Simple Summary
Gene editing has become a key technology in poultry research. Common methods include ZFNs, TALENs, CRISPR/Cas9 and others, among which CRISPR/Cas9 is the most widely used due to its simplicity and high efficiency. This paper reviews several common gene-editing techniques and their applications in enhancing disease resistance, improving production performance, and detecting diseases. Finally, the existing challenges and future opportunities are summarized.
Abstract
Gene editing technology, a revolutionary tool in molecular biology, enables precise modifications of genomic sequences and gene expression patterns, thereby conferring desired traits to cells or organisms. Since 2014, CRISPR/Cas9 has rapidly become the most widely used gene editing method in agricultural animals due to its high editing efficiency. Subsequently, the development of novel gene editing systems, such as base editors and prime editors, has provided enhanced precision and reduced off-target effects. These advancements have facilitated the transition of gene editing from laboratory research to clinical and agricultural applications. Gene editing has been extensively utilized to enhance production traits, improve disease resistance, facilitate disease detection, and establish disease models. This review outlines the development of gene editing technologies, discusses the advantages and limitations of key gene editing tools, and explores their applications in poultry. Furthermore, it examines the challenges and future prospects of gene editing in animal husbandry, including off-target effects, ethical concerns, and technical complexities.
1. Introduction
Gene editing technology utilizes molecular biology techniques to precisely alter genomic sequences and gene expression patterns, enabling cells or organisms to acquire desired traits. In recent years, gene editing has advanced rapidly, with traditional gene editing technologies, including the first-generation ZFN technology, the second-generation TALEN technology, and the third-generation CRISPR/Cas9 technology [1]. Since 2014, various Cas9 variants, as well as base editors and prime editing, have been developed [2]. With continuous technological advancements, gene editing has progressively expanded from laboratory research to clinical applications and has been extensively studied for modifying biological traits in agricultural animals, demonstrating significant potential for novel breed development and enhancement of livestock production efficiency. Compared to conventional breeding methods and genomics-assisted breeding approaches, gene editing-based breeding offers distinct advantages. For instance, while traditional breeding is limited to selecting existing traits within a population, gene editing enables the precise introduction of novel target traits. Moreover, this technology significantly accelerates the breeding process and circumvents the selection of undesirable traits caused by linkage disequilibrium in conventional breeding programs. Taking the CRISPR/Cas9 gene editing system as an example (Figure 1), genetic modification in agricultural animals begins with the identification of candidate genes based on research objectives, followed by the design of target-specific gRNAs using bioinformatics tools. Subsequently, molecular cloning techniques are employed to integrate the gRNA and Cas9 nuclease sequences into expression vectors, incorporating selectable markers to facilitate subsequent identification. Upon completion of vector construction, an appropriate delivery method must be selected.
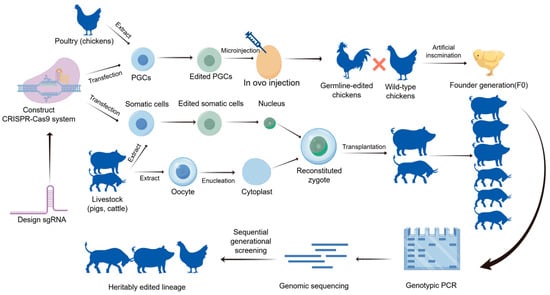
Figure 1.
The CRISPR/Cas9 gene editing process for livestock and poultry. The schematic illustrates the systematic workflow for generating gene-edited animals. Initially, specific sgRNAs are designed to construct the CRISPR/Cas9 expression vectors. For poultry (e.g., chickens), primordial germ cells (PGCs) are extracted, transfected in vitro, and then injected into developing embryos to generate chimeric founders (F0), which are subsequently crossed to produce heritable germline-edited lineages. For livestock (e.g., pigs, cattle), somatic cells are edited and utilized as donor cells for somatic cell nuclear transfer (SCNT). The reconstituted zygotes are transplanted into surrogate mothers. Finally, the heritably edited lineages are validated through genotypic PCR, genomic sequencing, and sequential generational screening to ensure the stability and accuracy of the targeted traits.
In livestock, plasmid delivery is typically achieved through somatic cell nuclear transfer (SCNT) or pronuclear microinjection of zygotes, with the edited positive embryos cultured in vitro to the blastocyst stage before being transferred into synchronized recipient females for gestation completion. In poultry, primordial germ cells (PGCs) serve as the primary editing vehicle, where in vitro-edited PGCs are transplanted into avian embryos to produce chimeric offspring. Postnatal individuals must undergo validation of target gene editing efficiency via PCR and sequencing techniques, complemented by whole-genome sequencing to assess potential off-target effects. To ensure stable inheritance of the edited traits, homozygous individuals are obtained through successive generations, ultimately establishing genetically edited breeds or lines with the desired phenotypes. Currently, gene editing technology has been applied to improve production traits, enhance disease resistance, and disease detection. This paper will review the development of gene editing technology, discuss the advantages and disadvantages of several gene editing techniques, and examine the application of gene editing in poultry animals, along with the challenges it faces in this field.
2. Development of Gene Editing Technology
Gene editing technology essentially involves the use of specially designed nucleases to create double-strand breaks (DSBs) at specific locations (S) in the genome of an organism. The cell’s own repair mechanisms, including non-homologous end joining (NHEJ) and homologous recombination (HDR), then repair the damaged genome, allowing for specific modifications such as insertions, deletions, or replacements at the target gene [3]. Traditional gene editing technologies include zinc finger nucleases (ZFNs), transcription activator-like effector nucleases (TALENs), and RNA-guided DNA endonucleases based on clustered regularly interspaced short palindromic repeats (CRISPR) and CRISPR-associated protein 9 (Cas9) (Figure 2) [4]. CRISPR/Cas9 technology has demonstrated significant advantages in terms of time and cost, with gene editing efficiency generally surpassing that of ZFNs and TALENs, making it the primary gene editing method in current agricultural animal research. Since 2014, it has been widely applied in livestock and poultry cell lines and species. Derived from CRISPR/Cas9, gene editing tools such as dCas9, nCas9, Cas12a (Figure 3), and novel gene editing technologies like base editors and prime editing have shown great potential in future animal husbandry applications due to their high targeting efficiency and low off-target activity (Figure 3). Gene editing technology has evolved over time, with several revolutionary breakthroughs since the 1970s, becoming an essential tool in life science research and medical applications. The development timeline is illustrated in Table 1, with key technologies introduced.
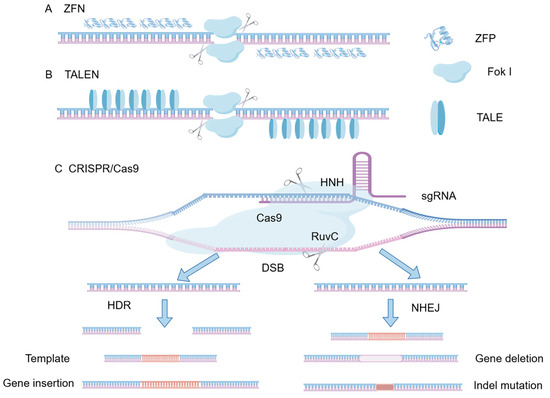
Figure 2.
Schematic diagram of the structure of ZFN, TALEN and CRISPR/Cas9. (A) Zinc Finger Nucleases (ZFNs) consist of customizable zinc-finger DNA-binding proteins fused with a FokI endonuclease cleavage domain. (B) Transcription Activator-Like Effector Nucleases (TALENs) utilize TALE repeat arrays for DNA recognition, similarly tethered to a FokI nuclease. (C) The CRISPR/Cas9 system employs a single guide RNA (sgRNA) to direct the Cas9 nuclease to a specific genomic locus adjacent to a Protospacer Adjacent Motif (PAM). The Cas9 enzyme utilizes its HNH and RuvC domains to introduce a double-strand break (DSB). The resulting DSBs are subsequently repaired either by Non-Homologous End Joining (NHEJ), typically leading to gene deletion or indel mutations, or by Homology-Directed Repair (HDR) in the presence of a repair template, enabling precise gene insertion or modification.
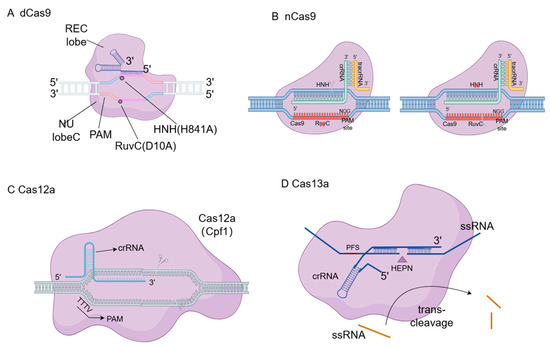
Figure 3.
Structural diagrams of dCas9, nCas9, Cas12a and Cas13a. (A) Dead Cas9 (dCas9) contains inactivating mutations in both the RuvC and HNH nuclease domains, allowing it to bind to target DNA without causing cleavage, which is utilized for gene regulation (CRISPRa/CRISPRi). (B) Nickase Cas9 (nCas9) possesses a mutation in one of the nuclease domains, generating single-stranded DNA nicks rather than double-strand breaks, thereby reducing off-target effects when paired with dual sgRNAs. (C) Cas12a (Cpf1) relies on a single crRNA and a T-rich PAM sequence to create staggered double-strand breaks distal to the PAM. (D) Cas13a specifically targets and cleaves single-stranded RNA (ssRNA), possessing a unique collateral trans-cleavage activity upon target recognition, which is widely utilized in diagnostic biosensors.

Table 1.
Development of Gene-Editing Technologies and Their Applications in Poultry.
2.1. CRISPR/Cas9
The limitations of the first two generations of gene editing technologies drove the development of the third generation, CRISPR/Cas9. CRISPR is a sequence of short palindromic repeats found in prokaryotic genomes, first discovered in E. coli in 1987 [21]. The CRISPR sequence has three parts: a leader region, conserved repeats, and variable spacers. Cas refers to CRISPR-associated proteins. The Cas9 nuclease complex consists of the Cas9 protein, a specific CRISPR RNA (crRNA), and a trans-activating CRISPR RNA (tracrRNA). CRISPR/Cas9 serves as a defense mechanism, allowing bacteria to resist viral infections and phage invasions [22]. The antimicrobial defense mechanism of the bacterial CRISPR/Cas9 system against exogenous pathogens comprises three distinct phases. When foreign DNA first invades the host, the Cas1 and Cas2 endonucleases recognize and cleave the viral genome, inserting fragments into the leader region of the bacterial CRISPR sequence while replicating the existing repeat sequences to form new repeats. When foreign DNA invades again, the leader region initiates the transcription of the CRISPR sequence into a precursor crRNA (pre-crRNA), which is then processed by the Cas9 protein to form crRNA. The crRNA and tracrRNA form a dimer and bind to the Cas9 protein, forming a crRNA-Cas9-tracrRNA complex. This complex is then degraded by RNase III, producing small crRNA guides that target and cleave the foreign DNA [1,23,24].
The artificially designed CRISPR/Cas9 system consists of a duplex RNA (tracrRNA/crRNA) and the Cas9 endonuclease bound to the RNA strand. The crRNA region is approximately 20 nucleotides long and contains a guide RNA (gRNA) that binds to the target DNA sequence. The tracrRNA consists of a 14-nucleotide anti-repeat region and three loops. The targeting specificity of the CRISPR/Cas9 system is partly determined by the base pairing between the gRNA and the target DNA, and partly by the Cas9 protein and a short DNA sequence known as the protospacer adjacent motif (PAM), typically composed of the NGG sequence, where N can be any of the four DNA nucleotides recognized by the Cas9 nuclease. The PAM sequence is located at the 3′ end of the target DNA. Before designing the gRNA, a suitable PAM sequence must be identified in the genome. Cas9 initiates the target site search process by detecting the preferred PAM sequence, allowing the gRNA to precisely bind to the target DNA. Crucially, the DNA targeting and hybridization mechanism of the CRISPR/Cas9 system exhibits distinct structural and functional polarity. The recognition process is highly directional; following PAM recognition at the 3′ end of the target DNA strand, the unwinding of the DNA double helix and the subsequent RNA-DNA heteroduplex formation (R-loop) propagate in a polar, zipper-like manner. This propagation proceeds strictly from the PAM-proximal “seed” region (typically the 8–12 nucleotides adjacent to the PAM) towards the PAM-distal 5′ end. This functional polarity fundamentally dictates the mismatch tolerance of the system: the PAM-proximal seed region is highly stringent and sensitive to base mismatches, whereas the PAM-distal end exhibits significantly greater flexibility. Understanding this directional polarity is essential for optimizing gRNA design to maximize on-target cleavage efficiency while mitigating off-target hybridization. The Cas9 helicase domain unwinds the double strand, while the nuclease domains (RuvC and HNH) execute the DSB in the DNA.
The CRISPR/Cas9 system was first proposed as an effective gene editing tool in 2012 and won the Nobel Prize in Chemistry in 2020 [25]. The CRISPR/Cas9 system is classified into types I, II, and III based on the characteristics of the Cas9 protein, with the type II prokaryotic CRISPR adaptive immune system proven to facilitate RNA-guided site-specific DNA cleavage. In 2013, Zhang et al. first used the type II CRISPR/Cas9 system to perform gene editing in mammalian cells [26]. The CRISPR/Cas9 system can recognize and cleave specific target DNA, creating double-strand breaks that induce the cell’s DNA repair mechanisms. In the absence of homologous DNA, repair occurs via NHEJ, leading to gene deletions [27]. When homologous DNA is present, repair occurs via HR, and by designing a single-stranded oligonucleotide (ssODN, or donor sequence), specific gene insertions can be achieved. Compared to ZFN and TALEN technologies, CRISPR is simpler, faster, and offers significant time and cost advantages. It also allows for simultaneous editing of multiple genes within a single cell, demonstrating high gene editing efficiency.
2.1.1. Nickase Cas9 (nCas9)
Nickase Cas9 (nCas9) is generated by activating one of the nuclear domains (RuvC or HNH) of Cas9, resulting in a single-strand nicking enzyme [28]. Compared to traditional Cas9, nCas9 only cleaves one strand of DNA, significantly reducing off-target effects and providing higher editing precision. By designing two adjacent sgRNAs, nCas9 can induce a “double-nicking” strategy, where double-strand breaks are only triggered when both nicks are present, reducing off-target rates by 50 to 1500 times [29]. The two gRNAs form cuts at the first and fifth bases after the recognition site [30].
2.1.2. dCas9
Dead Cas9 (dCas9) is a variant of Cas9 with deactivated nuclease activity through point mutations (e.g., D10A and H840A), retaining DNA-targeting capability but no longer cleaving DNA. This characteristic makes it an ideal tool for gene regulation. dCas9 can be fused with regulatory factors to activate or silence genes and modulate protein activity levels. For example, fusing dCas9 with transcriptional activators (e.g., VP64) or repressors (e.g., KRAB domain) enables gene activation (CRISPRa) or interference (CRISPRi) [31].
2.1.3. CRISPR/Cas12a
The CRISPR/Cas12a system, like CRISPR/Cas9, belongs to the type II CRISPR system but has significant differences, offering an alternative for gene editing. Unlike Cas9, Cas12a’s activity is primarily mediated by a short crRNA. In the Cas12a system, the specificity of target DNA recognition is guided by a longer spacer sequence of at least 22–23 nucleotides, enhancing recognition specificity. Cas12a recognizes PAM sequences rich in thymine (T) rather than the guanine (G)-rich PAMs required by Cas9, expanding the range of potential target sites. Cas12a generates staggered DSBs distal to the PAM, whereas Cas9 produces DSBs proximal to the PAM. Cas12a cleaves the target DNA strand 23 nucleotides downstream of the PAM region. Similar to Cas9, only some Cas12a homologs exhibit significant activity in eukaryotic cells. Cas12a, like Cas9, has a bilobed structure, including REC and NUC lobes. The Cas12a system uses the RuvCnuclease domain and a crRNA with RNase WED III to cleave both DNA strands, while Cas9 uses two domains for cleavage. In type II-A systems, Cas9 recognizes the PAM sequence located directly downstream of the protospacer, requiring tracrRNA, whereas Cas12a only requires crRNA to cleave the target strand [32,33].
2.1.4. CRISPR/Cas13
The CRISPR/Cas13 system is an RNA-targeting CRISPR system initially discovered in bacteria as a defense mechanism against RNA viruses. The CRISPR/Cas13 system achieves gene regulation by recognizing and cleaving single-stranded RNA (ssRNA) [34]. Upon binding to the target RNA, the Cas13 protein activates its nuclease activity, cleaving the target RNA and inhibiting its expression. It has been used to develop highly sensitive RNA detection tools.
2.2. Base Editor
Traditional gene editing technologies require the creation of DNA double-strand breaks, leading to unavoidable biological damage. Both DSB repair pathways, NHEJ and HDR, have limitations. In most somatic cells, NHEJ repair is more efficient than HDR but is prone to erroneous base insertions or deletions. In contrast, HDR can precisely introduce desired changes, including insertions, deletions, or replacements based on a DNA repair template [35]. However, HDR is limited to the S/G2 phase of the cell cycle and is inefficient in most therapeutically relevant cell types [35,36]. The development of single-base editing technology based on CRISPR/Cas9 has achieved new breakthroughs in gene editing tools. Currently developed single-base editing systems include cytosine base editors (CBEs), adenine base editors (ABEs), guanine base editors (GBEs), and prime editors (PEs) (Figure 4). These systems enable the replacement of different bases, such as A-to-G, C-to-T, C-to-G, and C-to-A. The avoidance of DSBs significantly reduces the occurrence of unintended mutations, such as base insertions, deletions, or frameshifts, making these systems highly efficient and precise. They hold great significance in the establishment of disease models and gene therapy.
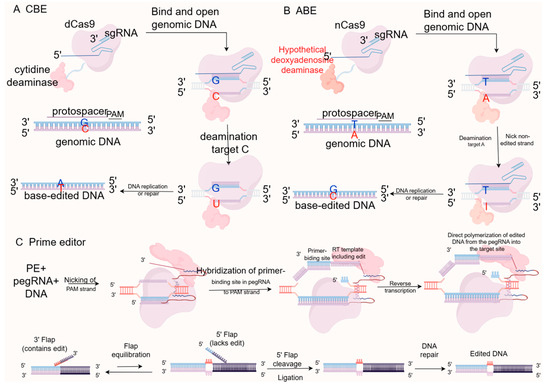
Figure 4.
Operating principle of CBE, ABE and PE. (A) Cytosine Base Editors (CBEs) fuse a cytidine deaminase with dCas9 or nCas9 to catalyze the conversion of cytosine (C) to uracil (U), ultimately resulting in a C-G to T-A transition during DNA repair. (B) Adenine Base Editors (ABEs) employ an evolved deoxyadenosine deaminase to convert adenine (A) to inosine (I), which is treated as guanine (G) by the polymerase, leading to an A-T to G-C transition. (C) Prime Editors (PEs) consist of an nCas9 fused to a reverse transcriptase (RT) and utilize a prime editing guide RNA (pegRNA). The pegRNA not only specifies the target site but also provides the RNA template containing the desired edits (insertions, deletions, or substitutions), enabling highly precise genome modifications without requiring donor DNA templates or inducing double-strand breaks.
2.3. Prime Editing
Prime editing (PE) systems achieve gene editing at target sites by fusing a Cas9 nickase with a reverse transcriptase and utilizing a prime editing guide RNA (pegRNA). The pegRNA consists of three parts: a single-guide RNA (sgRNA), a primer binding site (PBS), and a reverse transcription template (RT template) containing the desired edit [17]. The pegRNA guides nCas9 to create a single-strand nick at the target DNA site. The nicked DNA strand binds to the PBS sequence, and the reverse transcriptase synthesizes a new DNA sequence using the RT template, enabling base substitutions, insertions, or deletions. Prime editing can achieve 12 types of base substitutions, small insertions, and deletions, offering a new method to minimize off-target effects and improve target specificity in the genome compared to base editors [37]. However, prime editing systems and their applications still require improvements, such as addressing gRNA-dependent or independent off-target effects, high rates of insertions and deletions mediated by PE3, and the delivery of prime editing systems in adult animals. Further research on animal models is needed.
2.4. Gene Delivery Systems
The successful application of gene editing technology relies not only on efficient editing tools but also on effective delivery systems that transport editing components to target cells. The choice of delivery system and vector directly affects the efficiency, specificity, and safety of gene editing. Table 2 summarizes the principles and characteristics of different gene delivery systems and introduces several classic delivery vectors.
Table 2.
Principles and characteristics of different gene delivery systems.
2.4.1. Viral Vectors
Viral vectors are among the most commonly used tools for gene editing delivery, including lentivirus (Lentivirus) vectors, adenovirus (Adenovirus) vectors, and adeno-associated virus (AAV) vectors. AAV is widely used for in vivo gene editing due to its low immunogenicity and long-term expression characteristics. Lentiviruses, which can integrate into the host genome, are suitable for in vitro cell editing and stem cell research [45]. However, the biosafety of lentiviral vectors cannot be guaranteed, and AAV vectors can only carry approximately 4.7 kb of foreign DNA, limiting their application due to packaging capacity and potential immune responses [46].
2.4.2. Non-Viral Vectors
Non-viral vectors, due to their low immunogenicity and ease of large-scale production, are increasingly becoming an important choice for gene editing delivery. Lipid nanoparticles (LNPs) are currently the most commonly used non-viral vectors, capable of efficiently delivering CRISPR/Cas9 mRNA and sgRNA. Additionally, polymer nanoparticles such as PEI and PLGA have been extensively studied, with surface modifications further improving targeting and delivery efficiency [47]. Compared to viral vectors, non-viral vectors have lower transfection efficiency and potential cytotoxicity, limiting their in vivo applications.
2.4.3. Nanomaterial Vectors and Cell Delivery
In recent years, novel nanomaterial vectors, such as gold nanoparticles, carbon nanotubes, and metal–organic frameworks (MOFs), have shown great potential in gene editing delivery. These materials offer high loading capacity, good biocompatibility, and controllable release properties. MOFs, due to their degradability and high loading capacity, have been used to deliver CRISPR/Cas9 complexes, significantly improving editing efficiency. Additionally, modifying the surface of vectors with antibodies or ligands enables targeted delivery to specific cell types, enhancing the precision of gene editing. In cancer therapy, nanoparticles have successfully delivered the CRISPR/Cas9 system to tumor cells, significantly improving treatment efficacy and reducing off-target effects [48]. Cell-penetrating peptides (CPPs) have also been used to enhance the intracellular delivery efficiency of gene editing components [49].
The advancement of gene editing technologies has profoundly influenced the iterative innovation of techniques in the livestock sector. The following table presents a timeline of major milestones in the development of gene editing technology across various livestock species (Table 3).
Table 3.
A timeline of major milestones in gene editing technology for livestock.
Building on this broader context of livestock applications, the following section focuses specifically on the application of gene editing technology in poultry.
3. Applications of Gene Editing Technology in Poultry Animals
The development of CRISPR/Cas9-mediated genome editing technology is crucial for improving gene targeting efficiency, with the potential to rapidly enhance disease resistance, disease detection, and product quality traits in poultry and other agricultural animals, thereby improving the safety, controllability, and environmental benefits of gene-edited animal breeding (Figure 5). Table 4 summarizes the main applications of gene editing technology in poultry, organized by application area, target genes, methods, and achievements.
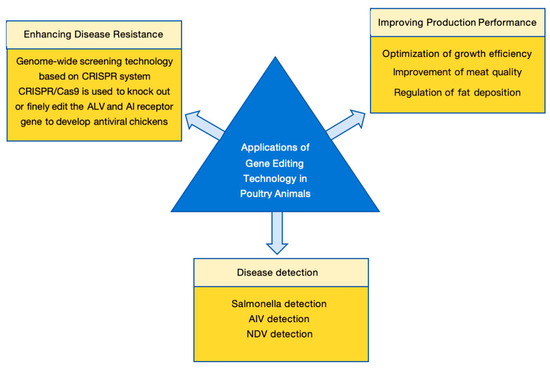
Figure 5.
Applications of gene editing in poultry animals. The application of gene editing technology in the poultry industry is primarily categorized into three strategic domains: (1) Enhancing disease resistance by targeting host susceptibility factors or viral receptors (e.g., ALV and AIV resistance); (2) Improving production performance, including the optimization of growth efficiency, modulation of fat deposition, and enhancement of meat quality traits; (3) Facilitating rapid and highly sensitive disease detection utilizing CRISPR-based diagnostic systems (e.g., CRISPR-Cas12a/Cas13a for Salmonella, AIV, and NDV surveillance).
Table 4.
Applications of Gene-Editing Technology in Poultry: Targets, Genes, Methods, and Achievements.
3.1. Enhancing Disease Resistance
Gene editing technology has shown significant advantages in the research and control of animal viral diseases. Strategies for controlling animal viral diseases include precise modification of the host genome to enhance resistance to viral infections and modification of the viral genome to reduce its replication and transmission capabilities. This technology has been widely applied in poultry for diseases such as Avian Leukosis, Newcastle disease virus (NDV) and Avian Influenza virus (AIV).
3.1.1. Genome-Wide Screening Technology Based on CRISPR System
In recent years, genome-wide CRISPR library screening technology has found widespread utility in diverse fields, including the development of disease-resistant strains, the establishment of disease models, and the design and advancement of vaccines—all achieved by precisely modulating the expression of host factors. The intricate crosstalk between viruses and host cells adds further complexity to disease treatment, rendering the identification of host factors involved in virus–host interactions a critical hurdle for precise therapeutic interventions. Benefiting from the progress of CRISPR gene-editing technology, this screening approach has been successfully applied to pinpoint key antiviral host factors in poultry. Numerous studies have confirmed that CRISPR library screening can effectively identify host factors that engage with viruses across multiple stages of infection, such as adsorption, endocytosis, and replication. These identified host factors offer valuable molecular insights for the prevention and treatment of viral diseases.
During the viral adsorption phase, genome-wide screening has uncovered core regulatory factors including LDLR, SLC35A1, and SLC35B2. These factors either act as viral receptors, participate in receptor biosynthesis, or compete with viruses for receptor binding (e.g., B4GALNT2), thereby governing the initial attachment of viruses to host cells.
In the viral endocytosis stage, screened factors such as SGMS1, IGDCC4, COG8, CYTH2, and PKCθ have been shown to be involved in transmembrane transport and intracellular vesicular trafficking, facilitating viral internalization into host cells and subsequent intracellular transport processes.
At the viral replication stage, factors like TMEM41B, the GARP/EARP complex, KXD1, SMPDL3B, and ZD17 have been discovered. These factors regulate intracellular membrane dynamics, autophagic processes, and lipid metabolism, ultimately influencing viral genome replication and viral particle assembly.
In the context of antiviral immune responses, two categories of key factors have been identified: one comprises antiviral factors (e.g., STAG2 activates the cGAS-STING signaling pathway, while JADE3 promotes the expression of the antiviral protein IFITM3), and the other includes factors that assist viruses in immune evasion (e.g., HSP90AB1 stabilizes viral proteins, and SLA-DM aids African swine fever virus in evading host immune surveillance) [50].
3.1.2. CRISPR/Cas9 Is Used to Knock out or Finely Edit the ALV and AI Receptor Gene to Develop Antiviral Chickens
Avian Leukosis
Avian Leukosis (AL) is a contagious tumor disease caused by Avian Leukosis virus (ALV), primarily affecting chickens and other poultry [71]. ALV belongs to the retrovirus family and integrates its genome to the host cell DNA, leading to cell transformation and tumor formation ALV is divided into several subgroups (e.g., A, B, C, D, E, and J) based on envelope protein differences, with subgroup J (ALV-J) being highly pathogenic and having a broad host range, making it the most harmful subgroup to the poultry industry in recent years [72]. Different ALV subgroups utilize different host receptors, such as TVA for subgroup A and the chicken type I Na+/H+ exchanger (chNHE1) for subgroup J. W38 is a key amino acid residue in chNHE1, and its deletion may confer resistance to ALV-J [51,73]. Gene editing technology offers a new strategy for controlling ALV by targeting the viral receptor gene or the viral genome to block ALV infection and replication. The CRISPR/Cas9 system, due to its high efficiency and precision, has become the preferred tool for ALV research. Lee et al. used CRISPR/Cas9 to introduce a W38 mutation in chNHE1 in primordial germ cells (PGCs) and transplanted them into rooster reproductive systems, producing W38-deficient gene-edited chickens that were completely resistant to ALV-J [51]. Koslová et al. introduced deletion mutations in TVA and TVC, creating DF-1 cells resistant to ALV-A and ALV-C, respectively [52]. In addition to editing host genes, CRISPR/Cas9 can directly target the ALV genome to disrupt its replication and transmission. Chen et al. used a replication-competent retroviral vector to deliver shRNA-mirs targeting ALV-B, significantly inhibiting ALV-B replication and spread in chicken cells [74].
Avian Influenza
Avian Influenza (AI) is a highly contagious disease caused by the Avian Influenza virus (AIV), which primarily affects poultry and wild birds. AIV belongs to the orthomyxoviridae family and is classified into multiple subtypes (e.g., H5N1, H7N9) based on surface protein hemagglutinin preprotein (HA) and neuraminidase (NA) [75]. Highly pathogenic Avian Influenza viruses (HPAIV) can cause severe respiratory illness and high mortality in poultry, posing a significant threat to global poultry production and public health [76]. AIV enters cells by binding its HA protein to sialic acid receptors on the surface of host cells, releasing RNA genomes, and utilizing host cell mechanisms for replication and transcription. In the process of viral replication, gene rearrangement may occur, leading to the emergence of new subtypes that are more transmissible and pathogenic. Therefore, Jin SePark et al. introduced human β-1,4N-acetylgalactosamine transferase 2 (B4GALNT2) into chicken fibroblasts to modify the sialic acid glycan chain containing α-2,3 ligation by transferring N-acetylgalactosamine to the subterminal galactose of the glycan chain, thereby playing a key role in preventing viruses from binding to the cell surface. Alewo et al. utilized CRISPR/Cas9 to generate homozygous gene-edited (GE) chickens containing two ANP32A amino acid substitutions to prevent viral polymerase interactions. Most vaccination strategies to control AIV have limited effectiveness, so AIV testing remains the main way to control the spread of the disease. Gene editing has been applied to AIV detection. Zhou et al. developed a rapid AIV detection system based on CRISPR/Cas12a, which demonstrates high sensitivity without the need for laboratory equipment [66]. CRISPR/Cas13a-based AIV detection systems have also been reported for H9N2 isoforms with high accuracy.
3.2. Improving Production Performance
As the core pillar of the global animal protein supply, the poultry industry has always been a core goal in the field of breeding to improve its production performance (such as optimization of growth efficiency, improvement of meat quality, and regulation of fat deposition). Although traditional breeding methods have achieved certain results in breed improvement, they have limitations such as long cycle, low accuracy, and strong dependence on genetic background, which are difficult to meet the development needs of modern poultry industry “efficient, high-quality and green”. The rise of gene editing technology, especially the CRISPR/Cas9 system, has provided a revolutionary tool for the precise improvement of poultry production performance—through the targeted modification of key functional genes, it can directly regulate core traits such as poultry growth and development, fat metabolism, etc., and break through the genetic bottleneck of traditional breeding.
3.2.1. Optimization of Growth Efficiency
Growth efficiency is the core economic trait of the poultry industry, which mainly depends on the growth rate, feed conversion rate and muscle growth potential. Relevant research focuses on key genes of growth regulation pathways and achieves precise trait improvement through gene editing.
Nawaz et al. systematically analyzed the genetic regulatory network of poultry growth by integrating genomics, transcriptomics, and single-cell RNA sequencing (scRNA-seq) data. The genetic regulatory network of poultry growth was systematically analyzed. The study clarified that myostatin (MSTN) is a key gene that negatively regulates skeletal muscle growth, and its knockout can lift the inhibition of muscle growth, increase the body weight of chickens by 26.9%, while inducing skeletal muscle hypertrophy and reducing abdominal fat deposition, achieving the dual effect of “muscle gain and fat loss”. In addition, key targets such as IGF2BP1, CAB39L, and LCORL were screened—IGF2BP1 by regulating cell proliferation, differentiation, and metabolism, while poultry weight and muscle weight were affected, and the deletion of promoter regions could significantly promote muscle growth. CAB39L is involved in energy balance and nutrient metabolism; editing this gene can optimize the activity of growth-related pathways. LCORL and LDB2 are localized in the growth-related QTL hotspots of chicken chromosomes 1 and 4, improving the size and growth rate of poultry by regulating mitochondrial function and skeletal muscle development. It was also reported that CRISPR/Cas9 technology was used to edit the above genes in chicken primitive germ cells (PGCs), and a transgenic strain with significantly improved growth rate and feed use efficiency was successfully cultivated, providing a complete technical solution for directed improvement of growth efficiency in targeted lines, including foundational studies often performed on commercial broiler backgrounds such as Ross 308 or specific local breeds (e.g., White Feather broilers) [54].
3.2.2. Improvement of Meat Quality
As a positive regulator of adipocyte differentiation, ZNF423 can promote intramuscular fat deposition by activating the expression of lipogenic marker genes such as PPARG, FABP4, and CEBPA, and intramuscular fat content is positively correlated with meat tenderness, juiciness, and flavor. NR2F2 inhibits excess fat accumulation by specifically binding to the distal promoter region of ZNF423 (−2356 to −2346 bp) through its DNA-binding domain (72–143 aa), thereby down-regulating the transcriptional expression of ZNF423. By regulating the expression balance of the two through gene editing technology, the goal of “reducing abdominal fat deposition and retaining an appropriate amount of intramuscular fat” can be achieved, effectively solving the negative correlation between fat content and meat quality in traditional breeding, and taking into account the growth efficiency and meat quality optimization [55].
PLIN1, as the most abundant structural protein on the surface of lipid droplets, maintains the dynamic balance of lipid storage and decomposition by regulating the binding efficiency of lipases (ATGL, HSL) to neutral lipids. CRISPR/Cas9-mediated PLIN1 gene knockout in chicken preadipocytes can significantly improve meat quality: on the one hand, it increases the proportion of unsaturated fatty acids in intramuscular fat, reduces the content of saturated fatty acids, and improves the nutritional value and flavor of meat. On the other hand, it inhibits excessive fat deposition and avoids the loss of tenderness caused by muscle fibers being wrapped in fat. At the same time, it can indirectly regulate the expression of genes related to muscle fiber development, such as MYOD1 and MYOG, optimize the shape and density of muscle fibers, and further improve the tenderness of meat [56].
The regulatory network composed of myogenic factors such as MYOD1, MYOG, and MYF5 directly participates in the determination and density regulation of skeletal muscle fiber type, regulates their spatiotemporal expression mode through gene editing technology, which can increase the proportion of slow-twitch muscle fibers and significantly improve the tenderness and juiciness of meat. Targeted editing of fatty acid metabolism-related genes such as FADS2 and PRKAG3 can further optimize the fatty acid composition of intramuscular fat, increase the content of unsaturated fatty acids, and enhance the nutritional value of meat [54].
3.2.3. Regulation of Fat Deposition
Li et al. systematically elucidated the mechanism of action of the NR2F2-ZNF423 transcriptional regulatory axis in fat deposition. Overexpression of NR2F2 as an inhibitor can significantly inhibit the differentiation of chicken preadipocytes (oil red O staining showed a decrease in lipid droplet formation); Overexpression of ZNF423 activates the fat differentiation pathway and promotes lipid droplet accumulation. Supplementation with ZNF423 in NR2F2-overexpressing cells can reverse the phenotype of fat deposition inhibition, confirming that the upstream and downstream regulatory relationship between the two is the core switch of fat deposition. Transcriptome analysis showed that PPAR signaling pathway and fatty acid synthesis pathway were significantly enriched in ZNF423 overexpressing cells, providing a pathway-level reference for multi-target co-editing [55].
Zhai et al. targeted the functional validation of the PLIN1 gene, revealing a novel pathway for the regulation of lipid droplet metabolism. PLIN1 balances basal lipolysis with hormone-stimulated lipolysis processes by regulating the activity of key lipolytic enzymes (ATGL, HSL). After CLISPR/Cas9 knockout of PLIN1, chicken preadipocytes showed the characteristics of “enhanced basal lipolysis and hormone-stimulated lipolysis inhibition”: the content of triglycerides (TG) was significantly reduced in the basal state, and the activities of ATGL and HSL were increased. After hormone stimulation, on the contrary, it effectively reduces the excessive accumulation of abdominal fat. At the same time, PLIN1 knockout cell morphology changed from spindle to elliptical, increasing proliferation and reducing early apoptotic activity, further inhibiting adipose tissue hyperproliferation and providing a complete cellular mechanism for the regulation of fat deposition [56].
Nawaz et al. found that growth signaling pathway genes (IGF2BP1, CAB39L) can indirectly affect fat distribution by regulating energy metabolism and nutrient absorption, reducing fat deposition to the abdomen, avoiding the negative correlation between growth efficiency and excessive fat deposition, and providing a multi-target option for “growth-fat” synergistic optimization [54].
3.3. Disease Detection
3.3.1. Salmonella Detection
Salmonella (especially Salmonella typhimurium) is one of the main pathogens causing contamination of poultry products, with high contamination rates and widespread routes, and rapid testing is critical to food safety. Jia et al. developed a CRISPR-SERS biosensor without amplification steps. The sensor targets the invasin A gene (invA) of Salmonella typhimurium, activates trans-cleavage activity after recognizing the target sequence via the Cas12a protein, cleaves the Rox-labeled single-stranded DNA reporter probe fixed on a silver nanoparticle-modified anodized aluminum (AAO) substrate, and then uses surface-enhanced Raman spectroscopy (SERS) to detect signal changes. The platform eliminates the need for DNA amplification steps, has a detection limit as low as 110 CFU/mL, has a recovery rate of 87.4–115.0% in chicken samples, and can specifically distinguish Salmonella from other pathogens such as Escherichia coli and Vibrio parahaemolyticus, and the entire detection process can be completed within 2 h, providing an efficient solution for on-site detection of Salmonella in the poultry production chain [57].
3.3.2. AIV Detection
Avian Influenza virus (AIV) is one of the most harmful poultry viruses with multiple subtypes and rapid mutations, and rapid detection is crucial for epidemic prevention and control. Xu et al. developed a CRISPR-Cas13a-mediated fluorescence detection platform to design specific crRNAs for the hemagglutinin gene (HA) of the Avian Influenza virus H9 subtype, and when viral RNA is present, Cas13a is activated and cleaves the fluorescently labeled RNA reporter probe, releasing a fluorescent signal. The method has a detection limit of 10 copies/μL, a detection time of only 1 h, and can distinguish between H9 subtypes and other Avian Influenza subtypes, making it suitable for rapid on-site screening [77].
3.3.3. NDV Detection
Newcastle disease virus (NDV) is an important pathogen that causes devastating diseases in poultry, Li et al. constructed an electrochemical sensor based on CRISPR-Cas12a, which targets the fusion protein gene (F gene) of Newcastle disease virus, and activates trans-cleavage activity after recognizing the target DNA through Cas12a, cutting the nucleic acid probe on the electrode surface, resulting in changes in electrochemical signals. The sensor has a detection limit of up to 5 copies/μL and can effectively resist matrix interference in poultry tissue samples, providing a reliable solution for the early diagnosis of Newcastle disease virus [78].
3.4. Applications in Commercial Poultry Lines vs. Purebred Lines
A critical consideration in the deployment of gene editing technology is its applicability to commercial poultry lines versus purebred lines. Currently, the vast majority of foundational gene-editing research and successful proof-of-concept studies in poultry are conducted using purebred experimental lines (such as White Leghorn layer lines) or specific laboratory cell lines (such as DF-1). The primary reason is that purebred lines possess well-characterized, stable, and homogenous genetic backgrounds, which facilitate the precise evaluation of the edited gene’s phenotypic effects without the confounding variables of hybrid genetics.
However, modern commercial poultry production relies entirely on highly complex, multi-way crossbred hybrid lines (e.g., commercial broilers like Ross 308 or Cobb 500) to maximize heterosis (hybrid vigor). Direct genetic editing of these commercial terminal hybrids is both technically impractical and biologically inefficient due to their complex genetic segregation and the inability to pass these traits reliably to the next generation (as commercial hybrids are not used for breeding).
Therefore, the successful integration of gene editing into commercial poultry requires a top-down introgression strategy within the commercial breeding pyramid. The target gene edits (e.g., the W38 mutation for ALV-J resistance or ANP32A edits for AIV resistance) must first be generated and established as homozygous traits in the purebred great-grandparent (GGP) or pedigree lines at the apex of the breeding pyramid. Once the desired edit is fixed in these purebred elite lines, the trait can be naturally cascaded down through the grandparent (GP) and parent stock (PS) generations via conventional crossbreeding. This strategy ensures that the millions of commercial birds at the base of the pyramid ultimately inherit the beneficial gene-edited traits (such as enhanced disease resistance) while fully preserving the finely tuned production traits and heterosis of the commercial hybrid lines. Consequently, while the editing process itself is strictly performed on purebred lines, its ultimate impact and economic value are profoundly realized in commercial poultry lines.
4. Risks and Challenges of Gene Editing Technology
Despite significant progress in the application of gene editing technology in agricultural animals, several challenges remain. Off-target effects compromise the biosafety of gene-edited animal products, ethical controversies and social acceptance limit the further market of gene-edited agricultural animals, and the technical complexity demands high expertise from researchers. Developing more precise gene editing tools, stricter off-target detection methods, establishing unified international regulatory frameworks, and creating more efficient delivery systems and simpler operational methods may address these issues.
4.1. Off-Target Effects and Safety
Off-target effects refer to accidental mutations introduced by gene editing tools at non-target sites, which can lead to cellular dysfunction and even cancer, which is a major risk of gene editing technology [79]. Although CRISPR/Cas9 systems offer higher targeting specificity compared to earlier technologies, there are still non-targeting risks in complex genomic environments [80]. Studies have shown that gRNA can bind to sequences with 3–5 base mismatches on the target DNA, causing Cas9 to cleavage at unintended sites, which in turn leads to genomic instability [81].
To reduce off-target effects, the researchers have developed several strategies. For example, high-fidelity Cas9 variants such as eSpCas9 and SpCas9-HF1 optimize Cas9 protein structure and significantly reduce the off-target rate [82]. The double scratch strategy has also been shown to reduce the rate of deviation from the target by 50 to 1500 times [83,84]. In addition, the use of Cas9 Nickase, a mutant that introduces only single-strand breaks rather than double-stranded breaks, can effectively enhance the specificity of editing, which significantly reduces off-target effects compared to traditional Cas9 by requiring two gRNAs to target opposite strands near the target editing site [85].
However, these methods cannot completely eliminate off-target effects. The long-term effects of non-target effects are unclear, especially in the germline and embryo editing areas. Therefore, the development of more accurate gene editing tools and more rigorous off-target detection methods are key directions for future research [86].
4.2. Ethical and Legal Issues
The rapid development of gene editing technology has sparked widespread ethical and legal debates, particularly in the field of human germline and embryo editing. In 2018, He Jiankui’s team announced the successful editing of the CCR5 gene in human embryos, triggering global ethical controversy [87]. Tomita et al. also pointed out that DSB-based gene editing in human embryos may cause chromosomal breaks, large fragment deletions, or rearrangements, and their safety has not been fully verified [88].
This event underscored the complexity of gene editing technology in terms of ethics and law, particularly in areas involving human life origins and genetic modification. The core of the ethical debate lies in the potential misuse and irreversibility of gene editing technology. Since there is no guarantee that gene-edited humans will not produce offspring, the consequences of germline editing may be inherited by future generations, with profound impacts on human society [89]. Additionally, the commercialization of gene editing technology may lead to the emergence of “designer babies,” exacerbating social inequality [90]. In the field of agricultural animals, even if off-target mutations are not introduced, gene editing may have unintended effects on the growth and reproductive performance of individuals.
Therefore, many countries and international organizations have called for strict ethical guidelines and legal frameworks to regulate the application of gene editing technology [91]. Legally, the regulation of gene editing technology is still in its exploratory stages. The European Union and the United States have significantly different regulatory policies for gene-edited crops, with the EU treating them as genetically modified organisms (GMOs) [92]. These policy differences not only limit the application of gene editing technology in animal husbandry but may also lead to international trade disputes. Therefore, establishing a unified international regulatory framework is an important future task [93].
4.3. Technical Complexity
The operational complexity of gene editing technology is one of the main obstacles to its widespread application. First, the design and optimization of gene editing tools require a high level of expertise and experience. For example, the design of sgRNAs must consider factors such as target sequence specificity, GC content, and secondary structure to ensure editing efficiency and accuracy [94].
Efficiently delivering gene editing components to target cells or tissues is also a core technical challenge. Different delivery methods have their respective advantages and limitations, and no universal optimal solution exists. For example, adeno-associated virus (AAV) vectors are limited by packaging capacity, leading researchers to develop dual AAV split strategies or use smaller Cas9 homologs such as SaCas9 and Cas12f [95]. Novel delivery platforms, such as bacterial extracellular contractile injection systems (eCIS), can specifically recognize target cells and deliver large molecules (e.g., SpCas9 protein and zinc-finger deaminases) with high specificity and low immunogenicity [96], providing new approaches for gene editing delivery.
DNA double-strand breaks (DSBs) are primarily repaired via non-homologous end joining (NHEJ) or homology-directed repair (HDR). NHEJ occurs efficiently in all cell cycle stages but tends to introduce random insertions or deletions (indels), whereas HDR requires a donor DNA template and is mostly limited to the S/G2 phase, with lower efficiency than NHEJ [81,95]. For scenarios requiring precise knock-in or point mutation repair, HDR’s low efficiency is a major bottleneck. Recently developed base editors (BEs) and prime editors (PEs) allow single-base conversions or small insertions/deletions without generating DSBs, bypassing HDR limitations, though they still have constraints such as deaminase-mediated off-target effects and limited editing windows. Notably, the NICER technique introduces multiple single-strand nicks near heterozygous mutation sites using nCas9, leveraging homologous chromosomes as endogenous repair templates for precise correction, with minimal DSB-related unintended mutations [88].
5. Conclusions
Gene editing technologies, particularly CRISPR/Cas9 systems, have shown significant potential in agricultural animal research. By precisely editing genomes, disease resistance, productivity, and disease detection can be rapidly improved. It has shown significant accuracy and efficiency in improving disease resistance (e.g., ALV and AIV) [97]. In studies aimed at improving animal performance, editing the MSTN gene reduced the deposition of avian fat in the abdomen to improve muscle mass and increase palatability [55]. These achievements not only improve the economic efficiency of livestock farming but also provide new solutions to global food security challenges. Although significant advances have been made in the application of gene editing technology in agricultural animals, it still faces several challenges, including off-target effects, ethical controversies, technical complexity, and social acceptance. Off-target effects are a major challenge of gene editing techniques. Reducing non-target effects requires more precise gene editing tools such as Cas9 variants, as well as the development of base and prime editors. In addition, more efficient delivery systems are needed to improve the efficiency and specificity of gene editing. Novel nanomaterial carriers, such as gold nanoparticles and metal–organic frameworks, have high loading capacity, good biocompatibility, and controlled release properties, which are key factors driving the application of CRISPR/Cas9 systems [98]. Cell-penetrating peptides (CPPs) enter cells through electrostatic interactions or endocytosis, making them suitable for delivering various macromolecules and cells, with low immunogenicity and toxicity. CPP-based delivery systems have great potential for in vivo applications [99]. The development of gene editing technology relies on interdisciplinary research. For example, integrated genomics, proteomics, and metabolomics can comprehensively assess the impact and potential risks of gene editing. Combining artificial intelligence and big data technology to optimize the design and delivery strategies of gene editing tools can improve editing efficiency and accuracy [100]. The ethical and legal controversies surrounding gene editing technology are also major obstacles to its widespread adoption. Genetic modification introduced into animals can have permanent deleterious effects on species unless genetic programs are used continuously to prevent deleterious mutations from entering the gene pool. The commercialization of gene editing technology may affect existing animal welfare. For example, some traits produced by gene editing can occur naturally in populations, while profit-driven genetic modification may ignore existing animal welfare issues [101]. Establishing a unified international regulatory framework to ensure the safety and fairness of gene editing technology can prevent unnecessary suffering in animals. In addition, strengthening public education and participation in gene editing technology is an important way to increase social acceptance. Gene editing technology has made significant progress in poultry breeding, providing new solutions to improve disease resistance, production performance, animal welfare, and reduce environmental pollution. However, gene editing technology still faces many challenges, including target bias, ethical controversies, technical complexity, and social acceptance. In the future, with the continuous advancement and optimization of technology, gene editing technology is expected to achieve wider applications in the field of agricultural animals and make important contributions to global food security and sustainable development.
Author Contributions
Conceptualization, R.L. and Q.W.; Literature search, R.L.; Literature curation, R.R. and Y.L.; Writing—original draft, R.L.; Writing—review and editing, R.R., M.T., Y.L. and H.W.; Visualization, R.R. and M.T.; Data verification, X.Z.; Technical input, X.Z.; Project oversight, Q.W.; Supervision, H.W.; Funding acquisition, X.L.; Manuscript structuring, X.L.; Final editing, X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by the Project of Chongqing Agricultural Science Innovation (NW202209), the Fundamental Research Funds for the Central Universities (XDJK2020C018), Chongqing Science and Enterprise Consortium Germplasm Resources Collection and Utilization and Variety Trial (KQ202301), Breeding of New Breeds (Synthetic Line) of High-Quality Chickens for Both Meat and Eggs (cstc2021 jscx-gksbX0004), and the National Natural Science Foundation of China (31802054), 2025 Chongqing Municipal College Student Innovation and Entrepreneurship Training Program (S202510635193).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study.
Acknowledgments
We sincerely thank the members of the College of Animal Science and Technology at Southwest University and the Chongqing Academy of Animal Sciences for their valuable discussions and technical support, and also thank the Figdraw platform for its assistance in creating the schematic diagrams.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
The following abbreviations are used in this manuscript:
| ZFN | Zinc Finger Nucleases |
| TALEN | Transcription Activator-Like Effector Nucleases |
| CRISPR | Clustered Regularly Interspaced Short Palindromic Repeats |
| Cas9 | CRISPR-associated protein 9 |
| DSBs | Double-Strand Breaks |
| NHEJ | Non-Homologous End Joining |
| HDR | Homology-Directed Repair |
| nCas9 | Nickase Cas9 |
| dCas9 | Dead Cas9 |
| PAM | Protospacer Adjacent Motif |
| crRNA | CRISPR RNA |
| tracrRNA | Trans-activating |
| gRNA | Guide RNA |
| ssODN | Single-Stranded Oligonucleotide |
| CBEs | Cytosine Base Editors |
| ABEs | Adenine Base Editors |
| GBEs | Guanine Base Editors |
| PEs | Prime Editors |
| pegRNA | Prime Editing Guide RNA |
| LNP | Lipid Nanoparticles |
| PEI | Polyethylenimine |
| PLGA | Poly (lactic-co-glycolic acid) |
| MOFs | Metal–Organic Frameworks |
| CPPs | Cell-Penetrating Peptides |
| PRRSV | Porcine Reproductive and Respiratory Syndrome Virus |
| CD163 | Cluster of Differentiation 163 |
| PRNP | Prion Protein Gene |
| MSTN | Myostatin |
| BLG | Beta-lactoglobulin |
| FGF5 | Fibroblast Growth Factor 5 |
| ATGL | Adipose Triglyceride Lipase |
| G0S2 | G0/G1 Switch Gene 2 |
| CH4 | Methane |
| TB | Tuberculosis |
| NRAMP1 | Natural Resistance-Associated Macrophage Protein 1 |
| SP110 | Sp110 Nuclear Body Protein |
| LEPR | Leptin Receptor |
| KISS1 | Kisspeptin1 |
| ALV | Avian Leukosis Virus |
| AIV | Avian Influenza Virus |
| HA | Hemagglutinin |
| NA | Neuraminidase |
| HPAIV | Highly Pathogenic Avian Influenza Virus |
| RNA | Ribonucleic Acid |
| DNA | Deoxyribonucleic Acid |
| mRNA | Messenger RNA |
| sgRNA | Single Guide RNA |
| shRNA | Short Hairpin RNA |
| miRNA | MicroRNA |
| siRNA | Small Interfering RNA |
| CRISPRa | CRISPR Activation |
| AAV | Adeno-Associated Virus |
| ALV-J | Avian Leukosis Virus Subgroup J |
| NDV | Newcastle Disease Virus |
| PGC | Primordial Germ Cell |
| SCNT | Somatic Cell Nuclear Transfer |
| SERS | Surface-Enhanced Raman Scattering |
| Rox | 6-Carboxy-X-Rhodamine |
| PS | Phosphate-Buffered Saline |
| GGP | Grandparent Generation Poultry |
| GP | Germline Progenito |
References
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Komal; Kaur, P.; Arora, N.; Sawale, J.A.; Singh, A. Transformative CRISPR-Cas9 Technologies: A Review of Molecular Mechanisms, Precision Editing Techniques, and Clinical Progress in Sickle Cell Disease. Curr. Drug Metab. 2024, 25, 734–741. [Google Scholar] [CrossRef] [PubMed]
- Gong, T.; Liu, D.; Wang, X.; Zhou, D.; Tang, L.; Wang, H.; Su, J.; Liang, P. Establishment of a CIB1 knockout human pluripotent stem cell line via CRISPR/Cas9 genome editing technology. Stem Cell Res. 2024, 81, 103580. [Google Scholar] [CrossRef]
- Gao, C. Genome engineering for crop improvement and future agriculture. Cell 2021, 184, 1621–1635. [Google Scholar] [CrossRef]
- Smith, H.O.; Welcox, K.W. A restriction enzyme from Hemophilus influenzae. J. Mol. Biol. 1970, 51, 379–391. [Google Scholar] [CrossRef]
- Jackson, D.A.; Symons, R.H.; Berg, P. Biochemical method for inserting new genetic information into DNA of simian virus 40: Circular SV40 DNA molecules containing lambda phage genes and the galactose operon of Escherichia coli. Proc. Natl. Acad. Sci. USA 1972, 69, 2904–2909. [Google Scholar] [CrossRef]
- Capecchi, M.R. Targeted gene replacement. Sci. Am. 1994, 270, 52–59. [Google Scholar] [CrossRef]
- Wilmut, I.; Schnieke, A.E.; McWhir, J.; Kind, A.J.; Campbell, K.H. Viable offspring derived from fetal and adult mammalian cells. Nature 1997, 385, 810–813. [Google Scholar] [CrossRef]
- Bibikova, M.; Carroll, D.; Segal, D.J.; Trautman, J.K.; Smith, J.; Kim, Y.-G.; Chandrasegaran, S. Stimulation of homologous recombination through targeted cleavage by chimeric nucleases. Mol. Cell. Biol. 2001, 21, 289–297. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.C.; Tan, S.; Qiao, G.; Barlow, K.A.; Wang, J.; Xia, D.F.; Meng, X.; Paschon, D.E.; Leung, E.; Hinkley, S.J.; et al. A TALE nuclease architecture for efficient genome editing. Nat. Biotechnol. 2011, 29, 143–148. [Google Scholar] [CrossRef]
- Mali, P.; Yang, L.; Esvelt, K.M.; Aach, J.; Guell, M.; DiCarlo, J.E.; Norville, J.E.; Church, G.M. RNA-guided human genome engineering via Cas9. Science 2013, 339, 823–826. [Google Scholar] [CrossRef]
- Dimitrov, L.; Pedersen, D.; Ching, K.H.; Yi, H.; Collarini, E.J.; Izquierdo, S.; Van De Lavoir, M.-C.; Leighton, P.A. Germline gene editing in chickens by efficient CRISPR-mediated homologous recombination in primordial germ cells. PLoS ONE 2016, 11, e0154303. [Google Scholar] [CrossRef]
- Oishi, I.; Yoshii, K.; Miyahara, D.; Kagami, H.; Tagami, T. Targeted mutagenesis in chicken using CRISPR/Cas9 system. Sci. Rep. 2016, 6, 23980. [Google Scholar] [CrossRef]
- Komor, A.C.; Kim, Y.B.; Packer, M.S.; Zuris, J.A.; Liu, D.R. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature 2016, 533, 420–424. [Google Scholar] [CrossRef]
- Gootenberg, J.S.; Abudayyeh, O.O.; Lee, J.W.; Essletzbichler, P.; Dy, A.J.; Joung, J.; Verdine, V.; Donghia, N.; Daringer, N.M.; Freije, C.A.; et al. Nucleic acid detection with CRISPR-Cas13a/C2c2. Science 2017, 356, 438–442. [Google Scholar] [CrossRef]
- Zhong, Z.; Zhang, Y.; You, Q.; Tang, X.; Ren, Q.; Liu, S.; Yang, L.; Wang, Y.; Liu, X.; Liu, B.; et al. Plant genome editing using FnCpf1 and LbCpf1 nucleases at redefined and altered PAM sites. Mol. Plant 2018, 11, 999–1002. [Google Scholar] [CrossRef] [PubMed]
- Anzalone, A.V.; Randolph, P.B.; Davis, J.R.; Sousa, A.A.; Koblan, L.W.; Levy, J.M.; Chen, P.J.; Wilson, C.; Newby, G.A.; Raguram, A.; et al. Search-and-replace genome editing without double-strand breaks or donor DNA. Nature 2019, 576, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Proudfoot, C.; Lillico, S.; Tait-Burkard, C. Genome editing for disease resistance in pigs and chickens. Anim. Front. Rev. Mag. Anim. Agric. 2019, 9, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Koslová, A.; Trefil, P.; Mucksová, J.; Reinišová, M.; Plachý, J.; Kalina, J.; Kučerová, D.; Geryk, J.; Krchlíková, V.; Lejčková, B.; et al. Precise CRISPR/Cas9 editing of the NHE1 gene renders chickens resistant to the J subgroup of avian leukosis virus. Proc. Natl. Acad. Sci. USA 2020, 117, 2108–2112. [Google Scholar] [CrossRef]
- Mehta, J. CRISPR-Cas9 gene editing for sickle cell disease and beta-thalassemia. N. Engl. J. Med. 2021, 384, 252–260. [Google Scholar]
- Ishino, Y.; Shinagawa, H.; Makino, K.; Amemura, M.; Nakata, A. Nucleotide sequence of the iap gene, responsible for alkaline phosphatase isozyme conversion in Escherichia coli, and identification of the gene product. J. Bacteriol. 1987, 169, 5429–5433. [Google Scholar] [CrossRef]
- Jiang, S.; Shen, Q.W. Principles of gene editing techniques and applications in animal husbandry. 3 Biotech 2019, 9, 28. [Google Scholar] [CrossRef]
- Deltcheva, E.; Chylinski, K.; Sharma, C.M.; Gonzales, K.; Chao, Y.; Pirzada, Z.A.; Eckert, M.R.; Vogel, J.; Charpentier, E. CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III. Nature 2011, 471, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Fineran, P.C.; Charpentier, E. Memory of viral infections by CRISPR-cas adaptive immune systems: Acquisition of new information. Virology 2012, 434, 202–209. [Google Scholar] [CrossRef]
- Ledford, H.; Callaway, E. Pioneers of revolutionary CRISPR gene editing win chemistry nobel. Nature 2020, 586, 346–347. [Google Scholar] [CrossRef]
- Cong, L.; Ran, F.A.; Cox, D.; Lin, S.; Barretto, R.; Habib, N.; Hsu, P.D.; Wu, X.; Jiang, W.; Marraffini, L.A. Multiplex genome engineering using CRISPR/cas systems. Science 2013, 339, 819–823. [Google Scholar] [CrossRef]
- Ma, S.; Chang, J.; Wang, X.; Liu, Y.; Zhang, J.; Lu, W.; Gao, J.; Shi, R.; Zhao, P.; Xia, Q. CRISPR/Cas9 mediated multiplex genome editing and heritable mutagenesis of BmKu70 in Bombyx mori. Sci. Rep. 2014, 4, 4489. [Google Scholar] [CrossRef]
- Ran, F.A.; Hsu, P.D.; Lin, C.-Y.; Gootenberg, J.S.; Konermann, S.; Trevino, A.E.; Scott, D.A.; Inoue, A.; Matoba, S.; Zhang, Y.; et al. Double nicking by RNA-guided CRISPR Cas9 for enhanced genome editing specificity. Cell 2013, 154, 1380–1389. [Google Scholar] [CrossRef]
- Khadempar, S.; Familghadakchi, S.; Motlagh, R.A.; Farahani, N.; Dashtiahangar, M.; Rezaei, H.; Gheibi Hayat, S.M. CRISPR-Cas9 in genome editing: Its function and medical applications. J. Cell. Physiol. 2019, 234, 5751–5761. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, S.J.; Ekman, M.B.; Manek, S.; Galandiuk, S. CRISPR-mediated gene editing for the surgeon scientist. Surgery 2019, 166, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, L.A.; Larson, M.H.; Morsut, L.; Liu, Z.; Brar, G.A.; Torres, S.E.; Stern-Ginossar, N.; Brandman, O.; Whitehead, E.H.; Doudna, J.A.; et al. CRISPR-mediated modular RNA-guided regulation of transcription in eukaryotes. Cell 2013, 154, 442–451. [Google Scholar] [CrossRef]
- Li, S.-Y.; Cheng, Q.-X.; Liu, J.-K.; Nie, X.-Q.; Zhao, G.-P.; Wang, J. CRISPR-Cas12a has both cis- and trans-cleavage activities on single-stranded DNA. Cell Res. 2018, 28, 491–493. [Google Scholar] [CrossRef]
- Swarts, D.C.; Jinek, M. Mechanistic insights into the cis- and trans-acting DNase activities of Cas12a. Mol. Cell 2019, 73, 589–600.e4. [Google Scholar] [CrossRef] [PubMed]
- Abudayyeh, O.O.; Gootenberg, J.S.; Konermann, S.; Joung, J.; Slaymaker, I.M.; Cox, D.B.T.; Shmakov, S.; Makarova, K.S.; Semenova, E.; Minakhin, L.; et al. C2c2 is a single-component programmable RNA-guided RNA-targeting CRISPR effector. Science 2016, 353, aaf5573. [Google Scholar] [CrossRef]
- San Filippo, J.; Sung, P.; Klein, H. Mechanism of eukaryotic homologous recombination. Annu. Rev. Biochem. 2008, 77, 229–257. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Staahl, B.T.; Alla, R.K.; Doudna, J.A. Enhanced homology-directed human genome engineering by controlled timing of CRISPR/Cas9 delivery. eLife 2014, 3, e04766. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chen, P.J.; Hussmann, J.A.; Yan, J.; Knipping, F.; Ravisankar, P.; Chen, P.-F.; Chen, C.; Nelson, J.W.; Newby, G.A.; Sahin, M.; et al. Enhanced prime editing systems by manipulating cellular determinants of editing outcomes. Cell 2021, 184, 5635–5652.e29. [Google Scholar] [CrossRef]
- Kwon, M.; Firestein, B.L. DNA transfection: Calcium phosphate method. Methods Mol. Biol. 2013, 1018, 107–110. [Google Scholar] [CrossRef]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. DEAE-Dextran Transfection; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2018. [Google Scholar] [CrossRef]
- Zhao, J.; Lever, A.M.L. Lentivirus-mediated gene expression. Methods Mol. Biol. 2007, 366, 343–355. [Google Scholar] [CrossRef]
- Temin, H.M. Retrovirus vectors: Promise and reality. Science 1989, 246, 983. [Google Scholar] [CrossRef]
- Zhang, D.; Das, D.B.; Rielly, C.D. Potential of microneedle-assisted micro-particle delivery by gene guns: A review. Drug Deliv. 2014, 21, 571–587. [Google Scholar] [CrossRef]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Electroporation. Cold Spring Harb. Protoc. 2019, 2019, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Gahl, R.F. Microinjection of live mammalian cells: A delivery method that provides added versatility to the study of cellular function. In Microinjection: Methods and Protocols; Liu, C., Du, Y., Eds.; Springer: New York, NY, USA, 2019; pp. 525–536. [Google Scholar]
- Leclerc, D.; Siroky, M.D.; Miller, S.M. Next-generation biological vector platforms for in vivo delivery of genome editing agents. Curr. Opin. Biotechnol. 2024, 85, 11. [Google Scholar] [CrossRef]
- Issa, S.S.; Shaimardanova, A.A.; Solovyeva, V.V.; Rizvanov, A.A. Various AAV serotypes and their applications in gene therapy: An overview. Cells 2023, 12, 785. [Google Scholar] [CrossRef] [PubMed]
- Hirschenberger, M.; Stadler, N.; Fellermann, M.; Sparrer, K.M.J.; Kirchhoff, F.; Barth, H.; Papatheodorou, P. CRISPA: A non-viral, transient Cas9 delivery system based on reengineered anthrax toxin. Front. Pharmacol. 2021, 12, 770283. [Google Scholar] [CrossRef]
- Allemailem, K.S.; Alsahli, M.A.; Almatroudi, A.; Alrumaihi, F.; Alkhaleefah, F.K.; Rahmani, A.H.; Khan, A.A. Current updates of CRISPR/Cas9-mediated genome editing and targeting within tumor cells: An innovative strategy of cancer management. Cancer Commun. 2022, 42, 1257–1287. [Google Scholar] [CrossRef]
- Nakase, I.; Tanaka, G.; Futaki, S. Cell-penetrating peptides (CPPs) as a vector for the delivery of siRNAs into cells. Mol. Biosyst. 2013, 9, 855–861. [Google Scholar] [CrossRef]
- Wang, K.; Ouyang, H.; Xie, Z.; Yao, C.; Guo, N.; Li, M.; Jiao, H.; Pang, D. Efficient generation of myostatin mutations in pigs using the CRISPR/Cas9 sys-tem. Sci. Rep. 2015, 5, 16623. [Google Scholar] [CrossRef] [PubMed]
- Carlson, D.F.; A Lancto, C.; Zang, B.; Kim, E.-S.; Walton, M.; Oldeschulte, D.; Seabury, C.; Sonstegard, T.S.; Fahrenkrug, S.C. Production of hornless dairy cattle from genome-edited cell lines. Nat. Biotechnol. 2016, 34, 479–481. [Google Scholar]
- Lee, H.J.; Lee, , K.Y.; Jung, , K.M.; Park, , K.J.; Lee, , K.O.; Suh, , J.-Y.; Yao, , Y.; Nair, , V.; Han, J.Y. Precise gene editing of chicken Na+/H+ exchange type 1 (chNHE1) confers resistance to avian leukosis virus subgroup J (ALV-J). Dev. Comp. Immunol. 2017, 77, 340-349. [Google Scholar] [CrossRef]
- Burkard, C.; Lillico, S.G.; Reid, E.; Jackson, B.; Mileham, A.J.; Ait-Ali, T.; Whitelaw, C.B.; Archibald, A.L. Precision engineering for PRRSV resistance in pigs: Macrophages from genome edited pigs lacking CD163 SRCR5 domain are fully resistant to both PRRSV genotypes while maintaining biological function. PLoS Pathog. 2017, 13, e1006206. [Google Scholar]
- Sun, Z.; Wang, M.; Han, S.; Ma, S.; Zou, Z.; Ding, F.; Li, X.; Li, L.; Tang, B.; Wang, H.; et al. Production of hypoallergenic milk from DNA-free beta-lactoglobulin (BLG) gene knockout cow using zinc-finger nucleases mRNA. Sci. Rep. 2018, 8, 15430. [Google Scholar]
- Chen, J.S.; Ma, E.; Harrington, L.B. CRISPR-Cas12a target binding unleashes indiscriminate single-stranded DNase activity. Science 2018, 360, 436–439. [Google Scholar]
- Koslová, A.; Kučerová, D.; Reinišová, M.; Geryk, J.; Trefil, P.; Hejnar, J. Genetic Resistance to Avian Leukosis Viruses Induced by CRISPR/Cas9 Editing of Specific Receptor Genes in Chicken Cells. Viruses 2018, 10, 605. [Google Scholar] [CrossRef]
- Tanihara, F.; Hirata, M.; Nguyen, N.T.; Sawamoto, O.; Kikuchi, T.; Otoi, T. One-Step Generation of Multiple Gene-Edited Pigs by Electroporation of the CRISPR/Cas9 System into Zygotes to Reduce Xenoantigen Biosynthesis. Int. J. Mol. Sci. 2021, 22, 2249. [Google Scholar] [CrossRef]
- Flórez, J.M.; Martins, K.; Solin, S.; Bostrom, J.R.; Rodríguez-Villamil, P.; Ongaratto, F.; Larson, S.A.; Ganbaatar, U.; Coutts, A.W.; Kern, D.; et al. CRISPR/Cas9-editing of KISS1 to generate pigs with hypogonadotropic hypogonadism as a castration free trait. Front. Genet. 2023, 13, 1078991. [Google Scholar] [CrossRef]
- Idoko-Akoh, A.; Goldhill, D.H.; Sheppard, C.M.; Bialy, D.; Quantrill, J.L.; Sukhova, K.; Brown, J.C.; Richardson, S.; Campbell, C.; Taylor, L.; et al. Creating resistance to avian influenza infection through genome editing of the ANP32 gene family. Nat. Commun. 2023, 14, 1–15. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, S.; Ma, Y.; Li, Y.; Deng, G.; Shi, J.; Wang, X. Rapid detection of avian influenza virus based on CRISPR-Cas12a. Virol. J. 2023, 20, 1–10. [Google Scholar] [CrossRef]
- Cuellar, C.J.; Amaral, T.F.; Rodriguez-Villamil, P.; Ongaratto, F.; Martinez, D.O.; Labrecque, R.; Losano, J.D.d.A.; Estrada-Cortés, E.; Bostrom, J.R.; Martins, K.; et al. Consequences of gene editing of PRLR on thermotolerance, growth, and male reproduction in cattle. FASEB BioAdvances 2024, 6, 223–234. [Google Scholar] [CrossRef]
- Feng, R.; Zhao, J.; Zhang, Q.; Zhu, Z.; Zhang, J.; Liu, C.; Zheng, X.; Wang, F.; Su, J.; Ma, X.; et al. Generation of Anti-Mastitis Gene-Edited Dairy Goats with Enhancing Lysozyme Expression by Inflammatory Regulatory Sequence using ISDra2-TnpB System. Adv. Sci. 2024, 11, e2404408. [Google Scholar] [CrossRef]
- Liu, P.; Jiang, J.; Chen, Y.; Gao, F.; Wang, S.; Yu, M.; Liu, Y.; Guo, R.; Zhang, L.; Xu, Z.; et al. Identification of Cables1 as a critical host factor that promotes ALV-J replication via genome-wide CRISPR/Cas9 gene knockout screening. J. Biol. Chem. 2024, 300, 107804. [Google Scholar] [CrossRef]
- Ding, J.; Liu, X.; Nawaz, A.H.; Leng, D.; Li, N.; Ge, D.; Li, D.; Feng, C. Integrative multi-omics analysis deciphers the regulatory mechanisms of egg weight traits in chickens. Poult. Sci. 2025, 104, 105813. [Google Scholar] [CrossRef]
- Hu, S.; Gan, M.; Wei, Z.; Shang, P.; Song, L.; Feng, J.; Chen, L.; Niu, L.; Wang, Y.; Zhang, S.; et al. Identification of host factors for livestock and poultry viruses: Genome-wide screening technology based on the CRISPR system. Front. Microbiol. 2024, 15, 1498641. [Google Scholar] [CrossRef]
- Park, J.S.; Lee, K.Y.; Han, J.Y. Precise Genome Editing in Poultry and Its Application to Industries. Genes 2020, 11, 1182. [Google Scholar] [CrossRef]
- Nawaz, A.H.; Ding, J.; Ali, M.; Leng, D.; Mukhtar, N.; Ali, A.; Feng, C. Decoding chicken growth regulation through multi-omics insights and emerging genetic tools for growth optimization. Poult. Sci. 2025, 104, 105542. [Google Scholar] [CrossRef]
- Li, X.; Sun, D.; Wang, Z.; Zhao, Q.; Liu, Y.; Hou, Z. Transcriptional regulatory mechanism of NR2F2 and ZNF423 in avian preadipocyte differentiation. Gene 2023, 897, 148106. [Google Scholar] [CrossRef]
- Zhai, G.; Pang, Y.; Zou, Y.; Wang, X.; Liu, J.; Zhang, Q.; Cao, Z.; Wang, N.; Li, H.; Wang, Y. Effects of PLIN1 Gene Knockout on the Proliferation, Apoptosis, Differentiation and Lipolysis of Chicken Preadipocytes. Animals 2022, 13, 92. [Google Scholar] [CrossRef]
- Jia, F.; Li, B.; He, Y.; Shen, Y.; Chen, J.; Li, X.; Li, Y. An amplification-free CRISPR-SERS biosensor for specific, sensitive and rapid detection of salmonella typhimurium in poultry. Food Sci. Techno. 2023, 189, 8. [Google Scholar]
- Wang, L.; Xie, T.; Zhou, X.; Yang, G.; Guo, Z.; Huang, Y.; Lamont, S.J.; Lan, X. LncIRF1 promotes chicken resistance to ALV-J infection. 3 Biotech 2023, 13, 367. [Google Scholar] [CrossRef]
- Xu, M.; Qian, K.; Shao, H.; Yao, Y.; Nair, V.; Ye, J.; Qin, A. 3′UTR of ALV-J can affect viral replication through promoting transcription and mRNA nuclear export. J. Virol. 2023, 97, e01152-23. [Google Scholar] [CrossRef] [PubMed]
- Feng, W.; Meng, W.; Cai, L.; Cui, X.; Pan, Z.; Wang, G.; Cheng, Z. Avian leukosis virus subgroup J induces its receptor--chNHE1 up-regulation. Virol. J. 2016, 13, 58. [Google Scholar] [CrossRef]
- Chen, M.; Granger, A.J.; VanBrocklin, M.W.; Payne, W.S.; Hunt, H.; Zhang, H.; Dodgson, J.B.; Holmen, S.L. Inhibition of avian leukosis virus replication by vector-based RNA interference. Virology 2007, 365, 464–472. [Google Scholar] [CrossRef]
- Spackman, E. A brief introduction to avian influenza virus. In Animal Influenza Virus; Spackman, E., Ed.; Springer: New York, NY, USA, 2020; Volume 2123, pp. 83–92. [Google Scholar]
- Alexander, D.J. An overview of the epidemiology of avian influenza. Vaccine 2007, 25, 5637–5644. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Wang, R.; Li, Y. Electrochemical biosensors for rapid detection of Escherichia coli O157:H7. Talanta 2017, 162, 511–522. [Google Scholar] [CrossRef]
- Li, F.; Ye, Q.; Chen, M.; Zhou, B.; Zhang, J.; Pang, R.; Xue, L.; Wang, J.; Zeng, H.; Wu, S.; et al. An ultrasensitive CRISPR/Cas12a based electrochemical biosensor for Listeria monocytogenes detection. Biosens. Bioelectron. 2021, 179, 113073. [Google Scholar] [CrossRef]
- Chen, S.-J. Minimizing off-target effects in CRISPR-Cas9 genome editing. Cell Biol. Toxicol. 2019, 35, 399–401. [Google Scholar] [CrossRef]
- Kim, H.; Lee, W.; Oh, Y.; Kang, S.-H.; Hur, J.K.; Lee, H.; Song, W.; Lim, K.-S.; Park, Y.-H.; Song, B.-S.; et al. Enhancement of target specificity of CRISPR–Cas12a by using a chimeric DNA–RNA guide. Nucleic Acids Res. 2020, 48, 8601–8616. [Google Scholar] [CrossRef]
- Barman, A.; Deb, B.; Chakraborty, S. A glance at genome editing with CRISPR-Cas9 technology. Curr. Genet. 2020, 66, 447–462. [Google Scholar] [CrossRef]
- Tyumentseva, M.; Tyumentsev, A.; Akimkin, V. CRISPR/Cas9 Landscape: Current State and Future Perspectives. Int. J. Mol. Sci. 2023, 24, 16077. [Google Scholar] [CrossRef]
- Shen, B.; Zhang, W.; Zhang, J.; Zhou, J.; Wang, J.; Chen, L.; Wang, L.; Hodgkins, A.; Iyer, V.; Huang, X.; et al. Efficient genome modification by CRISPR-Cas9 nickase with minimal off-target effects. Nat. Methods 2014, 11, 399–402. [Google Scholar] [CrossRef]
- Fu, Y.; Foden, J.A.; Khayter, C.; Maeder, M.L.; Reyon, D.; Joung, J.K.; Sander, J.D. High-frequency off-target mutagenesis induced by CRISPR-cas nucleases in human cells. Nat. Biotechnol. 2013, 31, 822–826. [Google Scholar] [CrossRef]
- Gallala, M. Application of CRISPR/Cas gene editing for infectious disease control in poultry. Open Life Sci. 2025, 20, 20251095. [Google Scholar] [CrossRef]
- Asmamaw Mengstie, M.; Teshome Azezew, M.; Asmamaw Dejenie, T.; Teshome, A.A.; Tadele Admasu, F.; Behaile Teklemariam, A.; Tilahun Mulu, A.; Mekonnen Agidew, M.; Adugna, D.G.; Geremew, H.; et al. Recent Advancements in Reducing the Off-Target Effect of CRISPR-Cas9 Genome Editing. Biol. Targets Ther. 2024, 18, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Cyranoski, D. The CRISPR-baby scandal: What’s next for human gene-editing. Nature 2019, 566, 440–442. [Google Scholar] [CrossRef]
- Tomita, A.; Sasanuma, H.; Owa, T.; Nakazawa, Y.; Shimada, M.; Fukuoka, T.; Ogi, T.; Nakada, S. Inducing multiple nicks promotes interhomolog homologous recombination to correct heterozygous mutations in somatic cells. Nat. Commun. 2023, 14, 5607. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, G.O.; Labude, M.K.; Zhu, Y.; Foo, R.S.-Y.; Xafis, V. International reporting mechanism for unethical germline gene editing experiments is needed. Trends Biotechnol. 2021, 39, 427–430. [Google Scholar] [CrossRef] [PubMed]
- Ormond, K.E.; Bombard, Y.; Bonham, V.L.; Hoffman-Andrews, L.; Howard, H.; Isasi, R.; Musunuru, K.; Riggan, K.A.; Michie, M.; Allyse, M. The clinical application of gene editing: Ethical and social issues. Pers. Med. 2019, 16, 337–350. [Google Scholar] [CrossRef]
- A, F.M.; B, A.N.; B, I.P. CRISPR/Cas9 gene-editing: Research technologies, clinical applications and ethical considerations—ScienceDirect. Semin. Perinatol. 2018, 42, 487–500. [Google Scholar]
- Eckerstorfer, M.F.; Dolezel, M.; Heissenberger, A.; Miklau, M.; Reichenbecher, W.; Steinbrecher, R.A.; Waßmann, F. An EU Perspective on Biosafety Considerations for Plants Developed by Genome Editing and Other New Genetic Modification Techniques (nGMs). Front. Bioeng. Biotechnol. 2019, 7, 31. [Google Scholar] [CrossRef] [PubMed]
- Lei, R.; Qiu, R. Ethical and regulatory issues in human gene editing: Chinese perspective. Biotechnol. Appl. Biochem. 2020, 67, 880–891. [Google Scholar] [CrossRef]
- Cai, W.; Liu, J.; Chen, X.; Mao, L.; Wang, M. Orthogonal chemical activation of enzyme-inducible CRISPR/Cas9 for cell-selective genome editing. J. Am. Chem. Soc. 2022, 144, 22272–22280. [Google Scholar] [CrossRef] [PubMed]
- van Haasteren, J.; Li, J.; Scheideler, O.J.; Murthy, N.; Schaffer, D.V. The delivery challenge: Fulfilling the promise of therapeutic genome editing. Nat. Biotechnol. 2020, 38, 845–855. [Google Scholar] [CrossRef]
- Kreitz, J.; Friedrich, M.J.; Guru, A.; Lash, B.; Saito, M.; Macrae, R.K.; Zhang, F. Programmable protein delivery with a bacterial contractile injection system. Nature 2023, 616, 357–364. [Google Scholar] [CrossRef]
- Wu, H.; Wang, Y.; Zhang, Y.; Yang, M.; Lv, J.; Liu, J.; Zhang, Y. TALE nickase-mediated SP110 knockin endows cattle with increased resistance to tuberculosis. Proc. Natl. Acad. Sci. USA 2015, 112, E1530-1539. [Google Scholar] [CrossRef]
- Hii, A.R.K.; Qi, X.; Wu, Z. Advanced strategies for CRISPR/Cas9 delivery and applications in gene editing, therapy, and cancer detection using nanoparticles and nanocarriers. J. Mater. Chem. B 2024, 12, 1467–1489. [Google Scholar] [CrossRef]
- Patel, P.; Benzle, K.; Pei, G.L. Cell-penetrating peptides for sustainable agriculture. Trends Plant Sci. 2024, 29, 1131–1144. [Google Scholar] [CrossRef]
- Abbasi, A.F.; Asim, M.N.; Dengel, A. Transitioning from wet lab to artificial intelligence: A systematic review of AI predictors in CRISPR. J. Transl. Med. 2025, 23, 153. [Google Scholar] [CrossRef]
- Shriver, A. Prioritizing the protection of welfare in gene-edited livestock. Anim. Front. Rev. Mag. Anim. Agric. 2020, 10, 39–44. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.