Preliminary Characterization of Yeasts from Bombino Bianco, a Grape Variety of Apulian Region, and Selection of an Isolate as a Potential Starter


 and
and 
Abstract
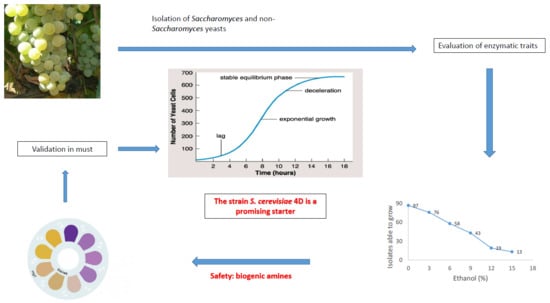
1. Introduction
2. Materials and Methods
2.1. Yeast Isolation
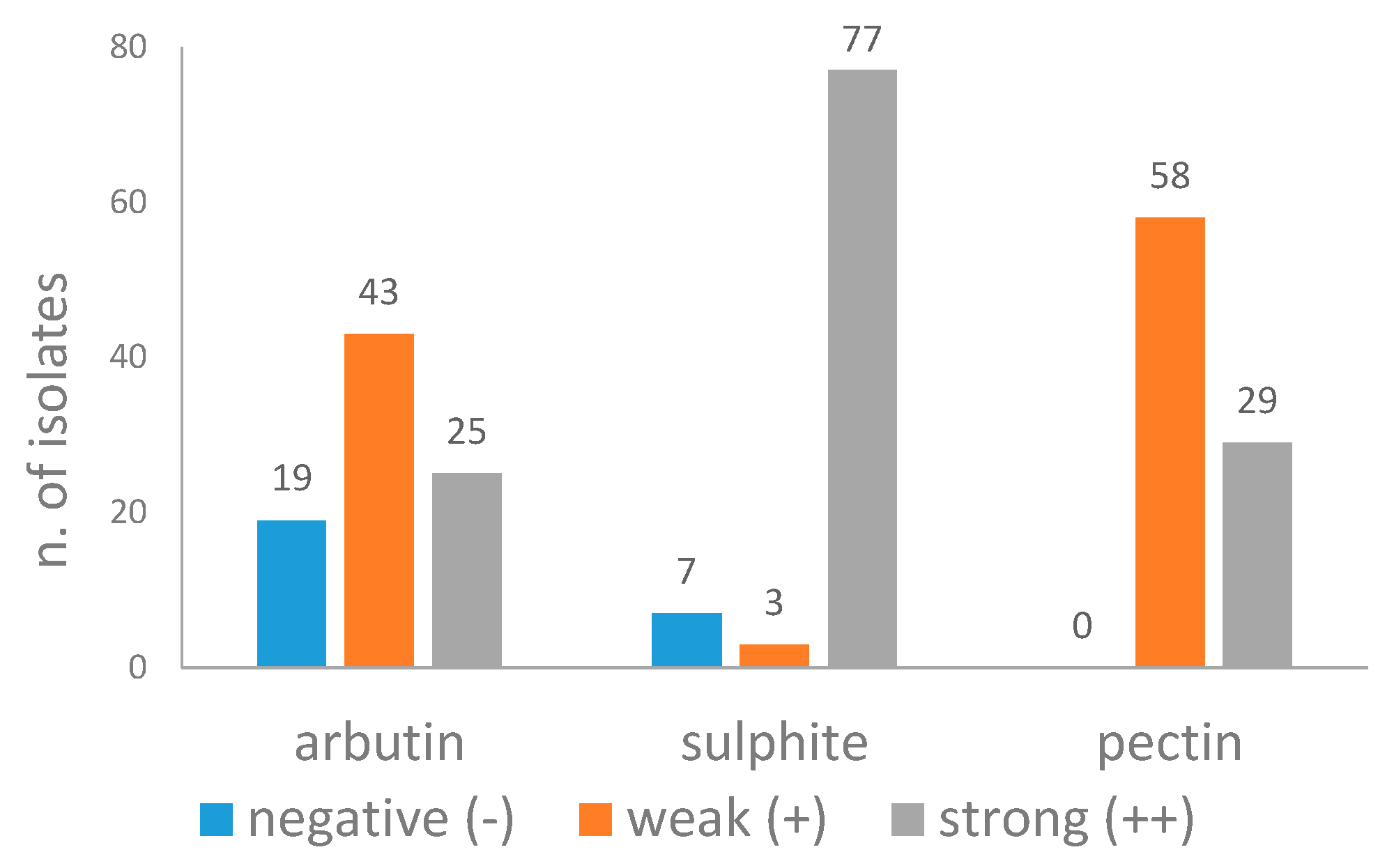
2.2. Arbutin Splitting Test
2.3. Hydrolysis of Pectins
2.4. Sulphite Production
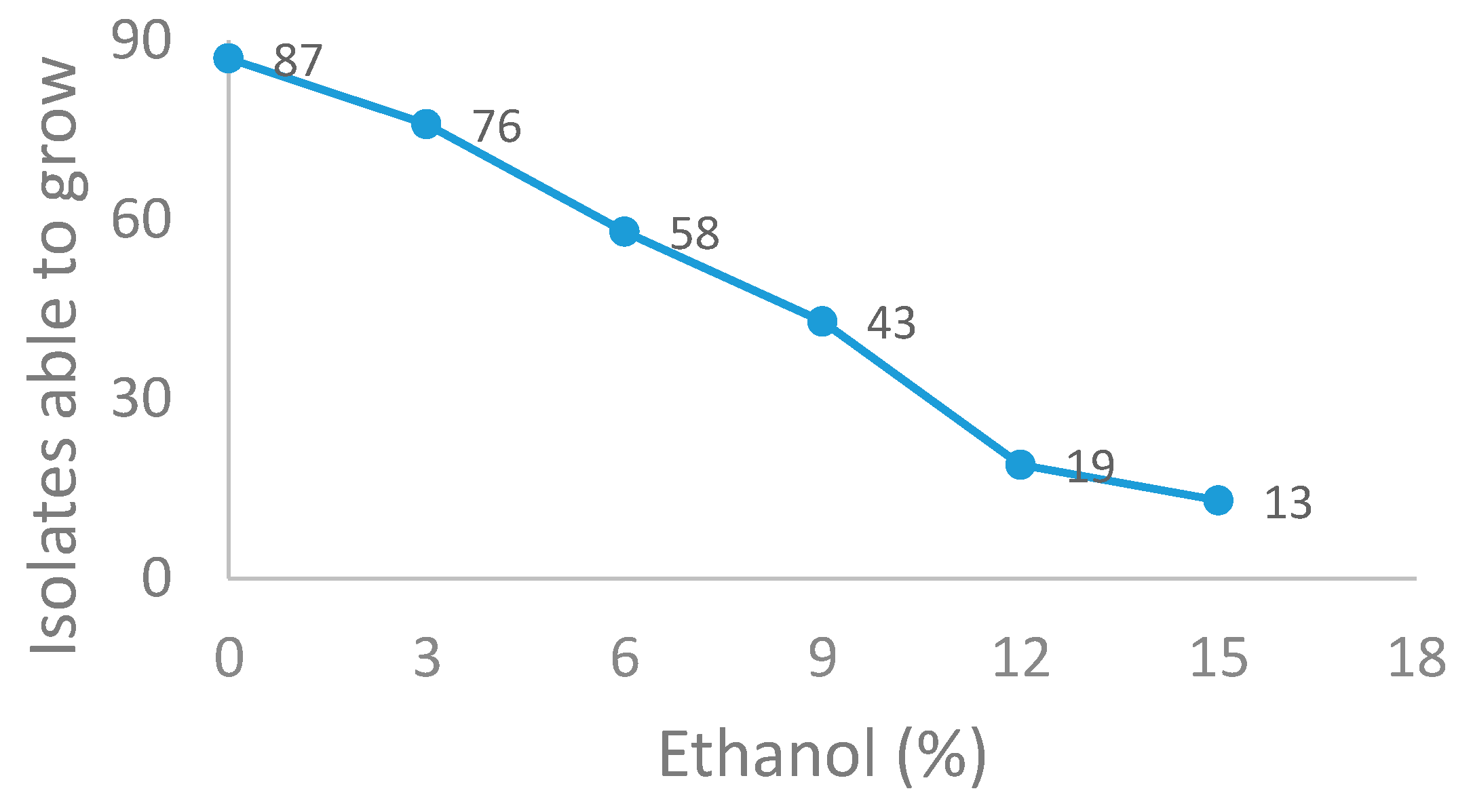
2.5. Resistance to Acetic Acid, Ethanol and SO2
2.6. Preliminary Identification
2.7. Amino Acid Decarboxylation
2.8. Lab-Scale Fermentations and Confirmation of the Technological Performances
2.9. Statistic
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tufariello, M.; Maiorano, G.; Rampino, P.; Spano, G.; Grieco, F.; Perrotta, C.; Capozzi, V.; Grieco, F. Selection of an autochthonous yeast starter culture for industrial production of Primitivo “Gioia del Colle” PDO/DOC in Apulia (Southern Italy). LWT 2019, 99, 188–196. [Google Scholar] [CrossRef]
- Petruzzi, L.; Capozzi, V.; Berbegal, C.; Corbo, M.R.; Bevilacqua, A.; Spano, G.; Sinigaglia, M. Microbial resources and enological significance: Opportunities and benefits. Front. Microbiol. 2017, 8, 995. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, C.; Berbegal, C.; Grieco, F.; Tufariello, M.; Spano, G.; Capozzi, V. Selection of indigenous yeast strains for the production of sparkling wines from native Apulian grape varieties. Int. J. Food Microbiol. 2018, 285, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Romano, P.; Capece, A. Wine microbiology. In Starter Cultures in Food Production, 1st ed.; Speranza, B., Bevilacqua, A., Corbo, M.R., Sinigaglia, M., Eds.; Wiley Blackwell: Oxford, UK, 2017; pp. 255–282. [Google Scholar]
- Ribéreau-Gayon, P.; Denis Dubourdieu, D.; Donèche, B.; Lonvaud, A. Handbook of Enology. In The Microbiology of Wine and Vinifications, 2nd ed.; John Wiley & Sons: New York, NY, USA, 2006; Volume 1, p. 497. [Google Scholar]
- Grieco, F.; Tristezza, M.; Vetrano, C.; Bleve, G.; Panico, E.; Grieco, F.; Mita, G.; Logrieco, A. Exploitation of autochthonous micro-organism potential to enhance the quality of Apulian wine. Ann. Microbiol. 2011, 61, 67–73. [Google Scholar] [CrossRef]
- Tristezza, M.; Vetrano, C.; Bleve, G.; Grieco, F.; Tufariello, M.; Quarta, A.; Mita, G.; Spano, G.; Grieco, F. Autochthonous fermentation starters for the industrial production of Negroamaro wines. J. Ind. Microbiol. Biotechnol. 2012, 39, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Garofalo, C.; Tristezza, M.; Grieco, F.; Spano, G.; Capozzi, V. From grape berries to wine: Population dynamics of cultivable yeasts associated to “Nero di Troia” autochthonous grape cultivar. World. J. Microbiol. Biotechnol. 2016, 32, 59. [Google Scholar] [CrossRef] [PubMed]
- Petruzzi, L.; Bevilacqua, A.; Corbo, M.R.; Speranza, B.; Capozzi, V.; Sinigaglia, M. A focus on quality and safety traits of Saccharomyces cerevisiae isolated from Uva di Troia grape variety. J. Food Sci. 2017, 82, 124–133. [Google Scholar] [CrossRef]
- Tristezza, M.; Fantastico, L.; Vetrano, C.; Bleve, G.; Corallo, D.; Grieco, F.; Mita, G.; Grieco, F. Molecular and technological characterization of Saccharomyces cerevisiae strains isolated from natural fermentation of Susumaniello grape must in Apulia, Southern Italy. Int. J. Microbiol. 2014, 897428. [Google Scholar] [CrossRef]
- Antonacci, D.; Perniola, R. la vitivinicoltura pugliese. Vigneevini 2005, XXXII, 116–122. [Google Scholar]
- Masino, F.; Montevecchi, G.; Arfelli, G.; Antonelli, A. Evaluation of the combined effects of enzymatic treatment and aging on lees on the aroma of wine from Bombino bianco grapes. J. Agric. Food Chem. 2008, 56, 9495–9501. [Google Scholar] [CrossRef]
- Baiano, A.; Varva, G.; De Gianni, A.; Terracone, C.; Viggiani, I.; Del Nobile, M.A. Effects of different vinification technologies on physico-chemical properties and antioxidant activity of ‘Falanghina’ and ‘Bombino bianco’ wines. Eur. Food Res. Technol. 2013, 237, 831–842. [Google Scholar] [CrossRef]
- Vincenzini, M.; Romano, P.; Farris, G.A. Microbiologia del Vino (Wine Microbiology); Casa Editrice Ambrosiana: Milan, Italy, 2006; p. 520. [Google Scholar]
- Petruzzi, L.; Bevilacqua, A.; Corbo, M.R.; Garofalo, C.; Baiano, A.; Sinigaglia, M. Selection of autochthonous Saccharomyces cerevisiae strains as wine starters using a polyphasic approach and ochratoxin A removal. J. Food Prot. 2014, 77, 1168–1177. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, A.; Lage, P.; Vilela, A.; Mendes-Faia, A.; Mendes-Ferreira, A. Phenotypic and metabolic traits of commercial Saccharomyces cerevisiae yeasts. AMB Express 2014, 4, 39. [Google Scholar] [CrossRef] [PubMed]
- Hernández, A.; Martin, A.; Aranda, E.; Pérez-Nevado, F.; Córdoba, M.G. Identification and characterization of yeast isolated from the elaboration of seasoned green table olives. Food Microbiol. 2007, 24, 346–352. [Google Scholar] [CrossRef]
- Sinigaglia, M.; Di Benedetto, N.; Bevilacqua, A.; Corbo, M.R.; Capece, A.; Romano, P. Yeasts isolated from olive mill wastewaters from southern Italy: Technological characterization and potential use for phenol removal. Appl. Microbiol. Biotechnol. 2010, 87, 2345–2354. [Google Scholar] [CrossRef]
- Bover-Cid, S.; Holzapfel, W.H. Improved screening procedure for biogenic amine production by lactic acid bacteria. Int. J. Food Microbiol. 1999, 53, 33–41. [Google Scholar] [CrossRef]
- Van Gerwen, S.J.C.; Zwietering, M.H. Growth and inactivation models to be used in quantitative risk assessments. J. Food Prot. 1998, 6, 1541–1549. [Google Scholar] [CrossRef]
- Baty, F.; Delignette-Muller, M.L. Estimating the bacterial lag time: Which model, which precision? Int. J. Food Microbiol. 2004, 91, 261–277. [Google Scholar] [CrossRef]
- Delignette-Muller, M.L.; Cornu, M.; Pouillot, R.; Denis, J.B. Use of the Bayesian modelling in risk assessment: Application to growth of Listeria monocytogenes and food flora in cold-smoked salmon. Int. J. Food Microbiol. 2006, 106, 195–208. [Google Scholar] [CrossRef]
- Tristezza, M.; Vetrano, C.; Bleve, G.; Spano, G.; Capozzi, V.; Logrieco, A.; Mita, G.; Grieco, F. Biodiversity and safety aspects of yeast strains characterized from vineyards and spontaneous fermentations in the Apulia Region, Italy. Food Microbiol. 2013, 36, 335–342. [Google Scholar] [CrossRef]
- Arevalo-Villena, M.; Briones-Perez, A.; Corbo, M.R.; Sinigaglia, M.; Bevilacqua, A. Biotechnological application of yeasts in food science. Starter cultures, probiotic, and enzyme production. J. Appl. Microbiol. 2017, 123, 1360–1372. [Google Scholar] [CrossRef] [PubMed]
- Pando Bedriñana, R.; Querol Simón, A.; Suárez Valles, B. Genetic and phenotypic diversity of autochthonous cider yeasts in a cellar from Asturias. Food Microbiol. 2010, 27, 503–508. [Google Scholar] [CrossRef] [PubMed]
- Fia, G.; Giovani, G.; Rosi, I. Study of β-glucosidase production by wine-related yeasts during alcoholic fermentation. A new rapid fluorimetric method to determine enzymatic activity. J. Appl. Microbiol. 2005, 99, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Mateo, J.J.; Di Stefano, R. Description of the beta-glucosidase activity of wine yeast. Food Microbiol. 1997, 14, 583–591. [Google Scholar] [CrossRef]
- Chaiyasut, C.; Pengkumsri, N.; Sirilun, S.; Peerajan, S.; Khongtan, S.; Sundaram Sivamaruthi, S. Assessment of changes in the content of anthocyanins, phenolic acids, and antioxidant property of Saccharomyces cerevisiae mediated fermented black rice bran. AMB Express 2017, 7, 114. [Google Scholar] [CrossRef]
- Mendes-Ferreira, A.; Mendes-Faia, A.; Leão, C. Survey of hydrogen sulphide production by wine yeasts. J. Food Prot. 2002, 65, 1033–1037. [Google Scholar] [CrossRef]
- Bevilacqua, A.; Beneduce, L.; Sinigaglia, M.; Corbo, M.R. Selection of yeasts as starter cultures for table olives. J. Food Sci. 2013, 78, M742–M751. [Google Scholar] [CrossRef]
- Merín, M.G.; Martín, M.C.; Rantsiou, K.; Cocolin, L.; de Ambrosini, V.I. Characterization of pectinase activity for enology from yeasts occurring in Argentine Bonarda grape. Braz. J. Microbiol. 2015, 46, 815–823. [Google Scholar] [CrossRef]
- Laulund, S.; Wind, A.; Derkx, P.M.F.; Zuliani, V. Regulatory and safety requirements for food cultures. Microorganisms 2017, 5, 28. [Google Scholar] [CrossRef]
- Galgano, F.; Caruso, M.; Condelli, N.; Favati, F. Agmatine in fermented foods. Front. Microbiol. 2012, 3, 199. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
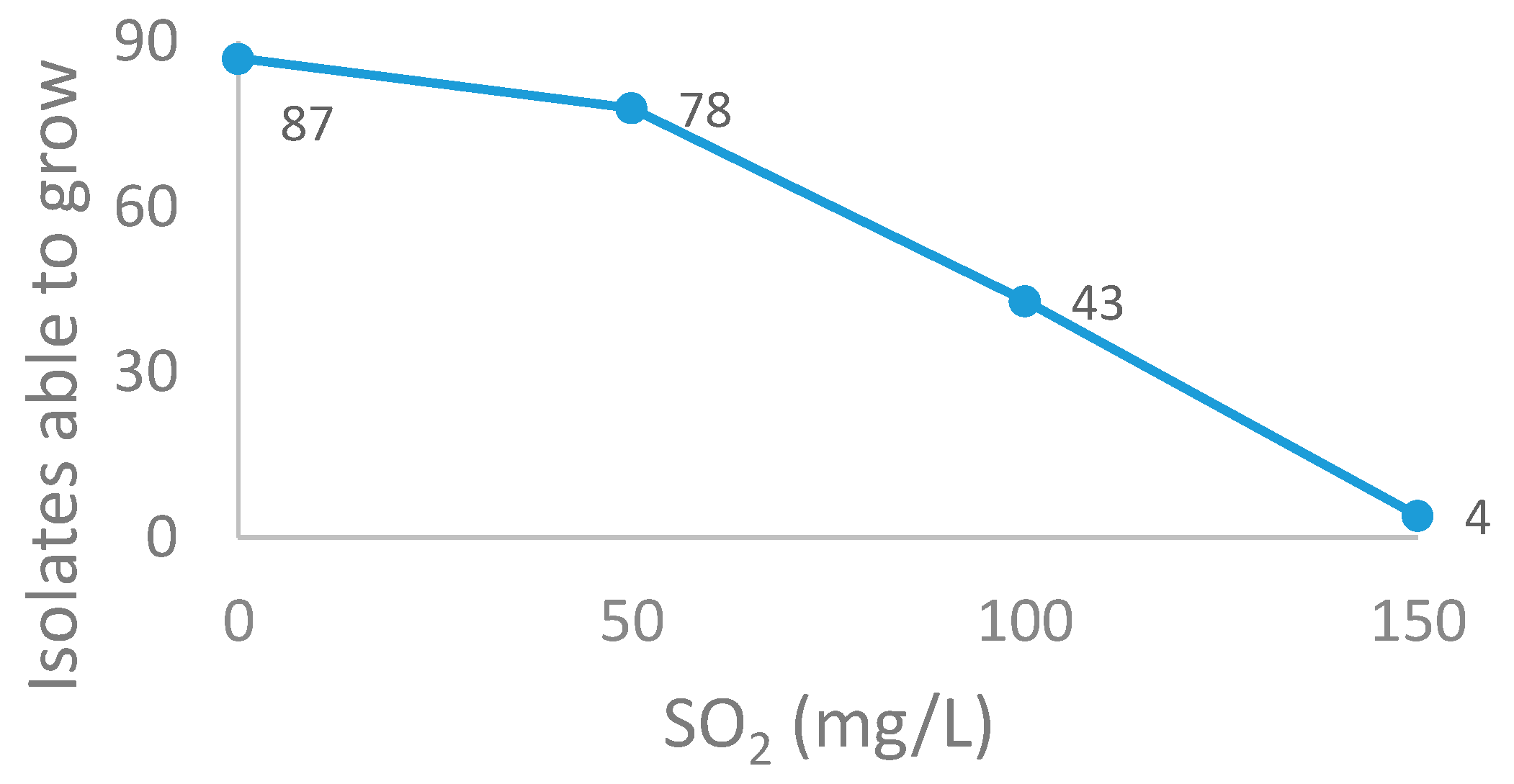{kind=link}
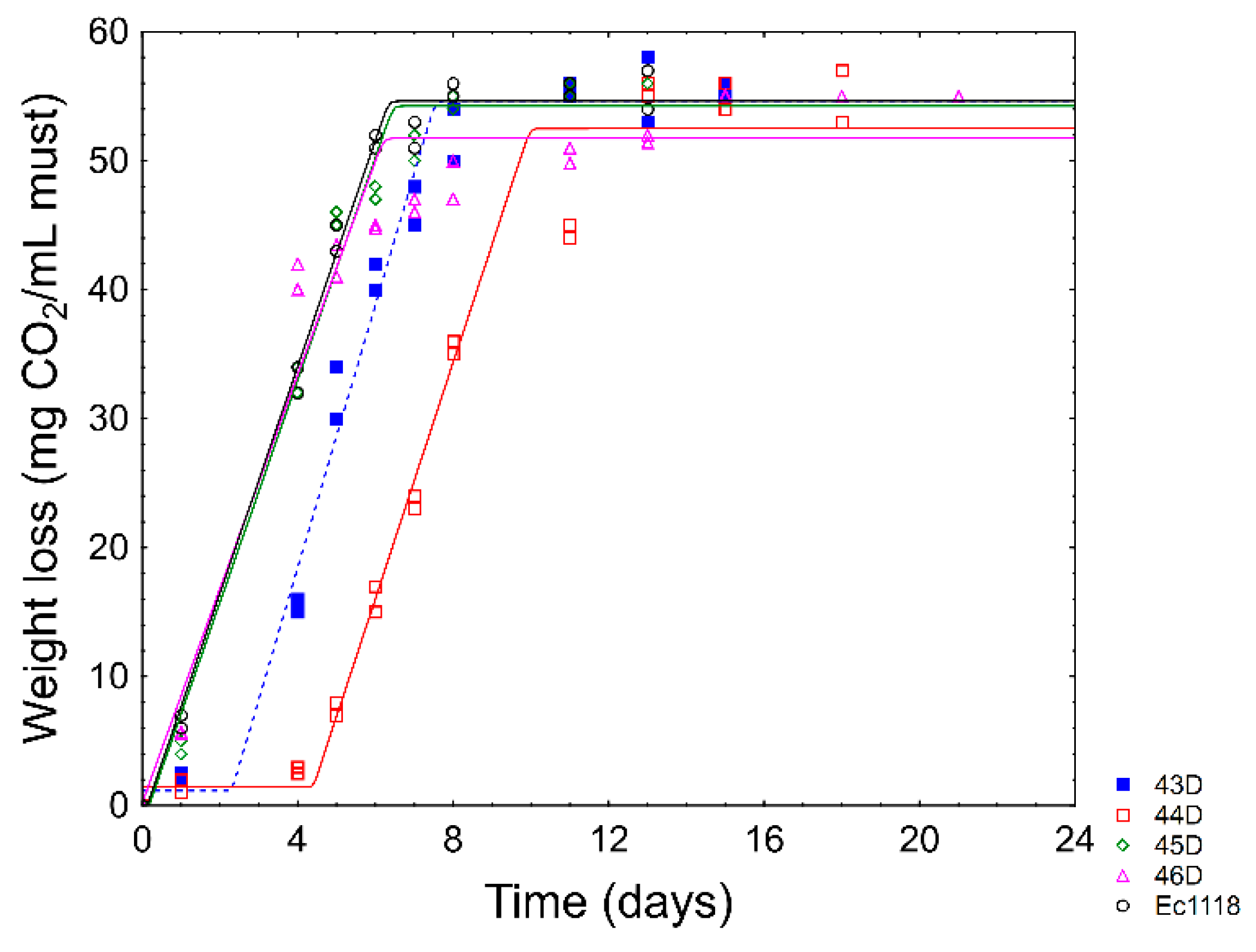{kind=link}
| Yeast Isolates | Cat | Arb | Sul | Pec | SO2 | EtOH | Acetic |
|---|---|---|---|---|---|---|---|
| 43D | ++ | + | − | ++ | 150 ppm | 15% | 0.10% |
| 44D | ++ | + | − | ++ | 150 ppm | 15% | 0.10% |
| 45D | + | + | + | ++ | 150 ppm | 15% | 0.10% |
| 46D | + | + | + | ++ | 150 ppm | 15% | 0.10% |
| Yeast Isolates | A | B | C | D | E | F | G | H | I |
|---|---|---|---|---|---|---|---|---|---|
| 43D | − | − | − | − | − | − | − | − | − |
| 44D | − | − | − | − | − | − | − | − | − |
| 45D | − | − | − | − | − | − | − | − | − |
| 46D | + | − | − | − | − | +/− | − | − | + |
| Yeast Isolates | Ethanol (g/L) | Glycerol (g/L) | Volatile Acidity (Acetic Acid, g/L) |
|---|---|---|---|
| 43D | 9.76 ± 0.98 | 5.64 ± 0.89 | 0.45 ± 0.09 |
| 44D | 8.78 ± 1.21 | 5.45 ± 0.63 | 0.36 ± 0.12 |
| 45D | 9.89 ± 0.06 | 6.12 ± 0.65 | 0.41 ± 0.07 |
| 46D | 10.25 ± 0.34 | 6.21 ± 0.09 | 0.42 ± 0.08 |
| Ec1118 | 11.20 ± 0.43 | 6.01 ± 0.11 | 0.39 ± 0.12 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Speranza, B.; Campaniello, D.; Petruzzi, L.; Sinigaglia, M.; Corbo, M.R.; Bevilacqua, A. Preliminary Characterization of Yeasts from Bombino Bianco, a Grape Variety of Apulian Region, and Selection of an Isolate as a Potential Starter. Fermentation 2019, 5, 102. https://doi.org/10.3390/fermentation5040102
Speranza B, Campaniello D, Petruzzi L, Sinigaglia M, Corbo MR, Bevilacqua A. Preliminary Characterization of Yeasts from Bombino Bianco, a Grape Variety of Apulian Region, and Selection of an Isolate as a Potential Starter. Fermentation. 2019; 5(4):102. https://doi.org/10.3390/fermentation5040102
Chicago/Turabian StyleSperanza, Barbara, Daniela Campaniello, Leonardo Petruzzi, Milena Sinigaglia, Maria Rosaria Corbo, and Antonio Bevilacqua. 2019. "Preliminary Characterization of Yeasts from Bombino Bianco, a Grape Variety of Apulian Region, and Selection of an Isolate as a Potential Starter" Fermentation 5, no. 4: 102. https://doi.org/10.3390/fermentation5040102
APA StyleSperanza, B., Campaniello, D., Petruzzi, L., Sinigaglia, M., Corbo, M. R., & Bevilacqua, A. (2019). Preliminary Characterization of Yeasts from Bombino Bianco, a Grape Variety of Apulian Region, and Selection of an Isolate as a Potential Starter. Fermentation, 5(4), 102. https://doi.org/10.3390/fermentation5040102