagation system a matter of global consequence.
The most significant challenge in Chinese strawberry propagation is the near-complete absence of a certified, disease-free, nursery supply chain. On-farm propagation remains the predominant practice, with growers maintaining their own mother plants from season to season without systematic virus testing or phytosanitary certification. This propagation structure, combined with the lack of adequate cold-storage infrastructure and certification programs, results in progressive viral and pathogen load accumulation, contributing to yield decline and quality inconsistency across successive cropping cycles [22].
Public research programs at Shandong Agricultural University, Beijing Academy of Agriculture and Forestry Sciences, and research institutes in Liaoning and Hebei have begun addressing virus elimination through shoot tip and anther culture techniques. A recent study from Shandong Agricultural University demonstrated a 98.39% virus-free rate in ‘Miaoxiang 7’ using anther culture, representing a significant methodological advance over the theoretical limit of 80–85% achievable by conventional shoot-tip culture [23].
California-based Lassen Canyon Nursery has established operations in China [24], indicating early-stage internationalization of professional nursery production practices. This development suggests that the Chinese industry is at an inflection point regarding the transition from on-farm propagation to professional nursery supply chains—a transition that parallels the evolution seen in European and North American industries several decades earlier.
2.4. Global Nursery Plant Production by Region
The scale of strawberry nursery plant production varies enormously across major producing regions, reflecting differences in industry structure, climate, regulatory environment, and certification requirements. Table 1 summarizes available data on strawberry nursery plant production volumes across key countries and regions. It should be noted that comprehensive, standardized nursery plant production statistics are not systematically collected at the international level; the figures presented below represent estimates derived from published industry reports, ISHS symposium proceedings, and national agricultural agency data. The distinction between formal certified plant production and informal on-farm propagation is particularly difficult to quantify, especially in countries, such as China, where the latter predominates.

Table 1.
Estimated annual strawberry nursery plant production and dominant propagation systems by major producing country/region. Sources: ISHS Acta Horticulturae symposium reports; FAO FAOSTAT (2023); California Strawberry Commission; national agricultural agencies.
3. Plug and Tray Plant Production Systems
3.1. Development and Advantages
In this review, the term “plug plant” refers to runner tips rooted individually in soilless substrate cells for short-term propagation (3–5 weeks), whereas “tray plant” describes an advanced Dutch system featuring multi-crown plants in modular trays designed for extended cold storage (10–11 months) and programmed production scheduling.
Plug plant technology emerged as a transformative alternative to traditional propagation, addressing many limitations of field-based systems. The system involves rooting unrooted runner tips directly into cellular trays containing peat-based or rockwool media under protected cultivation [34,35]. Tips are harvested from mother plants and immediately inserted into 42–72 cell trays, where they develop roots within 3–5 weeks under controlled environmental conditions. This approach eliminates the soil phase entirely, dramatically reducing soilborne disease risk.
The advantages of plug production are substantial. Disease incidence, particularly Phytophthora and Verticillium, is reduced by 80–95% compared to soil-grown plants, as the peat-based root medium provides no infection pathway for root-invading pathogens [36]. The primary infection site for Phytophthora is the meristematic region behind root caps; stem tissues, like stolons, are not infection avenues, making runner tips ideal propagules for disease avoidance [37]. Production cycles are compressed to 3–5 weeks in warm climates, compared to 12–16 weeks for traditional field production, enabling multiple production cycles annually [5,38].
Plug plants exhibit superior uniformity and transplant success rates of 95–98%, compared to 85–92% for bare-root plants [38]. The intact root ball minimizes transplant shock; controlled production conditions enable the precise management of plant architecture and development stage. Shipping and handling are simplified, as plug plants are more robust than bare-root plants, and transplanting can be mechanized more effectively. Water and fertilizer use efficiency is enhanced through precise fertigation management in the soilless system.
3.2. Production Protocols and Specifications
Optimal runner tip quality is critical for plug production success. Industry specifications recommend tips with two intact trifoliate leaves, a crown diameter of 4–10 mm, and the oldest trifoliate extending 6.5–10 cm from the crown base [5,39]. Tips should be harvested frequently (2–3 times weekly during peak season) to maintain size uniformity within acceptable ranges. Harvesting certified stock plants ensures genetic trueness and phytosanitary status, with foundation material typically sourced from nuclear stock programs [40].
Production environments range from simple shade structures in moderate climates to sophisticated climate-controlled greenhouses in extreme environments. Temperature management is critical during rooting, with optimal ranges of 18–24 °C promoting rapid root initiation while minimizing disease pressure [39]. Relative humidity of 85–95% is maintained during the first week through misting systems, then reduced to 70–80% to prevent fungal diseases while supporting continued root development. Light levels of 200–400 μmol m−2 s−1 provide sufficient photosynthesis without excessive transpiration stress.
Fertigation programs typically begin with low electrical conductivity (EC 0.5–0.8 dS m−1) during the first week to minimize osmotic stress, increasing to EC 1.2–1.8 dS m−1 as roots develop [41]. Balanced NPK ratios (20-20-20 or 15-15-15) support vegetative growth, with modifications based on tissue analysis and cultivar requirements [42,43]. Container size significantly affects plant development, with larger volumes (80–100 mL) producing more vigorous plants but requiring longer production cycles and increased substrate costs [44].
3.3. Economic Considerations
Plug plant production costs typically exceed bare-root plants by 30–100%, with per-plant costs ranging from USD 0.12–0.25 depending on scale, location, and cultivar [5]. However, total production costs may be comparable when accounting for reduced disease losses, improved transplant success, eliminated soil fumigation, and shortened production cycles. The capital investment for greenhouse infrastructure and climate control equipment represents a significant barrier for small-scale nurseries, although cooperative production facilities and contract propagation services have emerged to address this challenge. Market adoption has been strongest in high-value production systems, including protected cultivation, organic production, and areas with severe disease pressure or fumigant restrictions. Although plug plants cost 30–100% more per unit, the reduction in disease-related losses (up to 80–95%) and higher establishment rates (95–98%) often offset the increased initial cost. Plug plant systems are particularly suitable for regions with high soilborne disease pressure or fumigant restrictions, whereas micropropagation is most effective for nuclear stock maintenance and rapid cultivar multiplication.
3.4. Comparative Yield Performance: Bare-Root Versus Containerized Plants
Despite the widespread adoption of plug and tray plant systems, direct yield comparisons between bare-root and containerized production systems remain relatively limited in the published literature. The available evidence, however, consistently indicates that plug and tray plants provide advantages in early-season yield and transplant establishment, although total seasonal yield outcomes vary by region, cultivar, and production system.
Cocco et al. [32] conducted a two-season field comparison of bare-root and plug plants across three cultivars under tunnel culture in Southern Italy, revealing that plug plants demonstrated earlier fruiting onset and significantly higher early-season yields, with production concentrated in March and early April—a commercially advantageous window in Mediterranean markets. Fruit quality implications of these propagation differences are discussed in Section 3.7. Containerized strawberry transplants were first developed for Florida’s winter production system, demonstrating significant advantages over bare-root plants in establishment and early yield [13,45,46]. In studies conducted in Florida, Torres-Quezada et al. [26] found that for ‘Florida Radiance’, total seasonal yield was equivalent between bare-root and plug plants (averaging 6.1 Mg·ha−1), while for ‘Strawberry Festival’, plug plants resulted in 58% more fruit than bare-root plants. The water savings associated with plug plant establishment were also substantial—approximately 80% less irrigation water was required during the establishment phase compared to bare-root systems, representing a potential saving of over 820,000 gallons per acre per season [47].
Open-field yield benchmarks for bare-root systems typically range from 15–25 t·ha−1 in temperate regions under conventional plasticulture (planting densities of 50,000–70,000 plants·ha−1), while high tunnel and greenhouse production using containerized plants can reach 40–60 t·ha−1 at higher planting densities (up to 100,000 plants·ha−1). In soilless elevated bench systems in Korea and Japan, yields of 30–50 t·ha−1 are reported for major cultivars such as ‘Seolhyang’ and ‘Benihoppe’. In southern Brazil, a comparison of five plant types for the day-neutral cultivar ‘Albion’ under soilless substrate culture found no significant difference in total yield per hectare between bare-root and tray plant types under the agroclimatic conditions of that region, although cold-preserved tray plants with mature roots exhibited superior vegetative performance in terms of crown and stolon development—whether this vegetative advantage translates into yield differences under extended production seasons warrants further investigation. A summary of representative yield data across production systems and regions is presented in Table 2.
Table 2.
Representative yield data for bare-root (frigo/fresh), plug, and tray plant systems across major strawberry production regions. Data are compiled from published field studies and regional industry reports.
3.5. Tray Plant Systems and Dutch Innovations
Tray plant technology represents a sophisticated evolution of plug plant systems, developed primarily in the Netherlands and Belgium since the early 1990s. Unlike basic plug plants used immediately after rooting, tray plants are grown to a more advanced developmental stage with multiple crowns before cold storage, enabling programmed year-round production [43,48]. This system has become the dominant plant type for soilless strawberry production in western Europe, with approximately 100 million plants produced annually. The Netherlands expanded certified tray plant production from 30 ha in 2004 to 135 ha by 2014, reflecting rapid market adoption [48].
The Dutch tray plant production system begins with certified runner plants established as mother plants in March in bags or containers. Mother plants are typically grown outdoors on horizontal systems, although some cultivars utilize vertical systems under protected cultivation. Non-rooted stolon tips are propagated in July into peat-filled multi-cell trays (230–300 cc per cell), larger than standard plugs, to accommodate extended growth [48,49]. Plants are cultivated until late November or mid-December, during which they naturally undergo flower differentiation under the Netherlands’ maritime climate conditions. The favorable climate provides sufficient temperatures for continued flower differentiation through October and adequate chilling accumulation (600–800 h below 7 °C) in November–December [48,49].
A distinctive feature of Dutch tray plant production is the systematic use of flower dissection (flower mapping) for production management [50]. This technique, pioneered by researchers at Fragaria Holland and Pcfruit, involves microscopic examination of meristems to assess flower initiation and differentiation stages [48]. Flower dissections are performed in early September to evaluate flower bud development, enabling precise adjustment of nitrate nutrition during critical flower differentiation periods. The nutritional status during flower differentiation (August to mid-October) significantly affects subsequent fruit number, fruit quality, and production timing [51,52]. Low nitrogen levels during this period can result in smaller plants with fewer flowers per truss and extended flowering periods, while excessive nitrogen may cause abnormalities such as phyllody. Additional flower dissections at harvest in late November or December estimate production potential, guiding grower decisions on optimal planting dates and plant densities [48].
Cold-storage characteristics distinguish tray plants from other propagule types. The larger substrate volume and multi-crown architecture provide greater carbohydrate reserves, enabling storage periods of 10–11 months compared to 6–8 months for standard frigo plants [53]. The intact root system and substrate buffer minimize transplant shock and improve establishment rates. After storage, tray plants demonstrate superior regrowth vigor, yield potential, and fruit size compared to bare-root alternatives, producing approximately 20% more large fruits and thereby reducing harvest labor requirements [14].
The tray plant system enables sophisticated year-round production schedules through the staggered planting of cold-stored materials [14]. In heated greenhouses, three production cycles are achievable annually using short-day cultivars such as ‘Elsanta’ and ‘Sonata’: late December planting for March–May harvest (5.5 kg m−2), late May planting for July–August harvest (3.5 kg m−2), and August planting for October–December harvest (4.5 kg m−2) [14]. Alternative systems involve August planting with overwintering, first harvest in October–December, followed by chilling and photoperiodic treatments (night break) in early spring to trigger a second harvest from late April through June (approximately 7 kg m−2). Under polytunnels, the July planting of tray plants yields 3–3.5 kg m−2 for autumn harvest, with potential extension through November in heated structures [48].
Nutritional management protocols for tray plants have been extensively optimized through Dutch research programs [49]. During the nursery phase, nitrogen application timing critically influences plant architecture and reproductive development [51,52]. Early applications in July–August promote runner formation, while later applications during shortening photoperiods stimulate crown development and flower initiation. Fertigation programs utilize balanced nutrient solutions with a pH of 5.5 and electrical conductivity of 1.2 mS m−1 for everbearing cultivars and 1.4 mS m−1 for short-day types [52]. The potassium-to-calcium ratio is maintained at 0.65 during vegetative growth and increased to 0.8 during flowering and fruiting to support reproductive demands.
Recent research has focused on photoperiod manipulation and LED lighting effects on tray plant performance [50,54]. Artificial day-length shortening in summer can advance flower initiation timing, avoiding the need for year-old plants as transplant material [55,56]. LED lighting during winter production cycles increases inflorescence numbers and improves productivity, particularly for short-day cultivars, by simulating long-day conditions that enhance flowering potential [57]. Additionally, research on temperature-photoperiod interactions using meristem dissection has refined understanding of flowering dynamics in both short-day and everbearing cultivars, enabling more precise crop timing [48].
The tray plant system’s economic advantages derive from multiple factors beyond reduced disease pressure. The programmed production capability allows growers to target high-value market windows and maintain continuous supply chains. The 20% increase in marketable large fruit (>25 g) enhances both commercial packout percentages and harvest labor efficiency through reduced picking time per unit marketable weight. Multi-crown architecture generates more fruiting sites per plant, increasing yields per unit area. However, tray plants command premium prices of USD 0.30–0.45 per plant compared to USD 0.12–0.25 for standard plug plants, requiring growers to achieve higher yields and quality premiums to justify the investment [5]. Market adoption has been strongest in high-input greenhouse systems, where precise crop timing and consistent quality command market premiums.
3.6. Elevated Bench Propagation Systems in East Asia
Elevated bench propagation systems have emerged as a dominant commercial technology in East Asian strawberry production regions, particularly in Korea and Japan, where soilborne disease pressure necessitated alternatives to conventional field nurseries. These systems integrate controlled environment principles with systematic runner guidance, achieving disease suppression comparable to advanced plug plant technologies while maintaining the multiplication efficiency characteristic of runner-based propagation [57,58].
The fundamental architecture consists of benches elevated 1.0–1.5 m above ground level, supporting narrow cultivation beds (20–30 cm width) that accommodate mother plants at standardized spacing. This elevated configuration eliminates soil contact for both mother and developing daughter plants, interrupting splash dispersal pathways for Colletotrichum gloeosporioides and Phytophthora cactorum—the primary foliar and root pathogens limiting conventional nursery production [43,58]. Bench height enables ergonomic standing-posture operations, significantly reducing labor fatigue during the intensive runner guidance and plant assessment activities that characterize these systems.
Mother plants are established in soilless substrate within elevated beds during early spring (March–April), utilizing peat-based mixtures amended with perlite or coir to optimize physical properties [57]. As runners emerge from axillary meristems during the long-day conditions of late spring and early summer (May–June), they are systematically directed into individual rooting containers positioned adjacent to the bench structure. This guided placement ensures uniform daughter plant spacing, facilitates systematic quality assessment, and enables selective harvesting based on developmental stage and morphological characteristics.
Individual daughter plants root in standardized plastic containers (9–11 cm diameter of each cell, 130–160 mL volume per cell, 24–32 cells per container) filled with peat-based substrate formulations optimized for root development [48,55]. Fertigation management follows progressive concentration protocols established for strawberry plug production: electrical conductivity maintained at 0.6–0.8 dS m−1 during initial rooting (weeks 1–2), gradually increased to 1.0–1.4 dS m−1 during active vegetative growth (weeks 3–6), and reduced to 0.8–1.0 dS m−1 during the pre-harvest hardening phase [55,56]. This EC modulation optimizes adventitious root initiation under low osmotic-stress conditions, supports crown and leaf development during active growth, and promotes carbohydrate accumulation and stress tolerance before transplant harvest.
Environmental management during the critical summer rooting period employs 40–50% shade netting to reduce photosynthetic photon flux density to manageable levels, combined with evaporative cooling systems to maintain canopy temperatures below stress thresholds [48]. High-frequency irrigation (4–8 cycles daily depending on weather conditions) sustains consistent substrate moisture, while overhead misting during the first 7–10 days post guidance creates favorable microclimate conditions for adventitious root emergence from runner nodes [59]. The complete production cycle from mother plant establishment to daughter plant harvest typically spans 150–180 days, with the intensive rooting and development phase occupying 90–120 days.
The elevated bench configuration achieves dramatic disease suppression through multiple complementary mechanisms. Elimination of soil contact provides primary pathogen avoidance, while improved air circulation reduces leaf wetness duration and canopy-level relative humidity [58,60]. Comparative observations from commercial operations document 85–95% reduction in anthracnose (Colletotrichum) incidence and 90–98% reduction in Phytophthora infection rates compared to conventional soil-based nurseries.
Recent innovations focus on further optimization and automation. Controlled environment propagation methods using artificial lighting have been investigated to extend production windows and enhance transplant quality during suboptimal natural light conditions [61]. Automated environmental monitoring and precision fertigation controllers enable data-driven management protocols that optimize resource use efficiency while maintaining consistent quality output [55,56]. Integration with micropropagated mother plant systems provides certified disease-free starting material for the elevated bench production cycle [62].
Ongoing challenges include: (1) high capital costs limiting adoption by small-scale nurseries, necessitating cooperative production models or contract arrangements; (2) thermal management demands during increasingly frequent heat waves, requiring advanced cooling strategies; (3) substrate waste management as used growing media accumulates annually, driving research into locally-sourced alternatives; and (4) energy consumption for cooling and irrigation, motivating efficiency improvements [48].
Research efforts address these challenges through the development of regionally adapted substrate formulations, optimized environmental control strategies, and cooperative production models that enable small-scale operations to access advanced infrastructure through shared facilities or contract propagation services. The system exemplifies the successful adaptation of global plug plant concepts to regional production conditions, demonstrating how the thoughtful integration of disease management, quality control, and labor efficiency principles with cultivar-specific requirements, climate constraints, and market structures can drive sustained competitive advantage in nursery production.
3.7. Fruit Quality Implications of Propagation System
Beyond yield quantity, propagation methods can influence the fruit quality parameters that determine commercial value and consumer acceptance. This dimension of the plug versus bare-root comparison has received less systematic investigation than yield metrics; however, the available evidence suggests that propagation systems affect both phenological quality parameters and nutritional composition.
Cocco et al. [32] provided the most detailed published comparison of fruit quality between bare-root and plug plants across multiple cultivars in southern Italy, finding that anthocyanin and ellagitannin content—key determinants of fruit color, antioxidant capacity, and nutritional value—were on average higher in plug plant fruit than in bare-root fruit. Total soluble solids content, titratable acidity, and flesh firmness showed high year-to-year variability in both systems, suggesting that climatic conditions during the harvest period exert a stronger influence on these parameters than the propagation system, per se. The earlier fruiting of plug plants, while commercially advantageous for early-season price premiums, was associated with a reduction in per-fruit weight relative to bare-root plants in the same season, reflecting the trade-off between earliness and fruit size common to many intensively managed strawberry systems.
For F1 hybrid seed propagation (Section 8.1), fruit-quality variability represents a more substantive commercial concern. Seed-propagated populations exhibit phenotypic segregation in fruit size, shape, color, and flavor even within inbred lines, and this heterogeneity has been identified as a primary barrier to commercial adoption in fresh-market production, where uniformity is a strict requirement for premium retail channels. The ‘Yotsuboshi’ cultivar, developed through collaborative Japanese breeding programs for year-round seed propagation, demonstrated sufficient uniformity for commercial evaluation; however, challenges in achieving the aesthetic and organoleptic consistency of elite vegetatively propagated cultivars remain [30,63].
Virus status of the propagation material has a well-documented effect on fruit quality. Plants derived from virus-infected stock—particularly those carrying Strawberry mottle virus or Strawberry mild yellow edge virus—exhibit stunting, reduced fruit size, and altered sugar profiles relative to virus-free controls. The practical implication is that the quality advantage of containerized systems is inseparable from their phytosanitary superiority: certified, PCR-tested planting material consistently delivers more predictable fruit quality than field-propagated bare-root plants sourced from unmanaged or informally managed nurseries [30,64].
4. Micropropagation and Tissue Culture Systems
4.1. In Vitro Multiplication Protocols and Applications
Micropropagation has evolved from a specialized laboratory technique to a commercially viable propagation method for strawberry plants, particularly for nuclear stock maintenance and rapid cultivar multiplication. Modern protocols achieve 4–8-fold multiplication per subculture cycle (typically 4–6 weeks), enabling the production of millions of plants from single meristem explants within 12 months [65,66,67]. Meristem culture remains the foundation of virus elimination programs, with heat treatment (37 °C for 4–6 weeks) combined with meristem excision (0.2–0.5 mm) effectively eliminating major strawberry viruses including Strawberry mottle virus (SMoV), Strawberry mild yellow edge virus (SMYEV), and Strawberry crinkle virus (SCV) [68].
Multiplication protocols have undergone significant evolution over the past two decades. Traditional protocols used high concentrations of plant growth regulators (0.5–2.0 mg L−1 benzyladenine (BA) for shoot proliferation and 0.1–0.5 mg L−1 indole-3-butyric acid (IBA) for rooting), which created substantial problems, including hyperhydricity, somaclonal variation, and poor post-acclimatization performance [33,69,70,71,72]. Modern protocols have been fundamentally reprogrammed to address these limitations, now utilizing significantly reduced cytokinin concentrations (0.2–0.5 mg L−1 BA) in the proliferation stage and eliminating auxin application during rooting [73]
Contemporary strawberry micropropagation serves two distinct commercial applications requiring different optimization strategies. For nuclear stock maintenance and mother plant production, the reduced hormone protocol (0.2–0.5 mg L−1 BA, no auxin) ensures genetic stability and superior plant quality for subsequent runner production systems. Micropropagated mother plants grown in soilless vertical systems represent the optimal solution for producing plug plants with the highest phytosanitary status and true-to-type fidelity. These micropropagated mother plants demonstrate enhanced runner and tip production capacity compared to conventionally propagated stock [74,75]
The integration of micropropagated mother plants with conventional runner production has emerged as a powerful hybrid approach. Research demonstrates that micropropagated mother plants produce 50–100% more runners compared to conventionally propagated mother plants, with superior uniformity and vigor [76]. This system maintains the economic advantages of runner propagation while capturing the disease-free status and rapid multiplication potential of tissue culture. Elevated growing systems using sterile substrates enable high-density mother plant production (10–15 plants m−2) with minimal disease pressure [75].
4.2. Acclimatization and Field Performance
The successful acclimatization of micropropagated plantlets remains critical for commercial adoption [77,78]. Modern protocols achieve survival rates of 90–95% through gradual reduction of relative humidity (95% → 70% over 2–3 weeks) and a progressive increase in light intensity (50 → 400 μmol m−2 s−1) [33]. Substrate selection affects acclimatization success, with peat-perlite mixtures (3:1 or 2:1 v/v) providing optimal water retention and aeration [79]. Pre-acclimatization treatments, including exposure to higher CO2 concentrations (1000–1500 ppm) or brief drought stress, can enhance subsequent field performance [80]. Field evaluations generally show comparable or superior fruit yield and quality from micropropagated plants compared to conventional propagation, although some studies report delayed maturity in the establishment year [77,78,81].
4.3. Bioreactor Systems and Scaling
Temporary immersion bioreactor (TIB) systems represent a significant advance in commercial micropropagation capacity. These systems expose cultures to a liquid medium periodically (typically 15 min every 4–12 h) rather than continuously, improving gas exchange, reducing hyperhydricity, and enabling larger-scale production with reduced labor [75,76]. Multiplication rates in TIB systems often exceed traditional static culture by 50–100%, with some protocols achieving 8–12-fold multiplication per cycle [72]. The RITA® (Vitropic S.A. Saint-Mathieu-de-Tréviers, France) system and similar platforms have been successfully commercialized for strawberry plants, with some nurseries operating bioreactor facilities producing millions of plantlets annually.
Bioreactor systems for the massive propagation of strawberry plants can be the future, but it is important to develop highly efficient protocols for the production of embryogenic calli and subsequent differentiation of stable somatic embryo, by using bioreactors. In reality, not many protocols are now available; more research is needed to achieve this result. When it can be demonstrated that plants obtained with this system will result in a limited phenotypic variation, it will become the new frontier for a revolution in strawberry nursery production [72].
5. Environmental Control and Dormancy Regulation
5.1. Photoperiod and Temperature Interactions
An understanding of the environmental regulation of strawberry development has progressed substantially through controlled environment studies. The classic paradigm of short-day (SD) cultivars requiring short photoperiods (<14 h) and cool temperatures (<15 °C) for flower induction has been refined to reveal more nuanced genotype–environment interactions [54,82,83,84]. Recent phytotron studies demonstrate that dormancy induction in both short-day and everbearing cultivars requires the interaction of photoperiod, temperature, and exposure duration, with substantial cultivar variation in response thresholds [8,50,55].
Long photoperiods (>14 h), combined with moderate to warm temperatures (18–26 °C), strongly promote runner formation while suppressing flowering in short-day cultivars, providing the basis for efficient mother plant production [83]. The molecular mechanisms underlying photoperiodic control involve the circadian clock genes and flowering locus T (FT) homologs, with recent genomic studies identifying key regulatory pathways [85,86,87]. Short photoperiods (<12 h) at moderate temperatures (10–15 °C) induce dormancy characterized by reduced leaf and crown growth, shortened petioles, and decreased photosynthetic capacity [8,88].
5.2. Chilling Requirements and Manipulation
Chilling requirements for breaking dormancy and promoting flowering vary substantially among cultivar groups. Low-chill cultivars require 200–400 h below 7 °C, while high-chill types need 600–1000+ h [89]. The commercial manipulation of chilling has enabled production flexibility through techniques including: (1) night cooling using refrigerated air (12–16 h at 8–12 °C daily); (2) continuous cold storage in darkness (−2 to 2 °C); (3) continuous cold with supplemental lighting; (4) intermittent cold storage alternating with ambient conditions; and (5) crown cooling using chilled water circulation [90,91].
Optimal chilling temperatures range from −2 to 7 °C, with maximum efficacy typically around 2–5 °C [89]. Temperatures below −3 °C cause freezing injury, while those above 10 °C provide minimal chilling effect. Duration requirements depend on cultivar and subsequent forcing conditions, ranging from 2–4 weeks for low-chill types to 8–12 weeks for high-chill cultivars [90]. Light during chilling can modify plant responses, with low-intensity lighting (5–10 μmol m−2 s−1) helping to maintain plant viability during extended storage while providing a partial chilling effect [91].
5.3. Runner Production Optimization
Maximizing runner production in mother plant systems requires careful environmental manipulation. Long photoperiods (16–20 h) at 20–25 °C optimize stolon initiation and daughter plant formation [43,87,92]. LED lighting technology enables precise spectral control, with red:far-red ratios and blue light intensity affecting runner production. Higher red:far-red ratios (>1.2) and moderate blue light percentages (15–25%) promote vegetative growth and runner formation [14,93]. Growth regulators, particularly gibberellic acid (GA3) at 50–100 ppm, can enhance runner production by 30–50% in some cultivars, though effects are genotype-dependent and may reduce crown quality [91,94]. Nutritional management is critical, with higher nitrogen levels (150–200 ppm N) and adequate micronutrients supporting vigorous runner production, although excessive nitrogen can increase disease susceptibility [95].
By using microprogated mother plants, a new protocol for the production cycle, from the in vitro proliferation stage to the plug plants’ rooting, must be defined in order to achieve high-quality plug plants with respect to the plantation timing requested by the growers.
6. Cold Storage Systems and Frigo Plant Technology
6.1. Storage Protocols and Plant Quality
The cold storage of strawberry transplants enables production flexibility by decoupling propagation from planting schedules. Traditional frigo plants are bare-root plants harvested during dormancy, graded by crown diameter (A, A+, WB classes), and stored at −2 to 0 °C with high humidity (90–95%) for periods ranging from weeks to months [14]. Recent advances include tray plant storage, where rooted plug plants are cold-stored in their containers, maintaining root integrity and enabling faster field establishment [15].
Storage temperature management is critical for maintaining plant quality. Optimal temperatures range from −2 to 2 °C, with precise control (±0.5 °C) essential to prevent freezing injury below −3 °C or premature sprouting above 3 °C [53]. Recent research examining plug plant storage demonstrates that temperature fluctuations during storage significantly impact post-storage performance, with plants exposed to stable low temperatures (−0.5 to 1 °C) showing 15–25% better establishment than those experiencing variable temperatures [91]. Light exposure during storage has complex effects, with complete darkness maintaining dormancy longest, while low-intensity lighting (5 μmol m−2 s−1) can help to maintain photosynthetic capacity and improve post-storage recovery [96].
6.2. Post-Storage Management
The successful utilization of cold-stored plants requires careful post-storage management. Gradual warming (4–8 h at 10–15 °C) before field planting reduces transplant shock and improves establishment [97]. For plug plants, a 24–48 h acclimatization period with misting (RH 90–95%) facilitates the recovery of photosynthetic function and root activity [91]. Planting depth, irrigation scheduling, and protection from temperature extremes during establishment are critical for optimizing plant performance. Storage duration effects vary by cultivar, with most types tolerating 2–4 months of cold storage without significant quality loss, although some sensitive cultivars show reduced vigor after extended storage periods [98]. Crown diameter at storage correlates with post-storage flower and fruit development in predictable patterns, enabling the strategic timing of planting for desired harvest windows.
6.3. Implications for Nursery Scheduling and Programmed Transplant Supply
Modern strawberry nurseries increasingly operate as “programmed supply systems”, in which transplant quality and flowering readiness are intentionally aligned with planting windows [5,34,35]. Practical scheduling, therefore, requires integrating: (i) environmental regulation of runnering vs. flowering; (ii) runner-tip availability and rooting capacity; and (iii) cold-storage strategy (bare-root frigo vs. tray/plug storage) to match market-driven production calendars.
- (1)
- Environmental steering to separate “runnering mode” vs. “flowering-ready mode”
A core scheduling problem is that nurseries require high runner multiplication early but later need transplants that establish quickly and flower predictably after planting. The literature supports using long-day + warm conditions to promote runner formation [59,99,100], while shorter days and cooler conditions contribute to dormancy/flowering readiness depending on cultivar type [61,99,101]. Thus, a two-phase nursery schedule—(i) runner multiplication phase and (ii) transplant finishing/conditioning phase—can be justified as a general decision framework rather than cultivar-by-cultivar trial-and-error [61,101]. Scheduling becomes easier if nurseries explicitly classify cultivars into short-day vs. day-neutral/everbearing response groups and define a standard “finishing recipe” per group (photoperiod/temperature duration), rather than relying solely on calendar dates [61,99,101].
- (2)
- Scheduling the runner-tip rooting window (plug plants).
Plug plant systems allow rapid production cycles [5,34,35]; however, the plugging date and early nursery environment can strongly influence subsequent floral behavior, particularly in short-day cultivars [59,99,100]. Earlier plugging during mid-summer has been associated with a higher probability of fall flower induction in short-day types, indicating that nurseries can “steer” transplant developmental status by choosing plugging dates and early conditioning regimes for specific production targets.
For indoor or controlled propagation, light intensity and photoperiod can be used to increase propagation throughput and standardize quality [9,61], helping nurseries stabilize output when outdoor mother-plant runner supply is variable. Recent controlled-environment studies on indoor runner-tip propagation [59,102] provide operational ranges and highlight that propagation environments (PPFD and photoperiod) are not only biological variables but also levers for production planning.
A practical nursery plan benefits from defining (a) a weekly runner-tip harvest rhythm; (b) a fixed rooting cycle length (e.g., ~3–7 weeks, depending on cultivar and temperature) [32,42,63]; and (c) a target transplant “readiness state” (vegetative vs. flower-inductive) required by the receiving production system (field plasticulture vs. protected/off-season forcing) [5,101].
- (3)
- Cold storage as a scheduling buffer (bare-root frigo vs. tray/plug storage).
Cold storage enables decoupling propagation from planting, functioning as a “buffer” that absorbs weather risk and market-driven planting shifts. However, storage conditions (temperature stability and the presence/absence of low-intensity light) can affect post-storage performance and establishment [92,100,101,103]. Controlled studies on cold-storage temperature and light regimes in modern cultivars [56,90,92,104] indicate that maintaining stable low temperatures can improve post-storage performance and that storage protocols should be treated as part of the scheduling system, not merely logistics.
For plug/tray plants, storage retains root integrity and may support faster establishment compared with bare-root systems [5,34,35,105], which can translate into more predictable field performance when planting dates fluctuate. The industry implication is that nurseries can select a storage modality depending on whether the primary bottleneck is (i) planting-date uncertainty (favoring storage flexibility) or (ii) establishment risk (favoring rooted systems) [5,91,97,98].
A practical scheduling framework for nurseries involves defining the planting window and transplant type [5,34,35], back-calculating the rooting and conditioning timeline from the target planting date [34,35,44,101], selecting an environmental steering strategy aligned with cultivar response characteristics [61,99,101], and incorporating cold storage as a buffer when planting-date uncertainty is high [57,104], with attention to temperature stability and cultivar-specific cold sensitivity [90,91,97,98].
This “programmed supply” approach is increasingly relevant as nurseries face restrictions on soil fumigation, climate-driven variability, and the rising demand for uniform transplants compatible with protected and off-season production systems [5,12,35,39,68].
6.4. Cold Storage Duration and Its Effects on Subsequent Yield
The relationship between cold storage duration and post-storage yield performance represents a critical practical consideration for commercial nurseries and growers operating with frigo and tray plant systems. The capacity to store strawberry plants at −2 to 0 °C under controlled conditions enables the decoupling of nursery production from fruit production schedules, providing the programmed supply flexibility that is a defining advantage of these systems. However, the evidence consistently indicates that yield potential is inversely related to storage duration beyond certain thresholds. Strawberry plants stored for extended periods show a significant reduction in production capacity relative to plants planted out immediately after harvest [53]. The storage duration negatively affects runner tip viability and subsequent field performance, with progressive decline in transplant success rates and crown vigor as storage extends beyond six months [15]. The underlying mechanisms involve carbohydrate depletion, crown desiccation, and the accumulation of chilling injury in sensitive tissues during prolonged cold storage [96].
Duron and Gomez [88] investigated the effect of temperature and light conditions during the cold storage of strawberry transplants and runner tips, finding that storage conditions, not merely duration, significantly modulate post-storage recovery and yield potential. Plants stored under suboptimal temperature conditions or with inadequate darkness showed accelerated deterioration relative to those held under tightly controlled conditions, indicating that cold storage management practices are as important as storage duration in determining yield outcomes.
The Dutch tray plant system has addressed the storage duration problem more successfully than conventional frigo systems by conditioning plants for extended dormancy through precisely timed nitrogen management, short-day induction protocols, and chilling accumulation under controlled conditions. Tray plants produced in the Netherlands are routinely stored for up to 10–11 months without the severe yield penalties observed in conventionally stored frigo plants, enabling programmed planting schedules throughout the calendar year [14]. This distinction is commercially significant: the premium paid for Dutch tray plants over conventional frigo plants is justified in part by this enhanced storage tolerance and the associated production scheduling flexibility. Growers in the United Kingdom, Germany, and Scandinavia—where tray plants from the Netherlands are widely used—have demonstrated that yields from tray plants planted after extended storage can match or exceed those from fresh bare-root plants planted at optimal timing, provided storage protocols are followed correctly.
Nevertheless, the general principle that fresh plants planted immediately after harvest outperform long-term cold-stored plants under equivalent production conditions remains robust in the literature. This has important implications for China’s strawberry industry, where the absence of adequate cold-chain infrastructure compounds the yield losses associated with informal storage practices, and for emerging production regions in south and southeast Asia where cold-storage development is a priority for industry modernization.
7. Disease Management and Sustainable Approaches
7.1. Soilborne Disease Challenges
Soilborne pathogens remain the primary constraint in strawberry nursery production, with Phytophthora cactorum and Verticillium dahliae causing the most significant economic losses [106,107,108]. P. cactorum causes crown rot characterized by sudden wilting, reddish-brown crown discoloration, and plant death, with optimal infection occurring at 20–25 °C in saturated soils [18]. The pathogen produces motile zoospores that disperse through splash water, irrigation runoff, and contaminated soil, making field nurseries particularly vulnerable during wet conditions. Verticillium dahliae persists in soil as microsclerotia for 10–15 years, colonizing roots and vascular tissue, causing progressive wilting, stunting, and eventual plant death [109].
Additional pathogens complicating nursery production include Colletotrichum species, causing anthracnose crown rot, Fusarium oxysporum f. sp. Fragariae, causing fusarium wilt, Macrophomina phaseolina, causing charcoal rot, and Neopestalotiopsis species [106,107,108]. Viruses transmitted through vegetative propagation, including Strawberry mottle virus (SMoV), Strawberry mild yellow edge virus (SMYEV), and Strawberry crinkle virus (SCV), can reduce plant vigor by 20–40% and necessitate the regular introduction of virus-free nuclear stock [107,110].
7.2. Comparative Disease Incidence Across Propagation Systems
Quantitative comparisons of disease incidence across propagation systems demonstrate the phytosanitary advantages of containerized and soilless production systems. Table 3 summarizes reported disease incidence rates for key pathogens across bare-root field, plug, tray, and elevated bench propagation systems, based on published field studies and reviews in the literature.
Table 3.
Comparative disease incidence for key pathogens across strawberry propagation systems. Data derived from published field trials and reviews in the literature.
The primary mechanism of disease reduction in containerized systems is the elimination of the soil phase, which removes the primary infection pathway for root-invading pathogens, including Phytophthora cactorum, Verticillium dahliae, and Colletotrichum spp. The meristematic tissue behind the root caps represents the principal infection site for Phytophthora. Because the runner tips used in plug production are harvested before root development, they present no accessible infection surface for zoospore entry [2]. Soilless elevated bench systems achieve further pathogen exclusion by physically isolating the root zone from soil, irrigation return water, and splash dispersal—the primary vectors for Neopestalotiopsis spp., which has emerged as a significant nursery pathogen of concern in recent years [107]. In contrast, bare-root field systems remain fully exposed to these infection pathways, particularly in regions where methyl bromide alternatives provide incomplete control [109].
It should be noted that, while soilless systems virtually eliminate soil-borne pathogens, they do not preclude airborne diseases such as Botrytis cinerea and powdery mildew (Podosphaera aphanis). These pathogens can be more prevalent in enclosed greenhouse environments due to elevated humidity, and require dedicated management through ventilation, fungicide programs, and humidity control [111].
7.3. Integrated Disease Management
Sustainable disease management strategies integrate multiple approaches to reduce pathogen pressure. Cultural controls form the foundation, including: site selection avoiding fields with disease history; field sanitation; 3–5-year crop rotations; certified disease-free planting material; disease-free plug tray; removal of infected plants; fungicide treatment; and optimized irrigation management avoiding leaf wetness and soil saturation [3,112,113,114]. Biological control agents, including Trichoderma harzianum, Bacillus subtilis, and Streptomyces species, show promise for suppressing soilborne pathogens, with field efficacy of 40–60% reduction in disease incidence when integrated with other practices [114,115].
Regional disease pressure varies substantially across major production regions, reflecting differences in climate, soil conditions, and production systems. In the southeastern United States, the emergent pathogen Neopestalotiopsis has become the dominant constraint since 2017, with Florida nurseries reporting 40–50% losses in severely affected operations during the 2024–2025 season. This pathogen spreads rapidly through infected transplants from high-latitude nurseries in Atlantic Canada, with wet weather during tip harvest facilitating inoculum development. European strawberry production faces different disease profiles, with Fusarium oxysporum f. sp. fragariae race 2 emerging as a major threat in Spain, causing estimated annual losses of EUR 40–100 million. Asian production systems contend primarily with Colletotrichum and Phytophthora complexes, although elevated bench propagation systems have achieved 85–95% disease suppression compared to conventional field production in Korea and Japan.
The economic impacts of disease extend beyond direct crop losses to include increased production costs, market access restrictions, and supply-chain disruptions. North American strawberry nurseries faced unprecedented transplant shortages in 2024–2025, with major nurseries canceling orders or selling plants “at grower’s risk” due to Neopestalotiopsis contamination. The resulting supply disruptions forced the cancellation of regional strawberry festivals and left commercial producers without adequate planting stock. Certified disease-free plant programs command 15–30% price premiums but offer essential risk mitigation, particularly for operations targeting export markets with strict phytosanitary requirements. Investments in molecular diagnostics (PCR, qPCR) for nursery certification programs add approximately USD 0.15–0.30 per plant to production costs but enable market access and reduce downstream disease losses [107,110].
As illustrated in Figure 1, disease risk varies considerably across propagation systems—from high-risk conventional field bare-root systems to very low-risk tissue culture—and effective disease management requires a prevention-first IPM approach integrating certified planting material, cultural controls, biological agents, PCR-based monitoring, and targeted chemical intervention.
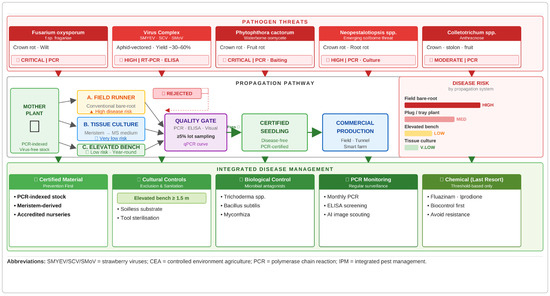
Figure 1.
Integrated overview of strawberry propagation strategies and disease management. Pathogen threats (red, top), propagation pathway (centre), and integrated disease management (green, bottom).
7.4. Innovative System Redesign
System-level redesign offers alternatives to chemical control. Elevated tray field systems that position plug trays 30–50 cm above ground level reduce Phytophthora infection by 40–70% by eliminating splash dispersal of zoospores from contaminated soil [75]. Three-year field trials comparing elevated systems to traditional ground-level production without fungicides demonstrated comparable or superior plant survival rates, with molecular monitoring confirming the absence of P. cactorum in air samples but presence in splash water at ground level [17]. Thermotherapy of runner tips (43–45 °C for 4–6 h) can eliminate certain pathogens without phytotoxicity [116]. Substrate-based production systems using sterilized growing media completely eliminate soilborne pathogen exposure during the propagation phase, although requiring clean water sources and rigorous sanitation protocols [25,116].
8. Emerging Technologies and Future Directions
8.1. F1 Hybrid Seed Propagation
Seed-propagated F1 hybrid cultivars represent a paradigm shift in strawberry propagation, eliminating the virus transmission risks inherent in vegetative propagation [30,63,114,117]. The concept was first commercialized with the Dutch cultivar ‘Karan’ in 1996, followed by Japanese cultivars ‘Chiba F-1 gou’ and ‘Yotsuboshi’, and more recently by global breeding companies [118,119,120,121]. Their development requires establishing multiple inbred lines through 4–6 generations of selfing to achieve sufficient homozygosity, then evaluating F1 combinations for heterosis effects on yield, fruit quality, and environmental adaptation [120].
Successful F1 cultivars, like ‘Yotsuboshi’, demonstrate unique flowering behavior, exhibiting both extra-early flower induction under short days with cool temperatures and everbearing characteristics under long days, enabling year-round production [63,120]. DNA marker systems for seed certification ensure genetic uniformity and prevent off-types [121,122]. Commercial seed production involves the controlled pollination of inbred parents, seed extraction, quality testing, and pelleting for precision sowing. Production costs are initially higher than vegetative propagation; however, the elimination of virus indexing, stock plant maintenance, and runner production infrastructure provides long-term economic advantages [120]. The seeds are more easily shipped, even by sea. Seed plants have higher phytosanitary quality. The seeds require high efficiency in the germination protocols in order to avoid losses [120].
8.2. Artificial Intelligence and Machine Learning Applications
The integration of AI and machine learning is revolutionizing environmental control, quality assessment, and decision-making in strawberry propagation. Computer vision systems using convolutional neural networks and vision transformers achieve 95–99% accuracy in automated assessment of runner tip quality, plant developmental stage, and disease detection, substantially exceeding human grading consistency [123]. These systems enable real-time sorting during propagation workflows, with commercially deployed systems processing 200–300 plants per hour. Deep learning models trained on multispectral imaging data detect physiological stress, nutrient deficiencies, and early disease symptoms 7–14 days before visual symptoms emerge, enabling preemptive intervention [101]. AI-driven climate control systems optimize temperature, humidity, photoperiod, and CO2 enrichment in real time, based on plant developmental stage and environmental conditions, achieving 15–25% improvements in energy efficiency, compared to rule-based control, while maintaining optimal growing conditions [124].
8.3. Vertical Farming and Controlled Environment Propagation
Controlled environment propagation using vertical farming infrastructure enables year-round production independent of seasonal constraints and climate limitations [118]. Multi-tier cultivation systems with LED lighting achieve 5–8× greater space productivity than conventional greenhouse systems, while providing precise environmental control [125]. Spectral optimization using red (600–700 nm) and blue (400–500 nm) LED combinations promotes compact morphology with enhanced root development, reducing production cycles by 20–30% compared to natural light systems [61]. Aeroponic and NFT (nutrient film technique) hydroponic systems enable automated fertigation with real-time adjustment of nutrient composition and pH, minimizing water consumption (90–95% reduction vs. field systems) while optimizing plant nutrition [125]. These systems completely eliminate soilborne disease exposure and reduce pesticide requirements by 80–90%, although capital costs of USD 800–1500 per m2 currently limit adoption to high-value production systems. Integration with renewable energy (solar PV, waste heat recovery) and automated climate control is improving economic viability [118,125].
8.4. Automation and Precision Agriculture
The integration of automation, sensors, and artificial intelligence is transforming propagation efficiency. Machine vision systems enable the automated grading of runner tips and plants based on morphological parameters, achieving 90–95% accuracy compared to manual sorting [101,125,126]. Environmental sensors and AI-driven climate control optimize growing conditions while minimizing energy consumption, with reinforcement learning algorithms outperforming rule-based control strategies by 15–25% in energy efficiency [126,127]. Automated propagation systems combining robotic tip harvesting, automated plugging, and climate-controlled growing environments are being commercialized, although capital costs currently limit adoption to large-scale operations. Deep learning models using multispectral imaging can detect early disease symptoms and predict plant performance, enabling the precision application of interventions [123,126].
9. Critical Analysis and Knowledge Gaps
9.1. Technology Adoption Barriers
Despite documented advantages, adoption rates of advanced propagation technologies remain limited globally outside specialized markets.
Initial capital requirements for plug plant systems (estimated USD 200,000–500,000) and micropropagation facilities (USD 500,000–2,000,000) exceed the financial capacity of the small-to-medium nurseries representing the majority of global production. Risk aversion in conservative agricultural sectors further limits investment, particularly when traditional systems remain functionally adequate in low disease pressure environments.
The successful operation of controlled environment systems requires expertise in climate control, fertigation management, and disease diagnostics often absent in traditional nursery operations. Training programs and technical support infrastructure lag technological availability, creating operational failure risks. Fragmented nursery industries in many regions lack the coordination for collective investments in shared propagation facilities or cooperative micropropagation laboratories that could overcome individual economic barriers.
9.2. Unresolved Scientific Questions
Comprehensive genomic and phenotypic tracking across extended propagation cycles is needed. While photoperiod, temperature, and chilling effects are individually well-studied [88,127], comprehensive models integrating their interactions across diverse cultivar backgrounds and production systems remain elusive. Machine learning approaches analyzing multi-year, multi-location datasets may reveal patterns not evident from controlled studies.
Pathogen evolution and adaptation to substrate-based production systems is understudied. Whether reduced selection pressure in disease-free propagation enables virulence increases upon field introduction requires investigation [2,3,110].
Empirical data on strawberry propagation system performance under projected 2040–2060 climate scenarios are minimal. Field trials manipulating temperature, CO2, and precipitation extremes would inform long-term adaptation planning.
9.3. Trade-Offs and System Selection Criteria
Evidence-based system selection requires explicit trade-off frameworks, as no propagation approach simultaneously optimizes cost, quality, flexibility, sustainability, and risk management. Cost–quality relationships present fundamental choices. Traditional systems minimize per-plant expenses but sacrifice uniformity and disease status, creating clear market segmentation where commodity production favors lower-cost approaches, while premium markets justify advanced system investments [118]. Flexibility carries substantial economic implications, with plug plant systems enabling rapid cultivar switching and schedule adjustments at cost premiums of 30–50%, compared to field nurseries requiring 12–16-week advance planning. Sustainability trade-offs prove particularly complex, as controlled environment systems reduce pesticide use (40–70%) and land requirements but increase energy consumption and infrastructure materials, yielding contradictory lifecycle conclusions depending on environmental impact weighting [118]. Risk distribution further complicates decisions. Advanced technologies reduce disease exposure but concentrate operational risks in equipment reliability and technical expertise, while traditional systems distribute risks across land areas at the cost of weather and pathogen vulnerability.
9.4. Data Gaps and Methodological Limitations
Critical methodological limitations constrain the translation of research findings into practical applications. Economic transparency remains severely limited, as most cost analyses are proprietary and published studies are often 5–10 years outdated, rendering them unsuitable for current decision making. Transparent, region-specific economic datasets employing standardized accounting methods are critically needed to enable evidence-based system selection. Similarly, temporal scope presents significant constraints, as typical study durations of 1–3 years prove insufficient for assessing the cumulative effects of repeated propagation cycles, genetic drift over multiple generations, or disease escape from supposedly pathogen-free systems. Structural biases in research design and reporting further complicate knowledge synthesis. Studies conducted at experimental scales of thousands of plants frequently fail to address whether findings translate to commercial operations managing millions of plants, yet few explicitly examine how system performance metrics change with operational scale. Publication bias compounds these challenges, as successful innovations are systematically overrepresented relative to failed approaches, potentially distorting adoption recommendations by creating an incomplete picture of technological reliability and real-world applicability.
9.5. Integration Challenges
Future competitiveness likely depends on integrated systems combining multiple technologies; however, integration introduces new challenges. Combining micropropagation, automated plug production, AI-driven climate control, and genomic selection requires a level of interdisciplinary expertise rarely concentrated in single organizations.
Precision propagation generates vast datasets (environmental sensors, plant imaging, genomic profiles); however, data integration across systems and platforms remains primitive. Standardized data formats and sharing protocols are needed.
Commercial systems increasingly involve proprietary technologies, cultivars, and protocols. Navigating IP landscapes while maintaining innovation momentum poses challenges for both researchers and industry.
10. Conclusions and Future Perspectives
Strawberry propagation has evolved significantly over the past two decades, driven by regulatory restrictions on soil fumigants, escalating disease pressures, and advances in controlled environment agriculture to diverse high-tech approaches tailored to specific production goals and constraints (Table 4) (Figure 2) [5]. Plug and tray plant systems have emerged as commercially viable alternatives, offering 80–95% disease reduction and 95–98% transplant survival rates despite higher production costs [12]. Environmental control of the photoperiod and temperature enables programmed transplant supply systems that align plant developmental stages with production schedules [9]. Integrated disease management, combining elevated growing systems, substrate-based production, fungicide treatment and biological controls, has reduced pathogen pressure by 40–70% without chemical fumigation [17].
Table 4.
Major advances in strawberry propagation systems over the past 20 years.
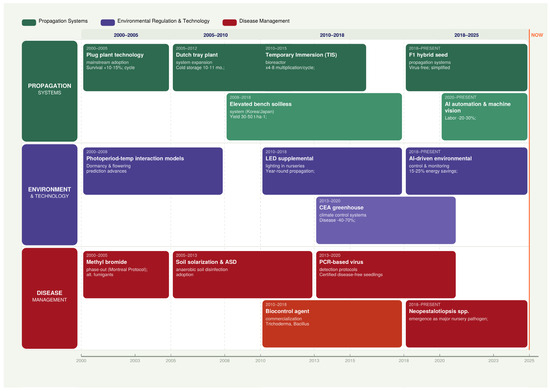
Figure 2.
Timeline of major developments in strawberry propagation methods, environmental regulation, and disease management strategies over the past 20 years (2000–2025). Events are organized into three thematic tracks: propagation systems (green), environmental regulation and technology (purple), and disease management (red). Timeline illustrating key milestones across three thematic tracks from 2000 to 2025. Vertical dashed lines denote major transitional years. Event positions are approximate and reflect the primary period of adoption or publication. MeBr = methyl bromide; CEA = controlled environment agriculture; TIS = temporary immersion system; ASD = anaerobic soil disinfestation; PCR = polymerase chain reaction.
Emerging technologies, including F1 hybrid seed propagation, AI-driven climate control, and machine vision systems, promise further transformation, although widespread adoption remains limited by capital investment barriers and technical knowledge requirements [61]. Micropropagation through temporary immersion bioreactors has achieved commercial-scale production with 50–100% higher multiplication rates while ensuring virus-free status [75]. However, significant challenges persist, including maintaining genetic integrity across propagation cycles, adapting systems to diverse climatic regions, and developing affordable solutions for small-medium operations [80]. The trajectory toward sustainability, quality assurance, and climate resilience is clear, although pathways remain diverse and scale-dependent.
Future research priorities must focus on reducing environmental footprints through energy-efficient systems and LED optimization [125], establishing transparent economic frameworks for evidence-based decision making, and creating modular integrated systems that combine micropropagation, controlled environment propagation, and AI management [125]. The strawberry propagation industry stands at an inflection point where technological capability exceeds current adoption, with success dependent on addressing economic and knowledge access barriers through targeted research, technology transfer programs, and adaptive business models that enable sustainable and climate-resilient production across all operational scales.
Author Contributions
Research conceptualization and analysis, Y.L.; writing—original draft preparation, Y.L.; writing—review and editing, H.Y.; searching for original articles and helping review editing. Y.Y. and B.M.; writing—review supervision, Y.Y.; project administration and funding acquisition. Y.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the Strawberry Science and Technology Plan Project of Global HortResearch Institute in Korea, Grant Agreement No. 2025IA001. The authors thank the Global HortResearch Institute for supporting this research initiative.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- FAOSTAT. Crops and Livestock Products. Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 15 January 2026).
- Maas, J.L. Compendium of Strawberry Diseases, 2nd ed.; APS Press: St. Paul, MN, USA, 1998. [Google Scholar] [CrossRef]
- Martin, F.N. Development of alternative strategies for management of soilborne pathogens currently controlled with methyl bromide. Annu. Rev. Phytopathol. 2003, 41, 325–350. [Google Scholar] [CrossRef] [PubMed]
- Voth, V.; Bringhurst, R.S. Culture and physiological manipulation of California strawberries. Am. Soc. Hortic. Sci. 1990, 25, 889–892. [Google Scholar] [CrossRef]
- Durner, E.F.; Poling, E.B.; Maas, J.L. Recent advances in strawberry plug transplant technology. HortTechnology 2002, 12, 545–550. [Google Scholar] [CrossRef]
- Boxus, P. Review on strawberry mass propagation. Acta Hortic. 1989, 265, 309–320. [Google Scholar] [CrossRef]
- Mukherjee, E.; Gantait, S. Strawberry biotechnology: A review on progress over past 10 years. Sci. Hortic. 2024, 338, 113618. [Google Scholar] [CrossRef]
- Rivero, R.; Remberg, S.F.; Heide, O.M.; Sønsteby, A. Environmental regulation of dormancy, flowering and runnering in two genetically distant everbearing strawberry cultivars. Sci. Hortic. 2021, 290, 110515. [Google Scholar] [CrossRef]
- Heide, O.M.; Sønsteby, A. Interactions of temperature and photoperiod in the control of flowering of latitudinal and altitudinal populations of wild strawberry (Fragaria vesca). Physiol. Plant. 2007, 130, 280–289. [Google Scholar] [CrossRef]
- Al-Madhagi, I. The habit of strawberry flowering is the key for runner propagation, where the photoperiod is the main environmental factor—A review. Adv. Hortic. Sci. 2023, 37, 433–449. [Google Scholar] [CrossRef]
- California Strawberry Commission. Production Guidelines. Available online: https://www.calstrawberry.com (accessed on 10 January 2026).
- Lieten, F. Recent advances in strawberry plug transplant technology. Acta Hortic. 1998, 513, 383–388. [Google Scholar] [CrossRef]
- Bish, E.B.; Cantliffe, D.J.; Chandler, C.K. Temperature conditioning and container size affect early season fruit yield of strawberry plug plants in a winter, annual hill production system. HortScience 2002, 37, 762–764. [Google Scholar] [CrossRef]
- Lieten, P. The strawberry nursery industry in the Netherlands. Acta Hortic. 2014, 1049, 99–106. [Google Scholar] [CrossRef]
- Hokanson, S.C.; Takeda, F.; Enns, J.M.; Black, B.L. Influence of plant storage duration on strawberry runner tip viability and field performance. HortScience 2004, 39, 1596–1600. [Google Scholar] [CrossRef]
- Ellis, M.A.; Converse, R.H.; Williams, R.N.; Williamson, B. Compendium of Raspberry and Blackberry Diseases and Insects, 2nd ed.; APS Press: St. Paul, MN, USA, 1991. [Google Scholar]
- Evenhuis, A.; Bac-Molenaar, J.A.; Pham, K.; Leiss, K.A. Sustainable disease control of Phytophthora cactorum in a strawberry nursery by adapting the growing system. Agronomy 2024, 14, 2809. [Google Scholar] [CrossRef]
- Pscheidt, J.W.; Ocamb, C.M. Pacific Northwest Plant Disease Management Handbook; Oregon State University: Corvallis, OR, USA, 2025; Available online: https://pnwhandbooks.org (accessed on 12 January 2026).
- UNEP. Montreal Protocol on Substances that Deplete the Ozone Layer. United Nations Environment Programme, 2020. Available online: https://www.unep.org/ozonaction/who-we-are/about-montreal-protocol (accessed on 8 January 2026).
- Fennimore, S.A.; Goodhue, R.E. Soil disinfestation with steam: A review of economics, engineering, and soil pest control. Int. J. Fruit Sci. 2016, 16, 71–83. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations (FAO). FAOSTAT Database. Strawberry Production Data, 2022–2023. Available online: https://www.fao.org/faostat/ (accessed on 6 March 2026).
- Yin, S.; Larson, K.D. Strawberry industry in China. Acta Hortic. 2009, 842, 132. [Google Scholar] [CrossRef]
- Tian, R.; Chen, S.; Guo, J.; Liu, K.; Li, Z.; Meng, L.; Zhang, X.; Gao, S.; Wei, H.; Luo, J.; et al. Establishing a virus-free rapid propagation system for strawberry ‘Miaoxiang 7’ through anther culture. Horticulturae 2026, 12, 227. [Google Scholar] [CrossRef]
- Lassen Canyon Nursery. Lassen Canyon Beijing. Available online: https://lassencanyonnursery.com/lassen-canyon-beijing/ (accessed on 24 February 2026).
- Zhang, Y.; Wang, G.; Chang, L.; Dong, J.; Zhong, C.; Wang, L. Current status of strawberry production and research in China. Acta Hortic. 2014, 1049, 19–28. [Google Scholar] [CrossRef]
- Torres-Quezada, E.A.; Zotarelli, L.; Whitaker, V.M.; Darnell, R.L.; Morgan, K.; Santos, B.M. Production techniques for strawberry plugs in West-Central Florida. HortTechnology 2020, 30, 238–247. [Google Scholar] [CrossRef]
- Martínez-Treceño, A.; Chomé, P.; Becerril, M.; Soria, C.; Medina, J.J.; López-Aranda, J.M. The strawberry nursery industry in Spain: An update. Acta Hortic. 2009, 842, 691–694. [Google Scholar] [CrossRef]
- López-Aranda, J.M.; Urbina, S.; Castellá, G.; Estrada, J.F.; Cotero, M.A. Large plot demonstrations for methyl bromide alternatives on strawberry in Baja California, Mexico. Acta Hortic. 2012, 926, 415–422. [Google Scholar] [CrossRef]
- Lee, W.S. Production of ever-bearing strawberry and production technology in Korea. Acta Hortic. 2014, 1049, 561–564. [Google Scholar] [CrossRef]
- Lee, H.S.; Choi, J.M.; Kim, T.I.; Kim, H.S.; Lee, I.H. Influence of bicarbonate concentrations in nutrient solution on the growth, occurrence of daughter plants and nutrient uptake in vegetative propagation of ‘Seolhyang’ strawberry. Korean J. Hortic. Sci. Technol. 2014, 32, 149–156. [Google Scholar] [CrossRef]
- Yoshida, Y.; Goto, T.; Hirai, M.; Masuda, M.; Nishimoto, K. Effects of the duration of chilling and forcing temperatures on flowering and runnering in everbearing strawberry ‘Elan’. Environ. Control Biol. 2012, 50, 29–36. [Google Scholar]
- Cocco, C.; Faedi, W.; Magnani, S.; Maltoni, M.L.; Stagno, F.; Quacquarelli, I.; Ferré, A.S.; Baruzzi, G. Effects of plug plants and bare-root plants on strawberry field performance, fruit quality traits and health-promoting compounds. J. Berry Res. 2020, 10, 189–203. [Google Scholar] [CrossRef]
- Morresi, V.; Capocasa, F.; Balducci, F.; Diamanti, J.; Mezzetti, B. Nursery propagation systems for high-quality strawberry (Fragaria × ananassa Duch.) plug plant production from micropropagated, soilless-grown mother plants. Horticulturae 2025, 11, 888. [Google Scholar] [CrossRef]
- Palombini, M.C.; Palencia, P.; Pavão, J.M.S.J.; Chiomento, J.L.T. Efficiency of strawberry cultivation under the effect of different types of plants in a soilless system in the high-altitude regions of Southern Brazil. Agronomy 2023, 13, 2179. [Google Scholar] [CrossRef]
- Poling, E.B.; Parker, K. Plug production of strawberry transplants. Adv. Strawb. Prod. 1990, 9, 37–39. [Google Scholar]
- Rowley, D.; Black, B.; Drost, D. Strawberry Plug Plant Production. USU Extension Publication Horticulture/High Tunnels/2010-02pr. 2010. Available online: https://digitalcommons.usu.edu/psc_facpub/511/ (accessed on 11 January 2026).
- Pritts, M.P.; Handley, D.T. Strawberry Production Guide for the Northeast, Midwest, and Eastern Canada; Northeast Regional Agricultural Engineering Service: Ithaca, NY, USA, 1998. [Google Scholar]
- Takeda, F.; Hokanson, S.C. Strawberry fruit and plug plant production in the greenhouse. Acta Hortic. 2003, 626, 133–139. [Google Scholar] [CrossRef]
- Hicklenton, P.R.; Reekie, J.Y. The effect of brief exposures to high temperatures on transplant performance of strawberry plug plants. Can. J. Plant Sci. 2000, 80, 613–618. [Google Scholar]
- Xu, X. Optimizing Environmental Parameters for Precision Indoor Propagation of Day-Neutral Strawberry. Master’s Thesis, North Carolina State University, Raleigh, NC, USA, 2019. [Google Scholar]
- Martin, R.R.; Tzanetakis, I.E. Characterization and recent advances in detection of strawberry viruses. Plant Dis. 2006, 90, 384–396. [Google Scholar] [CrossRef]
- Jun, H.J.; Jeon, E.H.; Kang, S.I.; Bae, G.H. Optimal nutrient solution strength for Korean strawberry cultivar ‘Daewang’ during seedling period. Korean J. Hortic. Sci. Technol. 2014, 32, 812–818. [Google Scholar] [CrossRef]
- Lee, H.C.; Kim, H.S.; Jang, W.S.; Nam, M.H.; Lee, I.H.; Lee, M.H.; Kim, T.I.; Lee, E.M.; Choi, J.M. Changes in chemical characteristics of drainage solution as influenced by years of root media use in successive hydroponic cultivations of ‘Seolhyang’ strawberries. Hortic. Sci. Technol. 2019, 37, 375–383. [Google Scholar] [CrossRef]
- Kim, D.Y.; Chae, W.B.; Kwak, J.H.; Park, S.H.; Cheong, S.R.; Choi, J.M.; Yoon, M.K. Effect of timing of nutrient starvation during transplant production on the growth of runner plants and yield of strawberry ‘Seolhyang’. Prot. Horti. Plant Fact. 2013, 22, 421–426. [Google Scholar] [CrossRef]
- Bish, E.B.; Cantliffe, D.J.; Hochmuth, G.J.; Chandler, C.K. Development of containerized strawberry transplants for Florida’s winter production system. Acta Hortic. 1997, 439, 461–468. [Google Scholar] [CrossRef]
- Albregts, E.E.; Howard, C.M. Strawberry production in Florida. In Florida Agricultural Experiment Station Bulletin 841; University of Florida: Gainesville, FL, USA, 1984. [Google Scholar]
- Hochmuth, G.; Cantliffe, D.J.; Chandler, C.; Stanley, C.; Bish, E.; Waldo, E.; Legard, D.; Duval, J. Containerized strawberry transplants reduce establishment-period water use and enhance early growth and flowering compared with bare-root plants. HortTechnology 2006, 16, 46–54. [Google Scholar] [CrossRef]
- Lieten, P. Advances in strawberry substrate culture during the last twenty years in the Netherlands and Belgium. Int. J. Fruit Sci. 2013, 13, 84–90. [Google Scholar] [CrossRef]
- Lieten, P. Strawberry production in Central Europe. Int. J. Fruit Sci. 2006, 5, 91–105. [Google Scholar] [CrossRef]
- Duralija, B.; Mesic, A.; Jemric, T. Cold Stored Tray Plants in Open Field Cultivation of Strawberries. Agric. Conspec. Sci. 2006, 71, 167–170. [Google Scholar]
- Lieten, P. The effect of nutrition prior to and during flower differentiation on phyllody and plant performance of short-day strawberry Elsanta. Acta Hortic. 2002, 567, 345–348. [Google Scholar] [CrossRef]
- Chaudhary, D.R.; Rasmussen, K.; Nyhuus, T.E.; Krüger, E.; Gislerød, H.R.; Torre, S. Effects of temperature and photoperiod on the flower potential in everbearing strawberry as evaluated by meristem dissection. Horticulturae 2021, 7, 484. [Google Scholar] [CrossRef]
- Kim, S.K.; Jeong, M.S.; Park, S.W.; Kim, M.J.; Na, H.Y.; Chun, C. Improvement of runner plant production by increasing photosynthetic photon flux during strawberry transplant propagation in a closed transplant production system. Korean J. Hortic. Sci. Technol. 2010, 28, 535–539. [Google Scholar]
- Lieten, F.; Kinet, J.M.; Bernier, G. Effect of prolonged cold storage on the production capacity of strawberry plants. Scientia Hortic. 1995, 60, 213–219. [Google Scholar] [CrossRef]
- Bradford, E.; Hancock, J.F.; Warner, R.M. Interactions of temperature and photoperiod determine expression of repeat flowering in strawberry. J. Am. Soc. Hortic. Sci. 2010, 135, 102–107. [Google Scholar] [CrossRef]
- Guttridge, C.G. Fragaria × ananassa. In CRC Handbook of Flowering; Halevy, A.H., Ed.; CRC Press: Boca Raton, FL, USA, 1985; Volume 3, pp. 16–33. [Google Scholar]
- Hidaka, K.; Dan, K.; Imamura, H.; Miyoshi, Y.; Takayama, T.; Sameshima, K.; Kitano, M.; Okimura, M. Effect of supplemental lighting from different light sources on growth and yield of strawberry. Environ. Control Biol. 2017, 55, 81–87. [Google Scholar] [CrossRef]
- Ruan, J.; Lee, Y.H.; Yeoung, Y.R. Flowering and fruiting of day-neutral and ever-bearing strawberry cultivars in high-elevation for summer and autumn fruit production in Korea. Hortic. Environ. Biotechnol. 2013, 54, 109–120. [Google Scholar] [CrossRef]
- Ruan, J.; Lee, Y.H.; Yeoung, Y.R.; Larson, K.D. Influence of short-day treatment on autumn fruit production of June-bearing strawberry cultivars. Hortic. Environ. Biotechnol. 2011, 52, 259–264. [Google Scholar] [CrossRef]
- Wei, H.; Liu, T.; Jeong, B.R. An optimal combination of the propagation medium and fogging duration enhances the survival, rooting and early growth of strawberry daughter plants. Agronomy 2020, 10, 557. [Google Scholar] [CrossRef]
- Lieten, P.; Gallace, N. Nitrogen fertilization in the nursery: Effect on strawberry tray plant yield and performance. Acta Hortic. 2021, 1309, 54. [Google Scholar] [CrossRef]
- Park, S.W.; Kim, S.K.; Kwack, Y.; Chun, C. Simulation of the number of strawberry transplants produced by an autotrophic transplant production method in a plant factory with artificial lighting. Horticulturae 2020, 6, 63. [Google Scholar] [CrossRef]
- Mori, T.; Kohori, J.; Kitamura, H.; Inokuchi, T.; Kato, I.; Sone, K.; Ishikawa, M.; Maeda, F.; Fukami, M.; Isobe, S.; et al. Development of F1 hybrid strawberry of seed propagation type named ‘Yotsuboshi’ by collaborative breeding among institutes. Hortic. Res. 2015, 14, 409–418. [Google Scholar] [CrossRef][Green Version]
- Martin, R.R.; Spiegel, S.; Jelkmann, W. Strawberry Mild Yellow Edge Disease. In Compendium of Strawberry Diseases, 2nd ed.; Maas, J.L., Ed.; APS Press: St. Paul, MN, USA, 1998; pp. 65–66. [Google Scholar]
- Anuradha, S.K.; Sehrawat, A.K.; Poonia, S.K.; Sandhya, B. Production of strawberry plant by in vitro propagation. Res. Crops 2016, 17, 545–549. [Google Scholar] [CrossRef]
- Kajic, V.; Cvjetkovic, B. The use of meristem tip culture to eliminate viruses from strawberry plants. Acta Hortic. 1989, 265, 595–598. [Google Scholar] [CrossRef]
- Badal, M.N.U.; Shoyeb, M.; Sarkar, M.A.R.; Rahman, S.; Rahman, S.M. Clonal propagation of strawberry (Fragaria × ananassa Duch.) through in vitro runner tip culture through incorporation of growth hormones. Biotechnol. J. Int. 2019, 22, 1–9. [Google Scholar] [CrossRef]
- Etienne, H.; Berthouly, M. Temporary immersion systems in plant micropropagation. Plant Cell Tissue Organ Cult. 2002, 69, 215–231. [Google Scholar] [CrossRef]
- Debnath, S.C. Bioreactors and molecular analysis in berry crop micropropagation—A review. Can. J. Plant Sci. 2011, 91, 147–157. [Google Scholar] [CrossRef]
- Hazarika, B.N. Morpho-physiological disorders in in vitro culture of plants. Scientia Hortic. 2006, 108, 105–120. [Google Scholar] [CrossRef]
- Neri, J.C.; Meléndez-Mori, J.B.; Tejada-Alvarado, J.J.; Vilca-Valqui, N.C.; Huaman-Huaman, E.; Oliva, M.; Goñas, M. An optimized protocol for micropropagation and acclimatization of strawberry (Fragaria × ananassa Duch.) variety ‘Aroma’. Agronomy 2022, 12, 968. [Google Scholar] [CrossRef]
- Quiroz, K.A.; Berríos, M.; Carrasco, B.; Retamales, J.B.; Caligari, P.D.; García-Gonzáles, R. Meristem culture and subsequent micropropagation of Chilean strawberry (Fragaria chiloensis (L.) Duch.). Biol. Res. 2017, 50, 20. [Google Scholar] [CrossRef]
- Capocasa, F.; Balducci, F.; Marcellini, M.; Bernardini, D.; Navacchi, O.; Mezzetti, B. Comparing nursery behavior, field plant yield and fruit quality of in vitro and in vivo propagated strawberry mother plants. Plant Cell Tissue Organ Cult. 2019, 136, 65–74. [Google Scholar] [CrossRef]
- Swartz, H.J.; Galletta, G.J.; Zimmerman, R.H. Field performance and phenotypic stability of tissue culture-propagated strawberries. J. Am. Soc. Hortic. Sci. 1981, 106, 667–673. [Google Scholar] [CrossRef]
- Karhu, S.; Hakala, K. Field performance of Fragaria × ananassa plants regenerated by tissue culture. Acta Hortic. 2000, 530, 197–202. [Google Scholar]
- Grout, B.W.W.; Aston, M.J. Transplanting of cauliflower plants regenerated from meristem culture. I. Water loss and water transfer related to changes in leaf wax and to xylem regeneration. Hortic. Res. 1978, 17, 1–7. [Google Scholar]
- Pospisilova, J.; Ticha, I.; Kadlecek, P.; Haisel, D.; Plzakova, S. Acclimatization of micropropagated plants to ex vitro conditions. Biol. Plant. 1999, 42, 481–497. [Google Scholar] [CrossRef]
- Nehra, N.S.; Kartha, K.K.; Stushnoff, C.; Giles, K.L. Effect of in vitro propagation methods on field performance of two strawberry cultivars. Euphytica 1994, 76, 107–115. [Google Scholar] [CrossRef]
- Li, Y.; Hu, J.; Wei, H.; Jeong, B.R. A long-day photoperiod and 6-benzyladenine promote runner formation through upregulation of soluble sugar content in strawberry. Int. J. Mol. Sci. 2020, 21, 4917. [Google Scholar] [CrossRef]
- Hytönen, T.; Elomaa, P.; Moritz, T.; Junttila, O. Gibberellin mediates daylength-controlled differentiation of vegetative meristems in strawberry (Fragaria × ananassa Duch.). BMC Plant Biol. 2009, 9, 18. [Google Scholar] [CrossRef]
- Gordon, T.R.; Kirkpatrick, S.C.; Hansen, J.; Shaw, D.V. Response of strawberry genotypes to inoculation with isolates of Verticillium dahliae differing in host origin. Plant Pathol. 2006, 55, 766–769. [Google Scholar] [CrossRef]
- Mouhu, K.; Kurokura, T.; Koskela, E.A.; Albert, V.A.; Elomaa, P.; Hytönen, T. The Fragaria vesca homolog of suppressor of overexpression of constans1 represses flowering and promotes vegetative growth. Plant Cell 2013, 25, 3296–3310. [Google Scholar] [CrossRef] [PubMed]
- Kurokura, T.; Mimida, N.; Battey, N.H.; Hytönen, T. The regulation of seasonal flowering in the Rosaceae. J. Exp. Bot. 2013, 64, 4131–4141. [Google Scholar] [CrossRef]
- Verheul, M.J.; Sønsteby, A.; Grimstad, S.O. Interaction of photoperiod, temperature, duration of short-day treatment and plant age on flowering of Fragaria × ananassa Duch. cv. Korona. Sci. Hortic. 2006, 107, 164–170. [Google Scholar] [CrossRef]
- Lieten, P.; Gallace, N. The effect of nitrogen on fruit shape and production in short day ‘Opera’ strawberry trayplants. Acta Hortic. 2017, 1156, 39. [Google Scholar] [CrossRef]
- Tanino, K.K.; Wang, J. Modeling chilling requirement and diurnal temperature differences on flowering and yield performance of strawberry crown production. HortScience 2008, 43, 2060–2065. [Google Scholar] [CrossRef]
- Li, Y.; Xiao, J.; Guo, G.; Jeong, B.R. Transplant pre-chilling induces earlier flowering and fruiting for forcing-cultured June-bearing strawberries. Scientia Hortic. 2021, 110371. [Google Scholar] [CrossRef]
- Duron, L.; Gomez, C. Evaluating the effect of temperature and light during cold storage of strawberry transplants and runner tips. Hortic. Environ. Biotechnol. 2025, 10, 1–13. [Google Scholar] [CrossRef]
- Kim, D.Y.; Kim, T.I.; Kim, W.S.; Kang, Y.I.; Yun, H.K.; Choi, J.M.; Yoon, M.K. Changes in growth and yield of strawberry (cv. Maehyang and Seolhyang) in response to defoliation during nursery period. Prot. Hortic. Plant Fact. 2011, 20, 283–289. [Google Scholar]
- Naznin, T.; Lefsrud, M.; Gravel, V.; Hao, X. Using different ratios of red and blue LEDs to improve the growth of strawberry plants. Acta Hortic. 2016, 1134, 125–130. [Google Scholar] [CrossRef]
- Braun, J.W.; Kender, W.J. Correlative bud inhibition and growth habit of the strawberry as influenced by application of gibberellic acid, cytokinin, and chilling during short daylength. J. Am. Soc. Hortic. Sci. 1985, 110, 28–34. [Google Scholar] [CrossRef]
- Nestby, R.; Lieten, F.; Pivot, D.; Raynal Lacroix, C.; Tagliavini, M. Influence of mineral nutrients on strawberry fruit quality and their accumulation in plant organs: A review. Int. J. Fruit Sci. 2006, 5, 139–156. [Google Scholar] [CrossRef]
- Palonen, P.; Buszard, D. Current state of cold hardiness research on fruit crops. Can. J. Plant Sci. 1997, 77, 399–420. [Google Scholar] [CrossRef]
- Savini, G.; Neri, D.; Zucconi, F.; Sugiyama, N. Strawberry growth and flowering: An architectural model. Int. J. Fruit Sci. 2005, 5, 29–50. [Google Scholar] [CrossRef]
- Durner, E.F.; Barden, J.A.; Himelrick, D.G.; Poling, E.B. Photoperiod and temperature effects on flower and runner development in day-neutral, Junebearing, and everbearing strawberries. J. Am. Soc. Hortic. Sci. 1984, 109, 396–400. [Google Scholar] [CrossRef]
- Park, S.W.; Kwack, Y.; Chun, C. Growth and propagation rate of strawberry transplants produced in a plant factory with artificial lighting as affected by separation time from stock plants. Hortic. Environ. Biotechnol. 2018, 59, 199–204. [Google Scholar] [CrossRef]
- Jung, D.H.; Kim, H.S.; Jhin, C.; Kim, H.J.; Park, S.H. Time-serial analysis of deep neural network models for prediction of climatic conditions inside a greenhouse. Comput. Electron. Agric. 2020, 173, 105402. [Google Scholar] [CrossRef]
- Choi, J.M.; Lee, H.S. Influence of Ca containing fertilizers on the growth of mother and daughter plants, and physiological disorders in propagation of ‘Seolhyang’ strawberry through plastic bag cultivation. Korean J. Hortic. Sci. Technol. 2013, 31, 50–55. [Google Scholar] [CrossRef][Green Version]
- Sønsteby, A.; Heide, O.M. Long-day control of flowering in everbearing strawberries. J. Hortic. Sci. Biotechnol. 2008, 83, 360–366. [Google Scholar] [CrossRef]
- Wilhelm, S.; Paulus, A.O. How soil fumigation benefits the California strawberry industry. Plant Dis. 1980, 64, 264–270. [Google Scholar]
- Choi, J.M.; Latigui, A.; Lee, C.W. Visual symptom and tissue nutrient contents in dry matter and petiole sap for diagnostic criteria of phosphorus nutrition for ‘Seolhyang’ strawberry cultivation. Hortic. Environ. Biotechnol. 2013, 54, 52–57. [Google Scholar] [CrossRef]
- Yoshida, Y. Strawberry production in Japan: History and progress in production technology and cultivar development. Int. J. Fruit Sci. 2013, 13, 103–113. [Google Scholar] [CrossRef]
- Fang, X.; You, M.P.; Barbetti, M.J. Reduced severity and impact of Fusarium wilt on strawberry by manipulation of soil pH, soil organic amendments and crop rotation. Eur. J. Plant Pathol. 2012, 134, 619–629. [Google Scholar] [CrossRef]
- Isobe, S.N.; Hirakawa, H.; Sato, S.; Maeda, F.; Ishikawa, M.; Mori, T.; Yamamoto, Y.; Shirasawa, K.; Kimura, M.; Fukami, M.; et al. Construction of an integrated high density simple sequence repeat linkage map in cultivated strawberry (Fragaria × ananassa) and its applicability. DNA Res. 2013, 20, 79–92. [Google Scholar] [CrossRef]
- Jelkmann, W.; Martin, R.R. Crinkle disease (Strawberry crinkle virus). In Compendium of Strawberry Diseases, 2nd ed.; Maas, J.L., Ed.; APS Press: St. Paul, MN, 1998; pp. 62–64. [Google Scholar]
- Xiao, C.L.; Chandler, C.K.; Price, J.F.; Duval, J.R.; Mertely, J.C.; Legard, D.E. Comparison of epidemics of botrytis fruit rot and powdery mildew of strawberry in large plastic tunnel and field production systems. Plant Dis. 2001, 85, 901–909. [Google Scholar] [CrossRef]
- Gaire, S.; Muzhinji, N.; Louws, F.J.; Adhikari, T.B. Neopestalotiopsis spp., an invasive fungal pathogen, is a major threat to strawberry production: The current status, challenges, and future directions. Front. Plant Sci. 2026, 17, 1725321. [Google Scholar] [CrossRef]
- Cosseboom, S.D.; Schoeneberg, A.; Johnson, C.S.; Peres, N.A.; Hu, M. Resistance to azoxystrobin and thiophanate-methyl is widespread in Colletotrichum spp. Isolates from the Mid-Atlantic Strawberry Fields. Plant Dis. 2020, 105, 2202–2208. [Google Scholar] [CrossRef]
- Sundelin, T.; Schiller, M.; Lubeck, M.; Jensen, D.F.; Paaske, K.; Petersen, B.D. First report of anthracnose fruit rot caused by Colletotrichum acutatum on strawberry in Denmark. Plant Dis. 2007, 89, 432. [Google Scholar] [CrossRef]
- Ávila-Hernández, J.G.; León-Ramírez, C.G.; Abraham-Juárez, M.R.; Tlapal-Bolaños, B.; Olalde-Portugal, V.; Délano-Frier, J.P.; Martínez-Antonio, A.; Aguilar-Zárate, P. Neopestalotiopsis spp.: A threat to strawberry production and management. Horticulturae 2025, 11, 288. [Google Scholar] [CrossRef]
- Bentvelsen, G.C.M.; Bouw, E.; Veldhuyzen van Zanten, J.E. Breeding strawberry (Fragaria × ananassa Duch.) from seed. Acta Hortic. 1997, 439, 149–154. [Google Scholar] [CrossRef]
- Fravel, D.R. Commercialization and implementation of biocontrol. Annu. Rev. Phytopathol. 2005, 43, 337–359. [Google Scholar] [CrossRef]
- Domínguez, P.; Miranda, L.; Soria, C.; de los Santos, B.; Chamorro, M.; Romero, F.; Daugovish, O.; Medina, J.J. Soil biosolarization for sustainable strawberry production. Agron. Sustain. Dev. 2014, 34, 821–829. [Google Scholar] [CrossRef]
- Fukao, S.; Ishikawa, M.; Fumi, M.; Toshikatsu, O. Methods of Cultivating the June-Bearing Seed Propagation Strawberry ‘Chiba F-1 gou’. CAFRC Res. Bull. 2012, 4, 1–6. [Google Scholar]
- Oliveira, M.S.; Wang, N.Y.; Peres, N.A. Multilocus phylogenetic analyses of Colletotrichum gloeosporioides species complex causing crown rot on strawberry in Florida. Phytopathology 2022, 112, 898–906. [Google Scholar] [CrossRef] [PubMed]
- Rho, I.R.; Woo, J.G.; Jeong, H.J.; Jeon, H.Y.; Lee, C.H. Characteristics of F1 hybrid and inbred lines in octaploid strawberry (Fragaria × ananassa Duch.). Plant Breed. 2012, 131, 550–554. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, X.; Ji, L.; Daggubati, S.; Shen, K. Deep learning for strawberry runner detection integrating ground and aerial imaging. Smart Agric. Technol. 2025, 12, 101290. [Google Scholar] [CrossRef]
- Lorch-Schierning, E.; Perera, A.; Baghalian, K. Using vertical farming systems for propagating everbearing strawberry: Physiological and technological feasibility assessment. J. Berry Res. 2024, 14, 31–44. [Google Scholar] [CrossRef]
- Whitaker, V.M.; Knapp, S.J.; Hardigan, M.A.; Edger, P.P.; Slovin, J.P.; Bassil, N.V.; Hytönen, T.; Mackenzie, K.K.; Lee, S.; Jung, S.; et al. A roadmap for research in octoploid strawberry. Hortic. Res. 2020, 7, 33. [Google Scholar] [CrossRef]
- Choi, K.Y.; Choi, E.Y.; Kim, I.S.; Lee, Y.B. Improving water and fertilizer use efficiency during the production of strawberry in coir substrate hydroponics using a FDR sensor-automated irrigation system. Hortic. Environ. Biotechnol. 2016, 57, 431–439. [Google Scholar] [CrossRef]
- Tehranifar, A.; Battey, N.H.; Cekic, C.; Taylor, S.; Shrives, K.J.; Hadley, P.; Greenland, A.J.; Darby, J.; Wilkinson, M.J. Characterization of the Fragaria chiloensis (L.) Duch. genome using variety-specific microsatellite primers. Euphytica 1998, 84, 245–251. [Google Scholar]
- Afonso, M.; Fonteijn, H.; Fiorentin, F.S.; Lensink, D.; Mooij, M.; Faber, N.; Polder, G.; Wehrens, R. Tomato fruit detection and counting in greenhouses using deep learning. Front. Plant Sci. 2020, 11, 571299. [Google Scholar] [CrossRef]
- Zhiwei, T.; Ma, W.; Yang, Q.; Duan, F. Application status and challenges of machine vision in plant factory—A review. Inf. Process. Agric. 2021, 9, 195–211. [Google Scholar] [CrossRef]
- He, Z.; Liu, Z.; Zhou, Z.; Karkee, M.; Zhang, Q. Improving picking efficiency under occlusion: Design, development, and field evaluation of an innovative robotic strawberry harvester. Comput. Electron. Agric. 2025, 237, 110684. [Google Scholar] [CrossRef]
- Park, Y.; Sethi, R.; Temnyk, S. Growth, flowering, and fruit production of strawberry ‘Albion’ in response to photoperiod and photosynthetic photon flux density of sole-source lighting. Plants 2023, 12, 731. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Mochizuki, T.; Mori, T.; Kohori, J.; Kitamura, H.; Inokuchi, T.; Kato, I.; Sone, K.; Ishikawa, M.; Maeda, F.; Fukami, M.; et al. ‘Yotsuboshi’, a new F1 hybrid strawberry of seed propagation type for year-round production. Acta Hortic. 2017, 1156, 53–58. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.