

In Vitro Gastrointestinal Bioaccessibility, Bioactivities and Colonic Fermentation of Phenolic Compounds in Different Vigna Beans
 ,
, 
Abstract

1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Chemicals and Reagents
2.3. Extraction of Phenolic Compounds from Raw Materials
2.4. In Vitro Gastrointestinal Digestion
2.5. In Vitro Colonic Fermentation
2.6. Estimation of Phenolic Content and Antioxidant Capacity
2.6.1. Determination of Total Phenolic Content (TPC)
2.6.2. Determination of Total Flavonoids Content (TFC)
2.6.3. Determination of Total Condensed Tannins (TCT)
2.6.4. 2,2′-Diphenyl-1-Picrylhydrazyl (DPPH) Antioxidant Assay
2.6.5. Ferric Reducing Antioxidant Power (FRAP) Assay
2.7. Quantification of Phenolic Compounds through High-Performance Liquid Chromatography Photodiode Array (HPLC-PDA)
2.8. Gastrointestinal Digestion and Colonic Fermentation Indices
2.8.1. Bioaccessibility of Phenolic Compounds
2.8.2. Recovery Index
2.8.3. Residual Intestinal Digesta Index
2.8.4. Residual Colonic Digesta Index
2.9. Short Chain Fatty Acids (SCFAs) Analysis
2.10. Statistical Analysis
3. Results and Discussion
3.1. Phenolic and Bioactivity Changes during In Vitro Digestion
3.1.1. Phenolic Estimation
3.1.2. Antioxidant Activities Estimation
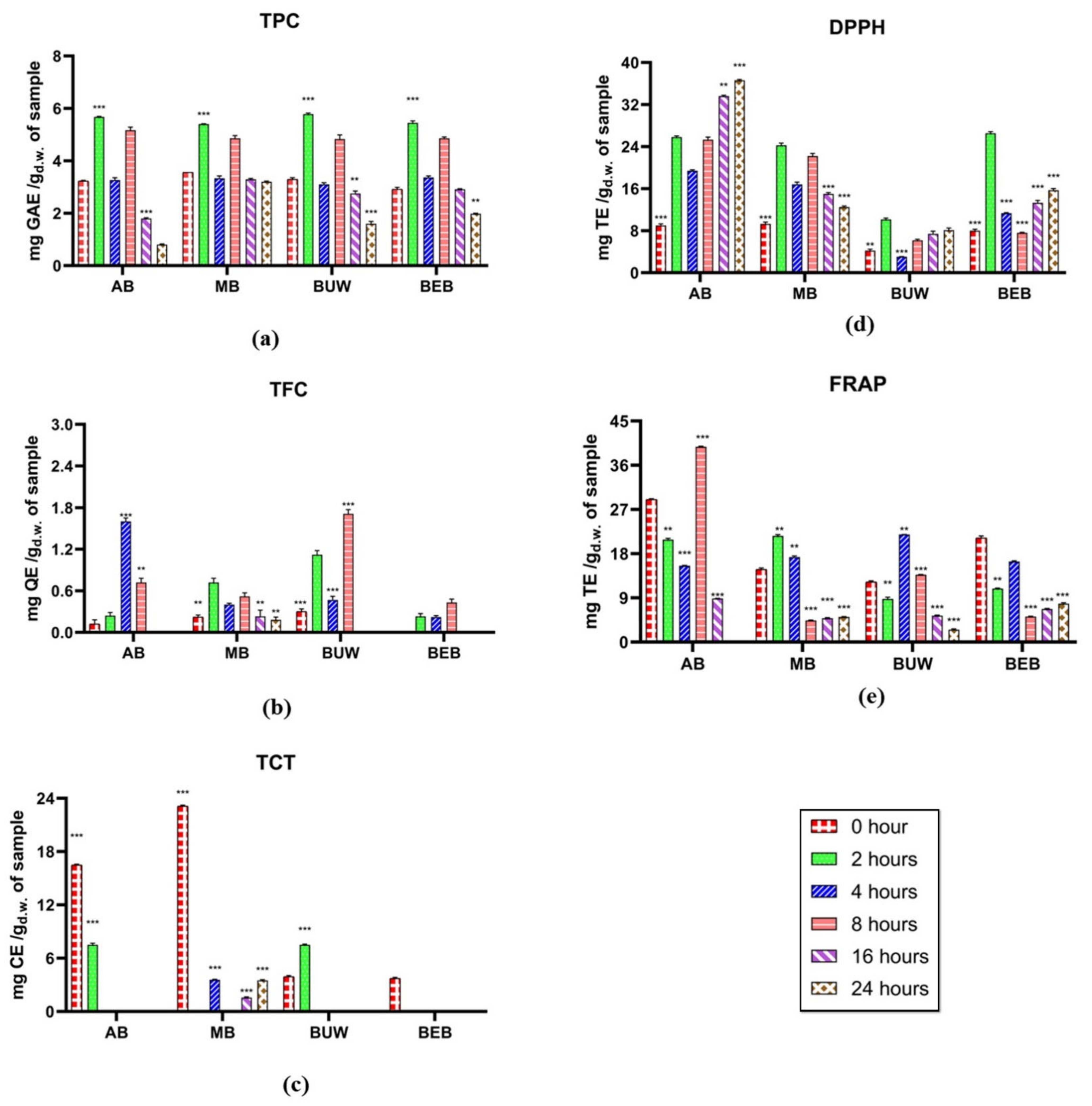
3.2. Phenolic and Bioactivity Changes during Colonic Fermentation
3.2.1. Phenolic Estimation
3.2.2. Antioxidant Capacity Estimation
3.3. Bioaccessibility of Individual Phenolic Compounds in Four Vigna Samples
3.4. Intestinal Recovery and Residual Index of Individual Phenolic Compounds in Four Vigna Samples
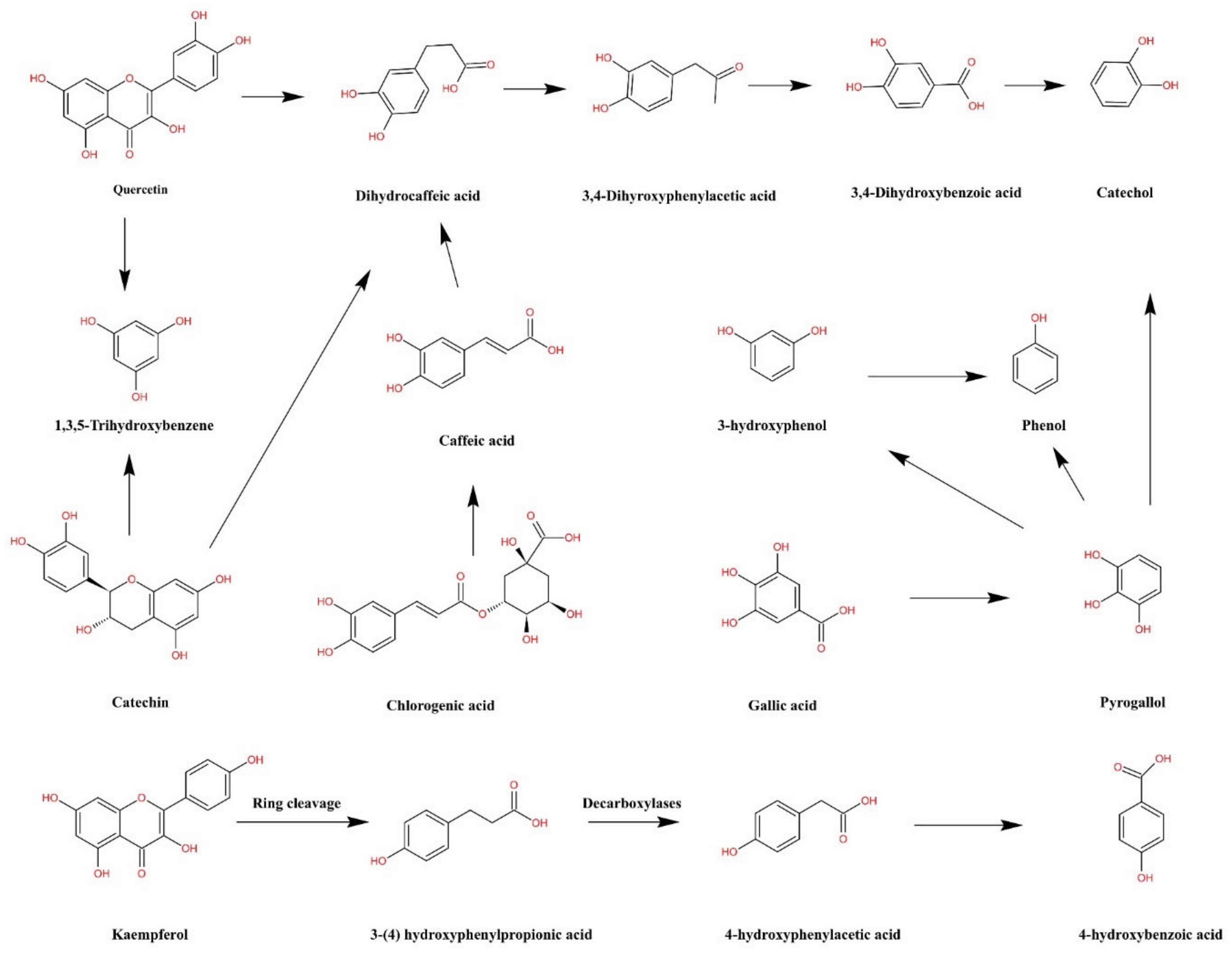
3.5. Short Chain Fatty Acids Production
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aitawade, M.; Sutar, S.; Rao, S.; Malik, S.; Yadav, S.; Bhat, K. Section Ceratotropis of subgenus Ceratotropis of Vigna (Leguminosae-Papilionoideae) in India with a new species from northern Western Ghats. Rheedea 2012, 22, 20–27. [Google Scholar]
- Singh, B. Cowpea: The food legume of the 21st century. Madison, WI: Crop Science Society of America. Inc. Doi. 2014, 10, 2014. [Google Scholar]
- Cheng, A.; Hou, C.; Sun, J.; Wan, F. Effect of steam explosion on phenolic compounds and antioxidant capacity in adzuki beans. J. Sci. Food Agric. 2020, 100, 4495–4503. [Google Scholar] [CrossRef] [PubMed]
- Girish, T.K.; Pratape, V.M.; Prasada Rao, U.J.S. Nutrient distribution, phenolic acid composition, antioxidant and alpha-glucosidase inhibitory potentials of black gram (Vigna mungo L.) and its milled by-products. Food Res. Int. 2012, 46, 370–377. [Google Scholar] [CrossRef]
- Senpuku, H.; Mohri, S.; Mihara, M.; Arai, T.; Suzuki, Y.; Saeki, Y. Effects of 7S globulin 3 derived from the adzuki bean [Vigna angularis] on the CSP- and eDNA- dependent biofilm formation of Streptococcus mutans. Arch. Oral Biol. 2019, 102, 256–265. [Google Scholar] [CrossRef] [PubMed]
- Winham, D.M.; Hutchins, A.M.; Johnston, C.S. Pinto Bean Consumption Reduces Biomarkers for Heart Disease Risk. J. Am. Coll. Nutr. 2007, 26, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Li, H.; Yi, J.; Zhang, J.; Che, H.; Cao, J.; Yang, L.; Zhu, C.; Jiang, W. Antioxidant properties of the mung bean flavonoids on alleviating heat stress. PLoS ONE 2011, 6, e21071. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Li, M.; Wu, H.; Liu, Z.; Barrow, C.; Dunshea, F.; Suleria, H.A.R. Bioaccessibility of phenolic compounds from sesame seeds (Sesamum indicum L.) during in vitro gastrointestinal digestion and colonic fermentation. J. Food Process. Preserv. 2022, 46, e16669. [Google Scholar] [CrossRef]
- Shi, M.; Wu, H.; Li, M.; Liu, Z.; Duan, X.; Barrow, C.; Dunshea, F.; Suleria, H.A.R. Assessment of the bioaccessibility of phenolics from Australian grown lettuces by in vitro simulated gastrointestinal digestion and colonic fermentation. Food Biosci. 2022, 48, 101754. [Google Scholar] [CrossRef]
- Quan, W.; Qie, X.; Chen, Y.; Zeng, M.; Qin, F.; Chen, J.; He, Z. Effect of milk addition and processing on the antioxidant capacity and phenolic bioaccessibility of coffee by using an in vitro gastrointestinal digestion model. Food Chem. 2020, 308, 125598. [Google Scholar] [CrossRef]
- Iglesias-Carres, L.; Mas-Capdevila, A.; Bravo, F.I.; Aragonès, G.; Arola-Arnal, A.; Muguerza, B. A comparative study on the bioavailability of phenolic compounds from organic and nonorganic red grapes. Food Chem. 2019, 299, 125092. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, A.; Goufo, P.; Barros, A.; Domínguez-Perles, R.; Trindade, H.; Rosa, E.A.S.; Ferreira, L.; Rodrigues, M. Cowpea (Vigna unguiculata L. Walp), a renewed multipurpose crop for a more sustainable agri-food system: Nutritional advantages and constraints. J. Sci. Food Agric. 2016, 96, 2941–2951. [Google Scholar] [CrossRef] [PubMed]
- Pavez-Guajardo, C.; Ferreira, S.R.S.; Mazzutti, S.; Guerra-Valle, M.E.; Sáez-Trautmann, G.; Moreno, J. Influence of In Vitro Digestion on Antioxidant Activity of Enriched Apple Snacks with Grape Juice. Foods 2020, 9, 1681. [Google Scholar] [CrossRef] [PubMed]
- Dong, R.; Liu, S.; Xie, J.; Chen, Y.; Zheng, Y.; Zhang, X.; Zhao, E.; Wang, Z.; Xu, H.; Yu, Q. The recovery, catabolism and potential bioactivity of polyphenols from carrot subjected to in vitro simulated digestion and colonic fermentation. Food Res. Int. 2021, 143, 110263. [Google Scholar] [CrossRef]
- Déprez, S.P.; Brezillon, C.; Rabot, S.; Philippe, C.; Mila, I.; Lapierre, C.; Scalbert, A. Polymeric Proanthocyanidins Are Catabolized by Human Colonic Microflora into Low-Molecular-Weight Phenolic Acids. J. Nutr. 2000, 130, 2733–2738. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Chang, S.K.C.; Zhang, Y.; Hsu, C.-Y.; Nannapaneni, R. Gut microbiota and short chain fatty acid composition as affected by legume type and processing methods as assessed by simulated in vitro digestion assays. Food Chem. 2020, 312, 126040. [Google Scholar] [CrossRef]
- Peng, D.; Zahid, H.F.; Ajlouni, S.; Dunshea, F.R.; Suleria, H.A.R. LC-ESI-QTOF/MS Profiling of Australian Mango Peel By-Product Polyphenols and Their Potential Antioxidant Activities. Processes 2019, 7, 764. [Google Scholar] [CrossRef]
- Gu, C.; Suleria, H.A.; Dunshea, F.R.; Howell, K. Dietary lipids influence bioaccessibility of polyphenols from black carrots and affect microbial diversity under simulated gastrointestinal digestion. Antioxidants 2020, 9, 762. [Google Scholar] [CrossRef] [PubMed]
- Mussatto, S.I.; Ballesteros, L.F.; Martins, S.; Teixeira, J.A. Extraction of antioxidant phenolic compounds from spent coffee grounds. Sep. Purif. Technol. 2011, 83, 173–179. [Google Scholar] [CrossRef]
- Suleria, H.A.; Barrow, C.J.; Dunshea, F.R. Screening and characterization of phenolic compounds and their antioxidant capacity in different fruit peels. Foods 2020, 9, 1206. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Liu, Z.; Lu, P.; Barrow, C.; Dunshea, F.R.; Suleria, H.A.R. Bioaccessibility and bioactivities of phenolic compounds from roasted coffee beans during in vitro digestion and colonic fermentation. Food Chem. 2022, 386, 132794. [Google Scholar] [CrossRef] [PubMed]
- Chait, Y.A.; Gunenc, A.; Bendali, F.; Hosseinian, F. Simulated gastrointestinal digestion and in vitro colonic fermentation of carob polyphenols: Bioaccessibility and bioactivity. LWT 2020, 117, 108623. [Google Scholar] [CrossRef]
- Mosele, J.I.; Macià, A.; Romero, M.-P.; Motilva, M.-J. Stability and metabolism of Arbutus unedo bioactive compounds (phenolics and antioxidants) under in vitro digestion and colonic fermentation. Food Chem. 2016, 201, 120–130. [Google Scholar] [CrossRef] [PubMed]
- Cárdenas-Castro, A.P.; Pérez-Jiménez, J.; Bello-Pérez, L.A.; Tovar, J.; Sáyago-Ayerdi, S.G. Bioaccessibility of phenolic compounds in common beans (Phaseolus vulgaris L.) after in vitro gastrointestinal digestion: A comparison of two cooking procedures. Cereal Chem. 2020, 97, 670–680. [Google Scholar] [CrossRef]
- Bohn, T. Dietary factors affecting polyphenol bioavailability. Nutr. Rev. 2014, 72, 429–452. [Google Scholar] [CrossRef] [PubMed]
- Lafarga, T.; Villaró, S.; Bobo, G.; Simó, J.; Aguiló-Aguayo, I. Bioaccessibility and antioxidant activity of phenolic compounds in cooked pulses. Int. J. Food Sci. Technol. 2019, 54, 1816–1823. [Google Scholar] [CrossRef]
- Pekkinen, J.; Rosa, N.N.; Savolainen, O.-I.; Keski-Rahkonen, P.; Mykkänen, H.; Poutanen, K.; Micard, V.; Hanhineva, K. Disintegration of wheat aleurone structure has an impact on the bioavailability of phenolic compounds and other phytochemicals as evidenced by altered urinary metabolite profile of diet-induced obese mice. Nutr. Metab. 2014, 11, 1. [Google Scholar] [CrossRef]
- Zhang, B.; Deng, Z.; Tang, Y.; Chen, P.X.; Liu, R.; Dan Ramdath, D.; Liu, Q.; Hernandez, M.; Tsao, R. Bioaccessibility, in vitro antioxidant and anti-inflammatory activities of phenolics in cooked green lentil (Lens culinaris). J. Funct. Foods 2017, 32, 248–255. [Google Scholar] [CrossRef]
- Wojtunik-Kulesza, K.; Oniszczuk, A.; Oniszczuk, T.; Combrzyński, M.; Nowakowska, D.; Matwijczuk, A. Influence of In Vitro Digestion on Composition, Bioaccessibility and Antioxidant Activity of Food Polyphenols—A Non-Systematic Review. Nutrients 2020, 12, 1401. [Google Scholar] [CrossRef] [PubMed]
- Tarko, T.; Duda-Chodak, A.; Zajac, N. Digestion and absorption of phenolic compounds assessed by in vitro simulation methods. A review. Rocz. Państwowego Zakładu Hig. 2013, 64, 79–84. [Google Scholar]
- Wróblewski, K.; Muhandiram, R.; Chakrabartty, A.; Bennick, A. The molecular interaction of human salivary histatins with polyphenolic compounds. Eur. J. Biochem. 2001, 268, 4384–4397. [Google Scholar] [CrossRef] [PubMed]
- Su, D.; Li, N.; Chen, M.; Yuan, Y.; He, S.; Wang, Y.; Wu, Q.; Li, L.; Yang, H.; Zeng, Q. Effects of in vitro digestion on the composition of flavonoids and antioxidant activities of the lotus leaf at different growth stages. Int. J. Food Sci. Technol. 2018, 53, 1631–1639. [Google Scholar] [CrossRef]
- Arranz, S.; Silván, J.M.; Saura-Calixto, F. Nonextractable polyphenols, usually ignored, are the major part of dietary polyphenols: A study on the Spanish diet. Mol. Nutr. Food Res. 2010, 54, 1646–1658. [Google Scholar] [CrossRef]
- Luo, J.; Cai, W.; Wu, T.; Xu, B. Phytochemical distribution in hull and cotyledon of adzuki bean (Vigna angularis L.) and mung bean (Vigna radiate L.), and their contribution to antioxidant, anti-inflammatory and anti-diabetic activities. Food Chem. 2016, 201, 350–360. [Google Scholar] [CrossRef] [PubMed]
- Price, M.L.; Hagerman, A.E.; Butler, L.G. Tannin content of cowpeas, chickpeas, pigeon peas, and mung beans. J. Agric. Food Chem. 1980, 28, 459–461. [Google Scholar] [CrossRef] [PubMed]
- Floegel, A.; Kim, D.-O.; Chung, S.-J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular antioxidant-rich US foods. J. Food Compos. Anal. 2011, 24, 1043–1048. [Google Scholar] [CrossRef]
- Halliwell, B.; Rafter, J.; Jenner, A. Health promotion by flavonoids, tocopherols, tocotrienols, and other phenols: Direct or indirect effects? Antioxidant or not? Am. J. Clin. Nutr. 2005, 81, 268S–276S. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound phenolics in foods, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef]
- Barros, N.V.d.A.; Abreu, B.B.d.; Rocha, M.d.M.; Araújo, M.A.d.M.; Moreira-Araújo, R.S.d.R. In vitro bioaccessibility of phenolic compounds and antioxidant activity in biofortified cowpea cultivars. Rev. Ciência Agronômica 2021, 52, 1. [Google Scholar]
- Alqurashi, R.M.; Alarifi, S.N.; Walton, G.E.; Costabile, A.F.; Rowland, I.R.; Commane, D.M. In vitro approaches to assess the effects of açai (Euterpe oleracea) digestion on polyphenol availability and the subsequent impact on the faecal microbiota. Food Chem. 2017, 234, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Gao, K.; Xu, A.; Krul, C.; Venema, K.; Liu, Y.; Niu, Y.; Lu, J.; Bensoussan, L.; Seeram, N.P.; Heber, D.; et al. Of the Major Phenolic Acids Formed during Human Microbial Fermentation of Tea, Citrus, and Soy Flavonoid Supplements, Only 3,4-Dihydroxyphenylacetic Acid Has Antiproliferative Activity. J. Nutr. 2006, 136, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Rechner, A.R.; Kuhnle, G.; Bremner, P.; Hubbard, G.P.; Moore, K.P.; Rice-Evans, C.A. The metabolic fate of dietary polyphenols in humans. Free. Radic. Biol. Med. 2002, 33, 220–235. [Google Scholar] [CrossRef]
- Saura-Calixto, F.; Serrano, J.; Goñi, I. Intake and bioaccessibility of total polyphenols in a whole diet. Food Chem. 2007, 101, 492–501. [Google Scholar] [CrossRef]
- Gonthier, M.-P.; Donovan, J.L.; Texier, O.; Felgines, C.; Remesy, C.; Scalbert, A. Metabolism of dietary procyanidins in rats. Free. Radic. Biol. Med. 2003, 35, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Goderska, K.; Gumienna, M.; Czarnecki, Z. Release of phenolic compounds from bean flour, bean-derived chips and black chokeberry juice and changes in their antioxidant activity during digestion in an in vitro gastrointestinal model. Pol. J. Food Nutr. Sci. 2008, 58, 4. [Google Scholar]
- Lucas-González, R.; Ángel Pérez-Álvarez, J.; Moscaritolo, S.; Fernández-López, J.; Sacchetti, G.; Viuda-Martos, M. Evaluation of polyphenol bioaccessibility and kinetic of starch digestion of spaghetti with persimmon (Dyospyros kaki) flours coproducts during in vitro gastrointestinal digestion. Food Chem. 2021, 338, 128142. [Google Scholar] [CrossRef] [PubMed]
- Panagopoulou, E.A.; Chiou, A.; Kasimatis, T.-D.; Bismpikis, M.; Mouraka, P.; Karathanos, V.T. Dried dates: Polar phenols and their fate during in vitro digestion. J. Food Meas. Charact. 2021, 15, 1899–1906. [Google Scholar] [CrossRef]
- Agudelo, C.D.; Luzardo-Ocampo, I.; Campos-Vega, R.; Loarca-Piña, G.; Maldonado-Celis, M.E. Bioaccessibility during In Vitro Digestion and Antiproliferative Effect of Bioactive Compounds from Andean Berry (Vaccinium meridionale Swartz) Juice. J. Agric. Food Chem. 2018, 66, 7358–7366. [Google Scholar] [CrossRef]
- Zhu, L.; Li, W.; Deng, Z.; Li, H.; Zhang, B. The Composition and Antioxidant Activity of Bound Phenolics in Three Legumes, and Their Metabolism and Bioaccessibility of Gastrointestinal Tract. Foods 2020, 9, 1816. [Google Scholar] [CrossRef] [PubMed]
- Bouayed, J.; Hoffmann, L.; Bohn, T. Total phenolics, flavonoids, anthocyanins and antioxidant activity following simulated gastro-intestinal digestion and dialysis of apple varieties: Bioaccessibility and potential uptake. Food Chem. 2011, 128, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Lafay, S.; Gil-Izquierdo, A. Bioavailability of phenolic acids. Phytochem. Rev. 2007, 7, 301. [Google Scholar] [CrossRef]
- McDougall, G.J.; Dobson, P.; Smith, P.; Blake, A.; Stewart, D. Assessing Potential Bioavailability of Raspberry Anthocyanins Using an in Vitro Digestion System. J. Agric. Food Chem. 2005, 53, 5896–5904. [Google Scholar] [CrossRef]
- Pineda-Vadillo, C.; Nau, F.; Dubiard, C.G.; Cheynier, V.; Meudec, E.; Sanz-Buenhombre, M.; Guadarrama, A.; Tóth, T.; Csavajda, É.; Hingyi, H. In vitro digestion of dairy and egg products enriched with grape extracts: Effect of the food matrix on polyphenol bioaccessibility and antioxidant activity. Food Res. Int. 2016, 88, 284–292. [Google Scholar] [CrossRef]
- Herrera-Cazares, L.A.; Hernández-Navarro, F.; Ramírez-Jiménez, A.K.; Campos-Vega, R.; de la Luz Reyes-Vega, M.; Loarca-Piña, G.; Morales-Sánchez, E.; Wall-Medrano, A.; Gaytán-Martínez, M. Mango-bagasse functional-confectionery: Vehicle for enhancing bioaccessibility and permeability of phenolic compounds. Food Funct. 2017, 8, 3906–3916. [Google Scholar] [CrossRef] [PubMed]
- Ordoñez-Díaz, J.L.; Moreno-Ortega, A.; Roldán-Guerra, F.J.; Ortíz-Somovilla, V.; Moreno-Rojas, J.M.; Pereira-Caro, G. In Vitro Gastrointestinal Digestion and Colonic Catabolism of Mango (Mangifera indica L.) Pulp Polyphenols. Foods 2020, 9, 1836. [Google Scholar] [CrossRef] [PubMed]
- Carnachan, S.M.; Bootten, T.J.; Mishra, S.; Monro, J.A.; Sims, I.M. Effects of simulated digestion in vitro on cell wall polysaccharides from kiwifruit (Actinidia spp.). Food Chem. 2012, 133, 132–139. [Google Scholar] [CrossRef]
- Goulas, V.; Hadjisolomou, A. Dynamic changes in targeted phenolic compounds and antioxidant potency of carob fruit (Ceratonia siliqua L.) products during in vitro digestion. LWT 2019, 101, 269–275. [Google Scholar] [CrossRef]
- Wang, C.; Wu, H.; Liu, Z.; Barrow, C.; Dunshea, F.; Suleria, H.A. Bioaccessibility and movement of phenolic compounds from tomato (Solanum lycopersicum) during in vitro gastrointestinal digestion and colonic fermentation. Food Funct. 2022, 13, 4954–4966. [Google Scholar] [CrossRef] [PubMed]
- Ortega, N.; Macià, A.; Romero, M.-P.; Reguant, J.; Motilva, M.-J. Matrix composition effect on the digestibility of carob flour phenols by an in-vitro digestion model. Food Chem. 2011, 124, 65–71. [Google Scholar] [CrossRef]
- Correa-Betanzo, J.; Allen-Vercoe, E.; McDonald, J.; Schroeter, K.; Corredig, M.; Paliyath, G. Stability and biological activity of wild blueberry (Vaccinium angustifolium) polyphenols during simulated in vitro gastrointestinal digestion. Food Chem. 2014, 165, 522–531. [Google Scholar] [CrossRef]
- Çalışkantürk Karataş, S.; Günay, D.; Sayar, S. In vitro evaluation of whole faba bean and its seed coat as a potential source of functional food components. Food Chem. 2017, 230, 182–188. [Google Scholar] [CrossRef]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.M.; De Souza, R.; Kendall, C.W.; Emam, A.; Jenkins, D.J. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 2006, 40, 235–243. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Compound | Oral Bioaccessibility (%) | Gastric Bioaccessibility (%) | Intestinal Bioaccessibility (%) | Colonic Bioaccessibility (%) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AB * | MB | BUW | BEB | AB | MB | BUW | BEB | AB | MB | BUW | BEB | AB | MB | BUW | BEB | ||
| 1 | Gallic acid | 179.12 | 99.30 | 84.82 | 76.32 | 91.91 | 98.81 | 84.63 | 75.91 | 92.11 | 98.77 | 84.75 | 76.52 | 97.02 | 108.96 | 94.57 | 81.18 |
| 2 | Protocatechuic acid | - | 100.61 | 0.00 | - | - | 100.70 | 0.00 | - | - | 101.15 | 0.00 | - | - | 103.83 | 103.29 | - |
| 3 | Caftaric acid | 94.51 | 100.11 | - | - | 99.46 | 0.00 | - | - | 95.00 | 0.00 | - | - | 0.00 | 0.00 | - | - |
| 4 | p-hydroxybenzoic acid | 123.82 | 121.26 | 100.23 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 100.13 |
| 5 | Chlorogenic acid | 102.46 | - | - | 101.82 | 101.35 | - | - | 100.41 | 101.20 | - | - | 100.70 | 101.14 | - | - | 0.00 |
| 6 | Coumaric acid | - | 98.87 | - | - | - | 0.00 | - | - | - | 0.00 | - | - | - | 0.00 | - | - |
| 7 | Kaempferol | - | - | 100.44 | 0.00 | - | - | 100.20 | 102.60 | - | - | 101.15 | 102.92 | - | - | 0.00 | 102.98 |
| Total Phenolic compounds | 157.25 | 179.85 | 106.48 | 79.20 | 102.35 | 32.07 | 66.40 | 130.75 | 100.76 | 32.14 | 66.94 | 102.09 | 167.54 | 84.95 | 62.90 | 96.76 | |
| No. | Compound | Intestinal Total Recovery (%) | Colonic Total Recovery (%) | Residual Intestinal Digesta Index (RID %) | Residual Colonic Digesta Index (RCD %) | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AB * | MB | BUW | BEB | AB | MB | BUW | BEB | AB | MB | BUW | BEB | AB | MB | BUW | BEB | ||
| 1 | Gallic acid | 191.54 | 206.89 | 177.41 | 159.08 | 188.82 | 208.35 | 183.00 | 158.52 | 99.43 | 108.12 | 92.65 | 82.56 | 91.79 | 99.38 | 88.43 | 77.33 |
| 2 | Protocatechuic acid | - | 207.74 | 106.36 | - | - | 103.83 | 204.02 | - | - | 106.58 | 106.36 | - | - | 0.00 | 100.73 | - |
| 3 | Caftaric acid | 95.00 | 0.00 | - | - | 95.50 | 0.00 | - | - | 0.00 | 0.00 | - | - | 95.50 | 0.00 | - | - |
| 4 | p-hydroxybenzoic acid | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 100.13 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| 5 | Chlorogenic acid | 101.20 | - | - | 200.91 | 101.14 | - | - | 102.12 | 0.00 | - | - | 100.21 | 0.00 | - | - | 102.12 |
| 6 | Coumaric acid | - | 0.00 | - | - | - | 0.00 | - | - | - | 0.00 | - | - | - | 0.00 | - | - |
| 7 | Kaempferol | - | - | 101.15 | 102.92 | - | - | 0.00 | 102.98 | - | - | 0.00 | 0.00 | - | - | 0.00 | 0.00 |
| Total Phenolic compounds | 213.68 | 170.25 | 146.02 | 183.85 | 216.51 | 99.31 | 87.22 | 128.38 | 112.92 | 138.11 | 79.08 | 81.76 | 48.97 | 14.36 | 24.32 | 31.62 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Bai, Q.; Zhou, J.; de Souza, T.S.P.; Suleria, H.A.R. In Vitro Gastrointestinal Bioaccessibility, Bioactivities and Colonic Fermentation of Phenolic Compounds in Different Vigna Beans. Foods 2022, 11, 3884. https://doi.org/10.3390/foods11233884
Li M, Bai Q, Zhou J, de Souza TSP, Suleria HAR. In Vitro Gastrointestinal Bioaccessibility, Bioactivities and Colonic Fermentation of Phenolic Compounds in Different Vigna Beans. Foods. 2022; 11(23):3884. https://doi.org/10.3390/foods11233884
Chicago/Turabian StyleLi, Minhao, Qian Bai, Jiajing Zhou, Thaiza Serrano Pinheiro de Souza, and Hafiz Ansar Rasul Suleria. 2022. "In Vitro Gastrointestinal Bioaccessibility, Bioactivities and Colonic Fermentation of Phenolic Compounds in Different Vigna Beans" Foods 11, no. 23: 3884. https://doi.org/10.3390/foods11233884
APA StyleLi, M., Bai, Q., Zhou, J., de Souza, T. S. P., & Suleria, H. A. R. (2022). In Vitro Gastrointestinal Bioaccessibility, Bioactivities and Colonic Fermentation of Phenolic Compounds in Different Vigna Beans. Foods, 11(23), 3884. https://doi.org/10.3390/foods11233884