Abstract
Plastic pollution has emerged as one of the most pervasive environmental challenges, with microplastics (MPs) widely distributed across marine ecosystems worldwide. This study aimed to assess the uptake of MPs by key fish and invertebrate species from different locations in the coastal zone of the Bulgarian Black Sea. Fish were collected during routine monitoring surveys in September–November 2024, while invertebrates were obtained via scuba diving. The presence of MPs in fish stomachs and invertebrate soft tissues, and their polymer composition, shape and size were analyzed using an Agilent 8700 LDIR Chemical Imaging System. Potential biological effects of ingested MPs were evaluated by an integrated Specific Oxidative Stress (SOS) index. The results revealed MP uptake levels comparable to those reported globally. Small-sized particles (<50 µm) with rounded shapes were most abundant across studied taxa. Polymer composition varied considerably depending on species and sampling region, indicating differences in exposure sources and environmental conditions. Oxidative stress levels in both fish and invertebrates showed substantial interspecific variation, and clear differences between the northern and southern region of the Bulgarian Black Sea. Overall, elevated uptake of MPs appears to contribute to oxidative stress in marine organisms, potentially affecting their health status, resilience, and adaptive capacity, as reflected by increased SOS index values.
Keywords:
adaptability; fish; marine invertebrates; marine pollution; oxidative stress; plastic polymers Key Contribution:
This work provides new evidence of species- and region-specific microplastic accumulation in the Bulgarian Black Sea, supported by comprehensive characterization of particle size, shape, and chemical composition. It further advances biomonitoring by demonstrating the potential of the Specific Oxidative Stress (SOS) index as an integrated tool to assess the biological effects of environmental conditions (including microplastic pollution).
1. Introduction
Marine ecosystems play an important role in sustaining human societies by providing a wide range of ecosystem services essential to well-being and economic development. They are a significant and crucial source of nourishment [1], support global trade and transportation networks [2], and also offer significant cultural and recreational benefits, enhancing quality of life. Beyond direct human uses, marine ecosystems also perform vital regulatory functions. Oceans play a central role in climate regulation by absorbing approximately a quarter of anthropogenic carbon dioxide emissions and over 90% of excess heat generated by greenhouse gas accumulation [3]. Coastal habitats act as natural buffers, protecting shorelines from erosion, storm surges, and extreme weather events while also serving as significant carbon sinks [4,5]. These ecosystems are therefore indispensable for both climate mitigation and adaptation strategies.
Despite their importance, marine ecosystems are increasingly threatened by a range of anthropogenic stressors. Pollution represents one of the most pervasive challenges, encompassing chemical contaminants, oil spills, and, more recently, plastic and microplastic debris. It is estimated that millions of tons of plastic waste enter the oceans annually, where it persists, fragments, and interacts with marine organisms at multiple trophic levels [6]. Such pollutants can cause physical harm, disrupt physiological processes, and act as vectors for toxic substances and invasive species. The cumulative effects of these pressures, often acting simultaneously with overfishing, habitat degradation, and climate change, are driving significant declines in marine biodiversity and marine ecosystem resilience. Researching and understanding the extent and impacts of pollution is therefore critical for the sustainable management and conservation of marine biodiversity and resources.
Plastic pollution has emerged as one of the most pervasive environmental challenges of the 21st century, with microplastics (MPs) (particles <5 mm) now widely detected across marine ecosystems worldwide [7,8]. Due to their small size and persistence, MPs are readily ingested by a wide range of organisms, from plankton to fish and mammals [9], including humans. Studies have shown that these are capable of bioaccumulating in organisms and transferring along trophic chains, with the potential for biomagnification within higher levels of the food web [10,11].
The Black Sea suffers the global trend of increasing marine pollution, with marine litter recognized as one of the major environmental pressures affecting its ecological status and overall environmental health [12,13,14]. Major contaminants include excess nutrients from agriculture and wastewater, heavy metals from industrial activities, persistent organic pollutants such as PCBs and pesticides, petroleum hydrocarbons, and increasing levels of plastic and MP debris [15,16,17]. Due to limited water exchange with the Mediterranean Sea, these pollutants tend to accumulate, particularly in coastal zones and sediments, leading to eutrophication, hypoxia, and long-term ecological degradation [18]. In recent years, MP contamination and their accumulation in biota have become an increasing concern in the Black Sea, too, contributing to additional stress on the already impacted ecosystems [19,20,21,22]. MPs were detected in water, sediments, and marine organisms [23,24,25,26,27,28,29,30,31]. These particles originate from diverse sources, including riverine inputs, coastal urbanization, fisheries, and tourism, and are influenced by regional hydrodynamics and environmental conditions [15,17,32].
In the Bulgarian part of the Black Sea, macroplastic and microplastic pollution is mainly driven by land-based sources, with river input as the dominant pathway, especially the Danube River, which transports large amounts of waste from a vast upstream catchment [24,32]. Coastal urban areas are also important sources of MPs due to population density, tourism, port activities, and wastewater discharge [25,29]. Fishing activities also introduce plastics, both directly (e.g., nets, ropes) and indirectly through the breakdown of gear [33]. Water and sediment studies show that MP levels in the Bulgarian part of the Black Sea are comparable to or higher than those reported in other Black Sea parts and the Mediterranean Sea and Baltic Sea, highlighting the severity of MP pollution [23,25,34,35,36,37,38,39]. Accumulation of MPs was observed in numerous marine species collected along the Bulgarian Black Sea coast [26,27,28,30,31].
Currently, increasing attention has been directed toward the biological effects of MPs on marine organisms. Practically, the effects of MPs are not directly related to their presence and quantity in the environment but depend on their bioaccumulation in the organism. Ingestion is the main, but not only, way for different MPs to enter an organism. Depending on their size, shape, and charge, MPs (and also nanoplastics) can penetrate biological barriers and accumulate in various organs, potentially causing adverse health effects through physical irritation, intoxication by the release of chemical additives and compounds from the plastic matrix, as well as by the effects of toxic substances (e.g., heavy metals) and pathogenic microorganisms adsorbed on their surfaces [9,40,41,42]. At the individual level, MPs’ impacts have been associated with alterations in feeding behavior, growth, development, reproduction, and lifespan. In turn, these changes may cascade to higher levels of biological organization, negatively affecting populations, communities, and ecosystem functioning [43,44,45].
One of the primary cellular mechanisms of MP-induced toxicity is the induction of oxidative stress (OS), which results from the imbalance between pro- and antioxidant processes [43,46,47,48]. The overproduction of reactive oxygen species (ROS) and/or inhibition of the antioxidant defense in cells can lead to lipid peroxidation, protein oxidation, and DNA damage. The ROS and oxidatively modified biomolecules may alter antioxidant enzyme activities, disrupt metabolic processes, reduce energy availability, and trigger signaling pathways, ultimately compromising organismal health and fitness [43,47,48]. It should be emphasized that oxidative stress (OS) is a general cellular response to a wide range of exogenous and endogenous factors and, by itself, does not allow clear discrimination between different stressors, particularly in field studies where multiple environmental factors act simultaneously. However, laboratory studies have demonstrated that MP intake can induce oxidative stress responses [49,50]. Under natural environmental conditions, MPs may therefore contribute to OS development and to the impairment of organism health, potentially acting in combination with other environmental stressors [43,47,48].
Research conducted in the Bulgarian sector of the Black Sea provides important insights into these processes. Recent findings indicate species-specific patterns of MP accumulation in both invertebrates and fish, with significant correlations observed between MP load and OS biomarkers such as catalase activity, glutathione levels, and lipid peroxidation [27,31]. These results suggest that MP pollution acts as a chronic environmental stressor, eliciting adaptive and potentially detrimental biological responses depending on species sensitivity and ecological conditions. Thus, oxidative stress biomarkers can serve as early warning indicators of physiological disturbances with potential downstream consequences at the ecosystem level [44,45,51].
The aim of this study was first to comprehensively assess the count, main polymer types, shape and size of MPs ingested by key fish and invertebrate species from the Bulgarian part of the Black Sea, and second, to evaluate the potential effects of MP uptake on their health status, resilience capacity, and adaptive potential, by using oxidative stress as an integrative biomarker within a systems-based approach. The obtained results are also expected to provide region-specific baseline data on the effects of contamination levels, including the MP uptake from the environment in marine species from the Bulgarian Black Sea. These findings may contribute to a better understanding of species-specific responses to contaminant exposure pathways, which can support the identification of reliable bioindicators, and inform more effective health and ecological risk assessment and future marine management strategies.
2. Materials and Methods
2.1. Sampling Area
Fish species were caught during routine monitoring surveys at certain trawling sites from the northern and southern regions of the Bulgarian part of the Black Sea, following a standardized methodology [52] (Figure 1). The sampling was performed using a standard bathypelagic otter midwater trawl (OTM) with a vertical mouth opening of 5 m, a total length of 25 m, and a mesh size of 6.5 mm. The invertebrate species were also collected by scuba diving. All specimens were collected during the same sampling period (September–November 2024) in order to reduce potential differences related to seasonal variations.
Figure 1.
Sampling polygons.
2.2. Study Species
Fish and invertebrates specimens were randomly gathered from the different stations’ trawling and diving catches. The key fish species selected for subsequent analyses included the demersal species, Mullus barbatus Linnaeus, 1758 (red mullet), Mesogobius batrachocephalus, Pallas, 1814 (toad goby), and Neogobius melanostomus (Pallas, 1814) (round goby), as well as pelagic species, Pomatomus saltatrix (Linnaeus, 1766) (bluefish), Trachurus mediterraneus (Steindachner, 1868) (horse mackerel), Sprattus sprattus (Linnaeus, 1758) (sprat), and Engraulis encrasicolus (Linnaeus, 1758) (European anchovy).
The studied invertebrate species of importance for ecosystems included Mytilus galloprovincialis Lamarck, 1819 (black mussels), Donax trunculus, Linnaeus, 1758 (white clams), Rapana venosa Valenciennes, 1846 (rapa whelk), and Palaemon adspersus Rathke, 1837 (prawn).
All samples were immediately shock-frozen and wrapped in aluminum foil on board and subsequently transferred to the laboratory in dry ice. Upon arrival, they were stored at −80 °C until further biochemical analysis.
2.3. Fish and Invertebrate Species Analyses
A total of 178 fish and 94 invertebrates were examined in the present study for both MP ingestion and OS. Prior to analysis, a part of the fish and invertebrates was thawed at room temperature. Their weight (W) and length (L) were recorded in grams and centimeters, respectively. For fish, Fulton’s condition index (K) was calculated according to the following formula: K = W/L3 × 100 [53]. Fulton’s condition factor was not calculated for the invertebrates because they exhibit substantial differences in body morphology, and, unlike fish, there is no universally standardized condition factor applicable across invertebrate taxa.
During biometric measurements and following dissection, soft tissue extraction, and sample processing, strict protocols were applied to minimize the risk of MP cross-contamination [54]. All specimens were subjected to identical, standardized sampling and processing procedures. Therefore, any artifacts potentially arising during collection, storage, or laboratory handling would have affected all samples uniformly and were unlikely to have biased the relative comparisons among species. In addition, individuals with relatively similar body size and morphology were selected, to minimize variability associated with age and developmental stage.
Species-specific ecological characteristics, biometric data, and Fulton’s condition factor of the investigated fish species are summarized in Table 1.

Table 1.
Biometric characteristics and Fulton’s condition index (K) of the studied fish species.
2.4. Microplastics Analyses
The presence of ingested microplastics in fish stomachs and also in soft tissues of invertebrates, along with their size, shape, and polymer type, was analyzed at an external certified laboratory (SGS Bulgaria Ltd., Varna, Bulgaria) using the Agilent 8700 LDIR Chemical Imaging System (Agilent Technologies, Inc., Santa Clara, CA, USA). The Agilent 8700 LDIR Chemical Imaging System is designed to classify microplastics based on their chemical composition using infrared spectroscopy (QCL-based LDIR technology). The system identifies polymers through comparison of the obtained infrared spectra with reference spectral libraries using the analytical software. The hit quality criteria applied in this study were based on the spectral matching score produced by the system software. Only particles with spectra showing an acceptable match quality above the instrument-recommended threshold were considered positively identified and included in the analysis. Spectra with low matching quality, ambiguous identification, or poor signal-to-noise ratio were excluded to minimize false-positive identification of microplastics. The size and shape of the particles were determined by the instrument software based on particle boundaries identified during imaging analysis. In this study, MPs were grouped into five size classes (<50, 51–100, 101–300, 301–1000, and 1001–5000 µm) and four shape categories: rounded particles, thin fibers, coarse fibers, and irregular fragments (Figure 2). The distinction between “thin” and “coarse” fibers was based on the software’s automatic classification according to aspect ratio, where thin fibers exhibited smaller diameters relative to their length, while coarse fibers showed larger diameters and a more robust morphology. These categories should be considered operational definitions generated by the analytical software.

Figure 2.
Representative shape categories of identified microplastics: rounded particles, thin fibers, coarse fibers, and irregular fragments, as classified by the Agilent 8700 LDIR Chemical Imaging System software Clarity, Version: 1.7.27.
In SGS, as a certified laboratory (https://www.sgs.com/en-bg/our-company/about-sgs/sgs-in-bulgaria, accessed on 14 April 2026), strict contamination prevention procedures are routinely applied. These measures included the use of filtered water and filtered reagents throughout the analytical procedure, rinsing all instruments and laboratory containers with filtered water prior to use, and covering the used vessels with foil whenever possible to reduce airborne contamination. In addition, laboratory personnel wore cotton laboratory coats and clothing to minimize the release of synthetic fibers. Furthermore, procedural blanks were included and processed under the same laboratory conditions as the environmental samples in order to assess potential background contamination and cross-contamination. The results from the blanks indicated negligible contamination and no consistent contamination pattern that could significantly affect the analytical results. All analyses were performed within the framework of the IAEA NUTEC Plastics Initiative (https://www.iaea.org/services/key-programmes/nutec-plastics, accessed on14 April 2026). Under the IAEA guidance document, a spectral “hit” is accepted only when it meets strict validation criteria: the spectrum is noise-controlled baseline sound, covers the required fingerprint region, matches a validated reference with confirmed diagnostic peaks, and is contamination-free, consistent with particle morphology, fully traceable, and reproducible across laboratories. The reference laboratories are the IAEA Environment Laboratories in Monaco, which operate two LDIR systems for microplastic analysis. Pictures from the analyses are presented in Appendix A.
2.5. Tissue Preparation for Oxidative Stress Analysis
After defrosting, biometric measurements of the studied individuals were taken and recorded. Each fish was dissected, and the liver and gills were carefully excised on ice. These two organs were selected because they are particularly vulnerable to oxidative stress induced by both exogenous and endogenous factors. Oxidative stress levels in the gills reflect the impact of direct environmental exposure and continuous contact with suspended particles, including MPs, whereas in the liver they reflect internal processing and detoxification processes, integrating systemic responses to ingested MPs and associated contaminants. Similarly, the soft tissues of the examined invertebrates were dissected from their shells and exoskeletons over ice. All organs and tissues were then finely minced and homogenized in 0.1 M potassium phosphate buffer (pH 7.4) to yield a 10% (w/v) tissue homogenate using a Potter–Elvehjem homogenizer equipped with a Teflon pestle (Thomas Scientific LLC, Swedesboro, NJ, USA). The homogenates were centrifuged at 3000× g for 10 min to obtain a post-nuclear fraction, which was subsequently used for determination of lipid peroxidation (LPO) and total glutathione (GSH) levels. An aliquot of the post-nuclear fraction was further centrifuged at 20,000× g for 20 min to obtain a post-mitochondrial supernatant. This fraction was used for the assessment of the activities of antioxidant enzymes: superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPx), glutathione reductase (GR), and glutathione-S-transferase (GST), as well as acetylcholinesterase (AChE). All procedures were conducted at 4 °C.
2.6. Biochemical Analyses
Oxidative stress biomarkers in the liver and gill supernatants were determined spectrophotometrically using commercial kits obtained from Sigma-Aldrich Co. LLC (St. Louis, MO, USA): Lipid Peroxidation (MDA) Assay Kit (Product No. MAK568), Glutathione Assay Kit (Product No. CS0260), Superoxide Dismutase Assay Kit (Product No. 19160), Catalase Assay Kit (Product No. CAT100), Glutathione Peroxidase Cellular Activity Assay Kit (Product No. CGP1) and Glutathione Reductase Activity Assay Kit (Product No. GRSA). All analyses were conducted strictly according to the manufacturer’s protocols, including calibration with the standard solutions supplied in each kit. Biomarker concentrations and enzyme activities were expressed per mg protein.
Acetylcholinesterase activity was determined according to Ellman’s method [55]. The reaction mixture consisted of 0.1 M potassium phosphate (K–PO4) buffer (pH 8.0), 0.045 M acetylthiocholine iodide as substrate, 0.008 M 5,5′-dithiobis-(2-nitrobenzoic acid) (DTNB), and an appropriate volume of fish or invertebrate tissue homogenate. Acetylcholinesterase catalyzes the hydrolysis of acetylthiocholine to thiocholine, which subsequently reacts with DTNB to produce the yellow-colored compound 5-thio-2-nitrobenzoate. The rate of formation of the colored product was monitored spectrophotometrically by recording the absorbance increase at 412 nm. Enzyme activity was expressed as units (U), defined as μmol of acetylthiocholine hydrolyzed per minute per mg of protein.
Protein concentrations in the samples were determined according to the method of Lowry [56], with bovine serum albumin used as a standard for calibration curve preparation.
To ensure analytical accuracy and reproducibility, all of the measurements were performed in triplicate, and the resulting mean values were used for subsequent statistical analyses.
2.7. Specific Oxidative Stress (SOS) Index
The Specific Oxidative Stress (SOS) index was applied as an integrated measure of oxidative status, following the methodology of Yakimov et al. [57] with some modifications [27,29]. Briefly, the SOS index is defined as the difference between the pro-oxidant score (ProO) and the antioxidant score (AntiO), SOS = ProO − AntiO, where ProO was determined as the mean of the standardized (Z-score) values of the assessed pro-oxidant biomarkers (LPO) (PrO = average (zLPO)) and AntiO was obtained as the mean of the Z-scores of the antioxidant parameters (GSH, SOD, CAT, and GPx) (AntiO = average (zSOD + zCAT + zGPX + zGSH). The standardization of the measured biological endpoints by the Z-score transformation ensured that variables with different units and scales became comparable. As a result, each biomarker contributes equally in relative terms to the composite indices, preventing parameters with larger numerical ranges from disproportionately influencing the final score. Specific Oxidative Stress values around zero indicate a balanced pro/antioxidant status, while negative or positive values reflect deviations below or above the site-specific mean, respectively, thereby characterizing the oxidative stress level of the organisms and their adaptive capacity.
2.8. Statistical Analyses
Statistical analyses were performed using the STATISTICA 10 software package (StatSoft Inc., Tulsa, OK, USA). Data are presented as mean ± standard deviation (SD). Prior to statistical testing, data distribution was assessed using the Shapiro–Wilk normality test. Depending on the distribution of the data, differences between groups were evaluated using either Student’s t-test for normally distributed datasets or the Mann–Whitney test for non-normally distributed data. Statistical significance was accepted at p < 0.05. Principal component analysis (PCA) was additionally applied to evaluate relationships among the studied variables.
3. Results
3.1. Biometric Characteristics of the Studied Marine Organisms
Table 1 summarizes the mean weight, total length, and condition factor (K) of the studied fish species sampled from stations of the trawling sites located in the northern and southern coastal regions. Biometric variation was primarily species-specific, mainly reflecting inherent differences among taxa.
When comparing individuals of the same fish species between the northern and southern regions, no statistically significant differences were observed in the measured parameters. The only exceptions were T. mediterraneus and S. sprattus, where individuals from the northern region showed significantly higher mean body weight, while S. sprattus also exhibited greater mean length compared to conspecifics from the southern region.
The highest condition factor (K) was calculated for N. melanostomus from both regions, whereas the lowest values were observed for E. encrasicolus and S. sprattus. Mean condition factor higher than 1 had only M. barbatus and N. melanostomus from both areas, and P. saltatrix from the northern region. A condition factor below 1 indicates relatively unfavorable environmental conditions, including limited resource availability and/or intensive exploitation. Accordingly, the results suggested that most of the studied fish species were in suboptimal condition, reflecting physiological stress with an adverse effect on their growth patterns and overall health status.
The mean weights of the studied invertebrates, as a morphological characteristic, are presented in Table 2. Although no statistically significant differences were observed between specimens from the northern and southern regions (except for D. trunculus), invertebrates from the northern coastal region generally exhibited a tendency toward higher body weights, which may reflect differences in local environmental conditions and/or resource availability. In contrast, D. trunculus from the southern region showed significantly higher values, indicating more favorable habitat suitability for this species.
Table 2.
Mean weights of the studied invertebrate species.
3.2. Microplastics in the Studied Fish and Invertebrates
3.2.1. MP Counts
MP uptake in the stomachs of fish and invertebrate soft tissues showed pronounced interspecific and spatial variability (Table 3).
Table 3.
MP counts in fish stomach and invertebrate soft tissues.
In the northern region, MP abundance in fish stomachs ranged from very low values in N. melanostomus to relatively high concentrations in T. mediterraneus, S. sprattus, and E. encrasicolus. Among invertebrates, MP counts in the soft tissues were generally low, with the highest value recorded in D. trunculus.
In the southern region, the highest number of particles in the stomach was recorded in S. sprattus and E. encrasicolus. The lowest concentration was observed in M. barbatus and N. melanostomus. In invertebrates, in the soft tissues of D. trunculus, higher MP abundance was observed, while R. venosa and M. galloprovincialis showed about 10 times lower values.
The results suggested spatial and taxonomic differences in MP intake between fish and invertebrates from the northern and southern regions. Among fish, several species from the southern region, including M. barbatus, M. batrachocephalus, and N. melanostomus, showed a higher MP number in their stomachs, suggesting stronger exposure to sediment-associated contamination in the south. In contrast, pelagic species such as T. mediterraneus and E. encrasicolus from the northern region exhibited higher values compared with those from the southern region, while S. sprattus shows similarly high levels in both areas. P. saltatrix displayed only minor regional variation.
Invertebrates showed a more consistent tendency toward higher values in the individuals from the southern region. This is particularly evident for D. trunculus and P. adspersus, whereas M. galloprovincialis and R. venosa from the north region showed slightly higher values than those from the southern region.
Overall, fish tend to exhibit higher and more variable MP intake than invertebrates, reflecting differences in feeding strategies and trophic transfer. The southern region appeared more impacted for benthic and sediment-associated species, while in some pelagic fish from northern regions, elevated levels were shown, indicating spatial heterogeneity in MP distribution and exposure pathways.
3.2.2. Size-Class Distribution of MPs in Fish and Invertebrates
The size distribution of MPs in fish and invertebrates showed a clear dominance of the smallest fraction across both regions (Table 4).
Table 4.
Size-class percentage distribution of MPs in fish stomachs and invertebrate soft tissues.
In species from the northern region, the highest portions of the <50 µm fraction were observed in S. sprattus and E. encrasicolus among fish, and in R. venosa among invertebrates. Larger particles (1001–5000 µm) were largely absent, with only trace amounts detected in a few species, namely P. saltatrix and D. trunculus.
In the species from the southern region, the highest portion of the <50 µm fraction was detected in M. batrachocephalus and in the studied invertebrates, except for M. galloprovincialis, where a more heterogeneous MP size distribution was observed.
Comparing the size-class distribution of MPs in fish and invertebrates from both regions, a decrease in the predominance of the smallest MPs was observed in N. melanostomus, S. sprattus, and E. encrasicolus, accompanied by an increased contribution presence of the intermediate size classes (51–300 µm). Only two fish species contained particles spanning all size classes: P. saltatrix from the northern region and N. melanostomus from the southern region.
3.2.3. Shape-Class Distribution of MPs in Fish and Invertebrates
The MPs’ shapes were classified based on the measured dimensions and aspect ratios of the detected particles, as determined by the analytical software. The distribution of MPs by shapes showed a clear dominance of rounded particles in biota from both regions, with some species-specific and regional differences (Table 5).
Table 5.
Percentage distribution of MPs by shape class in fish stomachs and invertebrate soft tissue.
In the northern region, rounded particles were >98% in species such as M. batrachocephalus, S. sprattus, and E. encrasicolus for fish and D. trunculus for invertebrates. Irregular fragments were also relatively common in some species, particularly M. galloprovincialis for invertebrates, and M. barbatus and N. melanostomus for fish, while fibers (thin and coarse combined) were generally minor components. All shape classes of MPs were found in three of the invertebrate species from the northern regions.
In the studied individuals from the southern region, rounded particles became even more dominant, reaching 100% in P. saltatrix and T. mediterraneus for fish, and R. venosa for invertebrates, and remaining above 96% in most other species. Fibrous particles were rare and largely absent in many species, with only minimal occurrences of thin or coarse fibers in fish species M. barbatus, N. melanostomus, and E. encrasicolus. Irregular fragments showed relatively higher proportions in N. melanostomus and E. encrasicolus.
A comparison of the shapes of MPs, found in animals from the northern and southern regions, revealed a shift toward a more uniform distribution in southern samples, which exhibited a higher proportion of rounded particles, reaching complete dominance in P. saltatrix and T. mediterraneus for fish, and R. venosa for invertebrates. In contrast, in the northern region samples, a more heterogeneous composition was presented, with a higher contribution of irregular fragments in several species.
3.2.4. Polymer Composition of MPs in Fish and Invertebrates
The polymer composition of MPs in fish stomachs and invertebrate soft tissues is shown in Table 6. In the biota from both the northern and southern region, polymer profiles of the MP uptake were highly variable among species. Dominant polymer types (>50%) in the specimens from the northern region were rubber in M. barbatus and M. galloprovincialis, PA in N. melanostomus and D. trunculus, and ABS in T. mediterraneus, and in those from the southern region were PA in M. barbatus, rubber in P. saltatrix and T. mediterraneus, and PP in P. adspersus. Overall, a dominance of rubber and PA was established.
Table 6.
Percentage distribution of MPs by polymer type in fish stomachs and invertebrate soft tissues.
In M. barbatus from the northern region, rubber MPs were dominant (58.82%), whereas the southern population was overwhelmingly dominated by polyamide (PA) (88.88%), indicating a strong regional shift in polymer type. In P. saltatrix from the northern region, a mixed composition of MPs (rubber, PE, PA, and PET) was established, while in the samples from the southern region, rubber was clearly dominant (67.26%), with a reduced contribution of other polymers. For T. mediterraneus, in the northern population, the dominant type of MPs was ABS (76.19%), whereas the southern population shifted toward an MP profile dominated by rubber (62.5%). In M. batrachocephalus from both regions, mixed polymer compositions were detected, but in the samples from the south, a more balanced distribution was displayed, with significantly reduced dominance of PU compared to those from the northern region. N. melanostomus from both northern and southern regions showed profiles dominated by PA. In S. sprattus from the northern region, a mixed profile (PE, PA, rubber, PU, and PP) was identified, while in the fish from the southern region, PU (32.99%) and PA (34.01%) were dominant, indicating a shift toward thermoplastic and synthetic elastomer materials. E. encrasicolus from both regions had mixed polymer profiles; however, those from the southern region displayed a higher presence of PU and PET compared to the more balanced northern region distribution in the biota dominated by PA and PP. In M. galloprovincialis from both regions, PET was the predominant polymer type, together with the rubber in the specimen from the north. In D. trunculus, the dominant polymer shifted from PA in the samples from the north (51.25%) to PP in the samples from the south (30.17%), indicating a clear regional replacement of main polymer types. In R. venosa, rubber became strongly dominant in the specimens from the south (50%) compared to a more PA-rich profile in the north. The northern P. adspersus showed a major shift from a mixed composition (rubber, PA, and ABS) to a profile dominated by PP (58.95%) in the southern samples.
Overall, the polymer type of the MPs, found in the biota from the southern region, is characterized by dominance of PA, rubber, PU, and PP (depending on species), whereas in the biota from the northern region a more heterogeneous and multi-polymer composition was identified, suggesting differences in sources, degradation stages, and/or exposure pathways between regions (Table 6).
The PCA analysis highlights the direction of maximum variance in the data cloud and, hence, the correlations of the studied variables with the PC axes. The prevailing MP polymer types in fish had an undefined relation with the northern and southern region.
As a whole, the PCA analyses of fish data (Figure 3a) showed high variance in the presence and distribution of MP polymers in fish stomachs from both the northern and southern Bulgarian Black Sea regions. The first PC axis (PC1) explains 51.59% of the variability in the data on MP polymer dispersion, reflecting differences between fish species from the northern and southern region, with a relatively well-defined positive correlation with MP polymers in M. batrachocephalus from the southern region. Some more clearly defined relations exist along the PC2 axis. Here, several fish species showed correlations with PA, PP, PA, PE and rubber (Figure 3a). On the other hand, PC2 is positively correlated with and reflects the accumulated MP polymers in the separate fish species originating from different habitats. This indicates the significant difference in the accumulated MP polymers in the different fish species and their habitats.
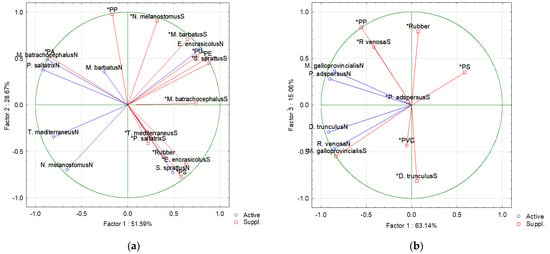
Figure 3.
Results of the PCA analyses of (a) fish and (b) invertebrate species from the northern (N) and southern (S) Bulgarian Black Sea, and the prevailing MP polymer types present in fish stomach and invertebrate soft tissues.
Results of the PCA analyses of the invertebrate species are presented in Figure 3b. PC1 accounts for 63.14% of the variation, and PC2 accounts for 15.06% of the variation. The variance of the relations between species, regions, and polymer type are unclear, as expressed by the correlations with PC1 axis. However, the PC2 axis has some more clearly defined relations between some MP polymer types and invertebrate species. In particular, R. venosa from the southern region seems related to PP and partly to rubber (positive PC2 end). On the negative PC2 end, D. trunculus (southern region) is grouped with PVC. This PCA analysis suggests that in the invertebrates, the main determinant of the accumulation of MP polymer types in the soft tissues appears to be the species (and its habitat).
3.3. Oxidative Stress Levels in Fish Organs and Invertebrate Soft Tissue
3.3.1. Oxidative Stress Levels in Fish Liver
The OS levels and the antioxidant defense biomarkers in the fish liver varied considerably among species and also between the northern and southern regions of the Bulgarian part of the Black Sea (Table 7A).
Table 7.
(A). Oxidative stress biomarkers in the liver of the studied fish species. (B). Oxidative stress biomarkers in the gills of the studied fish species.
In the northern region, LPO levels (expressed as nmoles malondialdehyde (MDA)/mg protein) were generally low in the demersal species such as M. barbatus and M. batrachocephalus, while markedly higher levels were observed in pelagic species, particularly E. encrasicolus and T. mediterraneus. Glutathione concentrations were highest in M. barbatus and T. mediterraneus, whereas small pelagic species such as S. sprattus and E. encrasicolus exhibited substantially lower levels. Antioxidant enzyme activities showed species-specific patterns. Catalase activity was relatively high in N. melanostomus and M. barbatus. Glutathione peroxidase activity was notably elevated in M. batrachocephalus and E. encrasicolus. Glutathione reductase activity showed the highest values in E. encrasicolus, N. melanostomus, and S. sprattus, whereas GST activity was exceptionally high in M. batrachocephalus and N. melanostomus. Acetylcholinesterase activity was highest in E. encrasicolus and lowest in P. saltatrix.
In the species from the southern region, LPO levels were substantially higher in N. melanostomus and E. encrasicolus compared to the other fish species. M. barbatus and M. batrachocephalus showed lower LPO values. Glutathione levels displayed variable trends, with T. mediterraneus exhibiting the highest concentrations, while P. saltatrix and S. sprattus showed markedly reduced levels. Antioxidant enzyme activities varied, as the SOD activity was particularly elevated in N. melanostomus, indicating enhanced OS response. Catalase activity was higher in M. barbatus. Glutathione peroxidase activity was highest in M. batrachocephalus and N. melanostomus, while GR activity showed a pronounced increase in M. barbatus and S. sprattus, suggesting intensified glutathione recycling. Glutathione-S-transferase activity was highest in M. batrachocephalus, consistent with the northern region. Acetylcholinesterase activity exhibited notable interspecific variability, with the highest values in E. encrasicolus and the lowest in N. melanostomus.
A comparison between the northern and southern region clearly revealed species-specific differences in OS responses. In several species, including M. barbatus, P. saltatrix, and N. melanostomus, southern populations exhibited statistically significantly higher LPO, as well as lower GSH levels, thus indicating elevated oxidative stress. This was often accompanied by increased antioxidant enzyme activities (e.g., SOD, CAT, GPx), suggesting activation of compensatory defense mechanisms, although not always sufficient to prevent oxidative damage. Some species, T. mediterraneus and M. batrachocephalus, showed more stable oxidative status between regions, reflecting a better physiological adaptation. Overall, the obtained results indicate a tendency toward higher oxidative pressure in fish from the southern region, with variable efficiency of antioxidant responses among species.
3.3.2. Oxidative Stress Levels in Fish Gills
The levels of OS biomarkers in fish gill tissues are presented in Table 7B. In the northern region, LPO showed the highest values in E. encrasicolus, T. mediterraneus, and S. sprattus, whereas lower levels were recorded in M. barbatus. The highest GSH concentrations were present in T. mediterraneus and P. saltatrix, while the lowest levels were observed in E. encrasicolus and S. sprattus. Superoxide dismutase activity peaked in S. sprattus. In E. encrasicolus, CAT activity was significantly lower, whereas GPx activity was the highest. GR showed elevated values in T. mediterraneus, S. sprattus, and E. encrasicolus. Glutathione-S-transferase activity was markedly higher in M. batrachocephalus compared to the other species. Acetylcholinesterase activity was highest in E. encrasicolus and lowest in S. sprattus.
In the southern sea region, LPO remained highest in E. encrasicolus, T. mediterraneus, and S. sprattus. Glutathione levels were highest in T. mediterraneus, while substantially lower values were measured in N. melanostomus and P. saltatrix. Superoxide dismutase activity was high in several species, particularly in M. barbatus and E. encrasicolus. Catalase activity was the highest in P. saltatrix. Glutathione peroxidase activity reached its highest level in E. encrasicolus. GR activity was markedly elevated in S. sprattus. Glutathione-S-transferase activity was highest in M. batrachocephalus. Acetylcholinesterase activity showed considerable variability, with the highest values in E. encrasicolus and the lowest in N. melanostomus.
A comparison between the northern and southern region revealed pronounced differences in OS responses in fish gills. Southern populations generally exhibited higher LPO, particularly in P. saltatrix, N. melanostomus, and M. barbatus, indicating elevated OS, while LPO remained similarly high in pelagic species such as S. sprattus and E. encrasicolus across both areas. Glutathione levels tended to decrease in the southern regions in several species (P. saltatrix, N. melanostomus, S. sprattus). Antioxidant enzyme activities showed a general upregulation in southern populations, including increased SOD and GPx in M. barbatus, E. encrasicolus, and N. melanostomus, as well as markedly elevated GR in S. sprattus. However, this response was not uniform, as CAT activity remained low and relatively unchanged across species. Notably, P. saltatrix and N. melanostomus displayed a combination of high LPO and reduced GSH, suggesting insufficient antioxidant protection. In contrast, T. mediterraneus and M. batrachocephalus showed comparatively moderate changes between regions, indicating a more stable oxidative balance. Overall, the results suggest increased oxidative pressure in the southern region, accompanied by species-specific and variably effective antioxidant responses in gill tissues.
3.3.3. Oxidative Stress Levels in Invertebrates
In Table 8, the OS biomarker values in the soft tissues of the studied invertebrates are presented. In the northern region, the lowest values of LPO were measured in R. venosa and the highest in M. galloprovincialis. Glutathione concentrations were highest in R. venosa, followed by P. adspersus. Superoxide dismutase and CAT activities were the highest in D. trunculus. Glutathione peroxidase activity showed extremely high values in D. trunculus. Glutathione reductase activity was significantly higher in M. galloprovincialis. GST activity was most pronounced in P. adspersus. Acetylcholinesterase activity varied substantially, with the highest values in R. venosa and the lowest in P. adspersus.
Table 8.
Oxidative stress biomarkers of the studied invertebrates.
In general, the pattern of the OS reaction of the invertebrate species from the southern region was similar to that of the northern region. LPO values were higher in M. galloprovincialis and P. adspersus. The GSH levels were the highest in R. venosa. Superoxide dismutase activity remained highest in D. trunculus, although slightly reduced compared to the northern region. Catalase activity showed minor variation, with the highest values again observed in D. trunculus. Glutathione peroxidase activity remained high in all species. Glutathione reductase showed a higher activity in D. trunculus. Glutathione-S-transferase activity remained significantly higher and stable in P. adspersus, and there was high AChE activity in R. venosa.
Overall, interregional comparisons indicate that invertebrates from the northern and southern regions exhibit similar OS responses, suggesting comparable adaptation to environmental conditions and relatively stable biochemical profiles across regions.
3.3.4. Integrated Oxidative Stress Indexes
The normalized OS indices (and AntiO) and the resulting integrated response (SOS) of the animals toward the exogenous and endogenous (including MPs uptake and tissue accumulation) factors are shown in Table 9. Positive SOS values, indicating elevated relative OS, were recorded in E. encrasicolus, S. sprattus, and T. mediterraneus, as well as in M. galloprovincialis and P. adspersus from both the northern and southern regions. All of these species exhibited positive prooxidant levels (ProO > 0) that could not be effectively counterbalanced by the antioxidant defense. In some cases, such as E. encrasicolus, S. sprattus, and M. galloprovincialis from both regions and T. mediterraneus from the southern region, antioxidant activity remained positive (Anti > 0) but insufficient to offset prooxidant processes. In contrast, suppressed antioxidant defense (Anti < 0) was observed in P. adspersus from both regions and in T. mediterraneus from the northern region, leading to a predominance of prooxidant processes and OS development. In P. saltatrix from the southern region, SOS > 0 was also calculated. Although prooxidant processes did not predominate (ProO < 0), the markedly low antioxidant capacity places this species in a vulnerable state, as even minor adverse environmental changes could impair its ability to mount an adequate protective response.
Table 9.
Normalized (z-transformed) pro-oxidant (Pro), antioxidant (Anti), and specific oxidative stress (SOS) indexes of the fish and invertebrate species.
The visualization of the SOS results (Figure 4) is presented in a coordinate system where the x-axis represents prooxidant processes, while the y-axis represents the levels of integrated antioxidant defense (including both enzymatic and non-enzymatic antioxidants). The resulting point reflects the overall OS status of the organism. Values close to zero indicate a balance between prooxidant and antioxidant processes.
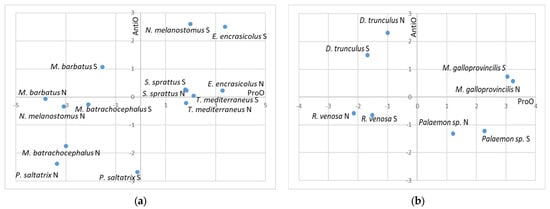
Figure 4.
Stress zones defined by values of the standardized oxidative stress indexes for prooxidant processes (ProO) and antioxidant processes (AntiO), and the distribution by zones of: (a) fish species; (b) invertebrate species. “N” refers to species from the northern region, while “S” refers to species from the southern region.
Organisms located in the upper-right quadrant are characterized by elevated prooxidant activity despite an activated antioxidant system, indicating that defensive mechanisms are insufficient to fully counteract oxidative pressure. In the upper-left quadrant, organisms exhibit enhanced antioxidant activity, likely induced by an external stressor, which effectively suppresses prooxidant processes, resulting in negative prooxidant values. Organisms positioned in the lower-left quadrant show generally low levels of both prooxidant and antioxidant activity, suggesting that they inhabit relatively stable and favorable environmental conditions that do not induce significant oxidative stress. In contrast, organisms in the lower-right quadrant experience the most severe oxidative stress conditions. In these cases, oxidative stress is strongly induced, leading to a suppression or exhaustion of antioxidant defenses. As a result, prooxidant processes dominate, either due to depletion of non-enzymatic antioxidants or inhibition of enzymatic antioxidant activity.
The obtained data indicated that among fish, T. mediterraneus from the northern region and P. adspersus from both regions are the most stressed species, as they fall within the lower right quadrant of the coordinate system, characterized by positive values for the prooxidant processes and negative values for the antioxidant defense. Additionally, S. sprattus from both regions, T. mediterraneus from the southern region, and E. encrasicolus from the northern region, as well as M. galloprovincialis from both regions, also exhibit compromised OS status, marked by very low antioxidant capacity and a predominance of prooxidant processes.
4. Discussion
The present study demonstrated substantial MP uptake by the studied key fish and invertebrate species in the Bulgarian sector of the Black Sea, with potential implications for organism health and overall ecosystem sustainability.
The obtained results present a detailed assessment of MPs taken up by key fish and invertebrate species, caught in the northern and southern Bulgarian Black Sea waters. All investigated specimens were proven to contain MPs. Our results showed MP counts ranging from 0.18 to 3.31 items/g across the different species and regions. Although many studies report MPs per individual rather than per gram, comparable research indicates that small pelagic fish such as anchovy and sprat typically contain around 1–4 particles per individual [58,59], which when recalculated, place the findings obtained herein within expected regional ranges. Differences in methodology, including digestion techniques and reporting units, make exact comparisons difficult, but in general, the data from our research align well with existing data in the literature.
One of the main results of this study is that the marine environmental and ecological conditions differ between the northern and southern regions of the Bulgarian Black Sea, with certain consequences for the marine organisms living there. We found statistically significant differences in the quantity of ingested MPs between only one fish species and only one invertebrate species from the northern and southern regions (Table 3), suggesting that microplastic pollution is widespread and relatively homogeneous across the study area. Notably, among the fish species, higher MP concentrations were observed in pelagic representatives compared to benthic ones. These findings are consistent with previous studies, which report that pelagic fish from the Bulgarian Black Sea, such as Belone belone (Linnaeus, 1761) and T. mediterraneus, exhibit a higher proportion of accumulated MPs [30]. This can be explained by the finding that small-sized MPs with low density are more abundant in the water column where pelagic fish feed [60]. The size of MPs can strongly influence their vertical distribution in the water column. Smaller particles (1–100 µm) show a more gradual decrease in abundance with depth, suggesting a more uniform distribution and longer residence time in the water column. In contrast, larger MPs (100–5000 µm) tend to accumulate within stratified layers, reflecting more limited vertical transport and faster settling [61]. Furthermore, fish species such as S. sprattus and E. encrasicolus are filter feeders or particulate feeders, increasing the likelihood of uptake of MPs that resemble plankton. In contrast, benthic species such as M. barbatus or N. melanostomus interact more with sediments, where MPs may be less bioavailable depending on local conditions. However, it should be noted that some studies have reported higher MP uptake in demersal fish, indicating that feeding strategy, habitat, and local contamination all influence the uptake [62,63,64].
In our study, the size-class distribution of MPs in the studied species showed a clear and widely reported pattern: particles <50 µm dominate across nearly all species and in both regions, often exceeding 70–95% of the total. This trend is consistent with previous observations in other fish from the Black Sea [30]. The current understanding is that smaller MPs are the most bioavailable and ecologically relevant fraction in marine ecosystems [59]. Larger plastic debris continuously breaks down into smaller particles through physical abrasion (waves, sediment friction), ultraviolet radiation, and biological degradation. As a result, the marine environment becomes progressively enriched in small MPs and even nanoplastics. Because of their small size, these particles remain suspended in the water column for longer periods and are more easily ingested by a wide range of marine organisms. Studies have shown that the abundance of MPs increases exponentially with decreasing particle size, meaning that the smallest fractions are numerically dominant in seawater and sediments [59]. In addition, smaller MPs are more easily ingested, both intentionally (as mistaken prey) and passively (via filtration), by a wider range of species. Pelagic fish such as S. sprattus and E. encrasicolus, studied herein, feed on plankton-sized particles, making them prone to ingesting small MPs (<50 µm) that resemble natural prey. The filter feeders like M. galloprovincialis and D. trunculus efficiently retain MPs due to their filtration mechanisms. Filtering large volumes of water that can retain particles from a few micrometers up to several hundred micrometers explains the more even distribution across size classes in the studied samples. In addition, smaller MPs also tend to persist longer in the digestive system, leading to their dominance in samples [65]. This prolonged retention in the digestive system can damage the mucosal lining and lead to translocation into tissues with subsequent pathological consequences.
Regarding MPs’ shapes, our results show a clear dominance of rounded particles across all species and both regions, often exceeding 90%, while fibers and irregular fragments occur only in low proportions or display species-specific variability. This predominance of rounded particles is consistent with the findings of Toschkova et al. [30], who reported that pellets were the most frequently isolated form in key fish species from the Bulgarian Black Sea, followed by irregular fragments, whereas fibers were the least abundant. However, many Black Sea surveys consistently identify fibers as the dominant MP shape in biota [19,21,22]. The degradation of plastic particles in the marine environment exposed to UV radiation, mechanical abrasion and chemical modifications progressively alters particle morphology, smoothing edges and transforming both fragments and fibers into more rounded forms over time [66]. Additionally, primary MPs such as industrial pellets and microbeads contribute directly to the abundance of rounded particles [58]. The observed predominance of rounded particles suggests a significant contribution of microbeads originating from personal care and cosmetic products, introduced into aquatic environments through wastewater effluents, especially in highly populated areas. Biologically, many organisms preferentially ingest rounded particles because they resemble natural prey and are easier to consume, while also being retained longer in the digestive tract compared to fibers. Together, these factors may lead to a dominance of rounded particles in biological samples [59].
The variability in MP shapes observed between species in this study may reflect differences in feeding behavior, habitat, and morphology. Species that exhibit a broader range of particle shapes, such as M. galloprovincialis, N. melanostomus, and M. barbatus, are typically non-selective feeders or interact with both the water column and sediments. These environments contain a more diverse mixture of MP shape types, including fibers from wastewater and irregular fragments from degraded plastics, resulting in higher variability in ingested shapes [67]. In contrast, pelagic species such as S. sprattus, E. encrasicolus, and P. saltatrix tend to feed on plankton in the water column, which biases ingestion toward small, rounded particles that resemble their natural prey. This leads to a much more uniform shape distribution dominated by rounded forms.
The polymer composition of MPs also exhibited spatial and species-specific variability. The results of the present study revealed a notable dominance of rubber and PA in many species, alongside variable contributions from common polymers such as PE, PP, and PET. Rubber-like particles in marine environments are commonly associated with tire wear particles, which are now recognized as a major source of MP pollution globally [68,69,70]. Urbanized coastal areas and regions influenced by river discharge (such as the Black Sea basin) tend to show elevated levels of rubber-derived particles [58,71]. In addition, marine sources such as fishing gear, boat coatings, and industrial materials may contribute to rubber inputs [69].
A high proportion of PA was observed in several species, notably in N. melanostomus and M. barbatus. Polyamide is mainly associated with synthetic textile fibers, indicating wastewater discharge as the primary source. These fibers are released during laundering and often pass through wastewater treatment systems, eventually reaching coastal waters. This pattern is consistent with studies identifying textile-derived microfibers as one of the most abundant MP polymer types in marine environments, especially in semi-enclosed basins such as the Black Sea [59]. The high proportion of PA in this study suggests a strong influence from urban wastewater and coastal population pressure.
Other polymers found reflect a mixture of consumer plastics and industrial materials [72,73]. For example, PE and PP, detected in species such as S. sprattus, E. encrasicolus, and D. trunculus, are among the most widely produced plastics globally and are commonly associated with packaging materials, bags, and containers. Polyethylene terephthalate, observed at high levels in some species (e.g., P. saltatrix, M. galloprovincialis), is typically derived from plastic bottles and textile fibers (PE). The presence of polymers such as PU, ABS, and POM indicates industrial sources, coatings, and engineered plastic products, further highlighting the diversity of anthropogenic inputs into the marine environment.
The PCA analyses of MP polymer type and size in the studied species suggested that there was no clear dominance of certain polymers associated with species diversity, lifestyle and occurrence. Our findings confirmed that MP accumulation in marine species depends on the habitat, lifestyle, food absorption, clearance rate, and physiological resilience of the species. It was established that spatial and taxonomic differences in MP uptake between the fish and invertebrates from the northern and southern regions were present. The differences in MPs by their polymer type ingested by the studied fish and invertebrates can largely be explained by high variability of feeding ecology and habitat. Microplastics detected in fish stomach contents generally reflect recent ingestion events. In pelagic species, MP exposure mainly originates from the water column and contaminated prey, whereas in benthic fish species it is more closely associated with sediments and sediment-related food resources. Soft tissues of filter-feeding invertebrates such as M. galloprovincialis can accumulate MPs directly from the surrounding environment through continuous filtration, while benthic invertebrates (e.g., D. trunculus) are more likely to be exposed to sediment-associated polymers. Consequently, pelagic fish may uptake more buoyant polymers such as PE and PP, whereas benthic organisms may accumulate larger and denser materials (as PET, PVC, and rubber), which tend to settle in bottom sediments [74,75]. This leads to differences in polymer composition between trophic groups. In addition, MPs ingested and retained in gills, particularly smaller particles, may potentially translocate across epithelial barriers into the blood stream and internal organs and tissues, which further contributes to the chemical diversity of MPs in organisms [76,77].
Variability in MP polymer types accumulated by the marine organisms from the northern and southern regions in this study most likely reflects local differences in pollution sources and hydrodynamics. The northern Bulgarian Black Sea region is strongly influenced by riverine inputs (e.g., the Danube), which transport a wide range of polymers from inland sources, including industrial and urban waste [32]. In contrast, the southern region may be more affected by local coastal activities, such as tourism, wastewater discharge, and maritime traffic, which can alter the relative contribution of polymers such as rubber and PA. For instance, there was an observed shift from PA dominance in D. trunculus from the northern region to PP in the clams of the southern region, as well as an increased contribution of rubber in R. venosa from the southern region (Table 6). The semi-enclosed nature of the Black Sea also promotes accumulation and patchy distribution of MPs, leading to site-specific polymer signatures [78,79,80].
What is new in the results of this study is the detailed assessment of the oxidative status of the studied species. Although the analyzed biomarkers provide valuable information regarding the physiological condition and stress status of the investigated organisms, they should not be interpreted as being specific indicators of MP exposure alone. Biomarker responses are generally associated with a broad range of environmental stressors, including chemical contamination, nutritional status, temperature fluctuations, hypoxia, and other anthropogenic pressures. Therefore, the observed biological responses in the present study likely reflect the combined influence of multiple environmental factors, among which MPs may represent one contributing stressor. The detected interspecific differences in OS reflect variations in the balance between prooxidant generation and antioxidant defense capacity, which is strongly influenced by ecological characteristics (including exposure to environmental pollutants), as well as species-specific biochemical and physiological traits. Species occupying pelagic niches, such as E. encrasicolus, S. sprattus, and T. mediterraneus, are generally more prone to oxidative imbalance. At least in part, the OS observed in these species may be related to MP exposure in their habitat (the water column) and MP uptake. In particular, species exhibiting higher MP uptake in the southern coastal area (e.g., M. barbatus, S. sprattus, E. encrasicolus, D. trunculus, and P. adspersus) also tended to show increased OS biomarkers and stronger shifts in antioxidant balance, as reflected by the elevated lipid peroxidation and altered enzymatic activities. This pattern is in line with the hypothesis that MPs can induce OS through mechanisms such as ROS overproduction, inflammation, and disruption of cellular redox homeostasis [81,82,83,84]. Similarly, controlled laboratory exposure experiments in fish and invertebrates have demonstrated that MP exposure can increase oxidative damage markers and activate the antioxidant defense, including LPO and SOD responses [27,47,85,86]. Under in situ conditions, responses of OS biomarkers reflect the combined influence of multiple environmental and methodological factors; however, when integrated with contamination data and ecological context, they can provide valuable early warning information on physiological disturbances also associated with MP exposure levels. Future studies should incorporate the assessment of additional environmental stressors and physicochemical parameters to better disentangle the relative contribution of MPs from other co-occurring stress factors. In addition, controlled laboratory exposure experiments would provide a more robust framework for evaluating the specific effects of MPs on the target organisms. Such experiments could help clarify dose–response relationships, uptake mechanisms, and the extent to which MPs alone contribute to the observed physiological and biochemical alterations.
Spatial differences in stress responses further indicated the importance of local environmental conditions. Variability between northern and southern regions likely reflects differences in pollution sources, hydrodynamics, and coastal pressures such as wastewater discharge and maritime activity. These factors influence not only MP abundance but also the mixture of co-occurring contaminants, which together determine the overall oxidative stress challenge faced by organisms. At present, there are no universally accepted, species-specific threshold values for the oxidative stress biomarkers that could clearly distinguish between normal physiological variation and stress-induced responses. In this respect, the integrative SOS index presented more concrete and comparable results on the stress-response reaction of the studied species. This index summarizes the overall redox status by integrating pro-oxidant and antioxidant responses into a single metric, thereby providing a more comprehensive indication of physiological state than individual biomarkers alone. Species such as T. mediterraneus, which showed higher MPs levels in the northern region, displayed a less consistent relationship with SOS patterns, suggesting species-specific differences in exposure routes, feeding ecology, and physiological resilience [87]. Overall, the co-occurrence of elevated MP uptake and increased SOS values in several southern region populations strongly supports the view that MPs may act as an additional environmental stressor contributing to oxidative imbalance in marine organisms, although the magnitude of this effect is clearly species dependent [27,47,88]. In addition, polymer type and particle characteristics may influence toxicity, as some polymers carry additives and pollutants (heavy metals and pathogens) that enhance oxidative processes [9,40,41]. Smaller MPs are more reactive due to their higher surface area, increasing their potential to induce excess ROS generation and cellular stress as reflected by SOS. Consequently, species accumulating more small MPs may be more prone to oxidative imbalance [59].
5. Conclusions
The present study demonstrates substantial MP uptake by key fish and invertebrate species from the Bulgarian Black Sea coast, providing important region-specific baseline data on MP contamination in marine biota. Bioaccumulation of MPs showed pronounced spatial and species-specific variability, influenced by habitat conditions and its degree of contamination, lifestyle, feeding behavior, and physiological characteristics. Higher MP accumulation was observed in selected pelagic fish species and in the wedge clam D. trunculus, while overall levels in the invertebrates remained relatively low. The identified MPs were dominated by small (<50 µm), rounded particles across all studied taxa. Polymer composition of MPs, found in the studied biota, differed between regions, with southern sites dominated by PA, rubber, PU, and PP, whereas northern sites exhibited a more heterogeneous polymer profile, indicating different sources and exposure pathways. Oxidative stress responses in both fish and invertebrates varied considerably among species and regions, indicating that marine organisms are exposed to multiple environmental stressors that may induce cellular imbalance and physiological stress, with MPs representing one of the possible contributing factors with subsequent ecological consequences. The observed biomarker responses should therefore be interpreted as indicators of general environmental stress rather than as specific responses to MP exposure alone. In this context, the SOS index has the potential to serve as a useful integrated tool for assessing the overall physiological status and stress burden in marine organisms under complex environmental conditions, including MP contamination and possible accumulation in organisms. Further field-based and controlled laboratory exposure experiments are needed to improve knowledge on uptake and accumulation of MPs in marine organisms and better elucidate the mechanisms underlying MP-induced oxidative stress and adaptive responses. Such studies would also enable a more accurate assessment of the specific contribution of MPs to the observed biological effects under conditions involving multiple environmental stressors. At the same time, they would contribute to validating the applicability of the SOS index as a reliable integrated biomarker for long-term ecological risk assessment and marine environmental monitoring.
Author Contributions
Conceptualization, N.C., A.A. and P.I.; methodology, A.A., E.T. and M.A. ; validation, N.C. and A.A.; formal analysis, A.A., E.T., M.A., S.K. and S.M.; investigation, A.A., E.T., M.A., S.K. and S.M.; resources, E.S., V.R., D.D. and Y.R.; data curation, A.A. and N.C.; writing—original draft preparation, A.A.; writing—review and editing, N.C., A.A. and P.I.; visualization, A.A.; supervision, N.C., A.A., P.I., K.S. and V.D.; project administration, A.A. and P.I.; funding acquisition, A.A. and P.I. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Scientific Fund, Sofia, Bulgaria, grant number Grant KP-06-N-61/10.
Institutional Review Board Statement
No laboratory experimental animals were involved in this study, and approval from the Local Ethics Committee was not required. Species sampling was conducted under the authorization of the Executive Agency for Fisheries and Aquaculture, Ministry of Agriculture and Food, Bulgaria, following the permit for the capture of fish and other aquatic organisms for scientific research purposes, No. 32/21.06.2023 and No. 36/24.07.2024, under contract No. D-156 dated 13 March 2023.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ABS | Acrylonitrile butadiene styrene |
| AChE | Acetylcholine esterase |
| AntiO | Antioxidant score |
| CAT | Catalase |
| GPx | Glutathione peroxidase |
| GR | Glutathione reductase |
| GSH | Total glutathione |
| GST | Glutathione-S-transferase |
| K | Fulton’s condition index |
| LPO | Lipid peroxidation |
| MDA | Malondialdehyde |
| MPs | Microplastics |
| nd | Not detected |
| OS | Oxidative stress |
| PA | Polyamide |
| PCA | Principal component analysis |
| PE | Polyethylene |
| PET | Polyethylene terephthalate |
| PMMA | Polymethyl methacrylate |
| POM | Polyoxymethylene |
| PP | Polypropylene |
| ProO | Pro-oxidant score |
| PS | Polystyrene |
| PTFE | Polytetrafluoroethylene |
| PU | Polyurethane |
| PVC | Polyvinyl chloride |
| ROS | Reactive oxygen species |
| SOD | Superoxide dismutase |
| SOS | Specific oxidative stress index |
Appendix A

Figure A1.
Analysis of MPs in E. encrasicolus: (a) filter image; (b) polymer identification analysis of MPs.


Figure A2.
Analysis of MPs in D. trunculus: (a) filter image; (b) polymer identification analysis of MPs.


Figure A3.
Analysis of MPs in P. adspersus: (a) filter image; (b) polymer identification analysis of MPs.


Figure A4.
Analysis of MPs in P. saltatrix: (a) filter image; (b) polymer identification analysis of MPs.

References
- FAO. The State of World Fisheries and Aquaculture 2022; Food and Agriculture Organization of the United Nations: Rome, Italy, 2022. [Google Scholar]
- UNCTAD; United Nations Conference on Trade and Development. Review of Maritime Transport 2023; United Nations: Geneva, Switzerland, 2023. [Google Scholar]
- Intergovernmental Panel on Climate Change. Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Barbier, E.B.; Hacker, S.D.; Kennedy, C.; Koch, E.W.; Stier, A.C.; Silliman, B.R. The value of estuarine and coastal ecosystem services. Ecol. Monogr. 2011, 81, 169–193. [Google Scholar] [CrossRef]
- Cunha, J.; Cardona, F.S.; Bio, A.; Ramos, S. Importance of protection service against erosion and storm events provided by coastal ecosystems under climate change scenarios. Front. Mar. Sci. 2021, 8, 726145. [Google Scholar] [CrossRef]
- United Nations Environment Programme. From Pollution to Solution: A Global Assessment of Marine Litter and Plastic Pollution; United Nations Environment Programme: Nairobi, Kenya, 2021. [Google Scholar]
- Marcharla, E.; Vinayagam, S.; Gnanasekaran, L.; Soto-Moscoso, M.; Chen, W.-H.; Thanigaivel, S.; Ganesan, S. Microplastics in marine ecosystems: A comprehensive review of biological and ecological implications and its mitigation approach using nanotechnology for the sustainable environment. Environ. Res. 2024, 256, 119181. [Google Scholar] [CrossRef]
- Acarer Arat, S. Microplastic pollution in marine ecosystems: Sources, impacts, and stakeholder-based solutions. Turk. J. Biol. 2025, 49, 421–440. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, O.A.; Jamal, M.T.; Almalki, H.S.; Alzahrani, A.H.; Alatawi, A.S.; Haque, M.F. Microplastic pollution in the marine environment: Sources, impacts, and degradation. J. Adv. Vet. Anim. Res. 2025, 12, 260–279. [Google Scholar] [CrossRef]
- Miller, M.E.; Hamann, M.; Kroon, F.J. Bioaccumulation and biomagnification of microplastics in marine organisms: A review and meta-analysis of current data. PLoS ONE 2020, 15, e0240792. [Google Scholar] [CrossRef]
- Nikhil, V.G.; Abisha, C.; Raghavan, R.; Ali, A.P.H.; Ranjeet, K.; Varghese, G.K. Bioaccumulation and trophic transfer of microplastics in oceanic food webs. Mar. Pollut. Bull. 2026, 224, 119113. [Google Scholar] [CrossRef]
- European Commission. Developing Optimal and Open Research Support for the Black Sea (DOORS); Horizon 2020 Project No. 101000518; CORDIS Project Page; European Commission: Brussels, Belgium, 2020; Available online: https://cordis.europa.eu/projects (accessed on 10 May 2026).
- European Environment Agency (EEA). Contaminants in Europe’s Seas: Moving Towards a Clean, Non-Toxic Marine Environment; EEA Report No. 25/2018; Publications Office of the European Union: Luxembourg, 2019; Available online: https://www.eea.europa.eu/en (accessed on 10 May 2026).
- National Institute for Marine Research and Development “Grigore Antipa” (NIMRD). ANEMONE Project: Assessing the Vulnerability of the Black Sea Marine Ecosystem to Human Pressures. Available online: https://www.anemoneproject.eu/?page_id=246 (accessed on 10 May 2026).
- BSC. State of the Environment of the Black Sea (2009–2014/2015); Black Sea Commission: Istanbul, Türkiye, 2019. [Google Scholar]
- Pokazeev, K.; Sovga, E.; Chaplina, T. Pollution in the Black Sea: Observations About the Ocean’s Pollution; Springer: Cham, Switzerland, 2021. [Google Scholar] [CrossRef]
- Strokal, M.; Strokal, V.; Kroeze, C. The future of the Black Sea: More pollution in over half of the rivers. Ambio 2023, 52, 339–356. [Google Scholar] [CrossRef]
- Rangel-Buitrago, N.; Rizzo, A.; Neal, W.; Mastronuzzi, G. Sediment pollution in coastal and marine environments. Mar. Pollut. Bull. 2023, 192, 115023. [Google Scholar] [CrossRef]
- Aytan, U.; Esensoy, F.B.; Senturk, Y.; Arifoğlu, E.; Karaoğlu, K.; Ceylan, Y.; Valente, A. Plastic occurrence in commercial fish species of the Black Sea. Turk. J. Fish. Aquat. Sci. 2022, 22, TRJFAS20504. [Google Scholar] [CrossRef]
- Bilgin Fıçıcılar, B.; Aydın, M.; Korkmaz, K. Microplastic contamination in commercial fish from the Central Black Sea. J. Ecol. Eng. 2025, 26, 108–119. [Google Scholar] [CrossRef]
- Gedik, K.; Gozler, A.M. Hallmarking microplastics of sediments and Chamelea gallina inhabiting southwestern Black Sea: A hypothetical look at consumption risks. Mar. Pollut. Bull. 2022, 174, 113252. [Google Scholar] [CrossRef]
- Ciucă, A.-M.; Stoica, E.; Barbeș, L. First Report of Microplastic Ingestion and Bioaccumulation in Commercially Valuable European Anchovies (Engraulis encrasicolus, Linnaeus, 1758) from the Romanian Black Sea Coast. J. Mar. Sci. Eng. 2025, 13, 394. [Google Scholar] [CrossRef]
- Berov, D.; Klayn, S. Microplastics and floating litter pollution in Bulgarian Black Sea coastal waters. Mar. Pollut. Bull. 2020, 156, 111225. [Google Scholar] [CrossRef]
- Georgieva, S.K.; Peteva, Z.V.; Stancheva, M.D. Evaluation of abundance of microplastics in the Bulgarian coastal waters. BioRisk 2023, 20, 59–69. [Google Scholar] [CrossRef]
- Bobchev, N.; Berov, D.; Klayn, S.; Karamfilov, V. High microplastic pollution in marine sediments associated with urbanised areas along the SW Bulgarian Black Sea coast. Mar. Pollut. Bull. 2024, 209, 117150117150. [Google Scholar] [CrossRef]
- Alexandrova, A.; Ignatova-Ivanova, T.V.; Bachvarova, D.G.; Toschkova, S.G.; Doichinov, A.H.; Ibryamova, S.F.; Chipev, N.H. Pilot Screening and Assessment of Microplastic Bioaccumulation in Wedge Clams Donax trunculus Linnaeus, 1758 (Bivalvia) from the Bulgarian Black Sea Coast. Acta Zool. Bulg. 2022, 74, 569–577. [Google Scholar]
- Alexandrova, A.; Mihova, S.; Tsvetanova, E.; Andreeva, M.; Pramatarov, G.; Petrov, G.; Chipev, N.; Doncheva, V.; Stefanova, K.; Grandova, M.; et al. Microplastic Bioaccumulation and Oxidative Stress in Key Species of the Bulgarian Black Sea: Ecosystem Risk Early Warning. Microplastics 2025, 4, 50. [Google Scholar] [CrossRef]
- Ibryamova, S.; Toschkova, S.; Bachvarova, D.; Lyatif, A.; Stanachkova, E.; Ivanov, R.; Natchev, N.; Ignatova-Ivanova, T. Assessment of the bioaccumulation of microplastics in the Black Sea mussel Mytilus galloprovincialis L., 1819. J. IMAB 2022, 28, 4676–4682. [Google Scholar] [CrossRef]
- Zlateva, I.; Raykov, V.; Alexandrova, A.; Ivanova, P.; Chipev, N.; Stefanova, K.; Dzhembekova, N.; Doncheva, V.; Slabakova, V.; Stefanova, E.; et al. Effects of anthropogenic and environmental stressors on the current status of red mullet (Mullus barbatus L., 1758) populations inhabiting the Bulgarian Black Sea waters. Nat. Conserv. 2023, 54, 55–79. [Google Scholar] [CrossRef]
- Toschkova, S.; Ibryamova, S.; Bachvarova, D.C.; Koynova, T.; Stanachkova, E.; Ivanov, R.; Natchev, N.; Ignatova-Ivanova, T. The assessment of the bioaccumulation of microplastics in key fish species from the Bulgarian aquatory of the Black Sea. BioRisk 2024, 22, 17–31. [Google Scholar] [CrossRef]
- Pramatarov, G.I.; Tsvetanova, E.R.; Ilinkin, V.M.; Andreeva, M.N.; Alexandrova, A.V.; Chipev, N.H. Effects of Microplastics and Metal Pollution on Bivalves from the Bulgarian Black Sea Sublittoral, with Comments on their Adaptive Capacity. Acta Zool. Bulg. 2025, 77, 107–119. [Google Scholar] [CrossRef]
- Zlateva, I.; Ricker, M.; Slabakova, V.; Slavova, K.; Doncheva, V.; Staneva, J.; Stanev, E.; Popov, I.; Gramcianinov, C.; Raykov, V. Analysis of terrestrial and riverine sources of plastic litter contributing to plastic pollution in the Western Black Sea using a Lagrangian particle tracking model. Mar. Pollut. Bull. 2024, 209, 117108. [Google Scholar] [CrossRef] [PubMed]
- Syversen, T.; Lilleng, G. Microplastics derived from commercial fishing activities. In Environmental Sciences; IntechOpen: London, UK, 2023. [Google Scholar] [CrossRef]
- Cincinelli, A.; Martellini, T.; Guerranti, C.; Scopetani, C.; Chelazzi, D.; Giarrizzo, T. A Potpourri of Microplastics in the Sea Surface and Water Column of the Mediterranean Sea. Trends Anal. Chem. 2019, 110, 321–326. [Google Scholar] [CrossRef]
- Pojar, I.; Stănică, A.; Stock, F.; Kochleus, C.; Günther, R.; Gayer, G.; Lechthaler, S.; Lenz, R.; Labrenz, M.; Laforsch, C. Sedimentary microplastic concentrations from the Romanian Danube River to the Black Sea. Sci. Rep. 2021, 11, 2000. [Google Scholar] [CrossRef] [PubMed]
- Jemaa, S.; Mahfouz, C.; Kazour, M.; Lteif, M.; Hassoun, A.E.R.; Ghsoub, M.; Amara, R.; Khalaf, G.; Fakhri, M. Floating Marine Litter in Eastern Mediterranean From Macro to Microplastics: The Lebanese Coastal Area as a Case Study. Front. Environ. Sci. 2021, 9, 699343. [Google Scholar] [CrossRef]
- Narloch, I.; Gackowska, A.; Wejnerowska, G. Microplastic in the Baltic Sea: A review of distribution processes, sources, analysis methods and regulatory policies. Environ. Pollut. 2022, 315, 120453. [Google Scholar] [CrossRef]
- Savuca, A.; Nicoara, M.N.; Faggio, C. Comprehensive Review regarding the Profile of the Microplastic Pollution in the Coastal Area of the Black Sea. Sustainability 2022, 14, 14376. [Google Scholar] [CrossRef]
- Glevitzky, M.; Dumitrel, G.-A.; Rusu, G.I.; Toneva, D.; Vergiev, S.; Corcheş, M.-T.; Pană, A.-M.; Popa, M. Microplastic Pollution on the Beaches of the Black Sea in Romania and Bulgaria. Appl. Sci. 2025, 15, 4751. [Google Scholar] [CrossRef]
- Liu, S.; Huang, J.; Zhang, W.; Shi, L.; Yi, K.; Yu, H.; Zhang, C.; Li, S.; Li, J. Microplastics as a vehicle of heavy metals in aquatic environments: A review of adsorption factors, mechanisms, and biological effects. J. Environ. Manag. 2022, 302, 113995. [Google Scholar] [CrossRef]
- Bidegain, G.; Sestelo, M.; Luque, P.L.; Uriarte, I.; Iriarte, A.; Villate, F. The Role of Microplastics in Marine Pathogen Transmission: Retrospective Regression Analysis, Experimental Design, and Disease Modelling. J. Mar. Sci. Eng. 2022, 10, 1837. [Google Scholar] [CrossRef]
- He, S.; Jia, M.; Xiang, Y.; Song, B.; Xiong, W.; Cao, J.; Peng, H.; Yang, Y.; Wang, W.; Yang, Z.; et al. Biofilm on Microplastics in Aqueous Environment: Physicochemical Properties and Environmental Implications. J. Hazard. Mater. 2021, 424, 127286. [Google Scholar] [CrossRef]
- Hu, M.; Palić, D. Micro- and nano-plastics activation of oxidative and inflammatory adverse outcome pathways. Redox Biol. 2020, 37, 101620. [Google Scholar] [CrossRef] [PubMed]
- Steinberg, C.E.W. Stress Ecology: Environmental Stress as Ecological Driving Force and Key Player in Evolution; Springer: Dordrecht, The Netherlands, 2012. [Google Scholar]
- Lemos, M.F.L. Biomarker Studies in Stress Biology: From the Gene to Population, from the Organism to the Application. Biology 2021, 10, 1340. [Google Scholar] [CrossRef]
- Parolini, M.; De Felice, B.; Gazzotti, S.; Annunziata, L.; Sugni, M.; Bacchetta, R.; Ortenzi, M.A. Oxidative stress-related effects induced by micronized polyethylene terephthalate microparticles in the Manila clam. J. Toxicol. Environ. Health A 2020, 83, 168–179. [Google Scholar] [CrossRef]
- Kadac-Czapska, K.; Ośko, J.; Knez, E.; Grembecka, M. Microplastics and oxidative stress—Current problems and prospects. Antioxidants 2024, 13, 579. [Google Scholar] [CrossRef]
- Das, A. The emerging role of microplastics in systemic toxicity: Involvement of reactive oxygen species (ROS). Sci. Total Environ. 2023, 895, 165076. [Google Scholar] [CrossRef]
- Kelly, E.R.M.; Trujillo, J.E.; Setiawan, A.; Pether, S.; Burritt, D.; Allan, B.J.M. Investigating the metabolic and oxidative stress induced by biofouled microplastics exposure in Seriola lalandi (yellowtail kingfish). Mar. Pollut. Bull. 2024, 203, 116438. [Google Scholar] [CrossRef]
- Ren, H.-Y.; Ma, H.-C.; He, R.-P.; Gao, C.-C.; Wen, B.; Gao, J.-Z.; Chen, Z.-Z. Microplastics Induce Structural Color Deterioration in Fish Poecilia reticulata Mediated by Oxidative Stress. Fishes 2025, 10, 382. [Google Scholar] [CrossRef]
- Hook, S.E.; Gallagher, E.P.; Batley, G.E. The Role of Biomarkers in the Assessment of Aquatic Ecosystem Health. Integr. Environ. Assess. Manag. 2014, 10, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Spedicato, M.T.; Massutí, E.; Mérigot, B.; Tserpes, G.; Jadaud, A.; Relini, G. The MEDITS trawl survey specifications in an ecosystem approach to fishery management. Sci. Mar. 2019, 83, 9–20. [Google Scholar] [CrossRef]
- Nash, R.D.; Valencia, A.; Geffen, A.J. The Origin of Fulton’s Condition Factor: Setting the Record Straight. Fisheries 2006, 31, 236–238. [Google Scholar]
- Bessa, F.; Frias, J.; Kögel, T.; Lusher, A.; Andrade, J.; Antunes, J.; Sobral, P.; Pagter, E.; Nash, R.; O’Connor, I.; et al. Harmonized Protocol for Monitoring Microplastics in Biota; JPI Oceans: Brussels, Belgium, 2019. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Yakimov, L.; Tsvetanova, E.; Georgieva, A.; Petrov, L.; Alexandrova, A. Assessment of the oxidative status of Black Sea mussels (Mytilus galloprovincialis Lamarck, 1819) from Bulgarian coastal areas with the introduction of a specific oxidative stress index. J. Environ. Prot. Ecol. 2018, 19, 1614–1622. [Google Scholar]
- Aytan, Ü.; Koca, Y.Ş.; Pasli, S.; Güven, O.; Ceylan, Y.; Başaran, B. Microplastics in commercial fish and their habitats in the important fishing ground of the Black Sea: Characteristics, concentration, and risk assessment. Mar. Pollut. Bull. 2025, 221, 118434. [Google Scholar] [CrossRef]
- Manolaki, S.M.; Dimitriou, P.D.; Lampa, M.; Karakassis, I.; Papageorgiou, N. Microplastic Concentration in Mediterranean Commercial Fish: A Systematic Review. Microplastics 2025, 4, 100. [Google Scholar] [CrossRef]
- Chevalier, C.; Vandenberghe, M.; Pagano, M.; Pellet, I.; Pinazo, C.; Tesán Onrubia, J.A.; Guilloux, L.; Carlotti, F. Investigation of dynamic change in microplastics vertical distribution patterns: The seasonal effect on vertical distribution. Mar. Pollut. Bull. 2023, 189, 114674. [Google Scholar] [CrossRef]
- Zhao, S.; Kvale, K.F.; Zhu, L.; Zettler, E.R.; Egger, M.; Mincer, T.J.; Amaral-Zettler, L.A.; Lebreton, L.; Niemann, H.; Nakajima, R.; et al. The distribution of subsurface microplastics in the ocean. Nature 2025, 641, 51–61. [Google Scholar] [CrossRef]
- Rathore, C.; Saha, M.; Desai, A.; Gupta, P.; Naik, A. Driving aspects of microplastic uptake: Influence in the bentho-pelagic ecosystem and its associated ecological risks along the coast of Goa, India. Environ. Res. 2025, 278, 121589. [Google Scholar] [CrossRef]
- Sui, Y.; Zhang, Y.; Chen, S.; Shi, Y.; Zou, J.; Mohamed, M.; Qin, K.; Yu, Y.; Liu, Z.; Zheng, L.; et al. Microplastic ingestion by fish with different feeding habits from different habitats in a typical semi-enclosed Haizhou Bay. J. Ocean Univ. China 2025, 24, 343–354. [Google Scholar] [CrossRef]
- Kalaiselvan, K.; Padmavathy, P.; Velu, R.; Jeyashakila, R. Microplastics accumulation in pelagic and benthic species along the Thoothukudi coast, South Tamil Nadu, India. Mar. Pollut. Bull. 2023, 189, 114735. [Google Scholar] [CrossRef] [PubMed]
- Wright, S.L.; Thompson, R.C.; Galloway, T.S. The physical impacts of microplastics on marine organisms: A review. Environ. Pollut. 2013, 178, 483–492. [Google Scholar] [CrossRef]
- Hidalgo-Ruz, V.; Gutow, L.; Thompson, R.C.; Thiel, M. Microplastics in the Marine Environment: A Review of the Methods Used for Identification and Quantification. Environ. Sci. Technol. 2012, 46, 3060–3075. [Google Scholar] [CrossRef]
- Filimon, A.; Nenciu, M.I.; Ionescu, D.T.; Suciu, L.; Ciobanu, A.A.; Radu, C.M.; Lazăr, L.; Vasile, D.; Bodeanu, S.; Cadar, O.; et al. Microplastic contamination in Black Sea cetaceans. Animals 2024, 14, 886. [Google Scholar] [CrossRef] [PubMed]
- Kole, P.J.; Löhr, A.J.; Van Belleghem, F.G.A.J.; Ragas, A.M.J. Wear and tear of tyres: A stealthy source of microplastics in the environment. Int. J. Environ. Res. Public Health 2017, 14, 1265. [Google Scholar] [CrossRef]
- Sun, W.; Wang, B.; Ouyang, W.; Liu, Z.; Zhang, H. Tire wear particles in aquatic environments: A systematic review of sources, detection, distribution, and toxicological impacts. Ecotoxicol. Environ. Saf. 2025, 305, 119236. [Google Scholar] [CrossRef]
- Zhao, T.; Zhang, Y.; Song, Q.; Meng, Q.; Zhou, S.; Cong, J. Tire and road wear particles in aquatic organisms: A review of source, properties, exposure routes, and biological effects. Aquat. Toxicol. 2024, 273, 107010. [Google Scholar] [CrossRef]
- Wagner, S.; Hüffer, T.; Klöckner, P.; Wehrhahn, M.; Hofmann, T.; Reemtsma, T. Tire wear particles in the aquatic environment—A review on generation, analysis, occurrence, fate, and effects. Water Res. 2018, 139, 83–100. [Google Scholar] [CrossRef]
- Geyer, R.; Jambeck, J.R.; Law, K.L. Production, use, and fate of all plastics ever made. Sci. Adv. 2017, 3, e1700782. [Google Scholar] [CrossRef]
- Lewanska, M.; Barczynska, R. Microplastics from Food Packaging: Polymer Degradation Pathways, Environmental Distribution, and Effects on the Human Gastrointestinal Tract. Polymers 2025, 17, 2923. [Google Scholar] [CrossRef] [PubMed]
- Erni-Cassola, G.; Zadjelovic, V.; Gibson, M.I.; Christie-Oleza, J.A. Distribution of Plastic Polymer Types in the Marine Environment: A Meta-Analysis. J. Hazard. Mater. 2019, 369, 691–698. [Google Scholar] [CrossRef]
- Kowalski, N.; Reichardt, A.M.; Waniek, J.J. Sinking Rates of Microplastics and Potential Implications of Their Alteration by Physical, Biological, and Chemical Factors. Mar. Pollut. Bull. 2016, 109, 310–319. [Google Scholar] [CrossRef]
- Ullah, I.; Chen, H.; Wang, J.; Kaiser, H.; Basher, A.A.; Li, J.; Zhu, X. Impacts of Microplastics on the Early Life Stages of Fish: Sources, Mechanisms, Ecological Consequences, and Mitigation Strategies. Toxics 2025, 14, 27. [Google Scholar] [CrossRef]
- Guerrera, M.C.; Aragona, M.; Porcino, C.; Fazio, F.; Laurà, R.; Levanti, M.; Montalbano, G.; Germanà, G.; Abbate, F.; Germanà, A. Micro and Nano Plastics Distribution in Fish as Model Organisms: Histopathology, Blood Response and Bioaccumulation in Different Organs. Appl. Sci. 2021, 11, 5768. [Google Scholar] [CrossRef]
- Miladinova, S.; Macias, D.; Stips, A.; Garcia-Gorriz, E. Identifying distribution and accumulation patterns of floating marine debris in the Black Sea. Mar. Pollut. Bull. 2020, 153, 110964. [Google Scholar] [CrossRef]
- Stanev, E.V.; Ricker, M. The fate of marine litter in semi-enclosed seas: A case study of the Black Sea. Front. Mar. Sci. 2019, 6, 660. [Google Scholar] [CrossRef]
- Terzi, Y.; Özcan, A.; Yılmaz, F.; Çelik, A.; Kaya, S.; Demir, V.; Yücel, N.; Aydın, M. A comparative study of microplastic contamination and ecological risk in beach and bottom sediments along the southern Black Sea coast. Reg. Stud. Mar. Sci. 2026, 94, 104766. [Google Scholar] [CrossRef]
- Solomando, A.; Capó, X.; Alomar, C.; Álvarez, E.; Compa, M.; Valencia, J.M.; Pinya, S.; Deudero, S.; Sureda, A. Long-term exposure to microplastics induces oxidative stress and a pro-inflammatory response in the gut of Sparus aurata (Linnaeus, 1758). Environ. Pollut. 2020, 266, 115295. [Google Scholar] [CrossRef]
- De Sales-Ribeiro, C.; Brito-Casillas, Y.; Fernandez, A.; Caballero, M.J. An end to the controversy over the microscopic detection and effects of pristine microplastics in fish organs. Sci. Rep. 2020, 10, 12434. [Google Scholar] [CrossRef]
- Subaramaniyam , U.; Allimuthu, R.S.; Vappu, S.; Ramalingam, D.; Balan, R.; Paital, B.; Panda, N.; Rath, P.K.; Ramalingam, N.; Sahoo, D.K. Effects of microplastics, pesticides and nanomaterials on fish health, oxidative stress and antioxidant defense mechanisms. Front. Physiol. 2023, 14, 1217666. [Google Scholar]
- Jeong, C.-B.; Kang, H.-M.; Lee, M.-C.; Kim, D.-H.; Han, J.; Hwang, D.-S.; Souissi, S.; Lee, S.-J.; Shin, K.-H.; Park, H.G.; et al. Adverse Effects of Microplastics and Oxidative Stress-Induced MAPK/Nrf2 Pathway-Mediated Defense Mechanisms in the Marine Copepod Paracyclopina Nana. Sci. Rep. 2017, 7, 41323. [Google Scholar] [CrossRef]
- Ghosh, T. Microplastics bioaccumulation in fish: Its potential toxic effects on hematology, immune response, neurotoxicity, oxidative stress, growth, and reproductive dysfunction. Toxicol. Rep. 2024, 14, 101854. [Google Scholar] [CrossRef]
- Wang, J.; Wu, F.; Dong, S.; Wang, X.; Ai, S.; Liu, Z.; Wang, X. Meta-analysis of the effects of microplastics on fish: Insights into growth, survival, reproduction, oxidative stress, and gut microbiota diversity. Water Res. 2024, 267, 122493. [Google Scholar] [CrossRef]
- Galloway, T.S.; Cole, M.; Lewis, C. Interactions of microplastic debris throughout the marine ecosystem. Nat. Ecol. Evol. 2017, 1, 0116. [Google Scholar] [CrossRef]
- Auta, H.S.; Emenike, C.U.; Fauziah, S.H. Distribution and importance of microplastics in the marine environment: A review of the sources, fate, effects, and potential solutions. Environ. Int. 2017, 102, 165–176. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.