
Bioprocessing of Barley and Lentil Grains to Obtain In Situ Synthesis of Exopolysaccharides and Composite Wheat Bread with Improved Texture and Health Properties

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Liquid Sourdough Fermentation
2.3. pH and Viscosity Measurement
2.4. Enumeration of Cultivable Bacteria and Yeasts
2.5. Determination of Dextran, Sugars and Organic Acids
2.6. Bread Making Trials
2.7. Bread Technological Characterization
2.8. Breads Nutritional Characterization
2.9. Volatile Organic Compounds Profile of Breads
2.10. Bread Sensory Analysis
2.11. Statistical Analysis
3. Results and Discussion
3.1. pH and Viscosity in Preliminary Liquid Sourdough Formulations
3.2. Microbial Growth and Acidification in the Liquid Sourdough Formulations
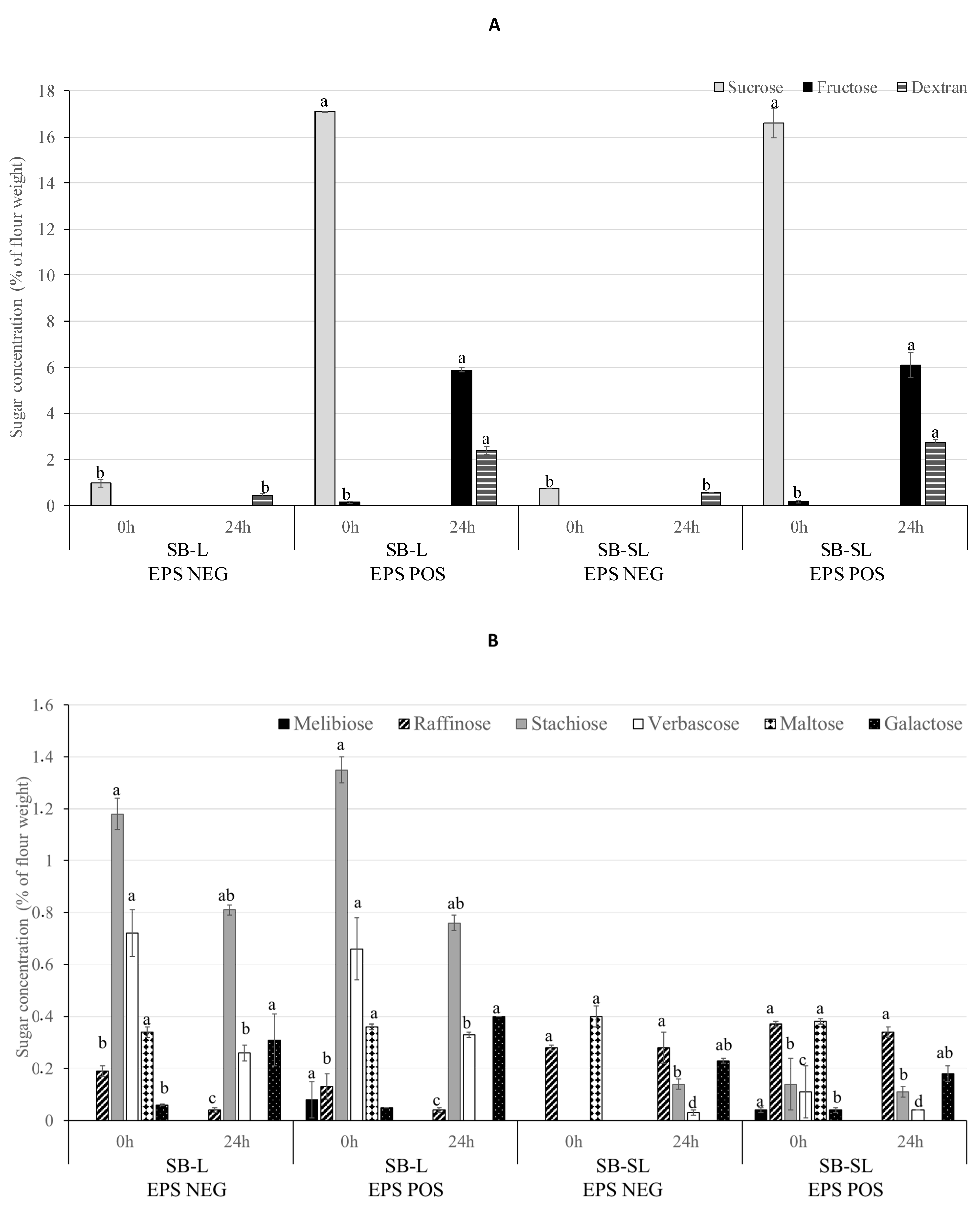
3.3. Sugars and Dextran Content in the Liquid Sourdough Formulations
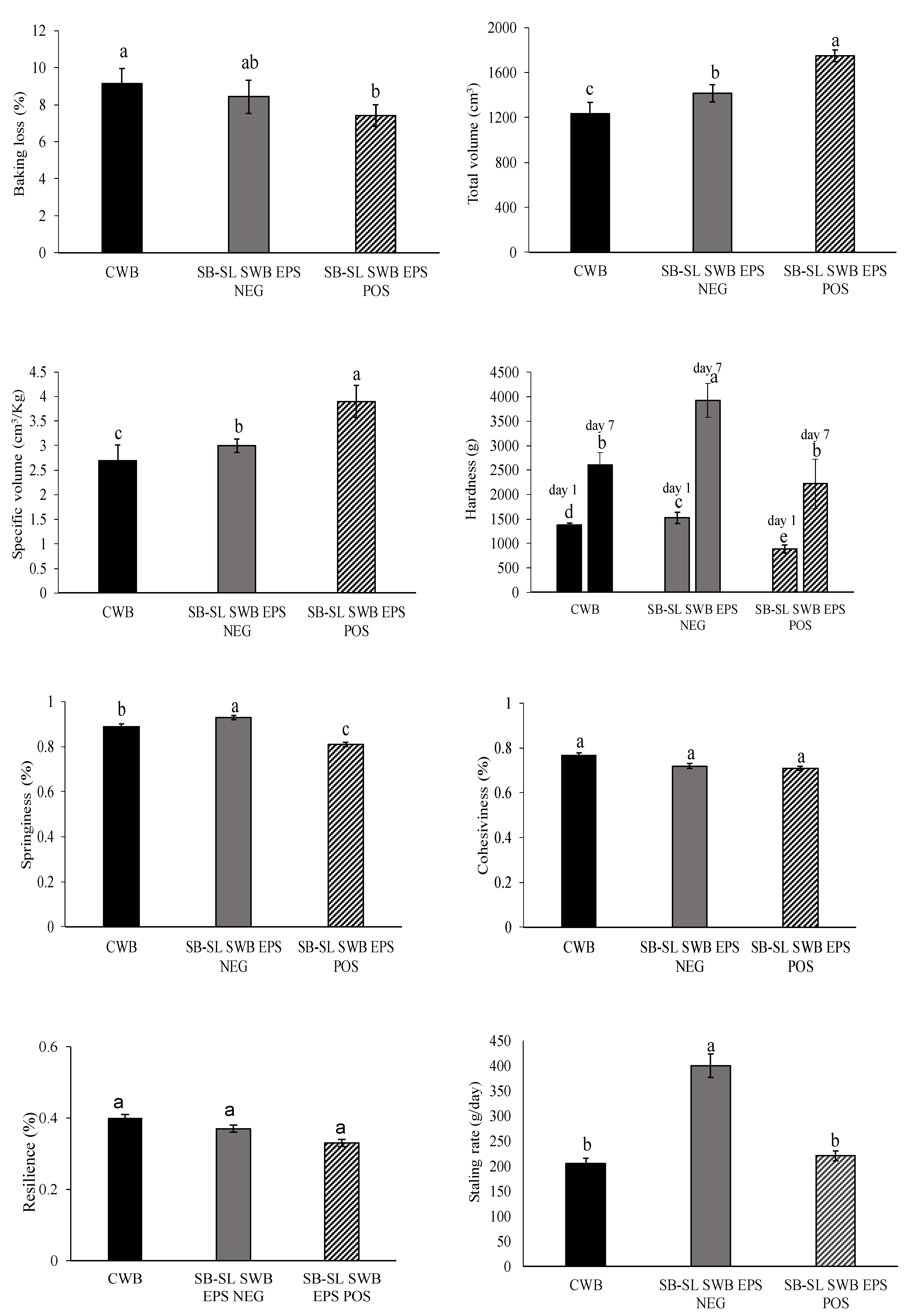
3.4. Breads Characterization
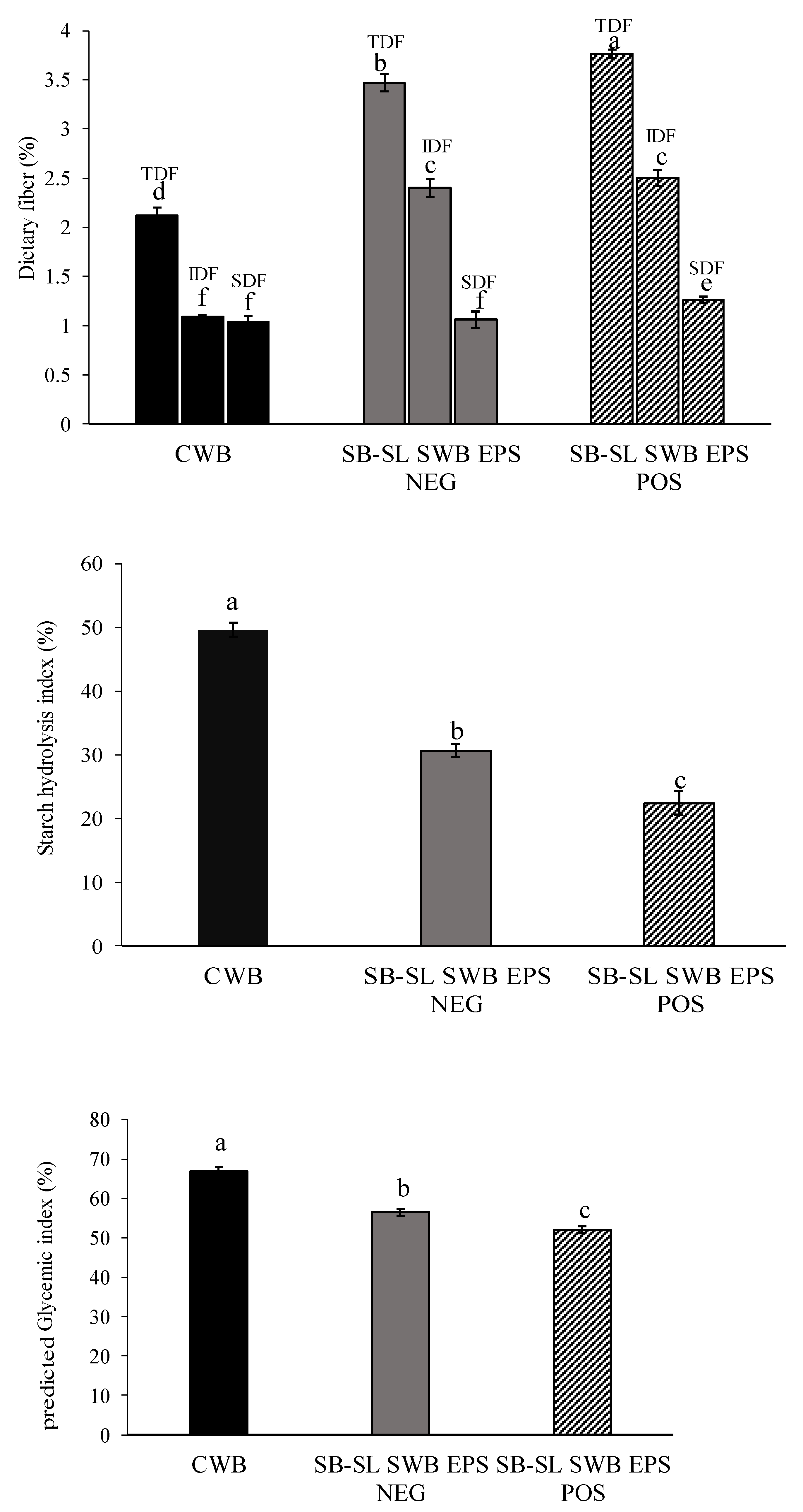
3.5. Dietary Fiber and Starch Hydrolysis Index
3.6. Volatile Organic Compounds
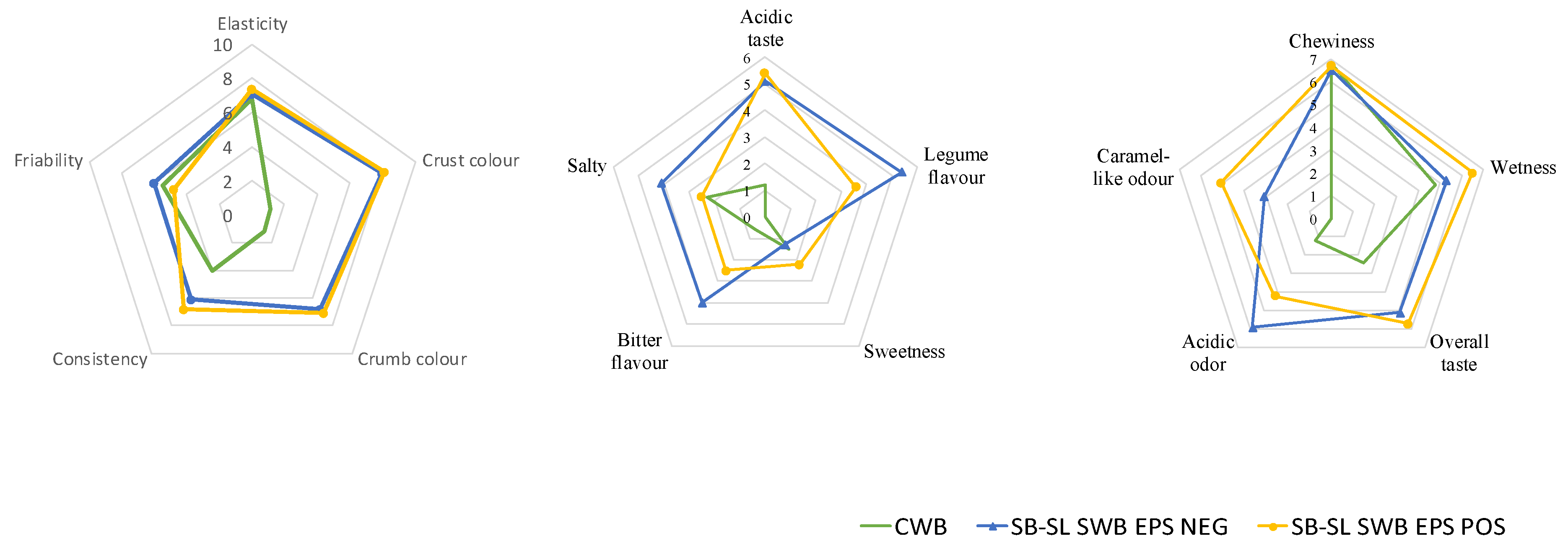
3.7. Sensory Profile
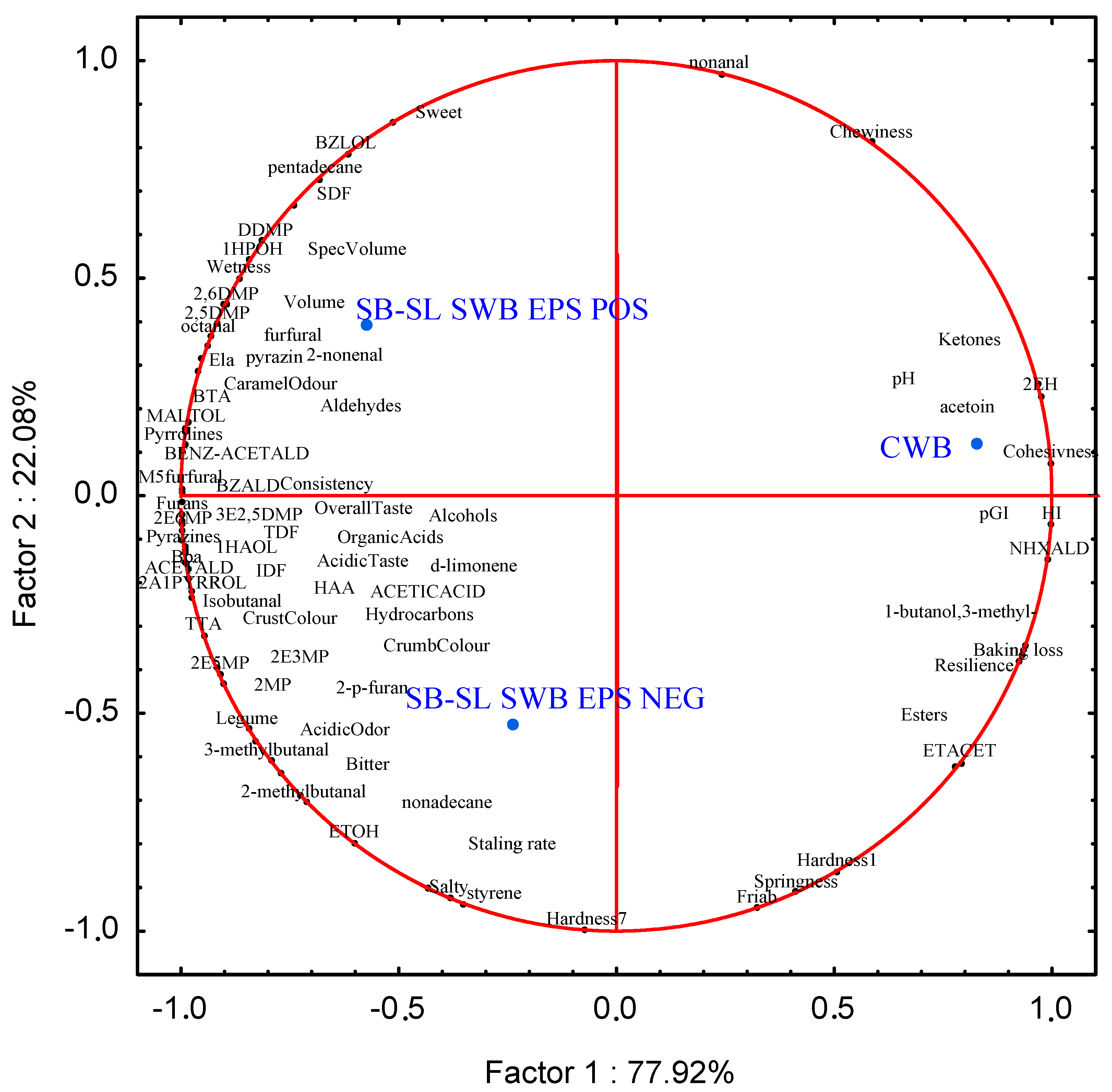
3.8. Correlations between VOCs, Technological, Nutritional and Sensory Features of Breads
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Nkhata, S.G.; Ayua, E.; Kamau, E.H.; Shingiro, J.B. Fermentation and germination improve nutritional value of cereals and legumes through activation of endogenous enzymes. Food Sci. Nutr. 2018, 6, 2446–2458. [Google Scholar] [CrossRef] [Green Version]
- Mathur, H.; Beresford, T.P.; Cotter, P.D. Health benefits of Lactic Acid Bacteria (LAB) Fermentates. Nutrients 2020, 12, 1679. [Google Scholar] [CrossRef] [PubMed]
- Röcken, W.; Voysey, P.A. Sourdough fermentation in bread making. J. Appl. Microbiol. 1995, 79, 38S–48S. [Google Scholar]
- Katina, K.; Liukkonen, K.H.; Kaukovirta-Norja, A.; Adlercreutz, H.; Heinonen, S.M.; Lampi, A.M.; Pihlava, J.-M.; Poutanen, K. Fermentation-induced changes in the nutritional value of native or germinated rye. J. Cereal Sci. 2007, 46, 348–355. [Google Scholar] [CrossRef]
- Leroy, F.; De Vuyst, L. Lactic acid bacteria as functional starter cultures for the food fermentation industry. Trends Food Sci. Technol. 2004, 15, 67–78. [Google Scholar] [CrossRef]
- Gobbetti, M.; De Angelis, M.; Di Cagno, R.; Calasso, M.; Archetti, G.; Rizzello, C.G. Novel insights on the functional/nutritional features of the sourdough fermentation. Int. J. Food Microbiol. 2019, 302, 103–113. [Google Scholar] [CrossRef]
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligné, B.; Gänzle, M.; Kort, R.; Pasin, G.; Pihlanto, A.; et al. Health benefits of fermented foods: Microbiota and beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Liukkonen, K.H.; Katina, K.; Wilhelmsson, A.; Myllymäki, O.; Lampi, A.M.; Kariluoto, S.; Piironen, V.; Heinonen, S.M.; Nurmi, T.; Adlercreutz, H.; et al. Process-induced changes on bioactive compounds in whole grain rye. Proc. Nutr. Soc. 2003, 62, 117–122. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Portincasa, P.; Montemurro, M.; Di Palo, D.M.; Lorusso, M.P.; De Angelis, M.; Bonfrate, L.; Genot, B.; Gobbetti, M. Sourdough fermented breads are more digestible than those started with baker’s yeast alone: An in vivo challenge dissecting distinct gastrointestinal responses. Nutrients 2019, 11, 2954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montemurro, M.; Coda, R.; Rizzello, C.G. Recent advances in the use of sourdough biotechnology in pasta making. Foods 2019, 8, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vuyst, L.; Harth, H.; Van Kerrebroeck, S.; Leroy, F. Yeast diversity of sourdoughs and associated metabolic properties and functionalities. Int. J. Food Microbiol. 2016, 239, 26–34. [Google Scholar] [CrossRef]
- Gobbetti, M.; Rizzello, C.G.; Di Cagno, R.; De Angelis, M. How the sourdough may affect the functional features of leavened baked goods. Food Microbiol. 2014, 37, 30–40. [Google Scholar] [CrossRef]
- Bockwoldt, J.A.; Fellermeier, J.; Steffens, E.; Vogel, R.F.; Ehrmann, M.A. β-Glucan production by Levilactobacillus brevis and Pediococcus claussenii for in situ enriched rye and wheat sourdough breads. Foods 2021, 10, 547. [Google Scholar] [CrossRef]
- Galle, S.; Arendt, E.K. Exopolysaccharides from sourdough lactic acid bacteria. Crit. Rev. Food Sci. Nutr. 2014, 54, 891–901. [Google Scholar] [CrossRef]
- Monsan, P.; Bozonnet, S.; Ecile Albenne, C.; Joucla, G. Homopolysaccharides from lactic acid bacteria. Int. Dairy J. 2001, 11, 675–685. [Google Scholar] [CrossRef]
- Korakli, M.; Gänzle, M.G.; Vogel, R.F. Metabolism by bifidobacteria and lactic acid bacteria of polysaccharides from wheat and rye, and exopolysaccharides produced by Lactobacillus sanfranciscensis. J. Appl. Microbiol. 2002, 92, 958–965. [Google Scholar] [CrossRef]
- Tieking, M.; Gänzle, M.G. Exopolysaccharides from cereal-associated lactobacilli. Trends Food Sci. Technol. 2005, 16, 79–84. [Google Scholar] [CrossRef]
- Lynch, K.M.; Zannini, E.; Coffey, A.; Arendt, E.K. Lactic acid bacteria exopolysaccharides in foods and beverages: Isolation, properties, characterization, and health benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 155–176. [Google Scholar] [CrossRef] [PubMed]
- Lynch, K.M.; Coffey, A.; Arendt, E.K. Exopolysaccharide producing lactic acid bacteria: Their techno-functional role and potential application in gluten-free bread products. Food Res. Int. 2018, 110, 52–61. [Google Scholar] [CrossRef]
- Salazar, N.; Gueimonde, M.; De Los Reyes-Gavilán, C.G.; Ruas-Madiedo, P. Exopolysaccharides produced by lactic acid bacteria and bifidobacteria as fermentable substrates by the intestinal microbiota. Crit. Rev. Food Sci. Nutr. 2016, 56, 1440–1453. [Google Scholar] [CrossRef]
- Saadat, Y.R.; Khosroushahi, A.Y.; Gargari, B.P. A comprehensive review of anticancer, immunomodulatory and health beneficial effects of the lactic acid bacteria exopolysaccharides. Carbohydr. Polym. 2019, 217, 79–89. [Google Scholar] [CrossRef]
- González-Garcinuño, Á.; Tabernero, A.; Domínguez, Á.; Galán, M.A.; Martin del Valle, E.M. Levan and levansucrases: Polymer, enzyme, micro-organisms and biomedical applications. Biocatal. Biotransform. 2018, 36, 233–244. [Google Scholar] [CrossRef]
- Hundschell, C.S.; Wagemans, A.M. Rheology of common uncharged exopolysaccharides for food applications. Curr. Opin. Food Sci. 2019, 27, 1–7. [Google Scholar] [CrossRef]
- Wolter, A.; Hager, A.S.; Zannini, E.; Galle, S.; Gänzle, M.G.; Waters, D.M.; Arendt, E.K. Evaluation of exopolysaccharide producing Weissella cibaria MG1 strain for the production of sourdough from various flours. Food Microbiol. 2014, 37, 44–50. [Google Scholar] [CrossRef]
- Valerio, F.; Bavaro, A.R.; Di Biase, M.; Lonigro, S.L.; Logrieco, A.F.; Lavermicocca, P. Effect of amaranth and quinoa flours on exopolysaccharide production and protein profile of liquid sourdough fermented by Weissella cibaria and Lactobacillus plantarum. Front. Microbiol. 2020, 11, 967. [Google Scholar] [CrossRef]
- Wang, Y.; Sorvali, P.; Laitila, A.; Maina, N.H.; Coda, R.; Katina, K. Dextran produced in situ as a tool to improve the quality of wheat-faba bean composite bread. Food Hydrocoll. 2018, 84, 396–405. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Compaoré-Sérémé, D.; Sawadogo-Lingani, H.; Coda, R.; Katina, K.; Maina, N.H. Influence of dextran synthesized in situ on the rheological, technological and nutritional properties of whole grain pearl millet bread. Food Chem. 2019, 285, 221–230. [Google Scholar] [CrossRef]
- Wang, Y.; Trani, A.; Knaapila, A.; Hietala, S.; Coda, R.; Katina, K.; Maina, N.H. The effect of in situ produced dextran on flavour and texture perception of wholegrain sorghum bread. Food Hydrocoll. 2020, 106, 105913. [Google Scholar] [CrossRef]
- Falcinelli, B.; Calzuola, I.; Gigliarelli, L.; Torricelli, R.; Polegri, L.; Vizioli, V.; Benincasa, P.; Marsili, V. Phenolic content and antioxidant activity of wholegrain breads from modern and old wheat (Triticum aestivum L.) cultivars and ancestors enriched with wheat sprout powder. Ital. J. Agron. 2018, 13, 297–302. [Google Scholar] [CrossRef]
- Marti, A.; Cardone, G.; Pagani, M.A.; Casiraghi, M.C. Flour from sprouted wheat as a new ingredient in bread-making. LWT 2018, 89, 237–243. [Google Scholar] [CrossRef] [Green Version]
- Benincasa, P.; Falcinelli, B.; Lutts, S.; Stagnari, F.; Galieni, A. Sprouted grains: A comprehensive review. Nutrients 2019, 11, 421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rico, D.; Peñas, E.; García, M.D.C.; Martínez-Villaluenga, C.; Rai, D.K.; Birsan, R.I.; Frias, J.; Martín-Diana, A.B. Sprouted barley flour as a nutritious and functional ingredient. Foods 2020, 9, 296. [Google Scholar] [CrossRef] [Green Version]
- Montemurro, M.; Pontonio, E.; Gobbetti, M.; Rizzello, C.G. Investigation of the nutritional, functional and technological effects of the sourdough fermentation of sprouted flours. Int. J. Food Microbiol. 2019, 302, 47–58. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Aguilar, C.; Dominguez-Pacheco, A.; Tenango, M.P.; Valderrama-Bravo, C.; Hernández, M.S.; Cruz-Orea, A.; Ordonez-Miranda, J. Lentil sprouts: A nutraceutical alternative for the elaboration of bread. J. Food Sci. Technol. 2019, 57, 1817–1829. [Google Scholar] [CrossRef]
- Perri, G.; Coda, R.; Rizzello, C.G.; Celano, G.; Ampollini, M.; Gobbetti, M.; De Angelis, M.; Calasso, M. Sourdough fermentation of whole and sprouted lentil flours: In situ formation of dextran and effects on the nutritional, texture and sensory characteristics of white bread. Food Chem. 2021, 355, 129638. [Google Scholar] [CrossRef] [PubMed]
- Galle, S.; Schwab, C.; Dal Bello, F.; Coffey, A.; Gänzle, M.G.; Arendt, E.K. Influence of in-situ synthesized exopolysaccharides on the quality of gluten-free sorghum sourdough bread. Int. J. Food Microbiol. 2012, 155, 105–112. [Google Scholar] [CrossRef]
- Katina, K.; Maina, N.H.; Juvonen, R.; Flander, L.; Johansson, L.; Virkki, L.; Tenkanen, M.; Laitila, A. In situ production and analysis of Weissella confusa dextran in wheat sourdough. Food Microbiol. 2009, 26, 734–743. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, P.; Arendt, E.; Gallagher, E. The increasing use of barley and barley by-products in the production of healthier baked goods. Trends Food Sci. Technol. 2013, 29, 124–134. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA) 2011a. Scientific opinion on the substantiation of health claims related to beta-glucans from oats and barley and maintenance of normal blood LDL-cholesterol concentrations (ID1236, 1299); increase in satiety leading to a reduction in energy intake (ID 851, 852), reduction of post-prandial glycemic responses (ID 821 824), and “digestive function” (ID 850) pursuant to Article 13(1) of Regulation (EC) No 1924/2006. EFSA J. 2011, 9, 2207. [Google Scholar]
- European Food Safety Authority (EFSA). Scientific opinion on the substantiation of a health claim related to barley beta-glucans and lowering of blood cholesterol and reduced risk of (coronary) heart disease pursuant to Article 14 of Regulation (EC) No 1924/2006. EFSA J. 2011, 9, 2471. [Google Scholar] [CrossRef]
- Food and Drug Administration, Department of Health and Human Services. Federal Register—71 FR 29248: Health Claims; Soluble Dietary Fiber from Certain Foods and Coronary Heart Disease Final Rule; FDA: Silver Spring, MD, USA, 2006; Volume 71, pp. 29248–29250. [Google Scholar]
- Faris, M.E.A.I.E.; Takruri, H.R.; Issa, A.Y. Role of lentils (Lens culinaris L.) in human health and nutrition: A review. Med. J. Nutr. Metab. 2013, 6, 3–16. [Google Scholar] [CrossRef]
- Santos, S.C.; Silva, B.; Valente, L.M.P.; Gruber, S.; Vasconcelos, M.W. The effect of sprouting in lentil (Lens culinaris) nutritional and microbiological profile. Foods 2020, 9, 400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Wang, Y.; Coda, R.; Säde, E.; Tuomainen, P.; Tenkanen, M.; Katina, K. In situ synthesis of exopolysaccharides by Leuconostoc spp. and Weissella spp. and their rheological impacts in fava bean flour. Int. J. Food Microbiol. 2017, 248, 63–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perri, G.; Calabrese, F.M.; Rizzello, C.G.; De Angelis, M.; Gobbetti, M.; Calasso, M. Sprouting process affects the lactic acid bacteria and yeasts of cereal, pseudocereal and legume flours. LWT 2020, 126, 109314. [Google Scholar] [CrossRef]
- Xu, Y.; Coda, R.; Holopainen-Mantila, U.; Laitila, A.; Katina, K.; Tenkanen, M. Impact of in situ produced exopolysaccharides on rheology and texture of fava bean protein concentrate. Food Res. Int. 2019, 115, 191–199. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Calasso, M.; Campanella, D.; De Angelis, M.; Gobbetti, M. Use of sourdough fermentation and mixture of wheat, chickpea, lentil and bean flours for enhancing the nutritional, texture and sensory characteristics of white bread. Int. J. Food Microbiol. 2014, 180, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Regulation (EC) No 1924/2006 of the European Parliament and of the Council of 20 December 2006 on Nutrition and Health Claims Made on Foods. Official Journal L 404, 30 December 2006, p. 9–25. Available online: https://eur-lex.europa.eu/eli/reg/2006/1924/corrigendum/2007-01-18/oj (accessed on 26 June 2021).
- AACC International. Approved Methods of the American Association of Cereal Chemists, 11th ed.; AACC: St. Paul, MN, USA, 2000. [Google Scholar]
- Katina, K.; Heiniö, R.L.; Autio, K.; Poutanen, K. Optimization of sourdough process for improved sensory profile and texture of wheat bread. LWT 2006, 39, 1189–1202. [Google Scholar] [CrossRef]
- Curiel, J.A.; Coda, R.; Centomani, I.; Summo, C.; Gobbetti, M.; Rizzello, C.G. Exploitation of the nutritional and functional characteristics of traditional Italian legumes: The potential of sourdough fermentation. Int. J. Food Microbiol. 2015, 196, 51–61. [Google Scholar] [CrossRef]
- Crowley, P.; Grau, H.; Arendt, E.K. Influence of additives and mixing time on crumb grain characteristics of wheat bread. Cereal Chem. 2000, 77, 370–375. [Google Scholar] [CrossRef]
- Liljeberg, H.; Åkerberg, A.; Björck, I. Resistant starch formation in bread as influenced by choice of ingredients or baking conditions. Food Chem. 1996, 56, 389–394. [Google Scholar] [CrossRef]
- Goñi, I.; Garcia-Alonso, A.; Saura-Calixto, F. A starch hydrolysis procedure to estimate glycemic index. Nutr. Res. 1997, 17, 427–437. [Google Scholar] [CrossRef]
- Tobaruela, E.D.C.; Santos, A.D.O.; de Almeida-Muradian, L.B.; Araujo, E.D.S.; Lajolo, F.M.; Menezes, E.W. Application of dietary fiber method AOAC 2011.25 in fruit and comparison with AOAC 991.43 method. Food Chem. 2018, 238, 87–93. [Google Scholar] [CrossRef]
- Pico, J.; Khomenko, I.; Capozzi, V.; Navarini, L.; Bernal, J.; Gómez, M.; Biasioli, F. Analysis of volatile organic compounds in crumb and crust of different baked and toasted gluten-free breads by direct PTR-ToF-MS and fast-GC-PTR-ToF-MS. J. Mass Spectrom. 2018, 53, 893–902. [Google Scholar] [CrossRef] [PubMed]
- Cavallo, N.; De Angelis, M.; Calasso, M.; Quinto, M.; Mentana, A.; Minervini, F.; Cappelle, S.; Gobbetti, M. Microbial cell-free extracts affect the biochemical characteristics and sensorial quality of sourdough bread. Food Chem. 2017, 237, 159–168. [Google Scholar] [CrossRef]
- Korakli, M.; Vogel, R.F. Structure/function relationship of homopolysaccharide producing glycansucrases and therapeutic potential of their synthesised glycans. Appl. Microbiol. Biotechnol. 2006, 71, 790–803. [Google Scholar] [CrossRef] [PubMed]
- Galle, S.; Schwab, C.; Arendt, E.; Gänzle, M. Exopolysaccharide-forming Weissella strains as starter cultures for sorghum and wheat sourdoughs. J. Agric. Food Chem. 2010, 58, 5834–5841. [Google Scholar] [CrossRef]
- Wisselink, H.W.; Weusthuis, R.A.; Eggink, G.; Hugenholtz, J.; Grobben, G.J. Mannitol production by lactic acid bacteria: A review. Int. Dairy J. 2002, 12, 151–161. [Google Scholar] [CrossRef]
- Zannini, E.; Mauch, A.; Galle, S.; Gänzle, M.; Coffey, A.; Arendt, E.K.; Taylor, G.P.; Waters, D.M. Barley malt wort fermentation by exopolysaccharide-forming Weissella cibaria MG1 for the production of a novel beverage. J. Appl. Microbiol. 2013, 115, 1379–1387. [Google Scholar] [CrossRef]
- Wang, T.L.; Domoney, C.; Hedley, C.L.; Casey, R.; Grusak, M.A. Can we improve the nutritional quality of legume seeds? Plant Physiol. 2003, 131, 886–891. [Google Scholar] [CrossRef] [Green Version]
- Duszkiewicz-Reinhard, W.A.N.D.A.; Gujska, E.; Khan, K. Reduction of stachyose in legume flours by lactic acid bacteria. J. Food Sci. 1994, 59, 115–117. [Google Scholar] [CrossRef]
- Yoon, M.Y.; Hwang, H.J. Reduction of soybean oligosaccharides and properties of α-D-galactosidase from Lactobacillus curvatus R08 and Leuconostoc mesenteriodes JK55. Food Microbiol. 2008, 25, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Waters, D.M.; Mauch, A.; Coffey, A.; Arendt, E.K.; Zannini, E. Lactic acid bacteria as a cell factory for the delivery of functional biomolecules and ingredients in cereal-based beverages: A review. Crit. Rev. Food Sci. Nutr. 2015, 55, 503–520. [Google Scholar] [CrossRef] [PubMed]
- Ferrero, C. Hydrocolloids in wheat breadmaking: A concise review. Food Hydrocoll. 2017, 68, 15–22. [Google Scholar] [CrossRef]
- Kohajdová, Z.; Karovičová, J.; Magala, M. Effect of lentil and bean flours on rheological and baking properties of wheat dough. Chem. Pap. 2013, 67, 398–407. [Google Scholar] [CrossRef]
- Fadda, C.; Sanguinetti, A.M.; Del Caro, A.; Collar, C.; Piga, A. Bread staling: Updating the view. Compr. Rev. Food Sci. Food Saf. 2014, 13, 473–492. [Google Scholar] [CrossRef] [Green Version]
- Gray, J.A.; Bemiller, J.N. Bread staling: Molecular basis and control. Compr. Rev. Food Sci. Food Saf. 2003, 2, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Farzaneh, V.; Ghodsvali, A.; Bakhshabadi, H.; Zare, Z.; Carvalho, I.S. The impact of germination time on some selected parameters through malting process. Int. J. Biol. Macromol. 2017, 94, 663–668. [Google Scholar] [CrossRef]
- Charoenthaikij, P.; Jangchud, K.; Jangchud, A.; Prinyawiwatkul, W.; No, H.K. Composite wheat–germinated brown rice flours: Selected physicochemical properties and bread application. Int. J. Food Sci. Technol. 2012, 47, 75–82. [Google Scholar] [CrossRef]
- De Angelis, M.; Damiano, N.; Rizzello, C.G.; Cassone, A.; Di Cagno, R.; Gobbetti, M. Sourdough fermentation as a tool for the manufacture of low-glycemic index white wheat bread enriched in dietary fibre. Eur. Food Res. Technol. 2009, 229, 593–601. [Google Scholar] [CrossRef]
- Henry, R.J. Pentosan and (1→ 3), (1→ 4)-β-glucan concentrations in endosperm and wholegrain of wheat, barley, oats and rye. J. Cereal Sci. 1987, 6, 253–258. [Google Scholar] [CrossRef]
- Wood, P.; Beer, M.U.; Butler, G. Evaluation of role of concentration and molecular weight of oat β-glucan in determining effect of viscosity on plasma glucose and insulin following an oral glucose load. Br. J. Nutr. 2000, 84, 19–23. [Google Scholar] [CrossRef] [Green Version]
- Marklinder, I.; Johansson, L. Sourdough fermentation of barley flours with varied content of mixed-linked (1→ 3),(1→ 4) β-d-glucans. Food Microbiol. 1995, 12, 363–371. [Google Scholar] [CrossRef]
- Mariotti, M.; Garofalo, C.; Aquilanti, L.; Osimani, A.; Fongaro, L.; Tavoletti, S.; Hager, A.S.; Clementi, F. Barley flour exploitation in sourdough bread-making: A technological, nutritional and sensory evaluation. LWT 2014, 59, 973–980. [Google Scholar] [CrossRef]
- Wood, P.J. Cereal β-glucans in diet and health. J. Cereal Sci. 2007, 46, 230–238. [Google Scholar] [CrossRef]
- Ruiz, J.A.; Quilez, J.; Mestres, M.; Guasch, J. Solid-phase microextraction method for headspace analysis of volatile compounds in bread crumb. Cereal Chem. 2003, 80, 255–259. [Google Scholar] [CrossRef]
- Plessas, S.; Bekatorou, A.; Gallanagh, J.; Nigam, P.; Koutinas, A.A.; Psarianos, C. Evolution of aroma volatiles during storage of sourdough breads made by mixed cultures of Kluyveromyces marxianus and Lactobacillus delbrueckii ssp. bulgaricus or Lactobacillus helveticus. Food Chem. 2008, 107, 883–889. [Google Scholar] [CrossRef]
- Hansen, A.; Schieberle, P. Generation of aroma compounds during sourdough fermentation: Applied and fundamental aspects. Trends Food Sci. Technol. 2005, 16, 85–94. [Google Scholar] [CrossRef]
- Heiniö, R.L.; Katina, K.; Wilhelmson, A.; Myllymäki, O.; Rajamäki, T.; Latva-Kala, K.; Liukkonen, K.H.; Poutanen, K. Relationship between sensory perception and flavour-active volatile compounds of germinated, sourdough fermented and native rye following the extrusion process. LWT 2003, 36, 533–545. [Google Scholar] [CrossRef]
- Pétel, C.; Courcoux, P.; Génovesi, N.; Rouillé, J.; Onno, B.; Prost, C. Free Sorting and association Task: A variant of the free sorting method applied to study the impact of dried sourdough as an ingredient on the related bread odor. J. Food Sci. 2017, 82, 985–992. [Google Scholar] [CrossRef]
- Zhao, C.J.; Schieber, A.; Gänzle, M.G. Formation of taste-active amino acids, amino acid derivatives and peptides in food fermentations—A review. Food Res. Int. 2016, 89, 39–47. [Google Scholar] [CrossRef]
- Poutanen, K.; Flander, L.; Katina, K. Sourdough and cereal fermentation in a nutritional perspective. Food Microbiol. 2009, 26, 693–699. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Code | Lactic Acid Bacteria | ΔLog a | Total Mesophilic Bacteria | Enterobacteriaceae | Yeasts | pH | TTA | Lactic Acid | Acetic Acid | FQ b |
|---|---|---|---|---|---|---|---|---|---|---|
| T0 h | ||||||||||
| SB-L EPS NEG | 6.81 ± 0.107 a | 6.62 ± 0.117 b | 1.77 ± 0.097 d | <1 c | 6.27 ± 0.113 a | 4.18 ± 0.093 d | n.d. c | n.d. | - | |
| SB-L EPS POS | 6.84 ± 0.058 a | 6.53 ± 0.143 b | 1.53 ± 0.176 d | 1.14 ± 0.176 b | 6.22 ± 0.044 a | 4.57 ± 0.117 d | n.d. | n.d. | - | |
| SB-SL EPS NEG | 6.78 ± 0.192 a | 6.74 ± 0.117 b | 5.44 ± 0.097 b | 2.35 ± 0.087 a | 6.07 ± 0.095 a | 5.4 ± 0.135 c | n.d. | n.d. | - | |
| SB-SL EPS POS | 6.71 ± 0.144 a | 6.72 ± 0.033 b | 5.53 ± 0.176 b | 2.29 ± 0.014 a | 6.15 ± 0.085 a | 5.6 ± 0.110 c | n.d. | n.d. | - | |
| T24 h | ||||||||||
| SB-L EPS NEG | 9.51 ± 0.044 a | 2.71 ± 0.035 a | 8.87 ± 0.111 a | 4.81 ± 0.063 c | <1 c | 4.62 ± 0.115 b | 11.6 ± 1.241 b | 18.00 ± 0.10 c | 4.38 ± 0.10 c | 4.11± 0.18 |
| SB-L EPS POS | 9.35 ± 0.148 a | 2.51 ± 0.055 a | 9.03 ± 0.201 a | 5.01 ± 0.212 c | <1 c | 4.53± 0.271 b | 12.4 ± 0.115 b | 17.20 ± 0.72 c | 5.45 ± 0.22 b | 3.16 ± 0.13 |
| SB-SL EPS NEG | 9.43 ± 0.152 a | 2.65 ± 0.082 a | 9.14 ± 0.257 a | 5.81 ± 0.091 a | <1 c | 4.44 ± 0.095 bc | 13.6 ± 0.241 ab | 22.00 ± 0.10 a | 5.40 ± 0.10 b | 4.07 ± 0.18 |
| SB-SL EPS POS | 9.55 ± 0.074 a | 2.84 ± 0.112 a | 9.23 ± 0.151 a | 6.01 ± 0.151 a | <1 c | 4.42 ± 0.045 bc | 14.2 ± 0.115 a | 20.20 ± 0.10 b | 7.80 ± 0.22 a | 2.59 ± 0.13 |
| Compounds | Odor | CWB | SL-SB SWB EPS NEG | SL-SB SWB EPS POS |
|---|---|---|---|---|
| Alcohols | ||||
| Ethanol | Alcoholic | 2.03 ± 0.096 b | 3.65 ± 0.074 a | 2.7 ± 0.109 b |
| 3-methylbutanol | Balsamic, alcoholic, malty | 0.65 ± 0.014 a | 0.52 ± 0.025 ab | 0.36 ± 0.004 b |
| 1-hexanol | Green grass, woody, sweet, flowery, mild | 0.18 ± 0.014 b | 1.00 ± 0.163 a | 1.18 ± 0.031 a |
| 1-heptanol | Green | n.d. 1 | 0.01 ± 0.001 b | 0.04 ± 0.015 a |
| 2-ethylhexanol | Green, vegetable | 0.27 ± 0.026 a | n.d. | n.d. |
| Benzylalcohol | Pleasant, aromatic | 0.34 ± 0.135 b | 0.39 ± 0.041 b | 0.90 ± 0.016 a |
| 1-nonanol | Citrus | 0.01 ± 0.003 b | 0.19 ± 0.089 a | 0.27 ± 0.005 a |
| Total | 3.56 ± 0.13 | 5.8 ± 0.63 | 5.45 ± 0.19 | |
| Organic Acids | ||||
| Acetic acid | Sour, acid, pungent | 0.05 ± 0.004 b | 0.50 ± 0.201 a | 0.56 ± 0.013 a |
| Butanoic acid | Sweaty, rancid | n.d. | 0.02 ± 0.001 | 0.03 ± 0 |
| Hexanoic acid | Sweaty, cheesy, fatty, goat-like | 0.04 ± 0.001 b | 0.26 ± 0.073 a | 0.28 ± 0.012 a |
| Total | 0.10 ± 0.02 b | 0.78 ± 0.03 a | 0.87 ± 0.43 a | |
| Aldehydes | ||||
| 2-methylpropanal | Malty | 0.1 ± 0.026 b | 0.19 ± 0.003 a | 0.19 ± 0.009 a |
| 2-methylbutanal | Almond, malty | 0.01 ± 0.001 b | 0.24 ± 0.078 a | 0.13 ± 0.015 ab |
| 3-methylbutanal | Malty, roasty, cucumber-like | 0.01 ± 0.003 b | 0.37 ± 0.122 a | 0.23 ± 0.01 ab |
| Hexanal | Green, grassy | 0.52 ± 0.017 a | 0.39 ± 0.069 b | 0.33 ± 0.015 b |
| Octanal | Citrus, flowery | 0.02 ± 0.029 b | 0.18 ± 0.161 ab | 0.34 ± 0.032 a |
| Nonanal | Citrus, soapy | 0.37 ± 0.036a | 0.26 ± 0.061b | 0.44 ± 0.016a |
| Furfural | Almond, soil | 0.26 ± 0.020b | 1.11 ± 0.430a | 1.90 ± 0.039a |
| Benzaldehyde | Almond, caramel | 0.07 ± 0.009b | 0.32 ± 0.08a | 0.38 ± 0.002a |
| 2-nonenal | Fatty, tallowy, green | 0.02 ± 0.006b | 0.17 ± 0.076ab | 0.28 ± 0.002a |
| Acetaldehyde | Fruity | 0.05 ± 0.003b | 0.23 ± 0.074a | 0.24 ± 0.003a |
| Benzeneacetaldehyde | flowery, honey-like | n.d. | 0.02 ± 0.008a | 0.03 ± 0.002a |
| Total | 1.43 ± 0.03b | 3.48 ± 0.72ab | 4.49 ± 0.10a | |
| Esters | ||||
| Ethylacetate | Sweet, fruity, pineapple | 0.09 ± 0.003ab | 0.11 ± 0.05a | 0.05 ± 0b |
| Total | 0.09 ± 0.00 ab | 0.11 ± 0.00 a | 0.05 ± 0.05 b | |
| Ketones | ||||
| Acetoin | Butter, butterscotch, cream, yogurt | 0.12 ± 0.005 a | 0.06 ± 0.015 b | 0.06 ± 0.009 b |
| Total | 0.12 ± 0.005 a | 0.06 ± 0.015 b | 0.06 ± 0.009 b | |
| Hydrocarbons | ||||
| d-limonene | Citrus | 0.04 ± 0.032 b | 0.18 ± 0.011 a | 0.19 ± 0.031 a |
| Styrene | Pungent | n.d. | 0.06 ± 0.013 a | 0.01 ± 0.021 b |
| Nonadecane | n.f. 2 | n.d. | 0.02 ± 0.014 a | 0.01 ± 0.017 a |
| Pentadecane | n.f. | n.d. | n.d. | 0.02 ± 0 ab |
| 4 h-pyran-4-one,2,3-dihydro-3,5 | Caramelized | n.d. | 0.01 ± 0.014 b | 0.05 ± 0.008 a |
| Total | 0.04 ± 0.032 b | 0.27 ± 0.07 a | 0.28± 0.03 a | |
| Furans | ||||
| 2-pentyl- furan | Butter, green bean, floral | 0.08 ± 0.024 b | 0.32 ± 0.278 ab | 0.28 ± 0.063 a |
| 2-furancarboxaldehyde,5-methyl | n.f. | 0.11 ± 0.015 b | 0.63 ± 0.29 a | 0.81 ± 0.002 a |
| Total | 0.21 ± 0.02 b | 0.95 ± 0.03 a | 1.09 ± 0.07 a | |
| Pyrazines | ||||
| 2-methylpyrazine | Roasted, burnt, sweet | 0.19 ± 0.004 b | 0.32 ± 0.065 a | 0.30 ± 0.004 a |
| 2,5-dimethylpyrazine | Crust-like, popcorn | 0.09 ± 0.01 b | 0.14 ± 0.041 ab | 0.20 ± 0.007 a |
| 2,6-dimethylpyrazine | Roasted | 0.06 ± 0 c | 0.12 ±0.051 b | 0.21 ± 0.009 a |
| 2-ethyl-6-methylpyrazine | Nutty | 0.03 ± 0.007 b | 0.15 ± 0.066 a | 0.17 ± 0.004 a |
| 2-ethyl-5-methylpyrazine | Baked | 0.03 ± 0.017 b | 0.07 ± 0.005 a | 0.06 ± 0.009 a |
| 2-ethyl-3-methylpyrazine | Nutty, roasted, sweety | 0.08 ± 0.011 b | 0.12 ± 0.016 a | 0.11 ± 0.001 a |
| 3-ethyl-2,5-dimethylpyrazine | Baked, earthy, potato-like | 0.02 ± 0.001 b | 0.08 ± 0.020 a | 0.09 ± 0.001 a |
| Pyrazinamide | n.f. | 0.02 ± 0.003 b | 0.03 ± 0.014 ab | 0.04 ± 0.001 a |
| Total | 0.62 ± 0.01 b | 1.17 ± 0.03 a | 1.27 ± 0.07 a | |
| Pyrrolines | ||||
| 2-acetyl-1-pyrroline | Cracker-like | 0.02 ± 0.010 b | 0.09 ± 0.040 a | 0.10± 0.013 a |
| 3-hydroxy-2-methyl-4-pyrone (maltol) | Caramel, sweet | 0.04 ± 0.014 b | 0.22 ± 0.15 a | 0.32 ± 0.047 a |
| Total | 0.06 ± 0.01 b | 0.31 ± 0.08 a | 0.42 ± 0.03 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perri, G.; Rizzello, C.G.; Ampollini, M.; Celano, G.; Coda, R.; Gobbetti, M.; De Angelis, M.; Calasso, M. Bioprocessing of Barley and Lentil Grains to Obtain In Situ Synthesis of Exopolysaccharides and Composite Wheat Bread with Improved Texture and Health Properties. Foods 2021, 10, 1489. https://doi.org/10.3390/foods10071489
Perri G, Rizzello CG, Ampollini M, Celano G, Coda R, Gobbetti M, De Angelis M, Calasso M. Bioprocessing of Barley and Lentil Grains to Obtain In Situ Synthesis of Exopolysaccharides and Composite Wheat Bread with Improved Texture and Health Properties. Foods. 2021; 10(7):1489. https://doi.org/10.3390/foods10071489
Chicago/Turabian StylePerri, Giuseppe, Carlo Giuseppe Rizzello, Marco Ampollini, Giuseppe Celano, Rossana Coda, Marco Gobbetti, Maria De Angelis, and Maria Calasso. 2021. "Bioprocessing of Barley and Lentil Grains to Obtain In Situ Synthesis of Exopolysaccharides and Composite Wheat Bread with Improved Texture and Health Properties" Foods 10, no. 7: 1489. https://doi.org/10.3390/foods10071489
APA StylePerri, G., Rizzello, C. G., Ampollini, M., Celano, G., Coda, R., Gobbetti, M., De Angelis, M., & Calasso, M. (2021). Bioprocessing of Barley and Lentil Grains to Obtain In Situ Synthesis of Exopolysaccharides and Composite Wheat Bread with Improved Texture and Health Properties. Foods, 10(7), 1489. https://doi.org/10.3390/foods10071489