Abstract
Baking bread is currently experiencing a profound transformation, propelled by an escalating interest in health and wellness, global cuisines, and sustainability efforts. The surface of fruits harbors various microbes, such as yeasts and lactic acid bacteria (LAB). In contrast to Saccharomyces cerevisiae, a commercial yeast, the microbial flora present on the surface of fruits allows bread with greater flavor, texture, and health benefits as seen in sourdough starters. In this research, sourdough starters were created using dried longan and raisin. The microbial flora of the sourdough starters and the fermentation profile were examined, and possible bacteriocin genes were identified. The dried longan starter exhibited bubbling, stickiness, and a drop in pH value from 5.80 to 4.45 at the third day of fermentation. Meanwhile, the raisin starter began bubbling on the sixth day with no change in fluid viscosity, and a small pH value decreased from 4.00 to 3.82. The dried longan starter contained Weissella cibaria, W. paramesenteroides, S. cerevisiae, Pediococcus pentosaceus, Torulaspora delbrueckii and Leuconostoc citreum. On the other hand, raisin starter only contained Candida krusei and T. delbrueckii. To detect antibacterial property in W. paramesenteroides and P. pentosaceus, four bacteriocins, Gassericin A (gaaA), Lactacin F (lafA), Plantaricin S (pls) and Pediocin (ped) were explored. Polymerase chain reaction (PCR) results revealed that gaaA and lafA were present in W. paramesenteroides, which suggested that this strain possessed antimicrobial properties.
1. Introduction
Compared to breads made with commercial yeast, breads baked using wild yeasts and lactic acid bacteria (LAB) provide benefits such as glycemic response, satiety, and gastrointestinal well-being. Additionally, they allow a valuable natural method for enhancing the texture, flavor, and stability of bread [1]. Yeast plays a significant role in the production of beverages and bread thanks to its capability to produce carbon dioxide and ethanol as well as aroma compounds. Especially, LABs are especially crucial in fermentation as they produce desirable acids, flavor compounds, and peptides that inhibit the growth of undesirable microorganisms [2].
Fruits are commonly used ingredients for making sourdough starters, owing to the widespread presence of yeast on their surfaces. Longan (Dimocarpus longan) is one of the most important fruit crops in Tainan, Taiwan. In addition to being consumed fresh, approximately 50% of longan production is dried to extend its storage life and intensify its flavor and aroma. Raisins are popular ingredients for homemade sourdough starters. They naturally harbor wild yeast and a diverse microbial flora which contribute to vibrant and resilient fermentation. For this study, we chose dried longan and dark sultana raisins as the primary materials for creating sourdough starters. Bacteriocins are proteinaceous or peptidic toxins produced by bacteria to inhibit the growth of similar or closely related bacterial strains [3]. Bacteriocins have been identified in many strains of LAB, and their structural genes are identified using a PCR array of primers [4]. Based on the results, we investigated the fermentation profiles of sourdough starters made of dried longan and raisins. The dominant microbes and organic acids produced were also investigated.
2. Materials and Methods
2.1. Preparing Sourdough Starters with Dried Fruit
In total, 50 g of dried fruits, 25 g of sugar, and 100 g of sterile water were added to a beaker. The ingredients were mixed with a stirring bar until the sugar had completely dissolved. Finally, the beaker was covered with plastic wrap and placed at 25 °C.
2.2. Observation of Sourdough Starters
The fermentation of the sourdough starters was observed and recorded on the top and at the side for 7 days. pH values were recorded every day.
2.3. Organic Acid Analysis
The samples were collected on the seventh day of fermentation. After the samples were filtered, they were centrifuged at 12,000 rpm for 30 min. The supernatant was collected and filtered using a PVDF 0.45 μm filter. Samples were then sent to the Organic and Health Food Science and Technology Development Center (OHC) Lab for organic acid analysis following the method of the National Standards of the Republic of China, CNS 12635 [5].
2.4. Bacterial Identification
In this study, colonies were isolated using three media: Luria–Bertani agar (LB agar), De Man, Rogosa, and Sharpe agar (MRS agar), and potato–dextrose agar (PDA). The samples were diluted using sterile water and spread-plated on three types of media sequentially. LB plates were grown in 37 °C, PDA at 25 °C, and MRS plates at 30 °C under anaerobic conditions.
2.5. Identification of Bacteriocin Genes
Genomic DNA was isolated from W. paramesenteroides and P. pentosaceus using Chelex100 chelating resin [6]. Four specific primer sequences for gaaA, lafA, pls, and ped were listed in Table 1 [7,8]. The PCR process was as follows: an initial denaturation step at 95 °C for 5 min, followed by 34 cycles of denaturation at 95 °C for 1 min, annealing at 57 °C for 45 s, and extension at 72 °C for 1 min, using the GeneAmp PCR System 9700. After the final extension at 72 °C for 5 min, the PCR products were analyzed on 1.5% agarose gels, and stained with ethidium bromide.

Table 1.
Primer for bacteriocin genes.
2.6. Fermentation Efficacy Test
A total of 25 g of the sourdough starter filtrate was thoroughly blended with 50 g of flour within a beaker. The initial and 41 h fermented dough heights were then meticulously measured and recorded.
3. Results
3.1. Observation of Sourdough Starters Made of Dried Longan and Raisin
The procedures outlined in Section 2.1 and Section 2.2 were performed for the preparation and observation of the sourdough starters. On the third day, noticeable bubbles were discerned within the sourdough starter containing dried longan (Figure 1), whereas a similar bubbling phenomenon manifested on the fifth day within the raisin-infused starter (Figure 2). Additionally, it is noteworthy that the dried longan exhibited a sticky texture starting from the third day of the fermentation process.
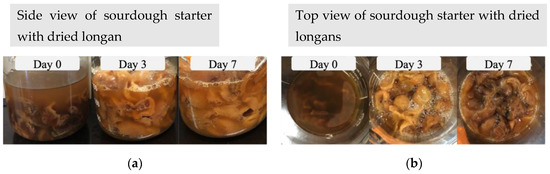
Figure 1.
Side view (a) and top view (b) of sourdough starter with dried longan during a 7-day fermentation.

Figure 2.
Side view (a) and top view (b) of sourdough starter with raisin during a 7-day fermentation.
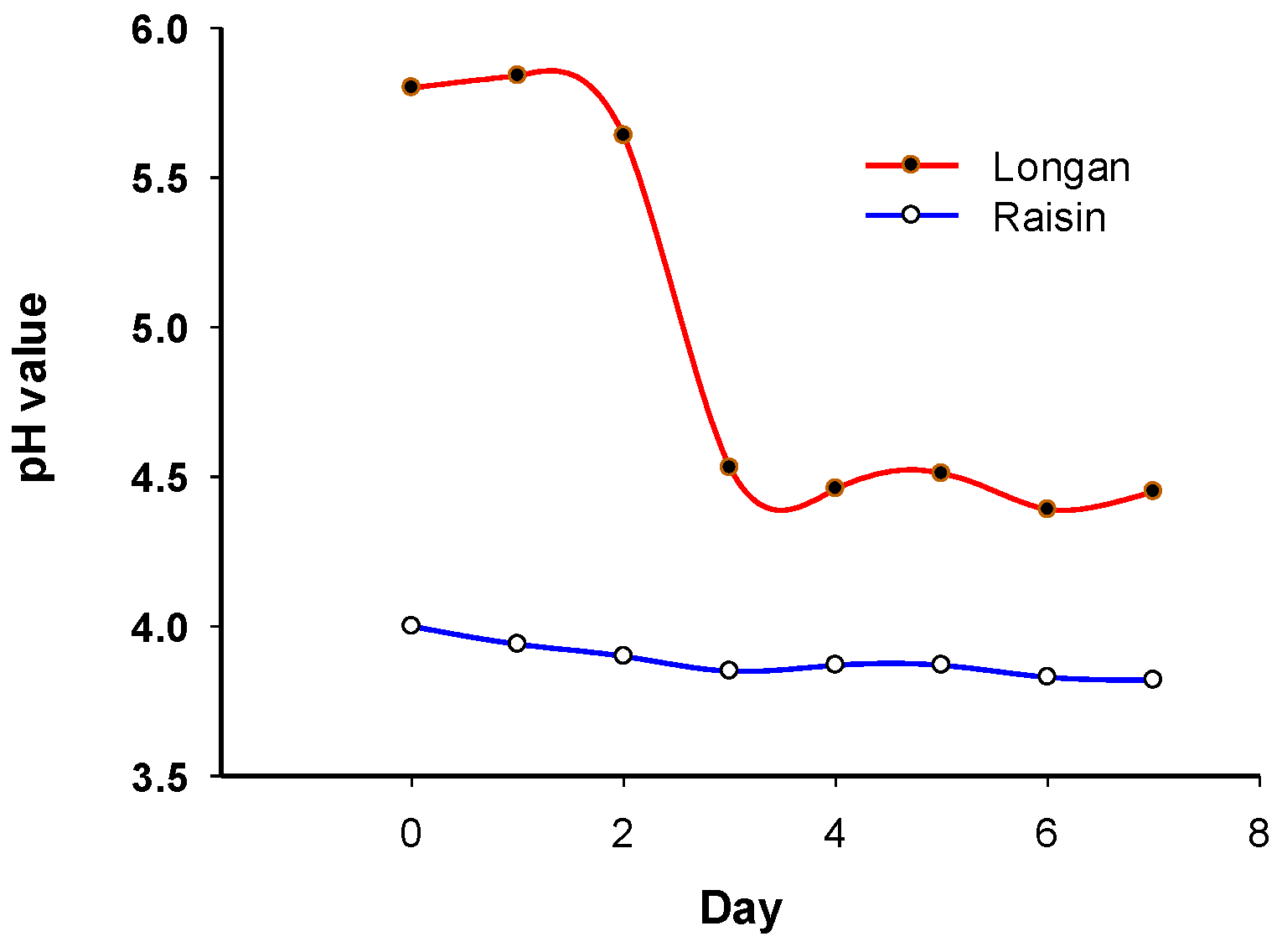
3.2. Change in pH of Sourdough Starters
pH values were monitored throughout the fermentation process. The pH of the sourdough starter with dried longan decreases from 5.80 to 4.45 during fermentation, particularly within the first three days (Figure 3). On the other hand, the pH of the sourdough starter with raisins decreased from 4.00 to 3.82 (Figure 3). There was a rapid drop in pH in the sourdough starter with dried longan compared to that with raisin.
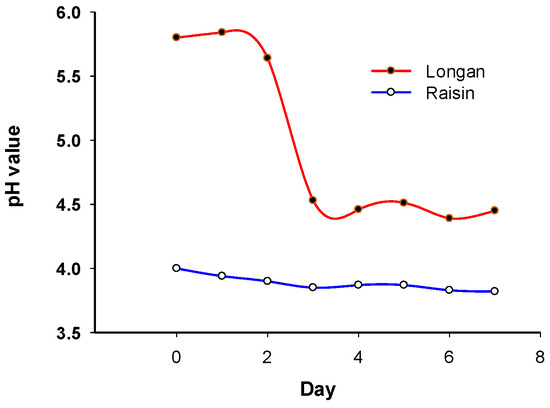
Figure 3.
pH change during fermentation.
3.3. Organic Acid
The result of the high-performance liquid chromatography (HPLC) analysis revealed that within the sourdough starter infused with dried longan, lactic acid emerged as the predominant metabolite in terms of concentration, accompanied by notable levels of acetic acid and citric acid. Conversely, in the sourdough starter incorporating raisin, acetic acid took precedence as the primary metabolite, followed by lactic acid in the second position (Table 2).
Table 2.
Organic acids in the sourdough starter with dried longan and raisin.
3.4. Bacterial Identification
Two sourdough starters were plated onto Luria–Bertani (LB) agar, and ten colonies from each sourdough starter were isolated and transferred to de Man, Rogosa, and Sharpe (MRS) agar or potato–dextrose agar (PDA) for further analysis. From the isolated colonies of the sourdough starter featuring dried longan, a total of four distinct yeast strains were identified, encompassing S. cerevisiae, Zygosaccharomyces (or Lachancea), and two strains of Torulaspora delbrueckii. Further, the analysis yielded the isolation of six LAB strains, namely Weissella paramesenteroides, Leuconostoc citreum, two strains of Pediococcus pentosaceus, and two strains of W. cibaria. In contrast, the sourdough starter incorporating raisin was home to two yeast species, specifically four strains of Candida krusei and an additional four strains of T. delbrueckii (see Table 3). These findings underscore the discernible differences in microbial profiles between the two sourdough starters. Notably, the sourdough starter containing dried longan displayed a higher diversity of yeast and LAB strains compared to the counterpart with raisin.
Table 3.
Species of strains found in sourdough starters with dried longan and raisin.
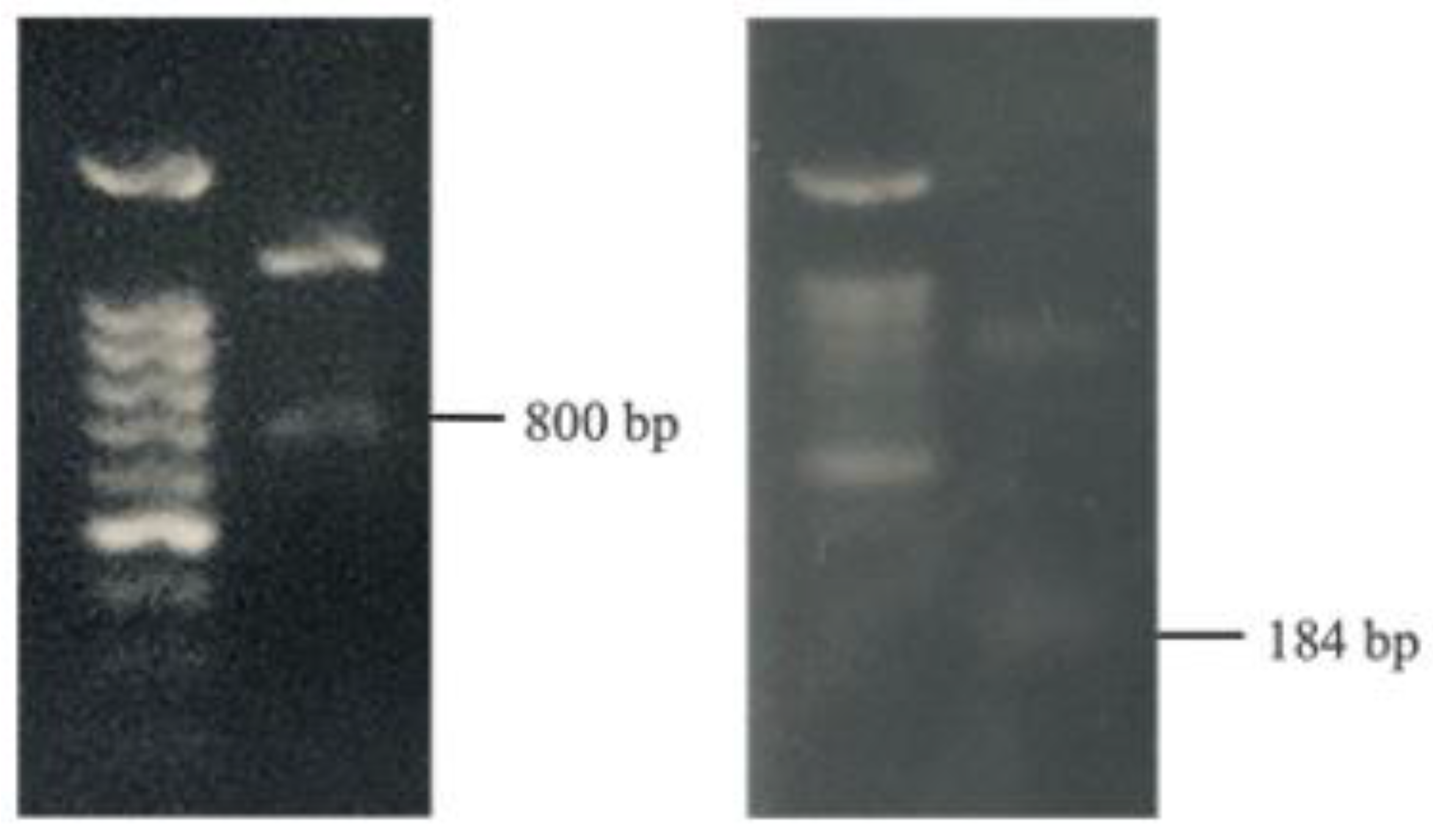
3.5. Identification of Bacteriocin Genes
The evaluation of potential antibacterial bacteriocins encompassed four types: Gassericin A (gaaA), Lactacin F (lafA), Plantaricin S (pls), and Pediocin (ped). Employing PCR, this analysis was conducted on both W. paramesenteroides and P. pentosaceus. Notably, the detection of the 800 bp gaaA gene and the 184 bp lafA gene within W. paramesenteroides provided tangible evidence of the strain’s inherent antibacterial properties (Figure 4).
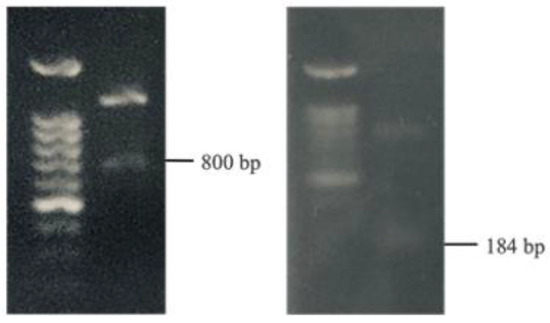
Figure 4.
PCR analysis of bacteriocin genes in W. paramesenteroides.
3.6. Fermentation Efficacy
Fermentation efficacy was assessed for the two distinct sourdough starters. Over a 41 h fermentation period, the volume of the sourdough starter containing dried longan underwent a doubling effect. In stark contrast, the volume of the sourdough starter enriched with raisins remained unaltered (see Figure 5). These findings underscore that the sourdough starter featuring dried longan underwent a more pronounced fermentation compared to its raisin-containing counterpart.

Figure 5.
Fermentation efficacy test of two sourdough starters. (A) at the beginning of fermentation, (B) after 41 h of fermentation.
4. Conclusions
This study delved into the fermentation dynamics of sourdough starters infused with dried longan and raisin. On the third day, the sourdough starter containing dried longan exhibited bubbling along with an augmented stickiness. Simultaneously, a notable decline in pH was observed. Conversely, the sourdough starter with raisin displayed bubbling on the fifth day, without any significant alteration in viscosity. These outcomes underscore the dissimilarities in the activity of microbial flora between the two sourdough starters, a disparity attributed to distinct metabolite production.
The sourdough starter enriched with dried longan harbored a more abundant microbial diversity in comparison to its raisin counterpart. This divergence in microbial flora and metabolite synthesis among the two sourdough starters underscores their potential uniqueness and individuality. Recognizing these distinctions and their potential implications serves as a foundation for further investigations into refining the fermentation process and enhancing the development of superior sourdough starters for baking applications.
Author Contributions
Conceptualization, M.-J.W.; methodology, Y.-C.L. and S.-H.T.; data curation, P.-S.W.; writing—original draft preparation, Y.-C.L.; writing—M.-J.W.; funding acquisition, M.-J.W. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by MOE STEM Project-Talent Cultivation Program in Dietary Supplements, Fermentation Industry and Food Safety, and MOST-110-2320-B-041-002-MY3.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ribet, L.; Dessalles, R.; Lesens, C.; Brusselaers, N.; Durand-Dubief, M. Nutritional benefits of sourdoughs: A systematic review. Adv. Nutr. 2023, 14, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Faria-Oliveira, F.; Diniz, R.; Godoy Santos, F.; Mezadri, H.; Castro, I.; Brandão, R. The Role of Yeast and Lactic Acid Bacteria in the Production of Fermented Beverages in South America; IntechOpen: London, UK, 2015; pp. 107–135. [Google Scholar]
- Negash, A.W.; Tsehai, B.A. Current Applications of Bacteriocin. Int. J. Microbiol. 2020, 2020, 4374891. [Google Scholar] [CrossRef] [PubMed]
- Macwana, S.J.; Muriana, P.M. A ‘bacteriocin PCR array’ for identification of bacteriocin-related structural genes in lactic acid bacteria. J. Microbiol. Methods 2012, 88, 197–204. [Google Scholar] [CrossRef] [PubMed]
- CNS 12635; Method of Test for Fruit and Vegetable Juices and Drinks—Determination of Organic Acids. CNS: Taipei, Taiwan, 2004.
- Walsh, P.S.; Metzger, D.A.; Higushi, R. Chelex 100 as a medium for simple extraction of DNA for PCR-based typing from forensic material. Biotechniques 2013, 54, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Hasannejad Bibalan, M.; Eshaghi, M.; Rohani, M.; Pourshafie, M.R.; Talebi, M. Determination of Bacteriocin Genes and Antibacterial Activity of Lactobacillus Strains Isolated from Fecal of Healthy Individuals. Int. J. Mol. Cell. Med. 2017, 6, 50–55. [Google Scholar] [PubMed]
- Zommiti, M.; Bouffartigues, E.; Maillot, O.; Barreau, M.; Szunerits, S.; Sebei, K.; Feuilloley, M.; Connil, N.; Ferchichi, M. In vitro Assessment of the Probiotic Properties and Bacteriocinogenic Potential of Pediococcus pentosaceus MZF16 Isolated From Artisanal Tunisian Meat “Dried Ossban”. Front. Microbiol. 2018, 9, 2607. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).