Corn Bioethanol Side Streams: A Potential Sustainable Source of Fat-Soluble Bioactive Molecules for High-Value Applications

 ,
,  , ,
, ,
Abstract
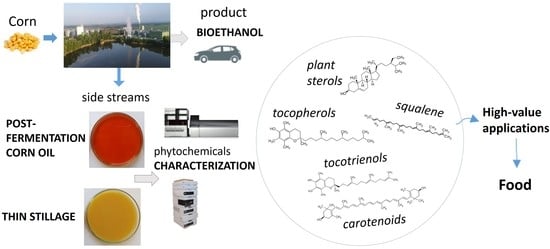
1. Introduction
2. Materials and Methods
2.1. Collection of Side Streams
2.2. Chemicals
2.3. Sample Treatment
2.4. Chemical Analyses
2.4.1. Analytical Procedures
2.4.2. High-Performance-Liquid-Chromatography (HPLC)
2.4.3. Liquid-Chromatography-Tandem Mass Spectrometry (LC-MS/MS)
2.5. Quality Assurance
2.6. Data Treatment
3. Results and Discussion
3.1. Phytosterols and Squalene
3.2. Tocopherols and Tocotrienols
3.3. Carotenoids
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- International Energy Agency. Renewables 2020—Analysis and Forecast to 2025; IEA: Paris, France, 2020. [Google Scholar]
- ePURE. European Renewable Ethanol—Key Figures. 2019. Available online: https://www.epure.org/media/2044/200813-def-pr-epure-infographic-european-renewable-ethanol-key-figures-2019_web.pdf (accessed on 20 November 2020).
- European Commission. Renewable Energy Progress Report. Report from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions; European Commission: Brussels, Belgium, 2017. [Google Scholar]
- European Commission. A sustainable Bioeconomy for Europe. Strengthening the Connection between Economy, Society and the Environment. Updated Bioeconomy Strategy 2018. Directorate-General for Research and Innovation; European Commission: Brussels, Belgium, 2018. [Google Scholar]
- European Commission. Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions. A New Circular Economy Action Plan for a Cleaner and More Competitive Europe; European Commission: Brussels, Belgium, 2020. [Google Scholar]
- Oreopoulou, V.; Tzia, C. Utilization of plant by-products for the recovery of proteins, dietary fibers, antioxidants, and colorants. In Utilization of By-Products and Treatment of Waste in the Food Industry; Oreopoulou, V., Russ, W., Eds.; Springer: New York, NY, USA, 2007; Volume 3, pp. 209–232. [Google Scholar] [CrossRef]
- Teixeira, A.; Baenas, N.; Dominguez-Perles, R.; Barros, A.; Rosa, E.; Moreno, D.A.; Garcia-Viguera, C. Natural bioactive compounds from winery by-products as health promoters: A review. Int. J. Mol. Sci. 2014, 15, 15638–15678. [Google Scholar] [CrossRef] [PubMed]
- Mirabella, N.; Castellani, V.; Sala, S. Current options for the valorization of food manufacturing waste: A review. J. Clean. Prod. 2014, 65, 28–41. [Google Scholar] [CrossRef]
- Ben-Othman, S.; Jõudu, I.; Bhat, R. Bioactives from agri-food wastes: Present insights and future challenges. Molecules 2020, 25, 510. [Google Scholar] [CrossRef]
- Sheng, S.; Li, T.; Liu, R. Corn phytochemicals and their health benefits. Food Sci. Hum. Wellness 2018, 7, 185–195. [Google Scholar] [CrossRef]
- Di Lena, G.; Ondrejíčková, P.; Pulgar, J.S.D.; Cyprichová, V.; Ježovič, T.; Lucarini, M.; Lombardi Boccia, G.; Ferrari Nicoli, S.; Gabrielli, P.; Aguzzi, A.; et al. Towards a Valorization of Corn Bioethanol Side Streams: Chemical Characterization of Post Fermentation Corn Oil and Thin Stillage. Molecules 2020, 25, 3549. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Di Lena, G.; Casini, I.; Lucarini, M.; Lombardi-Boccia, G. Carotenoid profiling of five microalgae species from large-scale production. Food Res. Int. 2019, 120, 810–818. [Google Scholar] [CrossRef]
- Winkler, J.K.; Rennick, K.A.; Eller, F.J.; Vaughn, S.F. Phytosterol and tocopherol components in extracts of corn distiller’s dried grain. J. Agric. Food Chem. 2007, 55, 6482–6486. [Google Scholar] [CrossRef]
- Majoni, S.; Wang, T. Characterization of oil precipitate and oil extracted from condensed corn distillers solubles. JAOCS J. Am. Oil Chem. Soc. 2010, 87, 205–213. [Google Scholar] [CrossRef]
- Seitz, L.M. Stanol and sterol esters of ferulic and p-coumaric acids in wheat, corn, rye, and triticale. J. Agric. Food Chem. 1989, 37, 662–667. [Google Scholar] [CrossRef]
- Harrabi, S.; St-Amand, A.; Sakouhi, F.; Sebei, K.; Kallel, H.; Mayer, P.M.; Boukhchina, S. Phytostanols and phytosterols distributions in corn kernel. Food Chem. 2008, 111, 115–120. [Google Scholar] [CrossRef]
- Esche, R.; Scholz, B.; Engel, K.H. Analysis of free phytosterols/stanols and their intact fatty acid and phenolic acid esters in various corn cultivars. J. Cereal Sci. 2013, 58, 333–340. [Google Scholar] [CrossRef]
- Ostlund, R.E., Jr.; Racette, S.B.; Okeke, A.; Stenson, W.F. Phytosterols that are naturally present in commercial corn oil significantly reduce cholesterol absorption in humans. Am. J. Clin. Nutr. 2002, 75, 1000–1004. [Google Scholar] [CrossRef]
- Phillips, K.M.; Ruggio, D.M.; Toivo, J.I.; Swank, M.A.; Simpkins, A.H. Free and esterified sterol composition of edible oils and fats. J. Food Compos. Anal. 2002, 15, 123–142. [Google Scholar] [CrossRef]
- Schwartz, H.; Ollilainen, V.; Piironen, V.; Lampi, A.-M. Tocopherol, tocotrienol and plant sterol contents of vegetable oils and industrial fats. J. Food Compos. Anal. 2008, 21, 152–161. [Google Scholar] [CrossRef]
- Normén, L.; Ellegård, L.; Brants, H.; Dutta, P.; Andersson, H. A phytosterol database: Fatty foods consumed in Sweden and the Netherlands. J. Food Compos. Anal. 2007, 20, 193–201. [Google Scholar] [CrossRef]
- De Jong, A.; Plat, J.; Bast, A.; Godschalk, R.W.L.; Basu, S.; Mensink, R.P. Effects of plant sterol and stanol ester consumption on lipid metabolism antioxidant status, and markers of oxidative stress, endothelial function, and low-grade inflammation in patients on current statin treatment. Eur. J. Clin. Nutr. 2008, 62, 263–273. [Google Scholar] [CrossRef]
- Woyengo, T.A.; Ramprasath, V.R.; Jones, P.J.H. Anticancer effects of phytosterols. Eur. J. Clin. Nutr. 2009, 63, 813–820. [Google Scholar] [CrossRef]
- Talati, R.; Sobieraj, D.M.; Makanji, S.S.; Phung, O.J.; Coleman, C.I. The comparative efficacy of plant sterols and stanols on serum lipids: A systematic review and meta-analysis. J. Am. Diet. Assoc. 2010, 110, 719–726. [Google Scholar] [CrossRef]
- Ghatak, S.B.; Panchal, S.J. Gamma-oryzanol—A multi-purpose steryl ferulate. Curr. Nutr. Food Sci. 2011, 7, 10–20. [Google Scholar] [CrossRef]
- Zhu, D.; Sánchez-Ferrer, A.; Nyström, L. Antioxidant Activity of Individual Steryl Ferulates from Various Cereal Grain Sources. J. Nat. Prod. 2016, 79, 308–316. [Google Scholar] [CrossRef]
- Jones, P.J.H.; Shamloo, M.; MacKay, D.S.; Rideout, T.C.; Myrie, S.B.; Plat, J.; Roullet, J.-B.; Baer, D.J.; Calkins, K.L.; Davis, H.R.; et al. Progress and perspectives in plant sterol and plant stanol research. Nutr. Rev. 2018, 76, 725–746. [Google Scholar] [CrossRef]
- Moreau, R.A.; Nyström, L.; Whitaker, B.D.; Winkler-Moser, J.K.; Baer, D.J.; Gebauer, S.K.; Hicks, K.B. Phytosterols and their derivatives: Structural diversity, distribution, metabolism, analysis, and health-promoting uses. Prog. Lipid Res. 2018, 70, 35–61. [Google Scholar] [CrossRef]
- Garcia-Llatas, G.; Cilla, A.; Alegría, A.; Lagarda, M.J. Bioavailability of plant sterol-enriched milk-based fruit beverages: In vivo and in vitro studies. J. Funct. Foods 2015, 14, 44–50. [Google Scholar] [CrossRef]
- Gies, M.; Servent, A.; Borel, P.; Dhuique-Mayer, C. Phytosterol vehicles used in a functional product modify carotenoid/cholesterol bioaccessibility and uptake by Caco-2 cells. J. Funct. Foods 2020, 68, 103920. [Google Scholar] [CrossRef]
- Reddy, L.; Couvreur, P. Squalene: A natural triterpene for use in disease management and therapy. Adv. Drug Deliv. Rev. 2009, 61, 1412–1426. [Google Scholar] [CrossRef]
- Popa, I.; Băbeanu, N.; Niță, S.; Popa, O. Squalene-Natural resources and applications. Farmacia 2014, 62, 840–862. [Google Scholar]
- Hossain, A.; Jayadeep, A. Determination of tocopherol and tocotrienol contents in maize by in vitro digestion and chemical methods. J. Cereal Sci. 2018, 83, 90–95. [Google Scholar] [CrossRef]
- Grilo, E.C.; Costa, P.N.; Gurgel, C.S.S.; Beserra, A.F.L.; Almeida, F.N.S.; Dimenstein, R. Alpha-tocopherol and gamma-tocopherol concentration in vegetable oils. Food Sci. Technol. 2014, 34, 379–385. [Google Scholar] [CrossRef]
- Gibreel, A.; Sandercock, J.R.; Lan, J.; Goonewardene, L.A.; Scott, A.C.; Zijlstra, R.T.; Curtis, J.M.; Bressler, D.C. Evaluation of value-added components of dried distiller’s grain with and phytosterols distributions in corn kernel. Food Chem. 2011, 111, 115–120. [Google Scholar] [CrossRef]
- Winkler-Moser, J.K.; Breyer, L. Composition and oxidative stability of crude oil extracts of corn germ and distillers grains. Ind. Crops Prod. 2011, 33, 572–578. [Google Scholar] [CrossRef]
- Saldeen, K.; Saldeen, T. Importance of tocopherols beyond α-tocopherol: Evidence from animal and human studies. Nutr. Res. 2005, 25, 877–889. [Google Scholar] [CrossRef]
- Moreau, R.A.; Hicks, K.B. The composition of corn oil obtained by the alcohol extraction of ground corn. JAOCS J. Am. Oil Chem. Soc. 2005, 82, 809–815. [Google Scholar] [CrossRef]
- Shin, E.-C.; Shurson, G.C.; Gallaher, D.D. Antioxidant capacity and phytochemical content of 16 sources of corn distillers dried grains with solubles (DDGS). Anim. Nutr. 2018, 4, 435–441. [Google Scholar] [CrossRef]
- Zingg, J.M. Modulation of signal transduction by vitamin E. Mol. Asp. Med. 2007, 28, 481–506. [Google Scholar] [CrossRef]
- Ahsan, H.; Ahad, A.; Siddiqui, W.A. A review of characterization of tocotrienols from plant oils and foods. J. Chem. Biol. 2015, 8, 45–59. [Google Scholar] [CrossRef]
- Sen, C.K.; Khanna, S.; Roy, S. Tocotrienols in health and disease: The other half of the natural vitamin E family. Mol. Asp. Med. 2007, 28, 692–728. [Google Scholar] [CrossRef]
- Khanna, S.; Parinandi, N.L.; Kotha, S.R.; Roy, S.; Rink, C.; Bibus, D.; Sen, C.K. Nanomolar vitamin E α-tocotrienol inhibits glutamate induced activation of phospholipase A2 and causes neuroprotection. J. Neurochem. 2010, 112, 1249–1260. [Google Scholar] [CrossRef]
- Peh, H.Y.; Tan, W.S.D.; Liao, W.; Wong, W.S.F. Vitamin E therapy beyond cancer: Tocopherol versus tocotrienol. Pharmacol. Ther. 2016, 162, 152–169. [Google Scholar] [CrossRef]
- Ramanathan, N.; Tan, E.; Loh, L.J.; Soh, B.S.; Yap, W.N. Tocotrienol is a cardioprotective agent against ageing-associated cardiovascular disease and its associated morbidities. Nutr. Metab. 2018, 15, 6. [Google Scholar] [CrossRef]
- Prasanth Kumar, P.K.; Jeyarani, T.; Gopala Krishna, A.G. Physicochemical characteristics of phytonutrient retained red palm olein and butter-fat blends and its utilization for formulating chocolate spread. J. Food Sci. Technol. 2016, 53, 3060–3072. [Google Scholar] [CrossRef]
- Bakota, E.L.; Winkler-Moser, J.K.; Hwang, H.-S.; Bowman, M.J.; Palmquist, D.E.; Liu, S.X. Solvent fractionation of rice bran oil to produce a spreadable rice bran product. Eur. J. Lipid Sci. Technol. 2013, 115, 847–857. [Google Scholar] [CrossRef]
- Obulesu, M.; Dowlathabad, M.R.; Bramhachari, P.V. Carotenoids and Alzheimer’s Disease: An insight into therapeutic role of retinoids in animal models. Neurochem. Int. 2011, 59, 535–541. [Google Scholar] [CrossRef]
- Wang, Y.; Cui, R.; Xiao, Y.; Fang, J.; Xu, Q. Effect of carotene and lycopene on the risk of prostate cancer: A systematic review and dose-response meta-analysis of observational studies. PLoS ONE 2015, 10, e0137427. [Google Scholar] [CrossRef]
- Murillo, A.G.; Fernandez, M.L. Potential of dietary non-provitamin a carotenoids in the prevention and treatment of diabetic microvascular complications. Adv. Nutr. 2016, 7, 14–24. [Google Scholar] [CrossRef]
- Cheng, H.M.; Koutsidis, G.; Lodge, J.K.; Ashor, A.; Siervo, M.; Lara, J. Tomato and lycopene supplementation and cardiovascular risk factors: A systematic review and meta-analysis. Atherosclerosis 2017, 257, 100–108. [Google Scholar] [CrossRef]
- Carpentier, S.; Knaus, M.; Suh, M. Associations between lutein, zeaxanthin, and age-related macular degeneration: An overview. Crit. Rev. Food Sci. Nutr. 2009, 49, 313–326. [Google Scholar] [CrossRef]
- Xavier, A.A.O.; Carvajal-Lérida, I.; Garrido-Fernández, J.; Pérez-Gálvez, A. In vitro bioaccessibility of lutein from cupcakes fortified with a water-soluble lutein esters formulation. J. Food Compos. Anal. 2018, 68, 60–64. [Google Scholar] [CrossRef]
- Ursache, F.M.; Andronoiu, D.G.; Ghinea, I.O.; Barbu, V.; Ioniţă, E.; Cotârleţ, M.; Dumitraşcu, L.; Botez, E.; Râpeanu, G.; Stănciuc, N. Valorizations of carotenoids from sea buckthorn extract by microencapsulation and formulation of value-added food products. J. Food Eng. 2018, 219, 16–24. [Google Scholar] [CrossRef]

{kind=link}
| Post-Fermentation Corn Oil | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lot | ERG | AVN a | STG + CAMP | β-SITO | STN | Σ STEROLS | SQUA | |||||||
| mg kg−1 Corn Oil | ||||||||||||||
| Free | Total | Free | Total | Free | Total | Free | Total | Free | Total | Free | Total | Free | Total | |
| 1 | 456 | 461 | 2081 | 2796 | 1178 | 2193 | 3073 | 7253 | 598 | - | 7387 | - | 867 | 832 |
| 2 | 413 | 433 | 1578 | 2636 | 1171 | 2616 | 3277 | 7652 | 529 | 4574 | 6968 | 17,911 | 875 | 843 |
| 3 | 408 | 411 | 1662 | 2248 | 1052 | 2186 | 3100 | 6742 | 540 | - | 6762 | - | 874 | 837 |
| 4 | 412 | 408 | 1618 | 2708 | 1113 | 2418 | 3289 | 7418 | 540 | 4277 | 6972 | 17,229 | 899 | 817 |
| 5 | 253 | 259 | 986 | 2131 | 1156 | 2358 | 4172 | 7238 | 569 | - | 7135 | - | 947 | 940 |
| 6 | 288 | 282 | 1412 | 2358 | 1149 | 2444 | 3528 | 7137 | 695 | 4638 | 7071 | 16,860 | 882 | 831 |
| 7 | 381 | 402 | 1428 | 2362 | 1034 | 2330 | 4180 | 7460 | 677 | 4829 | 7700 | 17,382 | 744 | 745 |
| 8 | 275 | 297 | 982 | 2066 | 983 | 2278 | 2834 | 6973 | 625 | 4379 | 5700 | 15,993 | 817 | 800 |
| 9 | 320 | 336 | 1353 | 2202 | 1081 | 2430 | 2803 | 7027 | 739 | 4472 | 6296 | 16,467 | 788 | 764 |
| 10 | 340 | 371 | 1107 | 2382 | 1022 | 2436 | 5250 | 7509 | 663 | 3134 | 8383 | 15,832 | 767 | 764 |
| 11 | 314 | 322 | 1151 | 2343 | 1017 | 2041 | 3449 | 6931 | - | - | - | - | 803 | 776 |
| Mean | 351 | 362 | 1396 | 2385 | 1087 | 2339 | 3541 | 7213 | 617 | 4329 | 7037 | 16,811 | 842 | 814 |
| sd | 66.7 | 66.6 | 333 | 236 | 69.6 | 158 | 730 | 280 | 73.8 | 556 | 731 | 759 | 62.4 | 54.2 |
| min | 253 | 259 | 982 | 2066 | 983 | 2041 | 2803 | 6742 | 529 | 3134 | 5700 | 15,832 | 744 | 745 |
| max | 456 | 461 | 2081 | 2796 | 1178 | 2616 | 5250 | 7652 | 739 | 4829 | 8383 | 17,912 | 947 | 940 |
| Thin Stillage | |||||||
|---|---|---|---|---|---|---|---|
| Lot | ERG | AVN a | STG + CAMP | β-SITO | STN | Σ STEROLS | SQUA |
| mg kg−1 Thin Stillage (Wet Mass Basis) | |||||||
| 1 | 11.3 | 61.3 | 54.0 | 170 | - | - | 19.2 |
| 2 | 5.16 | 31.4 | 29.5 | 88.3 | 50.6 | 205 | 10.1 |
| 3 | 8.33 | 38.0 | 40.4 | 135 | - | - | 16.2 |
| 4 | 7.62 | 45.6 | 41.3 | 141 | 86.6 | 322 | 15.3 |
| 5 | 5.20 | 35.8 | 34.8 | 103 | - | - | 12.9 |
| 6 | 5.12 | 32.2 | 30.1 | 110 | 63.1 | 241 | 12.0 |
| 7 | 4.87 | 18.0 | 35.0 | 101 | 47.4 | 207 | 11.4 |
| Mean | 6.80 | 37.5 | 37.9 | 121 | 61.9 | 244 | 13.9 |
| sd | 2.41 | 13.4 | 8.43 | 28.6 | 17.8 | 54.8 | 3.17 |
| min | 4.87 | 18.0 | 29.5 | 88.3 | 47.4 | 205 | 10.1 |
| max | 11.3 | 61.3 | 54.0 | 170 | 86.6 | 322 | 19.2 |
| mg kg−1 Thin Stillage (Dry Mass Basis) | |||||||
| 1 | 141 | 767 | 676 | 2134 | - | - | 240 |
| 2 | 63.7 | 388 | 364 | 1090 | 625 | 2532 | 125 |
| 3 | 106 | 483 | 514 | 1718 | - | - | 206 |
| 4 | 88.2 | 528 | 478 | 1632 | 1002 | 3728 | 178 |
| 5 | 60.9 | 420 | 409 | 1210 | - | - | 151 |
| 6 | 62.5 | 394 | 368 | 1349 | 771 | 2944 | 146 |
| 7 | 54.6 | 202 | 393 | 1138 | 532 | 2320 | 128 |
| Mean | 82.4 | 455 | 457 | 1467 | 732 | 2881 | 168 |
| sd | 31.7 | 172 | 112 | 379 | 205 | 621 | 42.7 |
| min | 54.6 | 202 | 364 | 1090 | 532 | 2320 | 125 |
| max | 141 | 767 | 676 | 2134 | 1002 | 3728 | 240 |
| Post-Fermentation Corn Oil | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lot | α-T | γ-T a | δ-T | Σ T | α-T3 | γ-T3 a | δ-T3 | Σ T3 | Σ (T + T3) | |||||||||
| mg kg−1 Corn Oil | ||||||||||||||||||
| Free | Total | Free | Total | Free | Total | Free | Total | Free | Total | Free | Total | Free | Total | Free | Total | Free | Total | |
| 1 | 179 | 175 | 700 | 706 | 25.2 | 18.2 | 904 | 899 | 216 | 133 | --- | 175 | 0.00 | 8.13 | - | 316 | - | 1215 |
| 2 | 195 | 204 | 753 | 773 | 24.8 | 25.6 | 973 | 1004 | 220 | 154 | --- | 196 | 0.00 | 8.45 | - | 358 | - | 1362 |
| 3 | 224 | 222 | 748 | 733 | 22.4 | 17.2 | 994 | 972 | 259 | 185 | --- | 214 | 0.00 | 6.08 | - | 405 | - | 1377 |
| 4 | 210 | 216 | 789 | 775 | 24.0 | 24.4 | 1023 | 1016 | 226 | 189 | --- | 241 | 0.00 | 7.72 | - | 438 | - | 1454 |
| 5 | 205 | 206 | 733 | 711 | 23.5 | 20.9 | 962 | 938 | 220 | 183 | --- | 214 | 0.00 | 8.31 | - | 406 | - | 1344 |
| 6 | 224 | 212 | 737 | 701 | 21.6 | 26.1 | 982 | 939 | 220 | 178 | --- | 202 | 0.00 | 4.10 | - | 384 | - | 1323 |
| 7 | 224 | 238 | 725 | 766 | 23.3 | 21.6 | 972 | 1026 | 208 | 174 | --- | 203 | 0.00 | 9.56 | - | 386 | - | 1412 |
| 8 | 201 | 224 | 727 | 731 | 28.3 | 30.1 | 956 | 985 | 196 | 175 | --- | 210 | 0.00 | 8.52 | - | 394 | - | 1379 |
| 9 | 188 | 235 | 694 | 716 | 22.3 | 17.1 | 904 | 969 | 181 | 168 | --- | 189 | 0.00 | 8.80 | - | 365 | - | 1334 |
| 10 | 219 | 224 | 761 | 761 | 21.1 | 22.0 | 1002 | 1006 | 227 | 156 | --- | 193 | 0.00 | 5.23 | - | 354 | - | 1360 |
| 11 | 194 | 220 | 761 | 793 | 20.6 | 22.9 | 975 | 1036 | 169 | 162 | --- | 186 | 0.00 | 9.63 | - | 357 | - | 1393 |
| Mean | 206 | 216 | 739 | 742 | 23.4 | 22.4 | 968 | 981 | 213 | 169 | --- | 202 | 0.00 | 7.68 | - | 379 | - | 1359 |
| sd | 15.7 | 17.0 | 27.8 | 32.3 | 2.18 | 4.04 | 36.6 | 42.3 | 24.3 | 16.6 | --- | 17.9 | 0.00 | 1.78 | - | 33.0 | - | 60.5 |
| min | 179 | 175 | 694 | 701 | 20.6 | 17.1 | 904 | 899 | 169 | 133 | --- | 175 | 0.00 | 4.10 | - | 316 | - | 1215 |
| max | 224 | 238 | 789 | 793 | 28.3 | 30.1 | 1023 | 1036 | 259 | 189 | --- | 241 | 0.00 | 9.63 | - | 438 | - | 1454 |
| Thin Stillage | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Lot | α-T | γ-T a | δ-T | Σ T | α-T3 | γ-T3 a | δ-T3 | Σ T3 | Σ T + T3 |
| mg kg−1 Thin Stillage Wet Mass | |||||||||
| 1 | 3.95 | 16.5 | 0.57 | 21.0 | 2.99 | 3.96 | 0.05 | 6.99 | 28.0 |
| 2 | 1.89 | 8.21 | 0.22 | 10.3 | 1.32 | 2.00 | 0.00 | 3.32 | 13.6 |
| 3 | 3.63 | 14.0 | 0.29 | 17.9 | 3.14 | 4.12 | 0.00 | 7.26 | 25.2 |
| 4 | 3.77 | 13.6 | 0.39 | 17.8 | 3.11 | 3.90 | 0.00 | 7.02 | 24.8 |
| 5 | 2.70 | 9.80 | 0.40 | 12.9 | 2.30 | 2.90 | 0.00 | 5.20 | 18.1 |
| 6 | 2.87 | 10.3 | 0.26 | 13.4 | 2.31 | 2.83 | 0.00 | 5.14 | 18.5 |
| 7 | 2.85 | 10.5 | 0.46 | 13.8 | 2.00 | 2.55 | 0.00 | 4.55 | 18.4 |
| Mean | 3.09 | 11.8 | 0.37 | 15.3 | 2.45 | 3.18 | 0.01 | 5.64 | 20.9 |
| sd | 0.73 | 2.92 | 0.12 | 3.70 | 0.68 | 0.82 | 0.02 | 1.49 | 5.11 |
| min | 1.89 | 8.21 | 0.22 | 10.3 | 1.32 | 2.00 | 0.00 | 3.32 | 13.6 |
| max | 3.95 | 16.5 | 0.57 | 21.0 | 3.14 | 4.12 | 0.05 | 7.26 | 28.0 |
| mg kg−1 Thin Stillage Dry Mass | |||||||||
| 1 | 49.4 | 206 | 7.15 | 263 | 37.4 | 49.6 | 0.61 | 87.5 | 350 |
| 2 | 23.3 | 101 | 2.73 | 127 | 16.3 | 24.7 | 0.00 | 41.0 | 168 |
| 3 | 46.2 | 178 | 3.72 | 228 | 40.0 | 52.4 | 0.00 | 92.4 | 320 |
| 4 | 43.6 | 158 | 4.51 | 206 | 36.0 | 45.2 | 0.00 | 81.2 | 287 |
| 5 | 31.5 | 116 | 4.80 | 152 | 26.7 | 34.2 | 0.00 | 60.9 | 213 |
| 6 | 35.1 | 125 | 3.12 | 163 | 28.2 | 34.6 | 0.00 | 62.8 | 226 |
| 7 | 31.9 | 118 | 5.16 | 155 | 22.4 | 28.6 | 0.00 | 51.0 | 206 |
| Mean | 37.3 | 143 | 4.46 | 185 | 29.6 | 38.5 | 0.09 | 68.1 | 253 |
| sd | 9.39 | 38.5 | 1.48 | 48.6 | 8.65 | 10.7 | 0.23 | 19.4 | 67.0 |
| min | 23.3 | 101 | 2.73 | 127 | 16.3 | 24.7 | 0.00 | 41.0 | 168 |
| max | 49.4 | 206 | 7.15 | 263 | 40.0 | 52.4 | 0.61 | 92.4 | 351 |
| Post-Fermentation Corn Oil | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lot | Lutein | Zeaxanthin | β-cryptoxanthin | N.I.C. 1 | N.I.C. 2 | N.I.C. 3 | N.I.C. 4 | N.I.C. 5 | N.I.C. 6 | Total | ||||||||||
| mg kg−1 Corn Oil | ||||||||||||||||||||
| Free | Total | Free | Total | Free | Total | Free | Total | Free | Total | Free | Total | Free | Total | Free | Total | Free | Total | Σ Free | Σ Total | |
| 1 | 68.1 | 84.4 | 53.7 | 69.8 | 14.3 | 18.0 | 9.60 | 12.1 | 7.17 | 8.31 | 10.5 | 13.2 | 6.66 | 8.96 | 6.40 | 8.26 | 19.6 | 24.2 | 196 | 240 |
| 2 | 64.4 | 87.0 | 47.2 | 75.4 | 11.8 | 19.6 | 9.14 | 13.9 | 5.73 | 8.18 | 10.5 | 15.2 | 6.01 | 9.05 | 5.87 | 8.28 | 14.1 | 24.4 | 175 | 254 |
| 3 | 88.7 | 93.8 | 66.9 | 72.4 | 16.9 | 20.4 | 12.6 | 13.1 | 8.79 | 9.46 | 13.7 | 13.7 | 7.54 | 8.85 | 7.08 | 7.66 | 21.2 | 25.4 | 243 | 257 |
| 4 | 80.5 | 79.4 | 64.3 | 61.9 | 18.3 | 18.9 | 10.8 | 13.2 | 7.58 | 7.79 | 12.5 | 12.3 | 7.18 | 7.79 | 6.64 | 7.01 | 21.1 | 21.6 | 229 | 223 |
| 6 | 84.0 | 84.4 | 61.7 | 62.8 | 17.9 | 19.8 | 11.6 | 11.7 | 7.82 | 8.29 | 11.9 | 12.0 | 7.31 | 8.55 | 6.90 | 7.44 | 19.1 | 21.7 | 228 | 229 |
| 7 | 88.3 | - | 58.7 | - | 16.2 | - | 10.1 | - | 6.71 | - | 11.4 | - | 8.36 | - | 8.20 | - | 21.0 | - | 229 | - |
| Mean | 79.0 | 85.8 | 58.7 | 68.5 | 15.9 | 19.3 | 10.6 | 12.8 | 7.3 | 8.41 | 11.8 | 13.3 | 7.17 | 8.64 | 6.85 | 7.73 | 19.3 | 23.4 | 217 | 240 |
| sd | 10.4 | 5.28 | 7.27 | 5.92 | 2.46 | 0.95 | 1.28 | 0.89 | 1.04 | 0.63 | 1.23 | 1.29 | 0.80 | 0.51 | 0.79 | 0.55 | 2.73 | 1.71 | 25.6 | 14.9 |
| min | 64.4 | 79.4 | 47.2 | 61.9 | 11.8 | 18.0 | 9.14 | 11.7 | 5.73 | 7.79 | 10.5 | 12.0 | 6.01 | 7.79 | 5.87 | 7.01 | 14.0 | 21.6 | 175 | 223 |
| max | 88.7 | 93.8 | 66.9 | 75.4 | 18.3 | 20.4 | 12.6 | 13.9 | 8.79 | 9.46 | 13.7 | 15.2 | 8.36 | 9.05 | 8.20 | 8.28 | 21.2 | 25.4 | 243 | 257 |
| Thin Stillage | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Lot | Lutein | Zeaxanthin | β-cryptoxanthin | N.I.C. 1 | N.I.C. 2 | N.I.C. 3 | N.I.C. 4 | N.I.C. 5 | N.I.C. 6 | Total | ||||||||||
| Free | Total | Free | Total | Free | Total | Free | Total | Free | Total | Free | Total | Free | Total | Free | Total | Free | Total | Σ Free | Σ Total | |
| mg kg−1 Thin Stillage Wet Mass | ||||||||||||||||||||
| 1 | 2.62 | 2.98 | 2.61 | 3.23 | 0.54 | 0.75 | 0.30 | 0.40 | 0.20 | 0.23 | 0.36 | 0.42 | 0.24 | 0.27 | 0.23 | 0.27 | 0.73 | 0.88 | 7.84 | 9.44 |
| 2 | 1.68 | 2.11 | 1.39 | 2.24 | 0.27 | 0.62 | 0.19 | 0.23 | 0.07 | 0.17 | 0.20 | 0.40 | 0.15 | 0.19 | 0.14 | 0.18 | 0.41 | 0.61 | 4.50 | 6.76 |
| 3 | 2.15 | 2.78 | 1.91 | 3.16 | 0.39 | 0.83 | 0.3 | 0.37 | 0.1 | 0.24 | 0.19 | 0.44 | 0.19 | 0.23 | 0.17 | 0.23 | 0.58 | 0.87 | 5.96 | 9.14 |
| 4 | 2.28 | 2.76 | 2.04 | 3.04 | 0.41 | 0.81 | 0.29 | 0.33 | 0.1 | 0.23 | 0.18 | 0.41 | 0.19 | 0.23 | 0.17 | 0.24 | 0.58 | 0.85 | 6.24 | 8.91 |
| 5 | 2.19 | 2.45 | 2.09 | 2.42 | 0.42 | 0.63 | 0.24 | 0.29 | 0.19 | 0.21 | 0.32 | 0.38 | 0.17 | 0.20 | 0.16 | 0.20 | 0.50 | 0.66 | 6.28 | 7.45 |
| 6 | 2.35 | 2.6 | 2.15 | 2.48 | 0.47 | 0.66 | 0.29 | 0.35 | 0.21 | 0.24 | 0.33 | 0.38 | 0.21 | 0.25 | 0.2 | 0.25 | 0.60 | 0.75 | 6.81 | 7.95 |
| 7 | 2.07 | 2.31 | 2.03 | 2.25 | 0.46 | 0.59 | 0.19 | 0.25 | 0.16 | 0.17 | 0.27 | 0.33 | 0.18 | 0.21 | 0.18 | 0.22 | 0.54 | 0.66 | 6.09 | 6.98 |
| Mean | 2.19 | 2.57 | 2.03 | 2.69 | 0.42 | 0.70 | 0.26 | 0.32 | 0.15 | 0.21 | 0.26 | 0.39 | 0.19 | 0.23 | 0.18 | 0.23 | 0.56 | 0.75 | 6.25 | 8.09 |
| sd | 0.28 | 0.30 | 0.36 | 0.44 | 0.08 | 0.09 | 0.05 | 0.06 | 0.06 | 0.03 | 0.08 | 0.04 | 0.03 | 0.03 | 0.03 | 0.03 | 0.1 | 0.12 | 1.00 | 1.08 |
| min | 1.68 | 2.11 | 1.39 | 2.24 | 0.27 | 0.59 | 0.19 | 0.23 | 0.07 | 0.17 | 0.18 | 0.33 | 0.15 | 0.19 | 0.14 | 0.18 | 0.41 | 0.61 | 4.50 | 6.76 |
| max | 2.62 | 2.98 | 2.61 | 3.23 | 0.54 | 0.83 | 0.30 | 0.40 | 0.21 | 0.24 | 0.36 | 0.44 | 0.24 | 0.27 | 0.23 | 0.27 | 0.73 | 0.88 | 7.84 | 9.44 |
| mg kg−1 Thin Stillage Dry Mass | ||||||||||||||||||||
| 1 | 32.7 | 37.4 | 32.7 | 40.4 | 6.81 | 9.37 | 3.82 | 4.99 | 2.52 | 2.91 | 4.53 | 5.31 | 2.98 | 3.44 | 2.88 | 3.34 | 9.11 | 10.99 | 98 | 118 |
| 2 | 20.8 | 26.1 | 17.1 | 27.7 | 3.34 | 7.69 | 2.38 | 2.84 | 0.87 | 2.15 | 2.46 | 4.93 | 1.83 | 2.30 | 1.69 | 2.25 | 5.00 | 7.49 | 56 | 83 |
| 3 | 27.4 | 35.4 | 24.2 | 40.2 | 4.95 | 10.5 | 3.77 | 4.70 | 1.22 | 3.00 | 2.36 | 5.61 | 2.36 | 2.95 | 2.11 | 2.90 | 7.40 | 11.1 | 76 | 116 |
| 4 | 26.4 | 31.9 | 23.6 | 35.2 | 4.72 | 9.33 | 3.35 | 3.88 | 1.15 | 2.64 | 2.04 | 4.75 | 2.18 | 2.71 | 2.02 | 2.78 | 6.75 | 9.89 | 72 | 103 |
| 5 | 25.8 | 28.7 | 24.5 | 28.4 | 4.94 | 7.43 | 2.80 | 3.45 | 2.21 | 2.49 | 3.77 | 4.44 | 2.02 | 2.37 | 1.91 | 2.35 | 5.82 | 7.70 | 74 | 87 |
| 6 | 28.7 | 31.8 | 26.2 | 30.3 | 5.68 | 8.08 | 3.59 | 4.22 | 2.62 | 2.89 | 4.05 | 4.65 | 2.58 | 3.03 | 2.42 | 3.04 | 7.28 | 9.16 | 83 | 97 |
| 7 | 23.3 | 26.0 | 22.7 | 25.3 | 5.20 | 6.62 | 2.16 | 2.78 | 1.78 | 1.86 | 3.06 | 3.69 | 2.05 | 2.39 | 1.97 | 2.42 | 6.07 | 7.40 | 68 | 78 |
| Media | 26.4 | 31.0 | 24.5 | 32.5 | 5.09 | 8.43 | 3.12 | 3.83 | 1.77 | 2.56 | 3.18 | 4.77 | 2.29 | 2.74 | 2.14 | 2.73 | 6.77 | 9.10 | 75 | 98 |
| sd | 3.83 | 4.39 | 4.63 | 6.13 | 1.05 | 1.35 | 0.68 | 0.86 | 0.70 | 0.43 | 0.95 | 0.62 | 0.39 | 0.42 | 0.39 | 0.40 | 1.33 | 1.61 | 13 | 16 |
| min | 20.8 | 26.0 | 17.1 | 25.3 | 3.34 | 6.62 | 2.16 | 2.78 | 0.87 | 1.86 | 2.04 | 3.69 | 1.83 | 2.30 | 1.69 | 2.25 | 5.00 | 7.40 | 56 | 78 |
| max | 32.7 | 37.4 | 32.7 | 40.4 | 6.81 | 10.5 | 3.82 | 4.99 | 2.62 | 3.00 | 4.53 | 5.61 | 2.98 | 3.44 | 2.88 | 3.34 | 9.11 | 11.1 | 98.1 | 118 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Lena, G.; Sanchez del Pulgar, J.; Lombardi Boccia, G.; Casini, I.; Ferrari Nicoli, S. Corn Bioethanol Side Streams: A Potential Sustainable Source of Fat-Soluble Bioactive Molecules for High-Value Applications. Foods 2020, 9, 1788. https://doi.org/10.3390/foods9121788
Di Lena G, Sanchez del Pulgar J, Lombardi Boccia G, Casini I, Ferrari Nicoli S. Corn Bioethanol Side Streams: A Potential Sustainable Source of Fat-Soluble Bioactive Molecules for High-Value Applications. Foods. 2020; 9(12):1788. https://doi.org/10.3390/foods9121788
Chicago/Turabian StyleDi Lena, Gabriella, Jose Sanchez del Pulgar, Ginevra Lombardi Boccia, Irene Casini, and Stefano Ferrari Nicoli. 2020. "Corn Bioethanol Side Streams: A Potential Sustainable Source of Fat-Soluble Bioactive Molecules for High-Value Applications" Foods 9, no. 12: 1788. https://doi.org/10.3390/foods9121788
APA StyleDi Lena, G., Sanchez del Pulgar, J., Lombardi Boccia, G., Casini, I., & Ferrari Nicoli, S. (2020). Corn Bioethanol Side Streams: A Potential Sustainable Source of Fat-Soluble Bioactive Molecules for High-Value Applications. Foods, 9(12), 1788. https://doi.org/10.3390/foods9121788