Abstract
Different cyanobacterial species have been shown to be a valuable source of biologically active compounds with immunomodulatory activity. To date, little is known about members of the genus Tolypothrix (Cyanophyceae). Therefore, the present study focuses on five Tolypothrix strains (T. tenuis PACC 5497, T. tenuis PACC 8648, T. distorta SAG 1482-2, T. distorta CCALA 194, Tolypothrix sp. PACC 5501) that were not previously evaluated for specific morphological characteristics and immunomodulatory potential toward human immune cells. Cyanobacterial cultures were studied by light and transmission electron microscopy (TEM). Peripheral blood leukocytes were isolated from patients with inflammatory conditions and treated ex vivo with Tolypothrix non-polar extract fractions. Following treatment, the cells were analyzed by flow cytometry, and cytokine concentrations in culture supernatants were quantified by ELISA. Light microscopy observations showed that the cultures established from four of the strains have morphological features that correspond to T. tenuis Kützing (1843) ex Bornet et Flahault 1887 and T. distorta Kützing (1843) ex Bornet et Flahault. TEM analyses indicated parietal arrangement of cellular thylakoids in all strains, but T. distorta CCALA194 and Tolypothrix sp. PACC 5501 also displayed fascicular thylakoid arrangement. Immunophenotypic analyses revealed significantly increased proportions of T, NK, and B lymphocytes in leukocyte cultures treated with Tolypothrix extracts compared to the untreated controls. The concentrations of proinflammatory cytokines were lower in the culture medium of treated cells, while levels of the anti-inflammatory cytokine interleukin-10 remained stable, except in cultures treated with T. distorta SAG 1482-2 extract. The present study provides the detailed morphological characteristics of five strains of the genus Tolypothrix and indicate that non-polar extract fractions derived from the strains exert immunomodulatory effects on human leukocytes.
1. Introduction
Cyanobacteriota (Cyanobacteria) are considered to be among the earliest photosynthetic organisms on Earth, which induced the oxygenation of water and atmosphere on the planet approximately 2.4 billion years ago [1]. Presently, representatives of this group constitute a major component of the phytoplankton in numerous freshwater and marine ecosystems [2] and thus, they hold a main position as the primary producers on Earth. To date, approximately 860 genera and more than 5700 species have been identified [3]. A significant number of cyanobacterial species have been shown to produce biologically active metabolites with various beneficial properties. Based on structural characteristics, biologically active compounds derived from cyanoprokaryotes are classified as pigments, lipids, polysaccharides, nucleosides, proteins and peptides, lipopeptides, phenolic compounds, alkaloids and terpenoids [4,5,6]. Generally, they exert diverse biological activities. A summarized instance includes different types of pigments (phycobiliproteins, chlorophylls, carotenoids, mycosporine amino acids (MAA)) that have demonstrated antioxidant, antitumor, antigenotoxic, antimutagenic, cardio protective and antiobesity activities; lipopeptides with antimalarial, anticancer, antiproliferative and/or antifungal properties; cyanobacterial nucleosides that have exerted antitumor, antifungal, antiviral activities; exopolysaccharides that have shown antioxidant, anti-inflammatory, or immunomodulatory properties, while sulfated polysaccharides have been shown to exhibit antiviral activity; phenolic compounds that have demonstrated antibacterial, antifungal, antitumor, and anti-inflammatory activities; lipids, mainly different types of saturated and unsaturated fatty acids, have exerted immunomodulatory and antitumor activities; and peptides with antimicrobial, antifungal and antitumor activity have been described, as well as alkaloids and terpenoids with anti-inflammatory, antitumor, antibacterial, antioxidant and UV-protective effects [4,5,6]. Recent advances in the methods for isolation of cyanobacterial metabolites, combined with molecular modeling and docking analyses, provide new opportunities for a more detailed investigation of cyanobacterial compounds and the development of products with potential applications in pharmacology, food industry, cosmetics, and biotechnology [7].
Currently, despite extensive research on biologically active substances produced by cyanobacteria, only 10–20% of the identified cyanobacterial metabolites have been comprehensively characterized with respect to their chemical structures and effects on living organisms [8]. The majority of detected compounds are commonly determined by preliminary bioassays or inferred through genomic and metabolomic analyses without complete elucidation of activity and structure. This considerable knowledge gap highlights the immense, largely untapped potential of cyanobacteria as a source of novel natural products with various beneficial effects. Continued efforts in compound isolation and bioactivity screening are therefore essential to fully explore the functional roles of extracts derived from different cyanobacterial species.
The genus Tolypothrix includes heterocytous filamentous cyanobacteria belonging to the order Nostocales. A significant number of representatives of this genus have not been examined in detail with regard to their potential use as sources of biologically active natural compounds [6]. Tolypothrix species are found in stagnant, slow-flowing or subaerial freshwater environments [9]. They typically form fasciculated colonies that appear as floating tufts or are attached to aquatic plants, rocks, and sediments and reproduce via hormogonia. The filaments are heteropolar, characterized by false branching and presence of a mucilaginous sheath, which contribute to their ecological adaptability. Heterocysts are present singly or in pairs, located intercalary and often at the base of false branches [10]. Akinetes are less commonly observed. Tolypothrix species are known to participate in nitrogen fixation and play an important role in the primary productivity and nutrient cycling of freshwater ecosystems [2]. Despite their ecological importance, systematic studies of biological activity focusing on Tolypothrix extracts remain limited compared with those on other genera within Nostocales, such as Nostoc, Anabaena, and Calothrix [4]. Several Tolypothrix strains were shown to be capable of producing secondary metabolites with antibacterial, antioxidant, antitumor properties; however, only a few of these bioactive compounds (for example, hassallidins and tjipanazoles) have been structurally characterized [6,11,12]. Certain species of the genus Tolypothrix have been shown to produce compounds with anti-inflammatory properties, which makes them interesting for the development of new agents including nutraceutics for the treatment of inflammatory diseases. An example of such a substance is tolypodiol isolated from T. nodosa [13]. Other examples are toyocamycin and tubercidin—nucleoside chemicals with anti-fungal properties isolated from T. tenuis [14]. Additionally, a small number of biosynthetic gene clusters determining the production of phosphonate, ectoine, phenazine, β-lactone and tetrapyrrole macrocycles were detected [4,15]. A screening study reported the antibacterial and radical-scavenging (antioxidant) activity of extracts derived from environmental isolates of Tolypothrix spp. [16]. All these findings highlight Tolypothrix species as a promising source of novel natural products with potential pharmacological applications. Still, the biological activity of compounds produced by Tolypothrix species and strains have not been evaluated which leaves much of the biosynthetic potential of this genus unexplored.
Our research group has recently shown that non-polar fractions of extracts derived from five Tolypothrix strains demonstrate important biological activities in vitro, including antitumor potential and other properties [17]. Experiments with a mouse macrophage model demonstrated that all extracts significantly decreased the production of proinflammatory cytokines by lipopolysaccharide-stimulated RAW264.7 cells. These findings indicated the anti-inflammatory activity of natural products obtained from the studied Tolypothrix strains. Chemical analysis showed the presence of 26 different fatty acids distributed differently in the individual strains [18]. Therefore, to support the pharmacological importance of these cyanobacterial samples, we proceeded our research by evaluating their effects on human immunocompetent cells and also by performing morphological analyses for strain characterization and determination of potential strain-specific features. The present study reports the morphometric characteristics and specific thylakoid arrangement in the cells of five Tolypothrix strains (Tolypothrix sp. PACC 5501, T. tenuis PACC 5497, T. tenuis PACC 8648, T. distorta SAG 1482-2, T. distorta CCALA 194), which were not previously described. Our research demonstrates for the first time the effects of non-polar Tolypothrix extract fractions on human peripheral blood leukocytes, based on analyses of the proportion of the main immune cell populations and the production of proinflammatory and anti-inflammatory cytokines.
2. Materials and Methods
2.1. Selected Cyanobacterial Strains
Five Tolypothrix strains (Nostocales, Cyanobacteriota) were included in the present study (Table 1). Four of them were identified to species level as Tolypothrix tenuis Kützing ex Bornet & Flahault 1886 (PACC 5497 and PACC 8648) and Tolypothrix distorta Kützing ex Bornet & Flahault 1886 (CCALA 194 and SAG 1482-2). One strain (Tolypothrix sp. PACC 5501) was not determined to species level. The strains were obtained from the following culture collections: PACC—Plovdiv Algal Culture Collection (Plovdiv, Bulgaria); CCALA—Culture Collection of Autotrophic Organisms (Třeboň, Czech Republic); and SAG—Sammlung von Algenkulturen (Göttingen, Germany).

Table 1.
Origin, maintenance and cultivation of the selected cyanobacterial strains.
2.2. Cultivation of Cyanobacteria and Preparation of Extracts
Z medium (Zehnder Medium for Cyanobacteria) [19] with pH modification in the alkaline range (pH 8.5–9.0) and ES (“Erddekokt + Salze”) medium (Basal medium) [20] were used for the cultivation and long-term maintenance of the cyanobacterial strains. Media requirements and the origin of the studied Tolypothrix strains are indicated in Table 1.
The cyanobacteria were inoculated in 25 cm2 culture flasks (TPP, Trasadingen, Switzerland)—1 mL of the delivered actively growing cultures was added to 9 mL culture medium. The expanded cultures were subsequently transferred to 75 cm2 flasks (3 mL inoculum and 27 mL culture medium). The Tolypothrix strains were grown in liquid alkaline Z-medium under sterile conditions. Cultivation was performed in a growth chamber equipped with a light source providing constant intensity and stable spectral composition, with automatic photoperiod control. A 12 h:12 h (light:dark) photoperiod was maintained using 40 W cool-white fluorescent lamps with an intensity of 10 µmol m−2 s−1.
The strains were cultured for six weeks under sterile conditions to accumulate the cyanobacterial biomass necessary for the study (morphological analysis, transmission electron microscopy (TEM), and extract preparation), to culture stage corresponding to the stationary phase under the applied conditions. Samples for classical morphological and TEM analyses were taken from actively growing cultures during the third week after inoculation.
To obtain cyanobacterial biomass, expanded cultures were harvested after the cultivation period, transferred to 50 mL centrifuge tubes and centrifuged for 15 min at 4000 rpm. Then, the cell pellet was frozen and lyophilized. Extracts of each Tolypothrix strain were derived from a total of 500 mg lyophilized cyanobacterial biomass. The material was mixed with 3 mL of methanol and vortexed for 1 min. This was followed by extraction for 20 min in an ultrasonic bath (Branson 5510R-DTH, Wilmington, NC, USA) at 40 °C, 25 kHz frequency. During this step, vortexing was performed every 3–5 min. After that, 6 mL of chloroform were added to the resulting suspension and mixed, which was followed by shaking for 20 min. Then, 3 mL of deionized water were added and the mixture was vortexed for 1 min. The extracts were centrifuged at 4000 rpm for 20 min. The methanol/chloroform (non-polar) and water/methanol (polar) fractions were separated and filtered through a 0.20 μm hydrophobic polytetrafluoroethylene (PTFE) membrane filter (Merck KGaA, Darmstadt, Germany). The organic solvents were removed from the samples by vacuum evaporation at 37 °C using Savant SpeedVac Concentrator (SAVANT Instruments Inc., Farmingdale, NY, USA). The dried extracts were dissolved in DMSO/water (1:1) and solutions with a final concentration of 5 mg/mL (w/v) were obtained. To achieve a final DMSO concentration of <1% for in vitro assays, working solutions were prepared with sterile Dulbecco’s modified Eagle medium (DMEM) or Dulbecco’s phosphate-buffered saline (DPBS) (both purchased from Gibco®, Life Technologies™, Paisley, Scotland, UK). The fatty acid composition of the obtained extract fractions was determined by gas chromatography in a previous study [17] and is discussed in the following sections in relation to the results reported in this article.
2.3. Microscopy Analyses
2.3.1. Morphometric Evaluations
Morphological analyses of the Tolypothrix strains were implemented with a Magnum-T microscope (Medline Scientific Ltd., Chalgrove, UK) and a high-resolution Si-3000 XLiCap digital camera (Medline Scientific Ltd., Chalgrove, UK). Pictures of all analyzed cultures were obtained at a magnification of 100–400×. The following valuable phenotypic features were monitored during the exponential growth phase of the strains: thallus (macroscopic view: color, growth), filaments (width), sheaths (width, color, structure), trichomes (shape, constrictions), cells (shape, dimensions), terminal cells (shape), heterocysts (arrangement, shape, size). Length (L) and width (W) measurements were performed on a minimum of 50 cells (measurement unit—µm).
2.3.2. Analyses of Thylakoid Arrangement
Cyanobacterial cultures were harvested and centrifuged at 4000 rpm for 5 min. The pelleted filaments were washed with 0.1 M cacodylate buffer and after that, they were fixed with 4% glutaraldehyde solution in 0.1 M cacodylate buffer at pH 7.2 for 4 h at 4 °C. The samples were further processed as previously described [21] and analyzed with a high-resolution transmission electron microscope HR STEM JEOL JEM 2100 (JEOL Ltd., Tokyo, Japan) operating at 200 kV, equipped with a CCD camera GATAN Orius 832 SC1000 (Gatan GmbH, Munich, Germany).
2.4. Isolation of Leukocytes and Cell Culture Conditions
In the present study, peripheral blood was obtained from 3 patients with inflammatory conditions (5 mL sample per patient; C-reactive protein/CRP/concentration in blood plasma > 10 mg/L). Prior to the sample collection, a written consent form has been signed by all participants. The experiment was performed in accordance with the Declaration of Helsinki and approved by the Local Ethical Committee at the University of Plovdiv “Paisii Hilendarski”, Bulgaria (protocol No. 7 from 10 June 2024).
Blood samples were collected from the cubital vein of the patients into BD Vacutainer® K2 EDTA tubes (Becton, Dickinson and Company, Oakville, ON, Canada). Then, the samples were centrifuged at 1000 rpm for 20 min at room temperature and the plasma was discarded. Red blood cells were lysed using 0.84% NH4Cl solution and the samples were washed twice with sterile DPBS (Gibco®, Life Technologies™, Paisley, Scotland, UK). The isolated leukocytes were centrifuged at 1000 rpm for 10 min and then, the cells were resuspended in DMEM supplemented with 10% heat-inactivated fetal bovine serum and a stabilized solution of antibiotic and antifungal agent (all purchased from Sigma Aldrich Chemie GmbH, Steinheim, Germany). This medium is denoted as supplemented DMEM. The isolated leukocytes (1 × 106 cells/mL) were plated in 12-well plates (TPP, Trasadingen, Switzerland) and treated for 48 h with 100 μg/mL of cyanobacterial extract, which was added to the culture medium (1 mL/well). Cells cultured for the same period (48 h) in supplemented DMEM without addition of the extract sample served as a negative non-treated control. Cells incubated for 48 h with 1 μg/mL phytohemagglutinin-L (PHA-L) (Sigma-Aldrich, Saint Louis, MO, USA) were used as a positive control. Leukocytes were cultured at 37 °C, 5% CO2, 95% ambient air, in a high-humidity incubator.
2.5. Immunophenotyping
At the end of the 48 h incubation period, treated and control cells were harvested by centrifugation at 1000 rpm for 10 min and were resuspended in FACS buffer (DPBS supplemented with 5% fetal calf serum and 0.05% NaN3). The cells were stained for 20 min at room temperature, with fluorochrome-conjugated anti-human antibodies grouped into the following 3 panels: (I) CD3 PE-Dazzle™ 594, CD4 FITC, CD8 PE, CD25 PE-Cy5 and CD279 PE-Cy7; (II) HLADR/DP FITC, CD19 PE, CD80 PE-Dazzle™ 594, CD11b PE-Cy5, and CD152 PE-Cy7; and III) CD3 PE-Dazzle™ 594, CD56 FITC, CD80 PE-Cy7 and CD16 PE (all from BioLegend®, San Diego, CA, USA). After that, the cells were washed twice and resuspended in 300 μL FACS buffer. The cells were analyzed by flow cytometry using a Cytomics FC500 instrument (Beckman Coulter Inc., Life Sciences, Indianapolis, IN, USA). All samples were analyzed in duplicate.
2.6. Analyses of Cytokine Levels
The concentration of interleukin (IL)-2, IL-6, IL-10, interferon (IFN)-γ, and tumor-necrosis factor (TNF)-α in the culture medium of non-treated cells and cells treated with Tolypothrix extracts after 48 h incubation was assayed with LEGEND MAX™ Kits Human IL-2, IL-6 IL-10, IFN-γ and TNF-α ELISA kits (BioLegend Inc., San Diego, CA, USA), according to the instructions of the manufacturer. All samples were analyzed in duplicate.
2.7. Statistics
Statistically significant differences between the test samples and the controls were determined by analysis of variance (ANOVA) and Fisher’s PLSD test for the ELISA and flow cytometry data, using StatView software version 5.0 (SAS Institute Inc., Cary NC, USA). In addition, to compare the effects between the extracts tested, we applied one-way analysis of variance (ANOVA), followed by Tukey’s post hoc test (IBM SPSS Statistics, version 28.0. Armonk, NY, USA). Prior to ANOVA, we checked assumptions for the normality of residuals by Shapiro–Wilk test and the homogeneity of variances by Levene’s test. All analyses used significance level α = 0.05. When ANOVA was significant, we conducted post hoc pairwise comparisons with Tukey’s Honest Significant Difference (Tukey HSD test). Adjusted p-values, confidence intervals, and mean differences were recorded for all group pairs. Group letters (a, b, c, d, e) were assigned for homogeneous subsets (groups were not significantly different), according to each test.
3. Results
3.1. Morphological Characteristics of the Tolypothrix Strains
The results from the biometric measurements of cells from the five Tolypothrix strains are summarized in Table 2. They indicate that T. distorta CCALA 194, T. distorta SAG 1482-2, T. tenuis PACC 5497 and T. tenuis PACC 8648 strains have typical corresponding species cell dimensions, concordant to the literature data [9].
Table 2.
Morphometric data for the selected Tolypothrix strains.
3.1.1. Morphological Description of the Strains
The essential morphological characteristics of taxonomic relevance for the studied strains are summarized in Table 3
. Only features that were commonly used for the identification of Tolypothrix species are included in the table. More variable characteristics, such as thallus coloration, macroscopic appearance, and growth pattern in culture flasks, are presented descriptively in this section. These features showed variation among strains under the applied cultivation conditions.
Table 3.
Morphological characteristics relevant for strain identification.
Tolypothrix distorta CCALA 194: Thallus olive-green, in the form of a gelatinous mass on the surface or film on the walls of culture flasks. Filaments were falsely branched and 12–14 μm wide (Figure 1A; Table 3). The sheath was firm, up to ±2 μm wide, and colorless to yellowish. The trichomes were cylindrical, distinctly constricted at cross-walls, and not widened or narrowed towards the ends. Cells are short and cylindrical, usually ½ shorter than their width (2.0–6.0 × 7.0–10.0 μm) and blue-green; end cells ± were rounded to spherical. The heterocysts were solitary, less frequently in pairs, usually at the base of branches, and spherical (8–10 μm) or short and cylindrical (6.0–10 × 8–10.0 μm).

Figure 1.
Morphology of studied Tolypothrix strains observed by light microscopy. (A). T. distorta CCALA 194; (B). T. distorta SAG 1482-2; (C). T. tenuis PACC 5497; (D). T. tenuis PACC 8648; and (E). Tolypothrix sp. PACC 5501. All pictures were taken at 400× magnification. Scale bars 50 µm.
Tolypothrix distorta SAG 1482-2: Thallus dark blue-green, in fascicles or in a cushion-like formation on the bottom and surface of the culture flasks. The filaments were straight or flexuous, rich falsely branched, and 10–12 μm wide (Figure 1B; Table 3). The sheaths were firm, thin, and colorless. The trichomes were cylindrical, it was unclear if they were constricted at cross-walls, and they were not widened or narrowed towards the ends. The cells were blue-green or olive-green, cylindrical, and usually shorter than their width (2.0–7.0 × 6.0–9.0 μm). Terminal cells ± were rounded to spherical. Heterocysts were usually at the base of the branches, single, less frequently in pairs, spherical or cylindrical (5.0–9.0 × 7–9.0 μm).
Tolypothrix tenuis PACC 5497: Thallus blackish-green to brownish, thin, and in the form of fascicles on the walls of the cultivation vessel and aerophilic above the nutrient medium. The filaments were up to 1 cm long, ±straight or slightly curved, up to 12 μm wide, with false branching. The sheaths were distinct, thin or thickened, colorless, and sometimes mucilaginous. The trichomes were cylindrical and slightly constricted at cross-walls (Figure 1C; Table 3). The cells were olive-green, cylindrical, ± isodiametric or slightly shorter than their width, 2.0–8.0 × 5.0–7.0 μm; terminal cells were rounded and conical, and longer than vegetative cells. The heterocysts were not observed.
Tolypothrix tenuis PACC 8648: Thallus olive-green, in the form of fascicles throughout the culture flasks. The filaments were long, ±straight or slightly curved, and up to 14 μm wide, with false branching. The sheaths were firm, distinctly colorless or yellowish. The trichomes were bright blue-green, cylindrical, and constricted at cross-walls (Figure 1D; Table 3). The cells were cylindrical, ±isodiametric or slightly shorter than their width (2.0–8.0 × 5.0–12.0 μm); terminal cells were rounded to almost spherical. The heterocysts were solitary and spherical (9.0 μm), and were observed very rarely.
Tholypothrix sp. PACC 5501: Thallus blackish-green, in the form of fascicles along the bottom and walls of the culture vessel and aerophilic above the liquid medium. The filaments were brownish-green, ±straight, and up to 8 μm wide, with false branching. The sheaths were thin, often mucilaginous to amorphous, and colorless. The trichomes were yellowish green, cylindrical, and clearly constricted at cross-walls (Figure 1E; Table 3). The cells were barrel-shaped to cylindrical, ±isodiametric, and slightly shorter or longer than the width, 2.0–7.0 × 4.0–7.0 μm; terminal cells were rounded and conical. The heterocysts were rare, solitary or in a row of up to three, almost spherical (6.0 μm).
3.1.2. TEM Analysis of Thylakoid Arrangement
Representative pictures for the three species (T. distorta, T. tenuis, Tolypothrix sp.) are shown on Figure 2. Cells from all five strains demonstrated the parietal (peripheral) arrangement of thylakoids (Figure 2A,D–F). A second type of thylakoid arrangement pattern was observed for T. distorta and Tolypothrix sp.—fasciculated thylakoids were irregularly grouped and located in the central part of the cellular cytoplasm (Figure 2B,C).
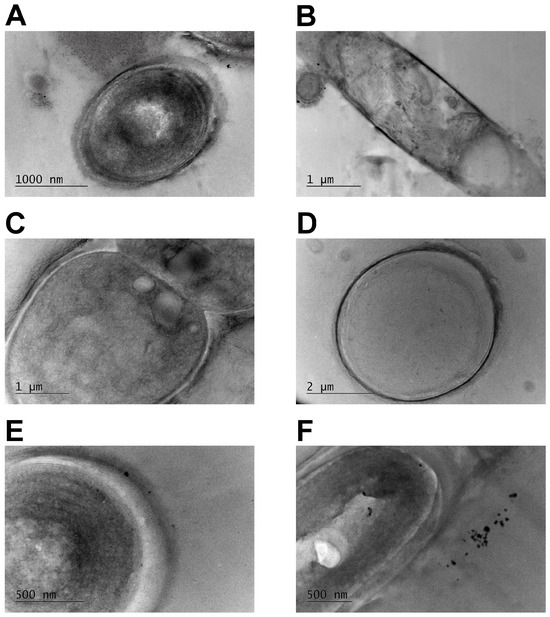
Figure 2.
Thylakoid arrangement in cells of studied Tolypothrix strains. (A,B) T. distorta CCALA 194; (C,D). Tolypothrix sp. 5501; and (E,F). T. tenuis PACC 8648.
3.2. Non-Polar Extract Fractions Obtained from Tolypothrix Strains Exert Immunomodulatory Properties
Our previous studies have demonstrated the anti-inflammatory activity of extracts derived from the five Tolypothrix strains. This was defined in vitro, using LPS-stimulated RAW264.7 mouse macrophages. After 24 h treatment of the cells with increasing concentrations of non-polar extract fractions (50, 100, and 200 µg/mL), we determined inhibitory effects on the production of the pro-inflammatory cytokines IL-6 and TNF-α [17]. The effects were consistent for all strains and doses that were tested. These findings stimulated additional studies aiming to further define the immunomodulatory potential of the non-polar fractions of cyanobacterial extracts, particularly on human immunocompetent cells. Thus, the main purpose of the present study was to evaluate the immunomodulatory activity of Tolypothrix sp. PACC 5501, T. tenuis PACC 5497, T. tenuis PACC 8648, T. distorta SAG 1482-2, and T. distorta CCALA 194 extracts on human leukocytes, in combination with a morphological description of the strains. Non-polar extract fractions were selected for the evaluations, based on previous results from in vitro assays with mouse macrophages and human cells lines that indicated significant biological activity, mainly for the non-polar extract fractions compared to the polar ones [17].
To determine the effects of extracts derived from different Tolypothrix strains on the levels of the main immune cell populations in the peripheral blood, we designed staining panels that allowed us to identify the subsets of T cells, B cells, antigen-presenting cells (APCs) and NK cells. To assess the T cell subsets in response to non-polar cyanobacterial extracts, we analyzed the expression of CD3, CD4, CD8, CD25, CD152 and CD279 markers by human leukocytes derived from patients with active inflammatory conditions. Immunophenotypic analyses of leukocytes treated for 48 h with Tolypothrix extracts demonstrated significantly higher levels of T lymphocytes compared to the control non-treated leukocytes (Figure 3A,B). The strongest effect was observed after treatment with Tolypothrix sp. 5501 and was more pronounced for the subpopulation of CD4+ T-lymphocytes. Overall, the elevated percentages of T cells after treatment with the Tolypothrix extract samples did not correlate with increased expression of the activation marker CD25 (Figure 3C), which indicates that the observed effect was not due to uncontrolled T-cell activation, and the treatment with cyanobacterial extracts is associated with an immune regulatory effect. Furthermore, there was a significant increase in the percentage of CD3+CD152+ T cells (Figure 3D). The strongest effect was observed under the influence of extracts from strains Tolypothrix sp. 5501, T. distorta SAG 1482-2 and strain T. tenuis PACC 8648. Increased levels of CD3+CD279+ T cells were also observed (Figure 3E). This result was evident for all cultures treated with Tolypothrix extracts, but not for the non-treated peripheral blood leukocytes.
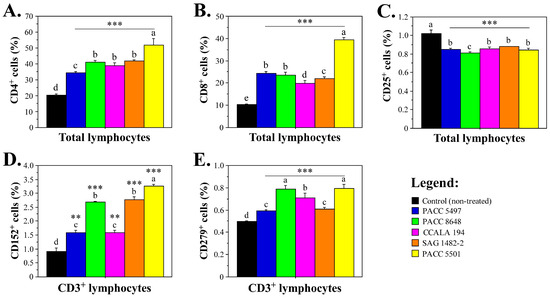
Figure 3.
Proportions of T cell subpopulations after ex vivo treatment of peripheral blood leukocytes with non-polar Tolypothrix extracts. (A). Percentages of CD4+ lymphocytes; (B). levels of CD8+ lymphocytes; (C). CD25-expressing lymphocyte levels; (D). CD152+ T cells; and (E). CD279+ T-lymphocytes. Asterisks (** p < 0.01, *** p < 0.001) denote significant differences between extracts versus control. The letters above the bars indicate statistical groupings—samples sharing the same letter are not significantly different by Tukey’s post hoc test.
Treatment with Tolypothrix non-polar extract fractions led to significantly increased levels of CD19+ B-lymphocytes, CD56+/CD16+ NK cells and CD80+ antigen-presenting cell subpopulations (Figure 4). The detected effects were more pronounced for the strains T. distorta CCALA 194 and Tolypothrix sp. PACC 5501, with regard to the levels of CD80+ and CD56+/CD16+ NK cells.
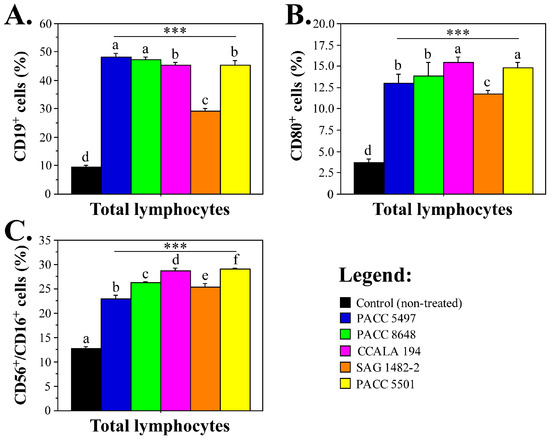
Figure 4.
Levels of B, NK and antigen-presenting cell populations after treatment with nonpolar Tolypothrix extracts. (A).—levels of B lymphocyte populations; (B).—CD80+ lymphocytes; and (C).—percentages of NK cells. Asterisks (*** p < 0.001) denote significant differences between extracts versus control. The letters above the bars indicate statistical groupings—samples sharing the same letter are not significantly different by Tukey’s post hoc test.
3.3. Tolypothrix Non-Polar Extract Fractions Inhibit Proinflammatory Cytokine Production by Human Leukocytes
Treatment with non-polar extract fractions of Tolypothrix stains influenced the levels of some pro-inflammatory cytokines, which are responsible for cell-mediated immunity and the initial activation of the immune system (Figure 5A–C,E). Significantly decreased IL-2 levels (Figure 5A) supported the hypothesis for an immunomodulatory effect of the extract treatment that does not induce excessive activation of T-cell populations.
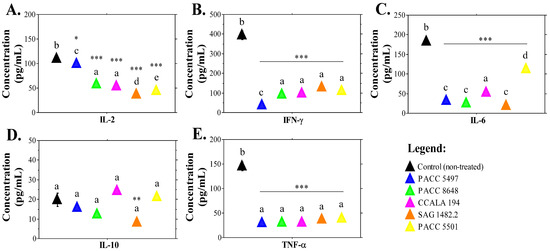
Figure 5.
Cytokine concentrations in the leukocyte culture medium after treatment with nonpolar Tolypothrix extract fractions. IL-2, IL-6, IL-10, IFN-γ and TNF-α levels in supernatants were analyzed by ELISA. (A).—concentration of IL-2; (B).—IFN-γ content in the cell culture medium of non-treated and treated with Tolypothrix extract leukocytes; (C).—IL-6 concentration; (D). levels of IL-10 in the cell culture medium after treatment with cyanobacterial extracts; and (E). TNF-α concentration in the culture medium of non-treated and treated with Tolypothrix extracts leukocytes. Asterisks (* p < 0.05, ** p < 0.01, *** p < 0.001) denote significant differences between extracts versus control. Letters above bars indicate statistical groupings—samples sharing the same letter are not significantly different by Tukey’s post hoc test.
When analyzing the data related to the modulation of cytokine levels, it should be taken into consideration that blood donors are patients with active inflammation. Therefore, the non-treated (control) cells showed high levels of pro-inflammatory cytokines. Figure 5 demonstrates that extracts from Tolypothrix reduced the production of pro-inflammatory cytokines (IL-6, IFN-γ and TNF-α) by human leukocytes. For IL-10, which is an anti-inflammatory cytokine, treatment with extracts from two strains (PACC 5501, CCALA 194) increased the concentration of the cytokine in the cell culture medium, suggesting anti-inflammatory potential. This was confirmed by the cytokine analyses that showed a lower content of IL-2, IL-6, TNF-α and IFN-γ in the culture supernatant of leukocytes treated with Tolypothrix extracts compared to the non-treated controls (Figure 5A–C,E). Only treatment with T. distorta SAG 1482-2 resulted in significantly reduced production of IL-10 (Figure 5D). Nonetheless, this sample, as well as the four other Tolypothrix samples, significantly reduced the production of three types of pro-inflammatory cytokines, which indicates their immune regulatory activity.
4. Discussion
Certain members of the genus Tolypothrix have been shown to produce compounds with anti-inflammatory properties, which makes them interesting for the development of new drugs providing effective treatment of inflammatory conditions, as well as nutraceuticals [5,12,22]. However, little is known about the biological activities, particularly about the immunomodulatory properties of substances derived from other Tolypothrix species. Thus, we have focused our research on five strains belonging to this genus—T. tenuis PACC 5497, T. tenuis PACC 8648, T. distorta CCALA 194, T. distorta SAG 1482-2 and Tolypothrix sp. PACC 5501. They were chosen based on recently reported data for their antitumor, antioxidant and anti-inflammatory potential [17]. The present study started with a morphological and ultrastructural assessment of material from expanded cultures of the selected strains aiming at detailed strain-specific description, which was not previously reported. These evaluations are an important step in strain validation and provide information for specific morphological and ultrastructural characteristics. The observed morphological variability among the studied strains may also reflect adaptive responses to environmental conditions; however, in the present study, these features are primarily considered in the context of strain identification and characterization.
Microscopy observations and morphometric measurements confirmed that the strains PACC 5497 and PACC 8648 belong to the species T. tenuis, while SAG 1482-2 and CCALA 194 belong to the species T. distorta. Morphometric results are consistent with the previously reported data for the cellular length and width of T. distorta (2.5–8.5 µm length, 6–12 µm width) and T. tenuis (2.3–6 µm length, 3–12 µm width) and correspond to the morphological description of both species [9].
In cultures of T. distorta, we observed the following variability of important morphological characters: thalli olive- to dark blue-green, and in fascicles or in a cushion-like formation on the bottom and surface of the culture flasks. The filaments were falsely branched and 10–14 μm wide. The sheaths were firm, up to ±2 μm wide, and colorless to yellowish. The trichomes were cylindrical, distinctly or unclearly constricted at cross-walls, and not widened or narrowed towards ends. The cells were blue-green or olive-green, cylindrical, and usually shorter (up to 1/2) than their width (2.0–7.0 × 6.0–10.0 μm); terminal cells ± were rounded to spherical. The heterocysts were usually at the base of the branches, single, less frequently in pairs, and spherical (8–10 μm) or short and cylindrical (5.0–10 × 7–10.0 μm). TEM analysis: parietal (peripheral) and fascicular arrangement of thylakoids [23].
Variations in thallus coloration likely reflect differences in pigment composition and physiological state, rather than stable taxonomic traits, as reported for other heterocytous cyanobacteria.
In cultures of T. tenuis, the thalli was olive-green to brownish, in the form of fascicles on the walls of the cultivation vessel and aerophilic above the liquid medium. The filaments were up to 1 cm long, ±straight or slightly curved, and up to 12 μm wide, with false branches. The sheaths were distinct, thin or thickened, colorless or yellowish, and sometimes mucilaginous. The trichomes were cylindrical and slightly constricted at cross-wall. The cells were bright blue-green to olive-green, cylindrical, ± isodiametric or slightly shorter than wide (2.0–8.0 × 5.0–7.0 μm). The terminal cells were rounded-conical or rounded to almost spherical. The heterocysts were not or were very rarely observed, and single spherical (9.0 μm). TEM analysis: parietal (peripheral) arrangement of thylakoids [23].
The strain Tolypothrix sp. PACC 5501 was not identified to the species level and, therefore, could not be included in the species-level comparison. We identified the following valuable morphological features: the filaments were brownish-green, ±straight, and up to 8 μm wide, with false branching. The sheaths were thin, often mucilaginous, and colorless. The trichomes were yellowish green, cylindrical, and clearly constricted at cross-walls. The cells were barrel-shaped to cylindrical, ±isodiametric, and slightly shorter or longer than the width (2.0–7.0 × 4.0–7.0 μm). The terminal cells were rounded and conical. Heterocysts were rare, solitary or in a row of up to three, and almost spherical (6.0 μm). TEM analysis: parietal (peripheral) and fascicular arrangement of thylakoids [23].
Although no direct relationship can be established within the scope of this study, the structural variability among strains may be associated with differences in metabolic profiles, which is consistent with the observed slight variation in the immunomodulatory activity.
Our studies proceeded with analyses of the potential immunomodulatory properties of extracts obtained from the selected Tolypothrix strains. A generally accepted strategy for evaluating the potential of cyanobacteria as producers of biologically active substances involves the study of “crude” extracts and fractions. Particularly, the non-polar extract fractions were used, due to their superior biological effects in terms of antitumor activity compared to polar fractions [17]. In vitro and/or in vivo test systems are used to evaluate different bioactive properties, including antitumor, antioxidant, anti-inflammatory and immunoregulatory activity. Specifically, anti-inflammatory effects are determined based on reduction in the levels of pro-inflammatory cytokines such as TNF-α, IL-1β and IL-6. Per example, low levels of TNF-α, IL-1β, and IL-6 in the culture medium were reported for LPS-stimulated RAW264.7 macrophages and mouse bone marrow-derived macrophages after treatment with lipid fractions from Nostoc commune var. spheroides and Spirulina platensis [24]. Reduced levels of TNF-α and NF-κB expression were determined in LPS-stimulated THP-1 cells after treatment with the two digalactosyldiacylglycerols isolated from Nodularia harveyana [25]. Similar to this effect, other authors reported the anti-inflammatory effect of nonpolar compounds, such as 7(E)-9-keto-octadec-7-enoic acid (C18 acid), isolated from cyanobacteria [26]. The compound suppressed iNOS expression and nitric oxide (NO) production in LPS-stimulated RAW264.7 cells—a frequently used in vitro model for studying inflammatory processes. Other cyanobacterial species have also been shown to be a source of bioactive compounds with anti-inflammatory activity. An example is Aphanothece halophytica, which produce mycosporine-2-glycine, a rare mycosporine-like amino acid that induced anti-inflammatory effects in LPS-stimulated RAW 264.7 macrophages [27]. Different genera of aquatic and terrestrial cyanobacteria produce scytonemin—a pigment that exerts anti-inflammatory activity in vitro by the down-regulation of NF-κB activity and inhibits skin inflammation by blocking the expression of inflammatory mediators [28]. Several research groups have reported the anti-inflammatory activity of cyanobacterial extracts or lipidic compounds, amino acids and peptides, polysaccharides and pigments based on the observed reduction/downregulation of proinflammatory cytokine production in vitro, using cell lines like RAW264.7, THP-1, human epidermal keratinocytes and others, or by demonstrating a reduction in edema, skin inflammation, lesions and itching using rodent models [29].
In our previous studies, non-polar fractions of Tolypothrix extracts significantly reduced the production of proinflammatory cytokines IL-6 and TNF-α by RAW264.7 mouse macrophages [17]. These findings paved the way for further evaluations of potential immunomodulatory effects of the Tolypothrix strains. The present research supported the data for anti-inflammatory activity and showed the important effects of the samples on human immune cells. Our results demonstrated significantly elevated levels of CD4+ T cells, CD8+ T cells, CD80+ lymphocytes, B cells, and NK cell populations in leukocyte cultures treated for 48 h with Tolypothrix non-polar extract fractions. The uniform enhancement of the percentages of the main leukocyte populations could be a sign for overactivation in response to the ex vivo cultivation in a medium containing cyanobacterial extracts. However, the percentage of CD25+ lymphocytes was significantly decreased in the Tolypothrix-treated cell cultures. CD25 is a subunit of the IL-2 receptor—an activation marker [18] expressed by immune and some non-immune cell types—but high levels of CD25 are specific for activated T lymphocytes and regulatory T cells [30]. The reduction in CD25+ lymphocytes together with significantly reduced IL-2 concentration in the cell culture medium following treatment with Tolypothrix extracts indicates that the test samples did not induce strong activation of the lymphocytes, which could eventually lead to immune reactions with negative effects. In support of this suggestion, we observed increased percentages of CD152+ and CD279+ T cells. CD152 (cytotoxic T-lymphocyte-associated antigen-4/CTLA-4/) is a main immune checkpoint regulator of T cell homeostasis and self-tolerance [31]. Increased transcriptomic expression of CD152 has been shown to be a predictive marker for a positive outcome for patients undergoing immunotherapy [32]. Therefore, the elevated numbers of CD152+ T cells after treatment with Tolypothrix extracts could be considered an indicator of immunomodulatory activity of the cyanobacterial samples. Similar indication has been provided by the detected enhanced proportion of T cells expressing CD279+—a programmed death 1 (PD-1) receptor that has been shown to exert immunoregulatory roles in T cell activation and tolerance [33]. Our findings for decreased levels of CD25+ lymphocytes and increased levels of CD152+ and CD279+ T cells, together with the decreased production of cytokines with proinflammatory activity (IL-6, IFN-γ and TNF-α) provide evidence for potential immunoregulatory activity of the selected Tolypothrix strains. Combined with the data for the elevated levels of the main immune cell populations, increased T cell expression of immune checkpoint regulators (CTLA-4 and PD-1) and sustained production of the anti-inflammatory cytokine IL-10, which was not reduced by treatment with four of the cyanobacterial extracts, our results prove potential to maintain anti-inflammatory responses and exclude the assumption for the induction of long-term immunosuppression with eventual severe consequences like recurrent opportunistic infections, de novo malignancies, renal failure, hypertension and autoimmunity [34]. The only extract that reduced IL-10 production was T. distorta SAG 1482.2. A possible reason for this different result could be the lack of stearidonic acid in this sample, which was determined in our previous studies [17]. Stearidonic acid (SDA) inhibits the action of cyclooxogenase-2 (COX-2) and arachidonate 5-lypoxigenase (ALOX5), which leads to a reduction in prostaglandins (e.g., PGE2) and leukotrienes that are important mediators of inflammatory responses [35]. In addition, SDA and its metabolites can inhibit nuclear factor kappa B (NF-kB), which reduces the expression of inflammatory cytokines (such as IL-6 and TNF-α) [36]. Previously, we identified long-chain fatty acids (ω-3 linolenic acid, ω-3 docosahexaenoic acid, and ω-9 oleic acid) as dominant nonpolar metabolites of Tolypothrix extracts [17]. These specific structures are known to exert anti-inflammatory effects. Oleic acid modulates membrane fluidity and NF-κB signaling, while ALA (alpha-linolenic acid) and DHA (docosahexaenoic acid) have been reported to downregulate pro-inflammatory genes and oxidative stress [17]. Thus, the fatty-acid profile aligns with our data for the cytokine levels. Strains that are richer in these unsaturated acids showed stronger inhibition of IL-6 and TNF-α.
Both our previous results [17] and those observed in this study indicate that Tolypothrix nonpolar extracts interrupt the inflammatory signaling cascade at multiple points. In LPS-stimulated RAW264.7 macrophages, extracts caused a clear dose-dependent scavenge of free radicals and decreased cytokine production (IL-6 and TNF-α). In parallel, ex vivo human peripheral leukocytes treated with the same extracts showed significantly lower IL-6, TNF-α, IFN-γ and IL-2, which was consistent with an overall suppression of pro-inflammatory signaling. This broad cytokine suppression suggests an upstream block of the NF-κB (Nuclear factor kappa B) and MAPK (Mitogen-activated protein kinase) pathways. Nonpolar compounds can inhibit NF-κB activation (suppressing iNOS and NO in LPS-stimulated cells) and can promote Nrf2/ARE antioxidant responses [17].
The limitations of the current data include the ex vivo/in vitro nature of the models. Our human leukocytes were from donors with existing inflammation, so baseline cytokines were high and the relative changes were modest. We did not directly measure NF-κB, MAPK phosphorylation or ROS/Nrf2 activation, so the proposed cascade is inferred. Future experiments should track NF-κB p65 nuclear translocation, MAPK activation, and Nrf2 target gene expression in treated cells, and test these extracts (or purified lipids) in animal models of inflammation to validate the pathway. In summary, Tolypothrix extracts appear to inhibit the LPS/TLR4→NF-κB/MAPK axis and boost antioxidant defenses, thereby reducing TNF-α, IL-6 and related cytokines.
The previously defined fatty acid composition of the studied Tolypothrix extracts [17] supports the observed inflammation-inhibitory potential. The fatty acids with highest levels in the samples were palmitic, oleic and linoleic acid. Palmitic acid is the most common saturated fatty acid. It has been shown to interact with Toll-like receptors and activate signaling pathways that promote inflammation [37]. However, in our experiments, we have observed opposite effects of Tolypothrix extracts on human leukocytes. This could be due to the complex composition of the samples that include a significant amount of unsaturated fatty acids, like oleic and linoleic acids, with proven inflammation inhibitory activity and mechanism of action involving activation of anti-inflammatory responses [38,39]. Higher levels of stearidonic acid were also present in the extracts obtained from T. tennuis PACC5497 and Tolypothrix sp. PACC5501. SDA contributes to this activity by inhibiting inflammatory cytokine production [36]. The extracts obtained from the two T. distorta strains also contained higher levels of palmitoleic acid. The ability of this fatty acid to counteract the proinflammatory activity of palmitic acid has been demonstrated [40]. Collectively, these findings support our hypothesis that unsaturated fatty acids with anti-inflammatory activity could inhibit the proinflammatory action of palmitic acid present in the samples and collectively contribute to the inflammation inhibitory potential of the Tolypothrix extracts. Further research will define the activities of purified individual compounds, possible synergistic effects of these compounds and their potential to serve as new candidate nutraceuticals.
5. Conclusions
The present study provides detailed morphological characteristics of five strains belonging to the genus Tolypothrix and demonstrates that non-polar extract fractions derived from T. tenuis PACC 5497, T. tenuis PACC 8648, T. distorta CCALA 194, T. distorta SAG 1482-2, and Tolypothrix sp. PACC 5501 are able to reduce proinflammatory cytokine production by human peripheral blood leukocytes and sustain the levels of the main immune cell populations without an induction of overactivation. An original contribution from our investigations is the demonstrated ability of Tolypothrix extracts to counteract the active inflammatory process and induce immunoregulatory lymphocyte phenotype ex vivo. These data indicate the important biological activity of the examined extract fractions and pave the way for future studies leading to identification of new targets for drug development and the establishment of natural product-based formulations that support the functions of the immune system.
Author Contributions
Conceptualization, B.D. and I.T.; methodology, B.D., D.B., I.T. and T.B.; software, B.D. and D.M.; validation, B.D., D.B., I.T., K.M. and T.B.; formal analysis, B.D., D.B., I.T., K.M. and T.B.; investigation, B.D., D.B., D.M., I.T., K.M. and T.B.; resources, B.D. and I.T.; data curation, B.D., D.B., D.M., I.T. and T.B.; writing—original draft preparation, T.B.; writing—review and editing, B.D., D.B., D.M., K.M., I.T. and T.B.; visualization, B.D. and I.T.; supervision, B.D. and I.T.; project administration, I.T.; funding acquisition, I.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Bulgarian National Science Found, grant number KP-06-N51/5, and project BG05SFPR001-3.004-0016-C01, “Support for the Development of Project-based Doctoral Studies at the University of Plovdiv ‘Paisii Hilendarski’”, under the “Education” Programme 2021–2027, co-financed by the European Social Fund Plus (ESF+).
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Research Ethics Committee at the University of Plovdiv (protocol No. 7 from 10 June 2024).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The original contributions presented in this study are included in the article.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ALA | Alpha-linolenic acid |
| ARE | Antioxidant response element |
| CCALA | Culture Collection of Autotrophic Organisms |
| CD | Cluster of differentiation |
| CRP | C-reactive protein |
| CTLA-4 | Cytotoxic T-lymphocyte-associated antigen-4 |
| DHA | Docosahexaenoic acid |
| DMEM | Dulbecco’s modified Eagle medium |
| DPBS | Dulbecco’s phosphate-buffered saline |
| IFN-γ | Interferon-gamma |
| IL | Interleukin |
| iNOS | Inducible nitric oxide synthase |
| LPS | Lipopolysaccharide |
| MAPK | Mitogen-Activated Protein Kinase |
| NF-κB | Nuclear factor kappa B |
| NO | Nitric oxide |
| Nrf2 | Nuclear factor erythroid 2-related factor 2 |
| PACC | Plovdiv algal culture collection |
| PD-1 | Programmed death-1 |
| ROS | Reactive oxygen species |
| SAG | Sammlung von Algenkulturen Göttingen |
| SDA | Stearidonic acid |
| TEM | Transmission electron microscopy |
| TNF-α | Tumor necrosis factor-alpha |
References
- Matsuo, T.; Ito-Miwa, K.; Hoshino, Y.; Fujii, Y.I.; Kanno, S.; Fujimoto, K.J.; Tsuji, R.; Takeda, S.; Onami, C.; Arai, C.; et al. Archaean green-light environments drove the evolution of cyanobacteria’s light-harvesting system. Nat. Ecol. Evol. 2025, 9, 599–612. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S. Chapter 1—Cyanobacterial imprints in diversity and phylogeny. In Advances in Cyanobacterial Biology; Singh, P.K., Kumar, A., Singh, V.K., Shrivastava, A.K., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 1–15. [Google Scholar]
- Guiry, M.D. How many species of algae are there? A reprise. Four kingdoms, 14 phyla, 63 classes and still growing. J. Phycol. 2024, 60, 214–228. [Google Scholar] [CrossRef]
- Jeong, Y.; Cho, S.H.; Lee, H.; Choi, H.K.; Kim, D.M.; Lee, C.G.; Cho, S.; Cho, B.K. Current Status and Future Strategies to Increase Secondary Metabolite Production from Cyanobacteria. Microorganisms 2020, 8, 1849. [Google Scholar] [CrossRef]
- Dixit, R.B.; Suseela, M.R. Cyanobacteria: Potential candidates for drug discovery. Antonie Van Leeuwenhoek 2013, 103, 947–961. [Google Scholar] [CrossRef]
- Sabat, S.; Patra, S.; Swain, S.; Bej, S.; Bishoyi, A.K.; Sahoo, C.R.; Padhy, R.N. Phycocompounds from Cyanobacteria: Exploring Synergistic Effects with Conventional Anticancer and Antimicrobial Properties. ACS Omega 2025, 10, 23957–23980. [Google Scholar] [CrossRef]
- Welker, M.; von Dohren, H. Cyanobacterial peptides—Nature’s own combinatorial biosynthesis. FEMS Microbiol. Rev. 2006, 30, 530–563. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Tiwari, S.P.; Rai, A.K.; Mohapatra, T.M. Cyanobacteria: An emerging source for drug discovery. J. Antibiot. 2011, 64, 401–412. [Google Scholar] [CrossRef]
- Komarek, J. Cyanoprokaryota. 3. Heterocytous genera. In Süswasserflora von Mitteleuropa/Freshwater Flora of Central Europe; Büdel, B., Gärtner, G., Krienitz, L., Schagerl, M., Eds.; Springer Spektrum: Berlin, Germany, 2013; pp. 1–1130. [Google Scholar]
- Komarek, J.; Johansen, J. Chapter 4—Filamentous Cyanobacteria. In Freshwater Algae of North America: Ecology and Classification; Wehr, J.D., Sheath, R.G., Kociolek, J.P., Eds.; Aquatic Ecology; Academic Press: Cambridge, MA, USA, 2015; pp. 135–235. [Google Scholar]
- Neuhof, T.; Schmieder, P.; Preussel, K.; Dieckmann, R.; Pham, H.; Bartl, F.; von Dohren, H. Hassallidin A, a glycosylated lipopeptide with antifungal activity from the cyanobacterium Hassallia sp. J. Nat. Prod. 2005, 68, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Kar, J.; Ramrao, D.P.; Zomuansangi, R.; Lalbiaktluangi, C.; Singh, S.M.; Joshi, N.C.; Kumar, A.; Kaushalendra; Mehta, S.; Yadav, M.K.; et al. Revisiting the role of cyanobacteria-derived metabolites as antimicrobial agent: A 21st century perspective. Front. Microbiol. 2022, 13, 1034471. [Google Scholar] [CrossRef]
- Prinsep, M.R.; Thomson, R.A.; West, M.L.; Wylie, B.L. Tolypodiol, an antiinflammatory diterpenoid from the cyanobacterium Tolypothrix nodosa. J. Nat. Prod. 1996, 59, 786–788. [Google Scholar] [CrossRef]
- Swain, S.S.; Paidesetty, S.K.; Padhy, R.N. Antibacterial, antifungal and antimycobacterial compounds from cyanobacteria. Biomed. Pharmacother. 2017, 90, 760–776. [Google Scholar] [CrossRef]
- Jin, X.; Zhang, Y.; Zhang, R.; Nguyen, K.U.; Lindsey, J.S.; Miller, E.S. Identification of Putative Biosynthetic Gene Clusters for Tolyporphins in Multiple Filamentous Cyanobacteria. Life 2021, 11, 758. [Google Scholar] [CrossRef]
- Zeeshan, M.; Suhail, S.; Biswas, D.; Farooqui, A.; Arif, J. Screening of selected cyanobacterial strains for phycochemical compounds and biologiacal activities in vitro. Biochem. Cell. Arch. 2010, 10, 163–168. [Google Scholar]
- Teneva, I.; Batsalova, T.; Moten, D.; Petkova, Z.; Teneva, O.; Angelova-Romova, M.; Antova, G.; Dzhambazov, B. Tolypothrix Strains (Cyanobacteria) as a Source of Bioactive Compounds with Anticancer, Antioxidant and Anti-Inflammatory Activity. Int. J. Mol. Sci. 2025, 26, 5086. [Google Scholar] [CrossRef]
- Adamczyk, M.; Bartosinska, J.; Raczkiewicz, D.; Kowal, M.; Surdacka, A.; Krasowska, D.; Michalak-Stoma, A.; Krasowska, D. The Expression of Activation Markers CD25 and CD69 Increases during Biologic Treatment of Psoriasis. J. Clin. Med. 2023, 12, 6573. [Google Scholar] [CrossRef]
- Staub, R. Ernärungsphysiologisch-autökologische Untersuchungen an der planktonische Blaualge Oscillatoria rubescens DC. Schweiz. Z. Hydrol. 1961, 23, 82–198. [Google Scholar]
- Georg-August-Universität Göttingen-Sammlung von Algenkulturen SAG: Media for Algal Cultures. List of Media and Recipes. Available online: https://www.uni-goettingen.de/de/list-of-media-and-recipes/186449.html (accessed on 3 February 2026).
- Teneva, I.; Belkinova, D.; Paunova-Krasteva, T.; Bardarov, K.; Moten, D.; Mladenov, R.; Dzhambazov, B. Polyphasic characterisation of Microcoleus autumnalis (Gomont, 1892) Strunecky, Komarek & J.R.Johansen, 2013 (Oscillatoriales, Cyanobacteria) using a metabolomic approach as a complementary tool. Biodivers. Data J. 2023, 11, e100525. [Google Scholar] [CrossRef] [PubMed]
- Bhatnagar, M.; Parwani, L.; Sharma, V.; Ganguly, J.; Bhatnagar, A. Exopolymers from Tolypothrix tenuis and three Anabaena sp. (Cyanobacteriaceae) as novel blood clotting agents for wound management. Carbohydr. Polym. 2014, 99, 692–699. [Google Scholar] [CrossRef]
- Mares, J.; Strunecky, O.; Bucinska, L.; Wiedermannova, J. Evolutionary Patterns of Thylakoid Architecture in Cyanobacteria. Front. Microbiol. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, F.; Reis, M.; Ferreira, L.; Grosso, C.; Ferraz, R.; Vieira, M.; Vasconcelos, V.; Martins, R. The Neuroprotective Role of Cyanobacteria with Focus on the Anti-Inflammatory and Antioxidant Potential: Current Status and Perspectives. Molecules 2024, 29, 4799. [Google Scholar] [CrossRef]
- Tena Perez, V.; Apaza Ticona, L.; Cabanillas, A.H.; Maderuelo Corral, S.; Rosero Valencia, D.F.; Quintana, A.M.; Ortega Domenech, M.; Rumbero Sanchez, A. Anti-inflammatory activity of glycolipids isolated from cyanobacterium Nodularia harveyana. Nat. Prod. Res. 2021, 35, 6204–6209. [Google Scholar] [CrossRef]
- Al-Awadhi, F.H.; Simon, E.F.; Liu, N.; Ratnayake, R.; Paul, V.J.; Luesch, H. Discovery and Anti-Inflammatory Activity of a Cyanobacterial Fatty Acid Targeting the Keap1/Nrf2 Pathway. Mar. Drugs 2023, 21, 553. [Google Scholar] [CrossRef]
- Tarasuntisuk, S.; Palaga, T.; Kageyama, H.; Waditee-Sirisattha, R. Mycosporine-2-glycine exerts anti-inflammatory and antioxidant effects in lipopolysaccharide (LPS)-stimulated RAW 264.7 macrophages. Arch. Biochem. Biophys. 2019, 662, 33–39. [Google Scholar] [CrossRef]
- Kang, M.R.; Jo, S.A.; Lee, H.; Yoon, Y.D.; Kwon, J.H.; Yang, J.W.; Choi, B.J.; Park, K.H.; Lee, M.Y.; Lee, C.W.; et al. Inhibition of Skin Inflammation by Scytonemin, an Ultraviolet Sunscreen Pigment. Mar. Drugs 2020, 18, 300. [Google Scholar] [CrossRef] [PubMed]
- Pagels, F.; Guedes, A.C.; Vasconcelos, V.; Lopes, G. Anti-inflammatory compounds from cyanobacteria. In The Pharmacological Potential of Cyanobacteria; Lopes, G., Silva, M., Vasconcelos, V., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 81–105. [Google Scholar]
- Peng, Y.; Tao, Y.; Zhang, Y.; Wang, J.; Yang, J.; Wang, Y. CD25: A potential tumor therapeutic target. Int. J. Cancer 2023, 152, 1290–1303. [Google Scholar] [CrossRef]
- Van Coillie, S.; Wiernicki, B.; Xu, J. Molecular and Cellular Functions of CTLA-4. Adv. Exp. Med. Biol. 2020, 1248, 7–32. [Google Scholar] [CrossRef]
- Krishnamurthy, N.; Nishizaki, D.; Lippman, S.M.; Miyashita, H.; Nesline, M.K.; Pabla, S.; Conroy, J.M.; DePietro, P.; Kato, S.; Kurzrock, R. High CTLA-4 transcriptomic expression correlates with high expression of other checkpoints and with immunotherapy outcome. Ther. Adv. Med. Oncol. 2024, 16, 17588359231220510. [Google Scholar] [CrossRef] [PubMed]
- Keir, M.E.; Butte, M.J.; Freeman, G.J.; Sharpe, A.H. PD-1 and its ligands in tolerance and immunity. Annu. Rev. Immunol. 2008, 26, 677–704. [Google Scholar] [CrossRef] [PubMed]
- Lopez, M.M.; Valenzuela, J.E.; Alvarez, F.C.; Lopez-Alvarez, M.R.; Cecilia, G.S.; Paricio, P.P. Long-term problems related to immunosuppression. Transpl. Immunol. 2006, 17, 31–35. [Google Scholar] [CrossRef]
- Funk, C.D. Prostaglandins and leukotrienes: Advances in eicosanoid biology. Science 2001, 294, 1871–1875. [Google Scholar] [CrossRef]
- Mansour, M.; van Ginkel, S.; Dennis, J.C.; Mason, B.; Elhussin, I.; Abbott, K.; Pondugula, S.R.; Samuel, T.; Morrison, E. The Combination of Omega-3 Stearidonic Acid and Docetaxel Enhances Cell Death over Docetaxel Alone in Human Prostate Cancer Cells. J. Cancer 2018, 9, 4536–4546. [Google Scholar] [CrossRef] [PubMed]
- Korbecki, J.; Bajdak-Rusinek, K. The effect of palmitic acid on inflammatory response in macrophages: An overview of molecular mechanisms. Inflamm. Res. 2019, 68, 915–932. [Google Scholar] [CrossRef] [PubMed]
- Alarcon-Gil, J.; Sierra-Magro, A.; Morales-Garcia, J.A.; Sanz-SanCristobal, M.; Alonso-Gil, S.; Cortes-Canteli, M.; Niso-Santano, M.; Martinez-Chacon, G.; Fuentes, J.M.; Santos, A.; et al. Neuroprotective and Anti-Inflammatory Effects of Linoleic Acid in Models of Parkinson’s Disease: The Implication of Lipid Droplets and Lipophagy. Cells 2022, 11, 2297. [Google Scholar] [CrossRef] [PubMed]
- Santa-Maria, C.; Lopez-Enriquez, S.; Montserrat-de la Paz, S.; Geniz, I.; Reyes-Quiroz, M.E.; Moreno, M.; Palomares, F.; Sobrino, F.; Alba, G. Update on Anti-Inflammatory Molecular Mechanisms Induced by Oleic Acid. Nutrients 2023, 15, 224. [Google Scholar] [CrossRef]
- Tsai, Y.W.; Lu, C.H.; Chang, R.C.; Hsu, Y.P.; Ho, L.T.; Shih, K.C. Palmitoleic acid ameliorates palmitic acid-induced proinflammation in J774A.1 macrophages via TLR4-dependent and TNF-alpha-independent signallings. Prostaglandins Leukot. Essent. Fat. Acids 2021, 169, 102270. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.