Screening for the Brewing Ability of Different Non-Saccharomyces Yeasts

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Nutrient Media
2.2. Yeast Sample Preparation
2.3. Phenolic Off-Flavor Test (POF Test)
2.4. Sensory Odor Test
2.5. Yeast Propagation and Wort Analysis
2.6. Fermentation Trials
2.7. Analysis of Produced Beers
2.8. Sensory Evaluation
3. Results
3.1. Overall Results
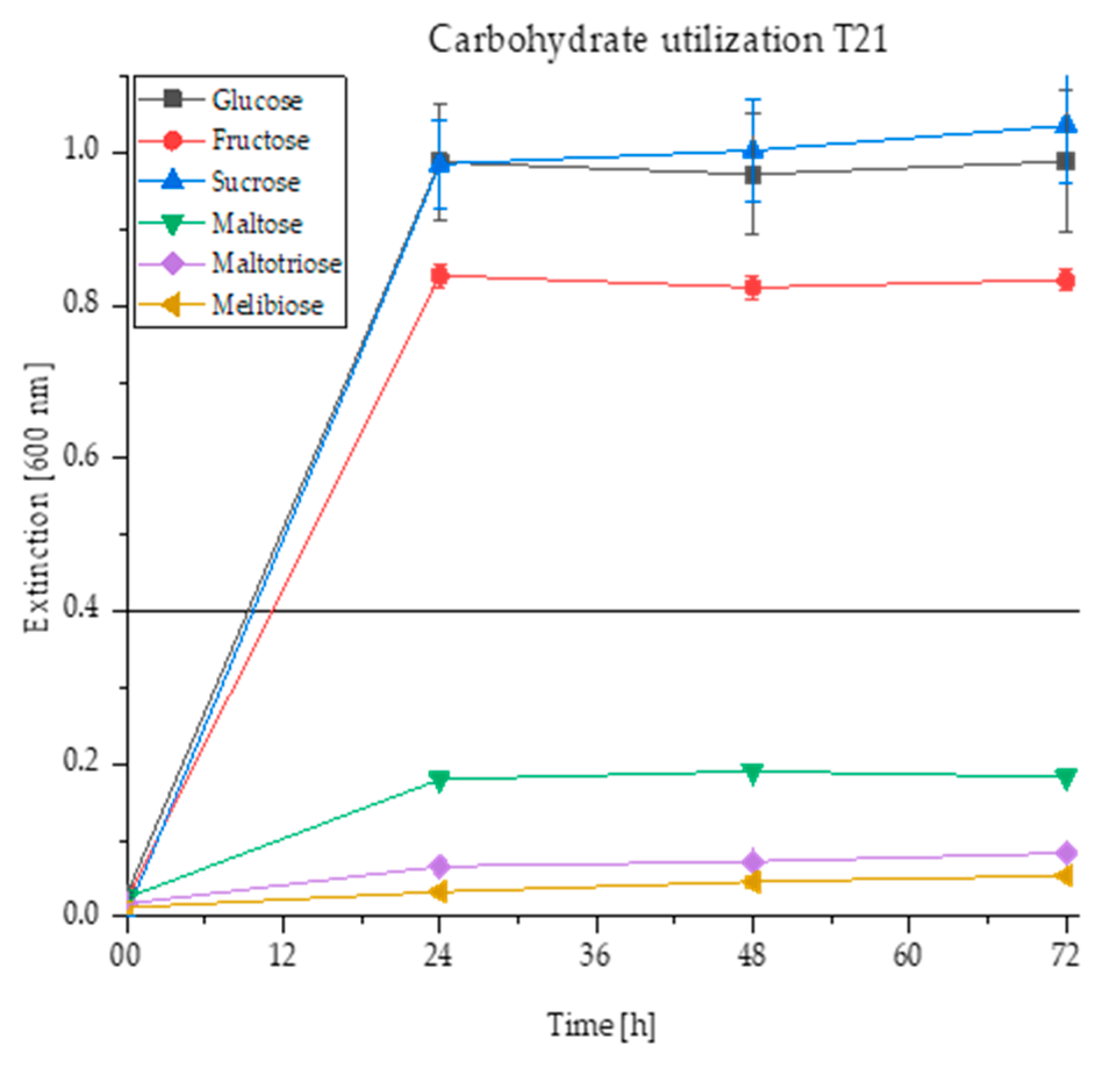
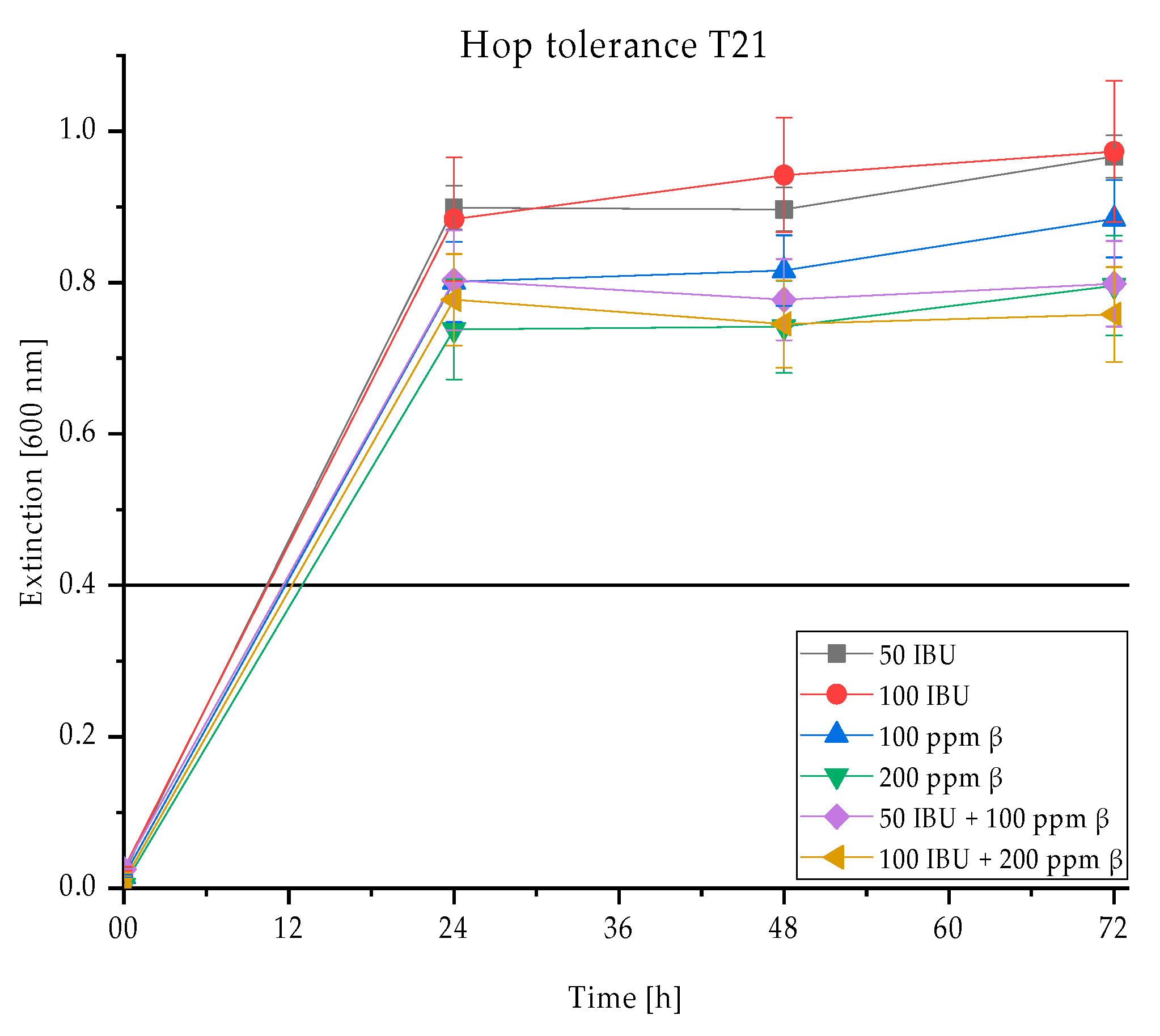
3.2. Exemplary Evaluation of Carbohydrate Utilisation, Ethanol and Hop Resistance
3.2.1. Carbohydrate Utilization
3.2.2. Ethanol Tolerance
3.2.3. Hop Tolerance
3.3. Results of the Sensory Odor Test
3.4. Fermentation Trials
3.4.1. Fermentation Process
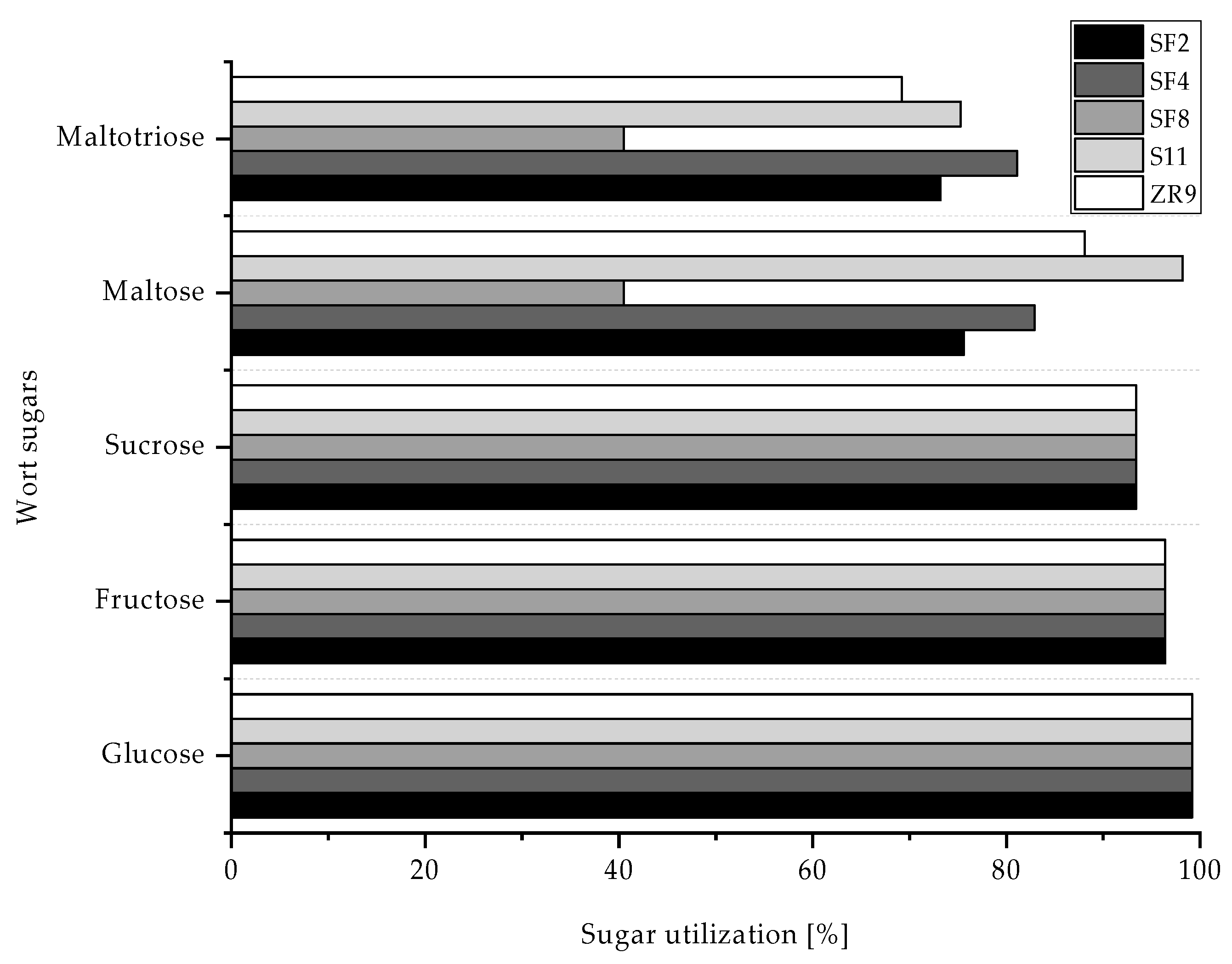
3.4.2. Wort Sugar Utilization
3.4.3. Volatile Compounds in Final Beers
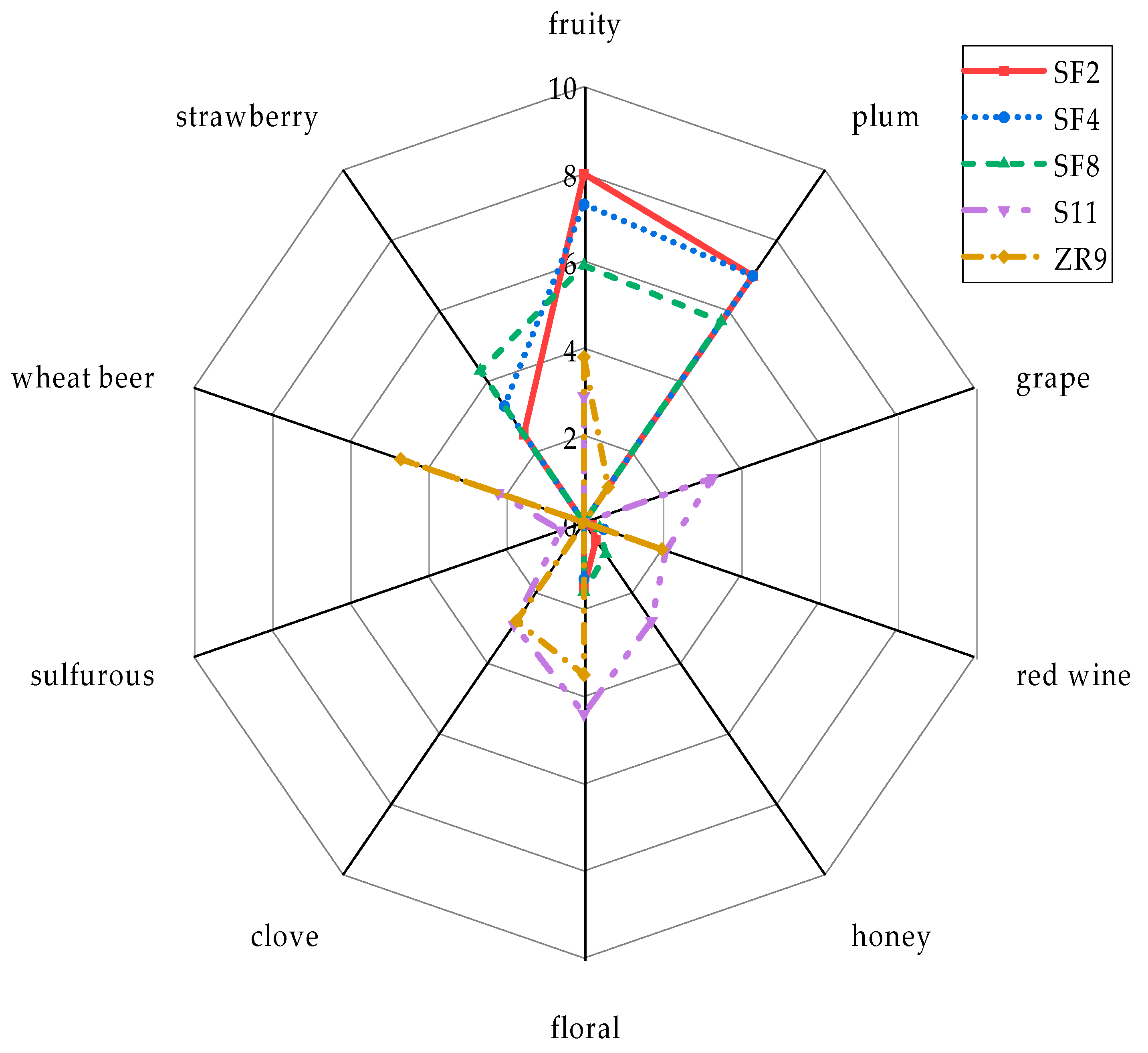
3.4.4. Aroma Profiles of Final Beers
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Piškur, J.; Rozpędowska, E.; Polakova, S.; Merico, A.; Compagno, C. How did Saccharomyces evolve to become a good brewer? Trends Genet. 2006, 22, 183–186. [Google Scholar] [CrossRef] [PubMed]
- Claussen, N.H. Eine Methode zur Anwendung von Hansens Reinzuchtsystem bei der Herstellung von englischen gelagerten Biersorten. Wochenschr. Brau 1904, 1904, 370–383. [Google Scholar]
- De Estela-Escalante, W.; Rosales-Mendoza, S.; Moscosa-Santillán, M.; González-Ramírez, J.E. Evaluation of the fermentative potential of Candida zemplinina yeasts for craft beer fermentation. J. Inst. Brew. 2016, 122, 530–535. [Google Scholar] [CrossRef]
- Michel, M.; Meier-Dörnberg, T.; Jacob, F.; Methner, F.-J.; Wagner, R.S.; Hutzler, M. Pure non-Saccharomyces starter cultures for beer fermentation with a focus on secondary metabolites and practical applications. J. Inst. Brew. 2016, 122, 569–587. [Google Scholar] [CrossRef]
- Basso, R.F.; Alcarde, A.R.; Portugal, C.B. Could non-Saccharomyces yeasts contribute on innovative brewing fermentations? Food Res. Int. 2016, 86, 112–120. [Google Scholar] [CrossRef]
- Michel, M.; Meier-Dörnberg, T.; Zarnkow, M.; Jacob, F.; Hutzler, M. Screening for the Brewing Ability of Non-Saccharomyces Yeast and Optimization of Fermentation Performance of One Torulaspora Delbrueckii Strain Found Suitable for Beer Production. In Proceedings of the World Brewing Congress, Denver, CO, USA, 13–17 August 2016. [Google Scholar]
- Ravasio, D.; Carlin, S.; Boekhout, T.; Groenewald, M.; Vrhovsek, U.; Walther, A.; Wendland, J. Adding Flavor to Beverages with Non-Conventional Yeasts. Fermentation 2018, 4, 15. [Google Scholar] [CrossRef]
- Narziß, L.; Back, W.; Miedaner, H.; Lustig, S. Untersuchung zur Beeinflussung der Geschmacksstabilität durch Variation technologischer Parameter bei der Bierherstellung. Monatsschrift für Brauwissenschaft 1999, 1999, 192–206. [Google Scholar]
- Stewart, G.G. The Horace Brown Medal Lecture: Forty Years of Brewing Research. J. Inst. Brew. 2009, 115, 3–29. [Google Scholar] [CrossRef]
- Vidgren, V.; Multanen, J.-P.; Ruohonen, L.; Londesborough, J. The temperature dependence of maltose transport in ale and lager strains of brewer’s yeast. FEMS Yeast Res. 2010, 10, 402–411. [Google Scholar] [CrossRef]
- Wickerham, L.J. A simple technique for the detection of melibiose-fermenting yeasts1. J. Bacteriol. 1943, 46, 501–505. [Google Scholar]
- Srinivasan, V.; Goldberg, D.; Haas, G.J. Contributions to the Antimicrobial Spectrum of Hop Constituents. Econ. Bot. 2004, 58, S230–S238. [Google Scholar] [CrossRef]
- Hazelwood, L.A.; Walsh, M.C.; Pronk, J.T.; Daran, J.-M. Involvement of vacuolar sequestration and active transport in tolerance of Saccharomyces cerevisiae to hop iso-alpha-acids. Appl. Environ. Microbiol. 2010, 76, 318–328. [Google Scholar] [CrossRef] [PubMed]
- Bamforth, C.W. Wort Composition and Beer Quality. In Brewing Yeast Fermentation Performance, 2nd ed.; Smart, K., Ed.; Blackwell Science: Oxford, UK; Malden, MA, USA, 2003; pp. 75–85. ISBN 9780470696040. [Google Scholar]
- Ding, J.; Huang, X.; Zhang, L.; Zhao, N.; Yang, D.; Zhang, K. Tolerance and stress response to ethanol in the yeast Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2009, 85, 253. [Google Scholar] [CrossRef]
- Narziss, L.; Back, W.; Gastl, M.; Zarnkow, M. Abriss der Bierbrauerei, 8th ed.; John Wiley & Sons Incorporated: Newark, NJ, USA, 2017; ISBN 9783527340361. [Google Scholar]
- Shinohara, T.; Kubodera, S.; Yanagida, F. Distribution of phenolic yeasts and production of phenolic off-flavors in wine fermentation. J. Biosci. Bioeng. 2000, 90, 90–97. [Google Scholar] [CrossRef]
- Montanari, L.; Perretti, G.; Natella, F.; Guidi, A.; Fantozzi, P. Organic and Phenolic Acids in Beer. LWT 1999, 32, 535–539. [Google Scholar] [CrossRef]
- Schwarz, K.J.; Stübner, R.; Methner, F.-J. Formation of styrene dependent on fermentation management during wheat beer production. Food Chem. 2012, 134, 2121–2125. [Google Scholar] [CrossRef]
- De Arruda Moura Pietrowski, G.; dos Santos, C.M.E.; Sauer, E.; Wosiacki, G.; Nogueira, A. Influence of fermentation with Hanseniaspora sp. yeast on the volatile profile of fermented apple. J. Agric. Food Chem. 2012, 60, 9815–9821. [Google Scholar] [CrossRef]
- Benito, S.; Palomero, F.; Morata, A.; Calderón, F.; Suárez-Lepe, J.A. New applications for Schizosaccharomyces pombe in the alcoholic fermentation of red wines. Int. J. Food Sci. Technol. 2012, 47, 2101–2108. [Google Scholar] [CrossRef]
- Amaya-Delgado, L.; Herrera-López, E.J.; Arrizon, J.; Arellano-Plaza, M.; Gschaedler, A. Performance evaluation of Pichia kluyveri, Kluyveromyces marxianus and Saccharomyces cerevisiae in industrial tequila fermentation. World J. Microbiol. Biotechnol. 2013, 29, 875–881. [Google Scholar] [CrossRef]
- Steensels, J.; Verstrepen, K.J. Taming wild yeast: Potential of conventional and nonconventional yeasts in industrial fermentations. Annu. Rev. Microbiol. 2014, 68, 61–80. [Google Scholar] [CrossRef]
- Hiralal, L.; Olaniran, A.O.; Pillay, B. Aroma-active ester profile of ale beer produced under different fermentation and nutritional conditions. J. Biosci. Bioeng. 2014, 117, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Verstrepen, K.J.; Derdelinckx, G.; Delvaux, F.R. Esters in beer-part 1: The fermentation process: More than ethanol formation. Cerevisia 2003, 28, 41–49. [Google Scholar]
- Vanderhaegen, B.; Neven, H.; Coghe, S.; Verstrepen, K.J.; Derdelinckx, G.; Verachtert, H. Bioflavoring and beer refermentation. Appl. Microbiol. Biotechnol. 2003, 62, 140–150. [Google Scholar] [CrossRef]
- Verstrepen, K.J.; Derdelinckx, G.; Dufour, J.-P.; Winderickx, J.; Thevelein, J.M.; Pretorius, I.S.; Delvaux, F.R. Flavor-active esters: Adding fruitiness to beer. J. Biosci. Bioeng. 2003, 96, 110–118. [Google Scholar] [CrossRef]
- Olaniran, A.O.; Hiralal, L.; Mokoena, M.P.; Pillay, B. Flavour-active volatile compounds in beer: Production, regulation and control. J. Inst. Brew. 2017, 123, 13–23. [Google Scholar] [CrossRef]
- Meilgaard, M.C. Flavor Chemistry of Beer: Part II: Flavor and Threshold of 239 Aroma Volatiles. MBAA Tech. Tech. Quart. Master. Brew. Assoc. Am. 1975, 12, 151–168. [Google Scholar]
- Pires, E.J.; Teixeira, J.A.; Brányik, T.; Vicente, A.A. Yeast: The soul of beer’s aroma—A review of flavour-active esters and higher alcohols produced by the brewing yeast. Appl. Microbiol. Biotechnol. 2014, 98, 1937–1949. [Google Scholar] [CrossRef]
- Sannino, C.; Mezzasoma, A.; Buzzini, P.; Turchetti, B. Non-conventional Yeasts for Producing Alternative Beers. In Non-conventional Yeasts: From Basic Research to Application; Sibirny, A., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 361–388. ISBN 978-3-030-21109-7. [Google Scholar]
- Olšovská, J.; Vrzal, T.; Štěrba, K.; Slabý, M.; Kubizniaková, P.; Čejka, P. The chemical profiling of fatty acids during the brewing process. J. Sci. Food Agric. 2019, 99, 1772–1779. [Google Scholar] [CrossRef]
- Wahyono, A.; Kang, W.-W.; Park, H.-D. Characterization and application of Torulaspora delbrueckii JK08 and Pichia anomala JK04 as baker’s yeasts. J. Food Nutr. Res. 2015, 54. [Google Scholar]
- Esslinger, H.M. Handbook of Brewing: Processes, Technology, Markets; John Wiley & Sons: Hoboken, NJ, USA, 2009; ISBN 3527623493. [Google Scholar]
- Sakamoto, K.; Konings, W.N. Beer spoilage bacteria and hop resistance. Int. J. Food Microbiol. 2003, 89, 105–124. [Google Scholar] [CrossRef]
- Kurtzman, C.P. (Ed.) The Yeasts. A taxonomic study, 5th ed.; Elsevier: Amsterdam, The Netherland, 2011; ISBN 9780123847072. [Google Scholar]
- Owens, J.D. Indigenous Fermented Foods of Southeast Asia; CRC Press: Boca Raton, FL, USA, 2014; ISBN 0429105916. [Google Scholar]
- Lee, S.M.; Jung, J.H.; Seo, J.-A.; Kim, Y.-S. Bioformation of Volatile and Nonvolatile Metabolites by Saccharomycopsis fibuligera KJJ81 Cultivated under Different Conditions-Carbon Sources and Cultivation Times. Molecules 2018, 23, 2762. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Procopio, S.; Becker, T. Flavor impacts of glycerol in the processing of yeast fermented beverages: A review. J. Food Sci. Technol. 2015, 52, 7588–7598. [Google Scholar] [CrossRef] [PubMed]
- Duong, C.T.; Strack, L.; Futschik, M.; Katou, Y.; Nakao, Y.; Fujimura, T.; Shirahige, K.; Kodama, Y.; Nevoigt, E. Identification of Sc-type ILV6 as a target to reduce diacetyl formation in lager brewers’ yeast. Metab. Eng. 2011, 13, 638–647. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, M. Entstehung und Beeinflussung qualitätsbestimmender Aromastoffe bei der Herstellung von Weißbier; Technische Universität München: Munich, Germany, 2005. [Google Scholar]
- Boulton, C.; Quain, D. Brewing Yeast and Fermentation; John Wiley & Sons: Hoboken, NJ, USA, 2008; ISBN 0470999403. [Google Scholar]
- Saerens, S.; Swiegers, J.H. Production of Low-Alcohol or Alcohol-Free beer with Pichia Kluyveri Yeast Strains. Google Patents EP2964742A2, 2016. [Google Scholar]
- Lindner, P. Schizosaccharomyces pombe n. sp., ein neuer Gährungserreger. Wochenschrift für Brauerei 1893, 1298–1300. [Google Scholar]
- Sefa-Dedeh, S.; Sanni, A.I.; Tetteh, G.; Sakyi-Dawson, E. Yeasts in the traditional brewing of pito in Ghana. World J. Microbiol. Biotechnol. 1999, 15, 593–597. [Google Scholar] [CrossRef]
- King, A.; Richard Dickinson, J. Biotransformation of monoterpene alcohols by Saccharomyces cerevisiae, Torulaspora delbrueckii and Kluyveromyces lactis. Yeast 2000, 16, 499–506. [Google Scholar] [CrossRef]
- Canonico, L.; Galli, E.; Ciani, E.; Comitini, F.; Ciani, M. Exploitation of Three Non-Conventional Yeast Species in the Brewing Process. Microorganisms 2019, 7, 11. [Google Scholar] [CrossRef]
- De Francesco, G.; Turchetti, B.; Sileoni, V.; Marconi, O.; Perretti, G. Screening of new strains of Saccharomycodes ludwigii and Zygosaccharomyces rouxii to produce low-alcohol beer. J. Inst. Brew. 2015, 121, 113–121. [Google Scholar] [CrossRef]
- Meilgaard, M.C.; Reid, D.S.; Wyborski, K.A. Reference standards for beer flavor terminology system. J. Am. Soc. Brew. Chem. 1982, 40, 119–128. [Google Scholar] [CrossRef]
- Andorrá, I.; Berradre, M.; Mas, A.; Esteve-Zarzoso, B.; Guillamón, J.M. Effect of mixed culture fermentations on yeast populations and aroma profile. LWT 2012, 49, 8–13. [Google Scholar] [CrossRef]
- Varela, C.; Barker, A.; Tran, T.; Borneman, A.; Curtin, C. Sensory profile and volatile aroma composition of reduced alcohol Merlot wines fermented with Metschnikowia pulcherrima and Saccharomyces uvarum. Int. J. Food Microbiol. 2017, 252, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Anfang, N.; Brajkovich, M.; Goddard, M.R. Co-fermentation with Pichia kluyveri increases varietal thiol concentrations in Sauvignon Blanc. Aust. J. Grape Wine Res. 2009, 15, 1–8. [Google Scholar] [CrossRef]
- Díaz, C.; Molina, A.M.; Nähring, J.; Fischer, R. Characterization and dynamic behavior of wild yeast during spontaneous wine fermentation in steel tanks and amphorae. Biomed Res. Int. 2013, 2013, 540465. [Google Scholar] [CrossRef] [PubMed]
- Varela, C.; Borneman, A.R. Yeasts found in vineyards and wineries. Yeast 2017, 34, 111–128. [Google Scholar] [CrossRef]
- Hutzler, M.; Riedl, R.; Koob, J.; Jacob, F. Fermentation and spoilage yeasts and their relevance for the beverage industry-a review. BrewingScience-Monatsschrift Für Brauwissenschaft 2012, 65, 33–50. [Google Scholar]
- Coton, E.; Coton, M.; Levert, D.; Casaregola, S.; Sohier, D. Yeast ecology in French cider and black olive natural fermentations. Int. J. Food Microbiol. 2006, 108, 130–135. [Google Scholar] [CrossRef]
- Ye, M.; Yue, T.; Yuan, Y. Effects of sequential mixed cultures of Wickerhamomyces anomalus and Saccharomyces cerevisiae on apple cider fermentation. FEMS Yeast Res. 2014, 14, 873–882. [Google Scholar] [CrossRef]
- Cañas, P.M.I.; García-Romero, E.; Manso, J.M.H.; Fernández-González, M. Influence of sequential inoculation of Wickerhamomyces anomalus and Saccharomyces cerevisiae in the quality of red wines. Eur. Food Res. Technol. 2014, 239, 279–286. [Google Scholar] [CrossRef]
- De Benedictis, M.; Bleve, G.; Grieco, F.; Tristezza, M.; Tufariello, M. An optimized procedure for the enological selection of non-Saccharomyces starter cultures. Antonie Van Leeuwenhoek 2011, 99, 189–200. [Google Scholar] [CrossRef]
- Loira, I.; Morata, A.; Comuzzo, P.; Callejo, M.J.; González, C.; Calderón, F.; Suárez-Lepe, J.A. Use of Schizosaccharomyces pombe and Torulaspora delbrueckii strains in mixed and sequential fermentations to improve red wine sensory quality. Food Res. Int. 2015, 76, 325–333. [Google Scholar] [CrossRef]
- Lee, P.-R.; Ong, Y.-L.; Yu, B.; Curran, P.; Liu, S.-Q. Evolution of volatile compounds in papaya wine fermented with three Williopsis saturnus yeasts. Int. J. Food Sci. Technol. 2010, 45, 2032–2041. [Google Scholar] [CrossRef]
- Yilmaztekin, M.; Erten, H.; Cabaroglu, T. Production of Isoamyl Acetate from Sugar Beet Molasses by Williopsis saturnus var. saturnus. J. Inst. Brew. 2008, 114, 34–38. [Google Scholar] [CrossRef]
- Erten, H.; Tanguler, H. Influence of Williopsis saturnus yeasts in combination with Saccharomyces cerevisiae on wine fermentation. Lett. Appl. Microbiol. 2010, 50, 474–479. [Google Scholar] [CrossRef]
- Son, E.Y.; Lee, S.M.; Kim, M.; Seo, J.-A.; Kim, Y.-S. Comparison of volatile and non-volatile metabolites in rice wine fermented by Koji inoculated with Saccharomycopsis fibuligera and Aspergillus oryzae. Food Res. Int. 2018, 109, 596–605. [Google Scholar] [CrossRef]
- Medeiros, A.B.P.; Pandey, A.; Christen, P.; Fontoura, P.S.G.; de Freitas, R.J.S.; Soccol, C.R. Aroma compounds produced by Kluyveromyces marxianus in solid state fermentation on a packed bed column bioreactor. World J. Microbiol. Biotechnol. 2001, 17, 767–771. [Google Scholar] [CrossRef]
- Fabre, C.E.; Duviau, V.J.; Blanc, P.J.; Goma, G. Identification of volatile flavour compounds obtained in culture of Kluyveromyces marxianus. Biotechnol. Lett. 1995, 17, 1207–1212. [Google Scholar] [CrossRef]
- Jiang, J. Identification of flavour volatile compounds produced by Kluyveromyces lactis. Biotechnol. Tech. 1993, 7, 863–866. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Number | Strain Abbr. | Species | Strain Number | Strain Abbr. | Species |
|---|---|---|---|---|---|
| YH837A-3D4 | CM1 | Cyberlindnera misumaiensis | YH725B-1F6 | M10 | Metschnikowia sp. |
| YH824B-1I7 | CM2 | Cyberlindnera misumaiensis | YH729A-1G1 | M11 | Metschnikowia sp. |
| YH837B-3D5 | CM3 | Cyberlindnera misumaiensis | YH730A-1G2 | M12 | Metschnikowia sp. |
| YHMH22AA-3H1 | CSa1 | Cyberlindnera saturnus | YH730B-1G3 | M13 | Metschnikowia sp. |
| WYSC 29 | DH1 | Debaryomyces hansenii | YH737A-1G4 | M14 | Metschnikowia sp. |
| WYSC 1664 | DH3 | Debaryomyces hansenii | YHMH50A-3B4 | NH2 | Nakazawaea holstii |
| DSMZ70244 | DH4 | Debaryomyces hansenii | YHMH50-3C5 | NH3 | Nakazawaea holstii |
| PI BB 151/BLQ 208 | DH8 | Debaryomyces hansenii | YH607A-1D1 | NH5 | Nakazawaea holstii |
| YH811A-3F4 | DH10 | Debaryomyces hansenii | YHAK1A-3I1 | PK1 | Pichia kluyveri |
| YH813A-3F5 | DH11 | Debaryomyces hansenii | PI S 4; Lu 36 | SF1 | Saccharomycopsis fibuligera |
| YH817-3F6 | DH12 | Debaryomyces hansenii | PI S 5; Lu 29/45 | SF2 | Saccharomycopsis fibuligera |
| YH832A-1I9 | DH13 | Debaryomyces hansenii | PI S 6; Lu 27 | SF3 | Saccharomycopsis fibuligera |
| YH883A-3E8 | DH14 | Debaryomyces hansenii | PI S 7; Lu 26 | SF4 | Saccharomycopsis fibuligera |
| YH885A-3E9 | DH15 | Debaryomyces hansenii | PI S 9; Lu 7/41 | SF5 | Saccharomycopsis fibuligera |
| YH887A-3E10 | DH16 | Debaryomyces hansenii | PI S 10; Lu 35 | SF6 | Saccharomycopsis fibuligera |
| YH888A-3F1 | DH17 | Debaryomyces hansenii | PI S 12; Lu 32 | SF8 | Saccharomycopsis fibuligera |
| CBS 5074 | H1 | Hanseniaspora uvarum | PI S 15; Lu 6/48 | SF10 | Saccharomycopsis fibuligera |
| CBS 2585 | H2 | Hanseniaspora uvarum | PI BB 132 | S2 | Schizosaccharomyces pombe |
| CBS 314T | H3 | Hanseniaspora uvarum | DSMBZ 70576 | S3 | Schizosaccharomyces pombe |
| CBS 1517 | H4 | Hanseniaspora uvarum | BLQ TUM S4 | S4 | Schizosaccharomyces pombe |
| RIBM A4 | H5 | Hanseniaspora uvarum | TUM G5S | S5 | Schizosaccharomyces pombe |
| RIBM A7 | H6 | Hanseniaspora uvarum | TUM G6S | S6 | Schizosaccharomyces pombe |
| RIBM A10 | H7 | Hanseniaspora uvarum | TUM G7S | S7 | Schizosaccharomyces pombe |
| RIBM A12 | H8 | Hanseniaspora uvarum | TUM G8S | S8 | Schizosaccharomyces pombe |
| RIBM A15 | H9 | Hanseniaspora uvarum | TUM G9S | S9 | Schizosaccharomyces pombe |
| RIBM A16 | H10 | Hanseniaspora uvarum | TUM G10S | S10 | Schizosaccharomyces pombe |
| YHMH56B-3C8 | KS2 | Kazachstania servazzii | TUM G11S | S11 | Schizosaccharomyces pombe |
| YHMH47B-3C4 | KS3 | Kazachstania servazzii | YH725A-1F5 | T21 | Torulaspora delbrueckii |
| BLQ TUM K1 | K1 | Kluyveromyces lactis | YH739A-1G6 | T22 | Torulaspora delbrueckii |
| TUM G2K | K2 | Kluyveromyces lactis | YH739B-1G7 | T23 | Torulaspora delbrueckii |
| TUM G3K | K3 | Kluyveromyces lactis | YH740B-1H1 | T24 | Torulaspora delbrueckii |
| TUM G4K | K4 | Kluyveromyces lactis | YH832D-3D3 | T25 | Torulaspora delbrueckii |
| TUM G5K | K5 | Kluyveromyces lactis | YH824A-1I6 | T26 | Torulaspora delbrueckii |
| TUM G6K | K6 | Kluyveromyces lactis | YH837D-3D7 | T27 | Torulaspora delbrueckii |
| TUM G7K | K7 | Kluyveromyces lactis | YHYF2-1D6 | T28 | Torulaspora delbrueckii |
| TUM G8K | K8 | Kluyveromyces lactis | YHMS8-1E2 | TM1 | Torulaspora microellipsoides |
| TUM G9K | K9 | Kluyveromyces lactis | YH601A-1C5 | WA1 | Wickerhamomyces anomalus |
| TUM G1KM | Km 1 | Kluyveromyces marxianus | YH601C-1C6 | WA2 | Wickerhamomyces anomalus |
| TUM G2KM | Km 2 | Kluyveromyces marxianus | YHB23-1D7 | WA3 | Wickerhamomyces anomalus |
| TUM G3KM | Km 3 | Kluyveromyces marxianus | YHDC1211B-3H3 | WA4 | Wickerhamomyces anomalus |
| TUM G4KM | Km 4 | Kluyveromyces marxianus | TUM WA25 | WA25 | Wickerhamomyces anomalus |
| TUM G5KM | Km 5 | Kluyveromyces marxianus | WYSC 137 | ZR1 | Zygosaccharomyces rouxii |
| TUM G6KM | Km 6 | Kluyveromyces marxianus | WYSC 84 | ZR2 | Zygosaccharomyces rouxii |
| TUM G7KM | Km 7 | Kluyveromyces marxianus | WYSC 82 | ZR3 | Zygosaccharomyces rouxii |
| TUM G8KM | Km 8 | Kluyveromyces marxianus | WYSC 83 | ZR4 | Zygosaccharomyces rouxii |
| TUM G9KM | Km 9 | Kluyveromyces marxianus | WYSC/G20 | ZR5 | Zygosaccharomyces rouxii |
| TUM G10KM | Km 10 | Kluyveromyces marxianus | WYSC/G 1673 | ZR6 | Zygosaccharomyces rouxii |
| CBS 3082T | LK1 | Lachancea kluyveri | WYSC/G 1998 | ZR7 | Zygosaccharomyces rouxii |
| YHMH44C-3C3 | M3 | Metschnikowia sp. | WYSC/G 2005 | ZR8 | Zygosaccharomyces rouxii |
| YHMS-1D4 | M4 | Metschnikowia sp. | WYSC/G 2091 | ZR9 | Zygosaccharomyces rouxii |
| YH750A-1H2 | M5 | Metschnikowia sp. | WYSC/G 2093 | ZR10 | Zygosaccharomyces rouxii |
| YH713B-1E5 | M6 | Metschnikowia sp. | WYSC/G 2142 | ZR11 | Zygosaccharomyces rouxii |
| YH721A-1F2 | M7 | Metschnikowia sp. | WYSC/G 2274 | ZR13 | Zygosaccharomyces rouxii |
| YH724A-1F3 | M8 | Metschnikowia sp. | WYSC/G 2325 | ZR14 | Zygosaccharomyces rouxii |
| YH724B-1F4 | M9 | Metschnikowia sp. | YHMH55A-3B10 | ZF1 | Zygotorulaspora florentina |
| Nutrient/Inhibiting Media | Supplier | Concentrations of Nutrient/Inhibiting Media | Yeast Carbon Base |
|---|---|---|---|
| D(+)-Glucose, anhydrous | Carl Roth, Karlsruhe, Germany | 10 g/L | 0 g/L |
| D(-)-Fructose BioChemica | AppliChem, Darmstadt, Germany | 10 g/L | 0 g/L |
| D(+)-Sucrose ≥ 99.0% | Merck, Darmstadt, Germany | 20 g/L | 0 g/L |
| D(+)-Maltose monohydrate 95% | Alfa Aesar, Karlsruhe, Germany | 55 g/L | 0 g/L |
| D-Maltotriose 98% | Alfa Aesar, Karlsruhe, Germany | 20 g/L | 0 g/L |
| D(+)-Melibiose ≥ 99.0% | Merck, Darmstadt, Germany | 10 g/L | 0 g/L |
| Ethanol 1% (v/v), undenatured | Serva, Heidelberg, Germany | 1% (v/v) | 23.3 g/L |
| Ethanol 2% (v/v), undenatured | Serva, Heidelberg, Germany | 2% (v/v) | 23.3 g/L |
| Ethanol 5% (v/v), undenatured | Serva, Heidelberg, Germany | 5% (v/v) | 23.3 g/L |
| Ethanol 8% (v/v), undenatured | Serva, Heidelberg, Germany | 8% (v/v) | 23.3 g/L |
| Ethanol 10% (v/v), undenatured | Serva, Heidelberg, Germany | 10% (v/v) | 23.3 g/L |
| IBU 50 (iso-α-acid), isomerized hop extract 30% | Hopsteiner, Mainburg, Germany | 3.3 g/L | 23.3 g/L |
| IBU 100 (iso-α-acid), isomerized hop extract 30% | Hopsteiner, Mainburg, Germany | 6.6 g/L | 23.3 g/L |
| β1: β-acid 100 ppm, beta-rich hop extract 40% | Hopsteiner, Mainburg, Germany | 0.1 g/L | 23.3 g/L |
| β2: β-acid 200 ppm, beta-rich hop extract 40% | Hopsteiner, Mainburg, Germany | 0.2 g/L | 23.3 g/L |
| βmix1: α- + β-stock-mix (50 IBU + 100 ppm β-acids) | Hopsteiner, Mainburg, Germany | 3.3 g/L, 0.1 g/L | 23.3 g/L |
| βmix2: α- + β-stock-mix (100 IBU + 200 ppm β-acids) | Hopsteiner, Mainburg, Germany | 6.6 g/L, 0.2 g/L | 23.3 g/L |
| Sugar Composition in g/L | |
|---|---|
| Fructose | 2.8 |
| Glucose | 11.9 |
| Sucrose | 1.5 |
| Maltose | 58.2 |
| Maltotriose | 12.7 |
| Analysis | MEBAK No. | Device |
|---|---|---|
| Sugar composition | MEBAK II 3.2.2.1.2 | HPLC UltiMate 3000 (Thermo Fisher Scientific, Waltham, MA, USA) |
| Original wort, ethanol content, extract content | MEBAK WBBM 2.9.6.3 | Bending vibration and NIR spectroscopy, Alcolyzer Plus with DMA 5000 X sample 122 (Anton-Paar GmbH, Ostfildern, Germany) |
| pH value | MEBAK WBBM 2.13 | pH meter with pH electrode, ProfiLine pH3210 pH meter (Xylem Inc., New York, NY, USA) |
| Fatty esters, 2-Phenylethanol | MEBAK WBBM 2.23.6 | GC-FID Clarus 580 (Perkin Elmer, Waltham, MA, USA), Column: 50m 0.32mm Phenomenex FFAP, 0.25 μm |
| Acetaldehyde, Ethylacetate, higher alcohols (n-Propanol, i-Butanol, Amylalcohols) | MEBAK WBBM 2.21.1 | GC-FID Clarus 580, Turbo Matrix 40, Head Space (Perkin Elmer, Waltham, MA, USA), Column: INNOWAX cross-linked polyethylene glycol, 60m × 0.32mm 0.5 μm |
| Diacetyl, 2,3-Pentandione | MEBAK WBBM 2.21.5.1 | GC-FID Clarus 580, Turbo Matrix 40, Head Space (Perkin Elmer, Waltham, MA, USA), Column: INNOWAX cross-linked polyethylene glycol, 60m × 0.32mm 0.5 μm |
| Strain Abbr. | Evaluation Odor 7 °P | Evaluation Odor 12 °P | Strain Abbr. | Evaluation Odor 7 °P | Evaluation Odor 12 °P |
|---|---|---|---|---|---|
| CM1 | isoamyl acetate | isoamyl acetate | M10 | neutral, yeasty | yeasty |
| CM2 | isoamyl acetate | honey, sweet | M11 | neutral, yeasty | yeasty |
| CM3 | fruity | yeasty | M12 | neutral, yeasty | yeasty |
| CSa1 | isoamyl acetate | isoamyl acetate | M13 | sour, yeasty | yeasty |
| DH1 | yeasty, lychee | fruity, banana | M14 | neutral, yeasty | neutral, yeasty |
| DH3 | fruity, pineapple | yeasty, strong | NH2 | fruity | overripe banana |
| DH4 | clove-like, acidic | clove-like, acidic, earthy | NH3 | pale, fruity | yeasty, fruity |
| DH8 | fruity | rice wine, fruity | NH5 | yeasty, diacetyl | fruity |
| DH10 | cardboard | cardboard, fruity | PK1 | yeasty, isoamyl acetate | yeasty, isoamyl acetate |
| DH11 | neutral | yeasty, fruity | SF1 | fruity | fruity, plum |
| DH12 | neutral | neutral | SF2 | fruity | fruity, plum |
| DH13 | neutral | yeasty, diacetyl | SF3 | fruity | fruity, plum, honey |
| DH14 | yeasty, wort-like | bready, caramel | SF4 | fruity | fruity, plum, honey |
| DH15 | fruity, sulfurous | neutral, earthy | SF5 | fruity | fruity, phenolic |
| DH16 | DMS | cardboard, DMS | SF6 | fruity | fruity, honey |
| DH17 | earthy, cooked | diacetyl | SF8 | fruity | fruity, honey, plum |
| H1 | fruity, acidic | acidic | SF10 | fruity | fruity, musty |
| H2 | fruity, acidic | acidic | S2 | sulfurous, pea soup | sulfurous, pea soup |
| H3 | fruity, acidic | acidic | S3 | fruity, sulfurous | sulfurous |
| H4 | fruity, acidic | acidic | S4 | fruity, sulfurous | sulfurous |
| H5 | neutral | acidic | S5 | malty, wort-like, sweet | sulfurous, sweet |
| H6 | neutral | acidic | S6 | sulfurous | sulfurous, pea soup |
| H7 | neutral | acidic | S7 | sulfurous | sulfurous, pea soup |
| H8 | sweet-sourish | acidic | S8 | sulfurous | sulfurous |
| H9 | sweet-sourish | acidic | S9 | sulfurous | sulfurous |
| H10 | sweet-sourish | acidic | S10 | fruity, sulfurous | fruity, sulfurous |
| KS2 | yeasty | yeasty, fruity | S11 | fruity | fruity, honey |
| KS3 | fruity, flowery | cooked vegetables | T21 | neutral | fruity |
| K1 | fruity, sulfurous | fruity | T22 | sour, cheesy | cheesy |
| K2 | fruity | fruity | T23 | neutral | fruity |
| K3 | fruity | fruity | T24 | neutral | fruity |
| K4 | fruity, pungent | fruity, pungent | T25 | neutral | fruity, apple, berry |
| K5 | fruity | fruity | T26 | neutral | fruity, mirabelle plum |
| K6 | fruity | fruity | T27 | neutral | yeasty |
| K7 | fruity | fruity | T28 | neutral, flowery | fruity, mirabelle plum |
| K8 | fruity, smoky | fruity, smoky | TM1 | yeasty, sweet | yeasty, acidic |
| K9 | fruity | fruity, apple | WA1 | wheat beer | phenolic, wheat beer |
| Km 1 | fruity | fruity | WA2 | vinous | phenolic, vinous |
| Km 2 | fruity | fruity | WA3 | musty | rotten eggs, phenolic |
| Km 3 | fruity | fruity | WA4 | sherry | phenolic, vinous |
| Km 4 | fruity | fruity | WA25 | fruity, pungent | fruity, banana |
| Km 5 | fruity, sweet | fruity, sweet | ZR1 | diacetyl | diacetyl |
| Km 6 | fruity | fruity | ZR2 | fruity, pineapple | fruity |
| Km 7 | fruity, sulfurous | fruity, sulfurous | ZR3 | fruity | fruity |
| Km 8 | fruity, musty | fruity, musty | ZR4 | neutral, yeasty | yeasty |
| Km 9 | fruity, musty | fruity, musty | ZR5 | fruity, acidic | fruity, musty |
| Km 10 | fruity | fruity | ZR6 | fruity, honey | fruity, honey |
| LK1 | neutral, acidic | slightly fruity, acidic | ZR7 | fruity, sweet | fruity, sweet |
| M3 | ethyl acetate | yeasty | ZR8 | fruity | fruity, tea-like |
| M4 | neutral, yeasty | neutral, yeasty | ZR9 | fruity, citrus | fruity |
| M5 | neutral, yeasty | neutral, yeasty | ZR10 | sweet-sourish, fruity | sweet-sourish, fruity |
| M6 | neutral, yeasty | neutral, yeasty | ZR11 | fruity, smoky | fruity, smoky |
| M7 | neutral, yeasty | neutral, yeasty | ZR13 | fruity, honey, sweet | fruity, honey, sweet |
| M8 | neutral | sour | ZR14 | fruity, sweet | fruity |
| M9 | neutral, yeasty | neutral, yeasty | ZF1 | fruity | fruity, pear, musty |
| Strain Abbr | Original Wort [%] | Apparent Attenuation [%] | Ethanol Content [% v/v] | pH Value | Glycerol [g/L] |
|---|---|---|---|---|---|
| SF2 | 12.73 ±σ = 0.07 | 39.53 ±σ = 0.33 | 2.65 ±σ = 0.03 | 4.37 ±σ = 0.01 | 0.24 ±σ = 0.00 |
| SF4 | 12.65 ±σ = 0.11 | 46.53 ±σ = 2.05 | 3.10 ±σ = 0.14 | 4.30 ±σ = 0.03 | 0.24 ±σ = 0.01 |
| SF8 | 12.78 ±σ = 0.07 | 22.57 ±σ = 1.11 | 1.52 ±σ = 0.08 | 4.51 ±σ = 0.04 | 0.33 ±σ = 0.01 |
| S11 | 12.54 ±σ = 0.06 | 86.43 ±σ = 1.79 | 5.73 ±σ = 0.13 | 4.39 ±σ = 0.01 | 0.43 ±σ = 0.04 |
| ZR9 | 12.86 ±σ = 0.06 | 58.07 ±σ = 4.20 | 3.95 ±σ = 0.30 | 4.39 ±σ = 0.05 | 0.47 ±σ = 0.01 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Methner, Y.; Hutzler, M.; Matoulková, D.; Jacob, F.; Michel, M. Screening for the Brewing Ability of Different Non-Saccharomyces Yeasts. Fermentation 2019, 5, 101. https://doi.org/10.3390/fermentation5040101
Methner Y, Hutzler M, Matoulková D, Jacob F, Michel M. Screening for the Brewing Ability of Different Non-Saccharomyces Yeasts. Fermentation. 2019; 5(4):101. https://doi.org/10.3390/fermentation5040101
Chicago/Turabian StyleMethner, Yvonne, Mathias Hutzler, Dagmar Matoulková, Fritz Jacob, and Maximilian Michel. 2019. "Screening for the Brewing Ability of Different Non-Saccharomyces Yeasts" Fermentation 5, no. 4: 101. https://doi.org/10.3390/fermentation5040101
APA StyleMethner, Y., Hutzler, M., Matoulková, D., Jacob, F., & Michel, M. (2019). Screening for the Brewing Ability of Different Non-Saccharomyces Yeasts. Fermentation, 5(4), 101. https://doi.org/10.3390/fermentation5040101