Abstract
The stability of soil organic carbon (SOC) in high-latitude permafrost regions plays a critical role in the global carbon balance. However, a systematic understanding of SOC pool fractions and their response to warming across different wetland types in the Great Hing’an Mountains remains lacking. In this study, soil samples were collected from forested, shrub, and herbaceous wetlands at depths of 0–60 cm and incubated at 5, 10 and 15 °C. A three-pool first-order kinetic model was employed to analyze SOC mineralization characteristics, carbon pool fractions, and influencing factors. The results showed that SOC mineralization rates exhibited a pattern of rapid increase followed by a peak and gradual decline over time, decreased with soil depth, and increased with temperature. The mineralization potential followed the order of shrub wetlands > herbaceous wetlands > forest wetlands. The temperature sensitivity (Q10) was lowest in the deep soil layer of shrub wetlands (1.2), whereas a deeper soil layer of forest wetlands exhibited the highest Q10 value (3.5). Across the three wetland types, SOC was dominated by the inert carbon pool (61–72%), with forest wetlands showing the highest proportion of inert carbon (72%). The active carbon pool in shrub wetlands was most sensitive to warming, while herbaceous wetlands had the largest inert carbon stock. Soil pH was significantly negatively correlated with the inert carbon pool, whereas soil moisture content showed a significantly positive correlation. Path analysis further revealed that SOC had the largest total effect on inert carbon accumulation, whereas available nitrogen and pH showed the strongest direct associations with Q10. Wetland type was indirectly associated with inert carbon stocks through its influence on soil moisture, pH, SOC, and available nitrogen. These results highlight that both direct and indirect pathways jointly influence SOC stability in permafrost wetlands. Overall, Wetland type and soil physicochemical properties jointly regulate SOC stability and its response to warming. These results suggest that although forest wetlands possess stronger carbon stability, their stable carbon pools may become increasingly vulnerable under climate warming.
1. Introduction
Permafrost soils store a substantial portion of the Earth’s soil organic carbon and therefore play a crucial role in the global carbon cycle and climate regulation [1,2]. The size and long-term stability of these carbon stocks have important implications for atmospheric CO2 concentrations and climate regulation, particularly under ongoing climate warming [3,4]. Wetlands, particularly permafrost peatlands, represent important carbon reservoirs where large amounts of organic carbon accumulate due to low temperatures and water-saturated conditions that limit microbial decomposition [5,6]. It has been estimated that approximately 2300 Pg of organic carbon is stored within the upper 3 m of soils globally [7], with more than one-third located in permafrost regions [8]. Although soil organic carbon in permafrost wetlands generally exhibits slow turnover rates under natural conditions, warming-induced changes in the balance between carbon inputs and mineralization outputs may gradually enhance CO2 release and contribute to climate feedback [9,10,11]. SOC stability, which reflects the resistance of organic carbon to microbial decomposition and environmental disturbance, therefore directly determines its long-term sequestration potential. Understanding the composition, stability, and environmental response mechanisms of SOC has thus become a central topic in global change research.
Due to the strong heterogeneity of SOC, its constituent fractions differ markedly in decomposition rates and turnover times. Multi-pool kinetic models have therefore been widely used to characterize SOC dynamics by partitioning soil carbon into pools with different degrees of stability [12,13]. Early SOC pool models were primarily developed for agricultural and temperate mineral soils, where decomposition processes are relatively well constrained. However, peatland and permafrost ecosystems differ substantially in hydrological conditions, oxygen availability, freeze–thaw processes, microbial communities, and organic matter composition, all of which strongly influence SOC turnover and stabilization mechanisms. In recent years, multi-pool approaches have increasingly been applied to peatland and permafrost ecosystems to evaluate carbon stability and mineralization processes under warming conditions [14,15,16]. These models commonly partition SOC into active, slow, and inert pools with distinct decomposition kinetics. Although two-pool models have been widely used to quantify SOC mineralization patterns and evaluate the effects of warming and soil depth [14,15,16], they are often insufficient to capture the multistage nature of SOC turnover [13]. Three-pool models may provide a more comprehensive representation of carbon dynamics and remain useful tools for quantitatively comparing SOC pool sizes and turnover characteristics across environmental gradients, despite simplifying the complexity of peatland biogeochemical processes. Previous studies have shown that land use, litter quality, clay content, and temperature changes can significantly influence the SOC pool distribution and decomposition dynamics [17,18,19,20,21]. Nevertheless, systematic assessments of SOC pool fractions, turnover time, and temperature sensitivity in high-latitude permafrost wetlands—particularly across different wetland types in the Great Hing’an Mountains—remain limited.
Temperature is widely recognized as a key factor regulating SOC decomposition in permafrost ecosystems. The temperature sensitivity coefficient (Q10) is widely used to quantify the response of SOC mineralization to warming and represents the proportional increase in decomposition rate associated with a 10 °C rise in temperature [22]. Previous studies suggest that SOC temperature sensitivity is closely related to substrate quality and carbon pool stability, with more stable carbon often exhibiting greater temperature sensitivity. However, the relationships between SOC pool stability and temperature sensitivity across different wetland types in the permafrost wetlands of the Great Hing’an Mountains remain poorly understood.
The permafrost region of the Great Hing’an Mountains represents the southernmost distribution area of high-latitude permafrost in Eurasia and constitutes the primary permafrost region in China [8]. This region contains extensive wetland ecosystems and stores substantial amounts of soil organic carbon, making it an important component of the regional and global carbon cycle. Due to its location at the southern boundary of the permafrost zone, the region is highly sensitive to climate warming and environmental changes, which may accelerate SOC mineralization and alter carbon cycling processes.
Therefore, typical wetland ecosystems in the Mohe region of the Great Hing’an Mountains were selected for this study. Through field sampling and laboratory incubation experiments combined with a three-pool first-order kinetic model, we quantified the size, turnover time, and stability of SOC pools under different wetland types, soil depths, and temperature conditions. In addition, the temperature sensitivity coefficient (Q10) was used to evaluate the response of SOC mineralization to warming. The objectives of this study were to: (1) characterize SOC mineralization dynamics under different wetland types and soil depths; (2) quantify the fractions and turnover characteristics of SOC pools using a three-pool kinetic model; (3) assess the temperature sensitivity of SOC mineralization and its relationships with environmental factors in permafrost wetlands in the Great Hing’an Mountains.
2. Materials and Methods
2.1. Study Area
Mohe is situated on the northern slope of the Great Hing’an Mountains, with geographic coordinates spanning from 122°00′ to 124°00′ E and 52°00′ to 54°00′ N (Figure 1). The area is marked by low mountains and undulating hills, with an average elevation of 500–600 m and a general topographic gradient sloping from south to north. The climate is classified as a temperate continental monsoon climate. Over the past two decades, the mean annual air temperature has been approximately −3.27 °C [23], and the annual precipitation ranges from 400 to 600 mm [24]. The frost-free period in this region lasts approximately 90 days, and it falls within the zone of discontinuous permafrost [25]. According to dominant vegetation structure, wetlands were classified into forest, shrub, and herbaceous types: forest wetlands are dominated by Larix gmelinii tree canopy; shrub wetlands by dwarf birch (Betula fruticosa) and willow (Salix spp.); and herbaceous wetlands by sedge (Carex spp.) and meadow marsh vegetation with little or no woody cover. All sampling sites were selected within the same permafrost zone under comparable environmental conditions: gentle slopes, seasonally waterlogged (though with vegetation-dependent moisture differences), and consistent active layer thickness and permafrost background, minimizing the influence of environmental heterogeneity on SOC dynamics [26].
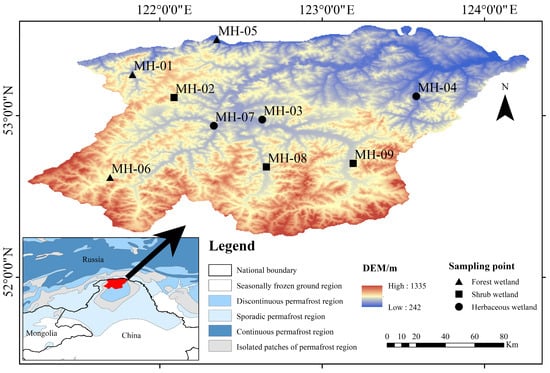
Figure 1.
Geographical location of soil sampling points and permafrost zonation in Mohe region Great Hing’an Mountains.
2.2. Sample Collection
In August 2024, organic peat soil samples were collected from three independent sampling sites for each of the three wetland types (forested, shrub, and herbaceous). At each site, one peat core was taken from depths of 0–20, 20–40 and 40–60 cm (no mixing of cores from different locations was performed) and then transported to the laboratory under refrigerated conditions. Visible plant residues, roots, and gravel were manually removed. One subsample was filtered through a 2 mm sieve and stored at 4 °C for laboratory incubation experiments; for each independent soil sample, three technical replicates were incubated, and the three technical replicates from the same sampling site were first averaged to obtain a site-specific value, so that all statistical analyses were based on the three field replicates (n = 3 per treatment). The other subsample was air-dried, ground, and sieved to a size of less than 2 mm for the analysis of soil physicochemical properties (Table S1 in Supplementary Materials).
2.3. Incubation Experiment
For each treatment, three fresh soil subsamples, each equivalent to 30 g of dry weight, after which samples were weighed and recorded [27], were incubated at three different temperature levels (5 °C, 10 °C and 15 °C), which represent the typical soil temperature range during the growing season in the study area and include a future warming scenario [28,29]. Each sampling site included three replicates, along with three empty bottles serving as blank controls. Before incubation, all soil samples were pre-incubated in the dark for three days to stabilize internal soil conditions and reduce disturbances from sample handling. Gas samples were collected on days 1, 2, 5, 10, 15, 21, 26, 32, 39, 46, 55, 63 and 72 of the incubation period to determine CO2 concentrations [30]. At the start of incubation, each bottle together with the soil was weighed to record the initial weight. Every three days, the bottles were reweighed, and deionized water was added to return the weight to the initial value, thereby maintaining soil moisture at a constant level throughout the experiment. Before each gas sampling event, the sealed incubation bottles were opened and placed in a ventilated area for 30 min, with air circulation facilitated by a fan to equilibrate gas concentrations inside and outside the bottles. Afterward, the three-way valve was closed, and the bottles were placed back into the temperature-controlled incubator. Following one hour of sealing, 20 mL of headspace gas was extracted using a syringe through the three-way valve and analyzed for CO2 concentration using a gas chromatograph (GC; Agilent 7890B, Santa Clara, CA, USA) [31]. The soil organic carbon mineralization rate represents the amount of organic carbon decomposed by microbial activity and released as CO2 per unit time. It is a dynamic indicator of SOC decomposition intensity and directly reflects the instantaneous carbon release capacity of soil carbon pools. The mineralization rate was calculated as follows [31]:
where F is the SOC mineralization rate (mg/kg/h), ρ is the CO2 concentration of the standard gas (g/L), ΔC is the difference in CO2 concentration between the sample and the blank control after 1 h of incubation (ppmv/h), V is the volume of the incubation bottle (mL), T is the incubation temperature (°C), and W is the dry weight of soil (kg).
The cumulative SOC mineralization represents the total amount of CO2 released from SOC decomposition over a given incubation period. It provides insight into the long-term carbon release potential of the soil carbon pool. Cumulative mineralization was calculated using the following equation [31]:
The temperature sensitivity of soil organic carbon mineralization, represented by the Q10 value, is calculated using an exponential model. Specifically, the method is as follows [32]:
In the equation, R denotes the CO2 emission rate (mg/kg/h); T represents the incubation temperature (°C); and α and β are model parameters.
2.4. Determination of SOC Pool Fractions
In this study, SOC was partitioned into active, slow, and inert pools based on kinetic modeling combined with acid hydrolysis measurements, rather than by physical density fractionation methods. Briefly, 2 mL of 13 mol/L H2SO4 was added to a 50 mL heat-resistant test tube containing soil, followed by shaking at room temperature for 6 h. Subsequently, 25 mL of deionized water was added to dilute the H2SO4 concentration to 1.0 mol/L. The tube was sealed and placed in an oven at 105 °C for hydrolysis for 3 h, during which heating was paused for 15 min every hour. After cooling, the samples were centrifuged at 3750 r/min for 15 min, and the supernatant was discarded. The remaining residue was rinsed with 5–10 mL of deionized water and centrifuged three times (the final rinse solution was retained). The soil residue and rinse solution were transferred to a flat-bottom funnel lined with filter paper and vacuum-filtered. The filtered soil samples were then oven-dried at 60 °C, and the inert carbon content was finally determined using a total organic carbon (TOC) analyzer [33].
Based on soil respiration incubation data combined with conventional measurements, a three-pool first-order kinetic model was applied to fit the size and turnover time of the active and slow SOC pools [30]. The model was fitted separately for each wetland type, soil depth, and temperature treatment, allowing model parameters to vary among soils:
where Csoct is the SOC content at time t; Ca and Ka represent the size and decomposition rate of the active carbon pool, respectively; Cs and Ks represent the size and decomposition rate of the slow carbon pool; and Cr and Kr represent the size and decomposition rate of the inert carbon pool. The decomposition rate constants Ka, Ks and Kr are inversely related to the mean residence time (MRT) of the corresponding carbon pools [30].
Because differences between laboratory incubation temperatures and field conditions affect SOC turnover time, laboratory-derived MRT values were converted to field-equivalent MRT values as follows [30]:
where MRT is the mean residence time, Q10 is the temperature sensitivity coefficient, and MAT is the mean annual temperature of the study area.
The decomposition rate of the inert carbon pool was calculated as [30]:
Assuming a turnover time of 1000 years for the inert carbon pool, the assumption of a 1000-year turnover time for the inert carbon pool has been widely adopted in previous SOC fractionation and kinetic modeling studies of recalcitrant carbon in wetland and permafrost soils [30,34], Kr was derived accordingly. With Csoct and Cr obtained from incubation experiments, the size of the slow carbon pool was calculated as Cs = Csoct − Cr − Ca. Substituting this relationship into the three-pool model yields:
Nevertheless, applying a three-pool model to a 72-day incubation introduces uncertainties, particularly for the slow and inert pools whose turnover times greatly exceed the experimental period. The estimated sizes and turnover times of these pools should therefore be interpreted as relative approximations rather than absolute values. Despite this limitation, the model remains useful for comparing SOC stability among different wetland types and temperature treatments.
In this study, SOC stability was evaluated based on three complementary criteria. First, SOC was partitioned into active, slow, and inert carbon pools; a higher proportion of inert pools indicates longer turnover time and greater stability. Second, mean residence times derived from the three-pool kinetic model were used to assess stability, with longer MRTs corresponding to slower decomposition rates and higher stability. Third, temporal patterns of SOC mineralization during laboratory incubation were examined; slower mineralization rates and later occurrence of peak respiration were interpreted as indicators of higher SOC stability [30].
2.5. Statistical Analysis
The three-pool first-order kinetic model was fitted using the Levenberg–Marquardt nonlinear optimization algorithm implemented in SAS 9.4, yielding estimates of Ca, Cs, Ka and Ks. Three-way analysis of variance (ANOVA) with second-order interactions and Fisher’s LSD post hoc test was used to examine the main and interactive effects of wetland type, soil depth, and temperature on peak mineralization rate and cumulative mineralization. Pearson correlation analysis was conducted to explore the relationships between SOC fractions and key environmental factors. All analyses were conducted separately for each incubation temperature (5 °C, 10 °C and 15 °C). Statistical significance was defined at p < 0.05 (significant) and p < 0.01 (highly significant). Correlation matrices were generated using SPSS 23.0, and heat maps were produced with Origin 2024. To systematically identify the key factors influencing the inert carbon pool and to disentangle their direct and indirect effects, Partial least squares path modeling (PLS-PM) was performed using RStudio 2024 with the plspm package. In the PLS-PM analysis, direct effects refer to standardized path coefficients linking environmental variables to the inert carbon pool, whereas indirect effects represent mediated influences through intermediate variables. Path diagrams and graphical processing were completed using Origin 2024.
3. Results
3.1. Characteristics of Soil Organic Carbon Mineralization
The mineralization rate exhibits a distinct pattern, initially rising rapidly and then gradually declining during the incubation period, reaching its peak between days 10 and 20 (Figure 2). Under temperatures of 5 °C, 10 °C and 15 °C, the mineralization rates across different wetland types follow the order of shrub wetland > herbaceous wetland > forest wetland. Shrub wetlands exhibited significantly higher mineralization rates than forest wetlands (p < 0.05). Furthermore, with increasing soil depth (from 0–20 cm to 40–60 cm), the peak mineralization rates of all wetland types decreased gradually, and the 0–20 cm layer showed greater mineralization potential than the deeper layers (p < 0.05).
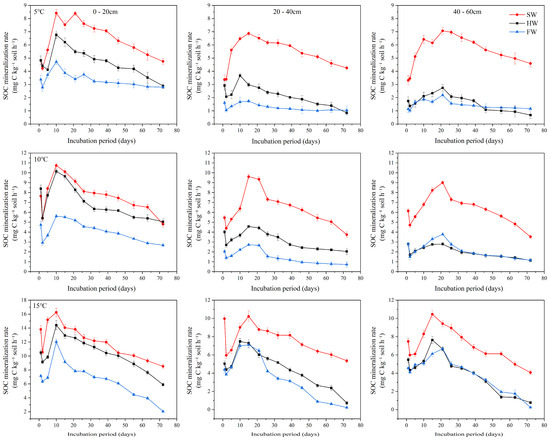
Figure 2.
Soil organic carbon mineralization rates at different soil depths (0–20 cm, 20–40 cm, 40–60 cm) in forested, shrub, and herbaceous wetlands under three temperature treatments (5 °C, 10 °C, 15 °C).
Throughout the 72-day incubation period, the cumulative SOC mineralization consistently increased (Figure 3). Under varying temperature conditions, the cumulative mineralization in shrub wetlands was always higher than in herbaceous wetlands and forest wetlands. At 5 °C, the greatest difference in cumulative mineralization was observed in the 40–60 cm soil layer, where the cumulative mineralization in the shrub wetland was 3.6 times and 3.9 times higher than in the herbaceous wetland and forest wetland, respectively. Under the same temperature conditions, the average cumulative mineralization in the upper soil layer (0–20 cm) was approximately 1.5 to 1.8 times higher than in the middle soil layer (20–40 cm), and 1.8 to 2.5 times higher than in the lower soil layer (40–60 cm). Three-way ANOVA revealed that wetland type, soil depth, and temperature each had significant main effects on both peak mineralization rate and cumulative mineralization (p < 0.01). The interaction between temperature and soil depth was also significant (p < 0.05).
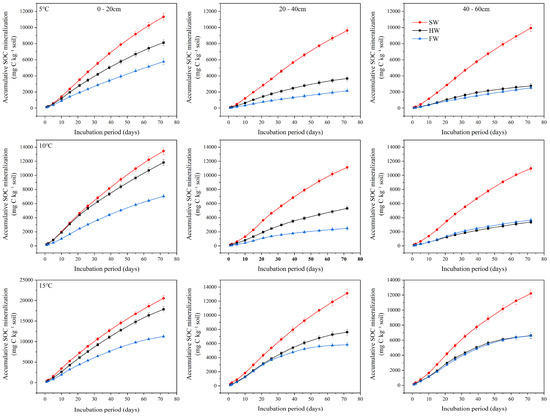
Figure 3.
Cumulative soil organic carbon mineralization over 72 days at different soil depths (0–20 cm, 20–40 cm, 40–60 cm) in forested, shrub, and herbaceous wetlands under 5 °C, 10 °C and 15 °C temperature treatments.
The fitted Q10 values ranged from 1.222 ± 0.007 to 3.493 ± 0.414 (Table 1). Apparent differences in Q10 were observed across vegetation types and soil depths. In shrub wetland soils, Q10 decreased with increasing soil depth, with the value in the 0–20 cm layer being 1.897 ± 0.274, and dropping to 1.222 ± 0.007 in the 40–60 cm layer. In herbaceous and forest wetland soils, Q10 reached its highest values in the 40–60 cm layer, which were higher than those in the 0–20 cm and 20–40 cm layers. Overall, both vegetation type and soil depth were associated with variations in the temperature sensitivity of soil organic carbon mineralization, with the deepest layer in forest wetland soils showing the highest Q10 values.

Table 1.
Temperature sensitivity of soil organic carbon mineralization and fitting parameters for different soil layers in forest wetlands, shrub wetlands, and herbaceous wetlands.
3.2. Soil Organic Carbon Pool Fractions and Turnover Time
The inert carbon pool constitutes the major component of soil organic carbon across all three wetland types, with contents ranging from 64.73 g/kg to 193.68 g/kg (Table 2). Among the different wetland types, forest wetlands exhibited the highest proportion of inert carbon, indicating relatively greater SOC stability (Figure 4). In contrast, herbaceous wetlands contained the highest absolute inert carbon content (193.68 g/kg), reflecting a larger stabilized carbon reservoir rather than necessarily a higher relative stability. As the temperature increased, the active carbon pool content in shrub and herbaceous wetlands showed an upward trend, while the change in forest wetlands was minimal.
Table 2.
Sizes and mean residence times of active, slow, and inert soil organic carbon pools at various soil depths (0–20 cm, 20–40 cm, 40–60 cm) in forested, shrub, and herbaceous wetlands under temperature treatments of 5 °C, 10 °C and 15 °C.
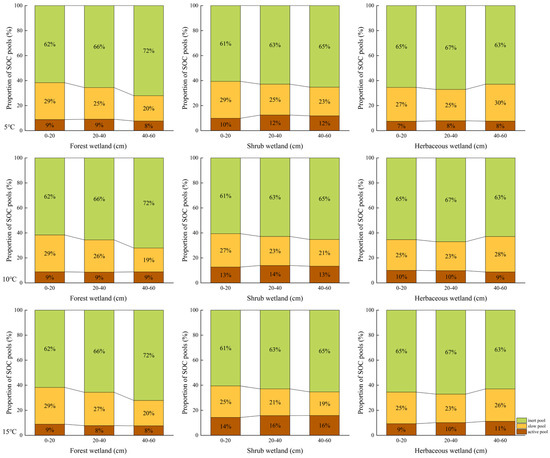
Figure 4.
Proportional contributions of soil organic carbon fractions (active, slow, inert) at different soil depths (0–20 cm, 20–40 cm, 40–60 cm) in forested, shrub, and herbaceous wetlands under 5 °C, 10 °C and 15 °C temperature treatments. Note: These values do not add up to exactly 100% due to rounding off by the plotting software, and the original data is ac-curate.
The turnover time of the active carbon pool follows a distinct vertical pattern: under the same wetland type and temperature conditions, the active carbon pool increases with soil depth, indicating slower decomposition of active carbon in deeper soils. Temperature elevation altered the carbon pool turnover times; with rising temperature, the turnover times of both the active and slow carbon pools showed a decreasing trend, which is mechanistically consistent with the observed increase in SOC mineralization rates. However, the extent of this temperature effect varied depending on wetland type and soil layer.
3.3. Correlation Between Soil Organic Carbon Fractions and Soil Physicochemical Properties
To identify the factors influencing soil organic carbon components in permafrost wetlands, a correlation analysis was conducted between organic carbon components and soil physicochemical properties at different temperatures (5 °C, 10 °C and 15 °C) (Figure 5). The results indicated that temperature significantly influenced these relationships. At all temperature treatments, soil pH exhibited a highly significant negative correlation with the inert carbon pool (p < 0.01). Soil moisture content showed a highly significant positive correlation with both the inert and slow carbon pools (p < 0.01). Bulk density did not show a significant correlation with any carbon component at 5 °C, but at 10 °C, it was significantly positively correlated with the decomposition rate of the slow carbon pool (p < 0.05), and the correlation was even stronger at 15 °C (p < 0.05). Additionally, the negative correlation between the active carbon pool and pH was significant only at 15 °C (p < 0.05).
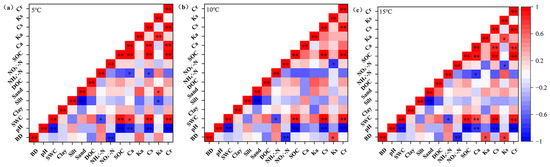
Figure 5.
Correlation analysis between soil organic carbon fractions and soil physicochemical factors at (a) 5 °C, (b) 10 °C and (c) 15 °C temperature treatments. Note: * indicates p < 0.05, statistically significant; ** indicates p < 0.01, highly statistically significant.
The PLS path model results (Figure 6a) revealed significant relationships among soil physicochemical properties, SOC fractions, and Cr accumulation. The direction and strength of the path coefficients indicated that SOC was strongly associated with Cr accumulation and may play an important role in regulating Cr dynamics. While carbon fractions and pH had no significant direct association with Cr, pH was indirectly associated with Cr through its relationship with SOC (Figure 6a). Available nitrogen and pH were the strongest direct predictors of Q10. The direct association of SOC on Q10 is not significant, whereas both available nitrogen and pH showed relatively strong direct associations with Q10. Wetland type was indirectly associated with both Cr and Q10 through its relationships with SWC, pH, available nitrogen, carbon fractions, and SOC. SWC showed a significant positive association with carbon fractions and SOC, and was indirectly associated with both Cr and Q10, positioning it as a key intermediate factor. In summary, in terms of total effect, SOC contributed the most to Cr, whereas available nitrogen and pH showed the greatest direct associations with Q10 (Figure 6b,c).
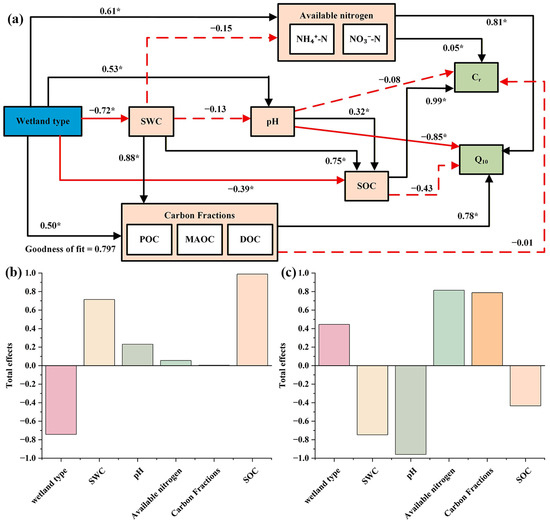
Figure 6.
(a) Path relationships between wetland soil physicochemical factors and the inert carbon pool and Q10, indicating positive (black) and negative (red) effects, with non-significant relationships represented by dashed lines, * indicates p < 0.05, statistically significant. (b) Total effects of various factors on Cr. (c) Total effects of various factors on Q10.
4. Discussion
4.1. Temperature Responses of Soil Organic Carbon Decomposition Rates in Different Wetland Types
The mineralization of soil organic carbon in typical wetlands of the Great Hing’an Mountains during incubation exhibited a characteristic dynamic pattern of initially increasing rapidly, then reaching a peak, and finally declining gradually, which is consistent with the theoretical framework of the three-pool model. During the initial stage of incubation, microorganisms preferentially utilize the readily decomposable components in the active carbon pool, such as soluble organic carbon, sugars, and amino acids, resulting in a rapid increase in the mineralization rate to a peak level [12,18]. As the active carbon pool is progressively depleted, the mineralization process gradually shifts toward dependence on the slow carbon pool, which possesses more complex chemical structures and slower turnover rates, leading to a decline in the mineralization rate that eventually stabilizes [12,18]. This process indicates that the chemical fractions of different carbon pools play a crucial role in regulating the short-term dynamics of SOC mineralization.
Increasing temperature significantly promoted the decomposition of SOC in different wetland soils. However, the magnitude of this response varied markedly among wetland types and soil depths. Overall, the stimulatory effect of warming on SOC decomposition followed the order of shrub wetland (SW) > herbaceous wetland (HW) > forest wetland (FW). Throughout the entire incubation period, the peak mineralization rate in the 0–20 cm soil layer of shrub wetlands increased most markedly, whereas the carbon release flux in forest wetlands remained consistently low, indicating a relatively weaker intensity of SOC decomposition. This difference may be associated with the quality of litter inputs [35]. Litter in shrub wetlands generally exhibits a lower C/N ratio and lower lignin content, which may make it more readily utilized by microorganisms, potentially explaining the higher mineralization rates are observed under warming conditions [35,36]. In contrast, coniferous litter in forest wetlands contains higher proportions of recalcitrant components such as waxes and phenolic compounds, which could constrain microbial decomposition activity and thus might reduce the intensity of SOC mineralization [35,36]. This pattern is consistent with observations in mountain peatlands, where sites with greater shrub (e.g., willow) cover showed distinct microbial community structures and higher decomposition potentials compared to those dominated by other vegetation types [37]. Vertically, the stimulatory effect of warming on SOC decomposition was markedly stronger in surface soils (0–20 cm) than in deeper soils (40–60 cm). This pattern may be attributed to the relatively abundant active carbon pool in surface soils, which also continuously receives fresh litter inputs [35,38]. In contrast, organic carbon in deeper soils is likely to be more stabilized (e.g., mineral-associated carbon or carbon protected within soil aggregates), which may result in a weaker response to temperature changes [35,38]. Wang et al. further demonstrated that this depth effect is so pronounced that it can override the influence of local stratigraphy on microbial community composition [37]. Overall, this pattern of a strong surface response and a weak deep response highlights significant spatial heterogeneity in how temperature influences soil carbon decomposition.
The results indicate that the temperature sensitivity of soil organic carbon (SOC) mineralization is higher in the middle and deep layers of forest wetlands, but generally lower in shrub wetlands. At first glance, this result appears to contradict the relatively low mineralization rates observed in forest wetlands; however, it actually reveals the multidimensional nature of temperature sensitivity [38,39]. The Q10 index reflects relative changes rather than the absolute amount of mineralization [22]. Although the deeper carbon pools in forest wetlands decompose slowly under low temperatures, their relatively stable chemical structures may contribute to stronger relative temperature responses [40]. It should be noted that the stability of carbon pools does not necessarily correspond to lower temperature sensitivity [38]. According to the carbon quality–temperature hypothesis, organic carbon with more complex structures and higher stability generally requires higher activation energy for decomposition, and therefore exhibits stronger intrinsic sensitivity to temperature changes [22]. Carlos A. Sierra et al. [41] further pointed out that under low-temperature (<0 °C) and extreme moisture conditions, although decomposition rates remain low, temperature sensitivity may instead be relatively high [41], which is consistent with the observations obtained in the deeper layers of forest wetlands (characterized by low temperature and high moisture) in this study.
This study also found that both the peak mineralization rate and cumulative mineralization at 15 °C were higher than those at 5 °C and 10 °C. This indicates that rising temperature to some extent alleviates the constraints of low temperature on microbial metabolic activity in soils, thereby accelerating the turnover of SOC [42,43]. It is noteworthy that warming not only promoted the consumption of the active carbon pool (Ca), but the increase in cumulative mineralization also suggests that a portion of the slow carbon pool (Cs), previously protected by physical or chemical mechanisms, may have been stimulated to participate in decomposition. This finding echoes the phenomenon of warming-induced activation of inert carbon observed by Jiang et al. [42] in northern peatlands, suggesting that under future climate warming scenarios, if environmental constraints (e.g., physical protection and moisture conditions) are partially alleviated by rising temperatures, the potential contribution of these stable pools to carbon release under warming conditions should not be overlooked. Furthermore, the stimulatory effect of warming on organic carbon mineralization observed in this study may be amplified over longer timescales [43]. In addition, it is possible that soil microbial communities in shrub wetlands are better adapted to readily decomposable substrates, which could lead to a stronger temperature response and greater carbon release [36]. Moreover, although litter in herbaceous wetlands is relatively easy to decompose, their relatively high inert carbon content (180.56–193.68 g·kg−1) may to some extent slow the overall decomposition process [44]. Therefore, vegetation type may jointly influence the intensity of the response of SOC decomposition to temperature changes in different wetland ecosystems by influencing both litter quality and microbial community structure.
4.2. Fractions of Soil Organic Carbon Pools and Factors Controlling Their Stability Across Wetland Types
Analysis of the soil organic carbon pool fractions revealed that the inert carbon pool dominated all three wetland types, comprising 61% to 72% of the total SOC. This finding aligns closely with the results of Zuo et al. [34], whose model estimates indicated that the inert carbon pool represents the majority of SOC (57% to 67%). Despite this overall similarity, clear differences in carbon pool characteristics were observed among wetland types: forest wetlands were characterized by a high proportional contribution of inert carbon, suggesting relatively greater SOC stability, in contrast, herbaceous wetlands possessed the largest absolute inert carbon stocks, indicating stronger carbon storage capacity, whereas shrub wetlands exhibited the highest carbon pool activity. Although forest wetlands had the lowest total SOC content, they exhibited the highest proportion of inert carbon (up to 72%). This pattern may be likely attributable to the slow humification of coniferous litter, which can favor the formation of highly condensed aromatic structures (e.g., humic substances and black carbon). Chen et al. [43] reported that during the late stages of litter decomposition, soil-derived microorganisms—particularly fungal taxa capable of degrading complex organic compounds—could be found. This mechanism aligns well with the high inert carbon pool proportions observed in forest wetlands in the present study, suggesting that the recalcitrant nature of coniferous litter not only slows early-stage decomposition but also promotes the formation of stable carbon pools during later stages of humification [45,46]. In contrast, shrub wetlands exhibited higher carbon pool activity and lower stability. This suggests that shrub wetlands may exhibit relatively greater carbon-release potential under climate warming scenarios. Supporting this interpretation, Jiang et al. [42] showed that glucose addition initially stimulated SOC mineralization (positive priming) but later suppressed it (negative priming), implying that inputs of labile carbon can alter carbon stability by regulating microbial community composition (e.g., fungal-to-bacterial ratios). This finding is consistent with the relatively high carbon pool activity and lower stability observed in shrub wetlands in the present study. Herbaceous wetlands, by contrast, exhibited the highest absolute inert carbon stocks and total SOC contents, reflecting strong carbon sequestration and storage capacity. However, this does not necessarily imply the highest relative SOC stability because the proportional contribution of inert carbon was lower than that in forest wetlands. Dense fibrous root systems provide continuous organic matter inputs that contribute to SOC accumulation and aggregate formation [18,35,47]. In addition, persistently waterlogged conditions in wetland soils suppress aerobic microbial decomposition, thereby favoring long-term carbon preservation. Together, these factors promote SOC accumulation and its progressive transformation into inert carbon forms.
Soil physicochemical factors appear to act synergistically in their associations with the carbon stability patterns described above, and these associations exhibit pronounced temperature dependence [46,48]. Correlation analyses suggest that soil acidity and moisture are two key environmental factors linked to SOC stability [49]. Soil pH showed an extremely significant negative correlation with the inert carbon pool (p < 0.01), and this relationship remained stable across all temperature treatments. The underlying mechanisms likely involve several pathways: under acidic conditions, the protonation of hydroxylated surfaces on Fe and Al oxides enhances the positive surface charge, thereby enhancing the specific adsorption of negatively charged aromatic organic compounds; meanwhile, acidic environments generally suppress extracellular enzyme activity, slowing organic matter decomposition [50]. Carbon stability in different ecosystems is subject to threshold effects; for example, excessive drying in peatlands can disrupt the positive role of woody vegetation in carbon storage [51]. Soil water content (SWC) showed a significant positive correlation with both the inert and slow carbon pools, underscoring the important role of moisture in maintaining carbon stability in permafrost regions by limiting aerobic microbial activity and influencing substrate transport and availability [52]. High SWC may suppress aerobic microbial decomposition by creating anaerobic conditions, and may also enhance the formation and persistence of soil aggregates, thereby physically protecting organic carbon [53,54]. Permafrost itself acts as a stable reservoir of water and ice, and its degradation will directly weaken these protective effects. Notably, the associations involving soil physical properties become increasingly important with rising temperature, suggesting a temperature dependence in how soil texture relates to carbon decomposition. In this work, bulk density was found to be positively associated with the decomposition rate of the slow carbon fraction at 10 °C, with this association becoming more pronounced at 15 °C. This trend indicates that, under relatively low thermal conditions, constraints on carbon breakdown are dominated by limitations in microbial activity rather than by substrate availability or physical protection mechanisms. Once warming activates microbial metabolism, soil physical structure—such as compaction-induced constraints on gas diffusion—becomes a dominant control on carbon turnover rates [55]. Consistent with this interpretation, the relationship between sand content and the decomposition rate of the slow carbon pool also shifted with increasing temperature, further illustrating the complex interactions among temperature, moisture, and texture. Under warming conditions, accelerated evaporation may exacerbate the poor water-holding capacity of sandy soils, causing moisture limitation to override aeration as the primary constraint on decomposition [56]. Together, these results demonstrate that the influence of soil texture on carbon stability does not operate in isolation, but is realized through its interaction with other environmental factors—particularly temperature and water availability [23,57].
This study further employed structural equation modeling (Figure 6) to explore the systematic associations underlying inert carbon pool stability and the temperature sensitivity (Q10) of soil respiration. Wetland type, as an upper-level driver, was indirectly associated with the accumulation efficiency of inert carbon and the response pattern of Q10 through its relationships with SWC, pH, available nitrogen, and carbon fractions [43]. Notably, although pH has no direct effect on inert carbon, it showed an indirect positive association with inert carbon through SOC [50]. The significant positive effect of SWC on carbon fractions and SOC suggests that moisture conditions may influence the availability and transformation processes of carbon substrates, which in turn may relate to both the binding sites for inert carbon and the temperature sensitivity of soil respiration [53,54]. More critically, Q10 was most directly associated with available nitrogen and pH rather than with SOC or carbon fractions, suggesting that nitrogen availability and pH may be more directly linked to the temperature sensitivity of organic carbon decomposition in permafrost wetlands, and that the total carbon pool size alone does not fully determine its temperature sensitivity. This path model integrates previously scattered correlations and suggests that inert carbon accumulation and Q10 are associated with a hierarchical network: wetland type corresponds to background conditions of hydrothermal, chemistry, and carbon substrates, while SOC and available nitrogen serve as the most direct contributing factors most directly associated with inert carbon and Q10, respectively, and the path model further reveals that direct, indirect, and total effects jointly operate in this hierarchical network. Future climate change, if it alters vegetation community structure or leads to permafrost degradation (with associated changes in hydrothermal regimes and pH), will profoundly affect the stability of the regional recalcitrant carbon pool and the soil carbon–climate feedback through this synergistic network [58].
Overall, carbon stability in permafrost wetlands appears to reflect the interactive influences of vegetation-derived inputs and the soil physicochemical environment, with temperature modulating the relative importance of individual environmental factors within this interaction. Herbaceous wetlands maintain carbon stability primarily through the combined effects of high soil moisture and acidic conditions. At the same time, our results suggest a temperature dependence in how soil texture relates to carbon decomposition, as the associations of bulk density and sand content with carbon decomposition varied with thermal conditions [34,59]. Collectively, these findings indicate that carbon stability in permafrost regions is associated with the coupled interactions among moisture, acidity, soil texture, and temperature. Accordingly, accurate predictions of carbon-cycle feedback under future climate warming must explicitly account for the interactions among these key environmental drivers [60].
5. Conclusions
This study investigated SOC mineralization in three typical marsh wetlands in the permafrost region of the Great Hing’an Mountains. SOC mineralization rates increased with temperature and decreased with soil depth. Shrub marshes, with a higher proportion of active carbon, showed the highest mineralization potential and rapid warming response. Forest marshes had the highest proportion of inert carbon (72%), indicating strong carbon stability, but their deeper stable carbon pools exhibited higher temperature sensitivity (Q10), posing a risk of activation. Soil pH (negative) and moisture (positive) were key factors significantly associated with inert carbon accumulation. Path analysis indicated that pH and soil water content influenced inert carbon accumulation indirectly, mediated by SOC. Under future warming, shrub marshes may act as rapid carbon sources, whereas forest marshes face potential risks from activation of stable carbon pools in deeper soils.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/su18115260/s1, Table S1: Soil physicochemical properties across wetland types and depths.
Author Contributions
Conceptualization, X.D.; Formal analysis, H.L.; Investigation, H.L., X.D., D.Y., H.M., R.Z., J.L. and F.Q.; Writing—original draft, H.L.; Writing—review and editing, X.D., M.L. and H.M.; Supervision, M.L.; Project administration, M.L.; Funding acquisition, X.D. and M.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was financially supported by Heilongjiang Provincial Natural Science Foundation of China—Distinguished Young Scholars Program (JQ2025D003); National Natural Science Foundation of China (42371119, 42501119); China Postdoctoral Science Foundation (2025M770323) and Natural Science Foundation of Heilongjiang Province (PL2024D006).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article and Supplementary Materials. Further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Man, H.R.; Dong, X.F.; Liu, C.; Wu, X.D.; Li, M.; Zheng, Z.C.; Jiang, Q.; Zang, S.Y. Spatial patterns and key drivers of soil organic carbon in northeastern China’s discontinuous permafrost zone. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2025, 18, 6738–6745. [Google Scholar] [CrossRef]
- Guo, Y.X.; Yu, G.H.; Hu, S.J.; Liang, C.; Kappler, A.; Jorgenson, M.T.; Guo, L.D.; Guggenberger, G. Deciphering the Intricate Control of Minerals on Deep Soil Carbon Stability and Persistence in Alaskan Permafrost. Glob. Change Biol. 2024, 30, e17552. [Google Scholar] [CrossRef]
- Mishra, U.; Hugelius, G.; Shelef, E.; Yang, Y.H.; Strauss, J.; Lupachev, A.; Harden, J.W.; Jastrow, J.D.; Ping, C.L.; Riley, W.J.; et al. Spatial heterogeneity and environmental predictors of permafrost region soil organic carbon stocks. Sci. Adv. 2021, 7, eaaz5236. [Google Scholar] [CrossRef]
- Guo, Z.Y.; Wang, Y.H.; Wan, Z.M.; Zuo, Y.J.; He, L.Y.; Li, D.; Yuan, F.H.; Wang, N.N.; Liu, J.Z.; Song, Y.Y.; et al. Soil dissolved organic carbon in terrestrial ecosystems: Global budget, spatial distribution and controls. Glob. Ecol. Biogeogr. 2020, 29, 2159–2175. [Google Scholar] [CrossRef]
- Song, Y.Y.; Wang, L.L.; Ma, X.Y.; Shi, F.X.; Wang, X.W.; Ren, J.S.; Jiang, L.; Hou, A.X.; Song, C.C. Effects of plant community diversity on soil microbial functional groups in permafrost peatlands of Greater hing’an Mountains, Northeast China. Wetl. Ecol. Manag. 2022, 30, 595–606. [Google Scholar] [CrossRef]
- Xue, J.; Zhang, Y.F.; Wang, S.J.; Du, Y.; Wang, D.X.; Zhang, H.; Song, Y.Y.; Wang, X.W.; Sun, X.X. Plant functional type control on soil microbial activity and carbon source utilization in permafrost peatland. Plant Soil 2025, 514, 807–821. [Google Scholar] [CrossRef]
- Jackson, R.B.; Lajtha, K.; Crow, S.E.; Hugelius, G.; Kramer, M.G.; Pineiro, G. The ecology of soil carbon: Pools, vulnerabilities, and biotic and abiotic controls. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 419–445. [Google Scholar] [CrossRef]
- Wu, H.B.; Zhang, Z.S.; Xue, Z.S.; Zhao, W.W.; Sang, L.; Wu, H.T.; Wang, W.F.; Guan, Q.; Lu, K.L. Predicting the potential geographical distribution of peatlands in Northeast China based on the ensemble model. Glob. Planet. Change 2025, 252, 104866. [Google Scholar] [CrossRef]
- Feng, Y.S.; Song, Y.Y.; Zhu, M.Y.; Li, M.T.; Gong, C.; Luo, S.Y.; Mei, W.K.; Feng, H.H.; Tan, W.W.; Song, C.C. Microbes drive more carbon dioxide and nitrous oxide emissions from wetland under long-term nitrogen enrichment. Water Res. 2025, 272, 122942. [Google Scholar] [CrossRef]
- Si, Q.; Chen, K.L.; Wei, B.; Zhang, Y.W.; Sun, X.; Liang, J.Y. Dissolved carbon flow to particulate organic carbon enhances soil carbon sequestration. Soil 2024, 10, 441–450. [Google Scholar] [CrossRef]
- Liu, Z.D.; Song, Y.Y.; Ma, X.Y.; Yuan, J.B.; Lou, Y.J.; Yang, C.; Tang, H.R.; Song, C.C. Deep soil microbial carbon metabolic function is important but often neglected: A study on the Songnen Plain reed wetland, Northeast China. Fundam. Res. 2022, 3, 833–843. [Google Scholar] [CrossRef]
- Townsend, A.R.; Vitousek, P.M.; Trumbore, S.E. Soil organic matter dynamics along gradients in temperature and land use on the island of Hawaii. Ecology 1995, 76, 721–733. [Google Scholar] [CrossRef]
- Zhu, L.Y.; Pan, J.J.; Zhang, W. Study on soil organic carbon pools and turnover characteristics along an elevation gradient in qilian mountain. Environ. Sci. 2013, 34, 668–675. (In Chinese) [Google Scholar] [CrossRef]
- Hatten, J.A.; Zabowski, D. Changes in soil organic matter pools and carbon mineralization as influenced by fire severity. Soil Sci. Soc. Am. J. 2009, 73, 262–273. [Google Scholar] [CrossRef]
- Rasmussen, C.; Southard, R.J.; Horwath, W.R. Mineral control of organic carbon mineralization in a range of temperate conifer forest soils. Glob. Change Biol. 2006, 12, 834–847. [Google Scholar] [CrossRef]
- Rey, A.; Pegoraro, E.; Jarvis, P.G. Carbon mineralization rates at different soil depths across a network of European forest sites (FORCAST). Eur. J. Soil Sci. 2008, 59, 1049–1062. [Google Scholar] [CrossRef]
- Collins, H.P.; Blevins, R.L.; Bundy, L.G.; Christenson, D.R.; Dick, W.A.; Huggins, D.R.; Paul, E.A. Soil carbon dynamics in corn-based agroecosystems: Results from carbon-13 natural abundance. Soil Sci. Soc. Am. Proc. 1999, 63, 584–591. [Google Scholar] [CrossRef]
- Wang, X.Y.; Li, L.; Xin, Z.J.; Li, X.H.; He, S.F.; Sun, X.Y. Spatial Variations in Organic Carbon Pools and Their Responses to Different Annual Straw Return Rates in Surface Paddy Soils in South China. Sustainability 2022, 14, 16875. [Google Scholar] [CrossRef]
- Ren, Z.B.; Li, C.J.; Wang, S.; Zhou, W.X.; Stringer, L.C.; Fu, B.J. Effects of land-use change on soil total carbon pool: A meta-analysis. Agric. Ecosyst. Environ. 2026, 396, 110021. [Google Scholar] [CrossRef]
- Schiedung, M.; Barré, P.; Peoplau, C. Separating fast from slow cycling soil organic carbon–A multi-method comparison on land use change sites. Geoderma 2025, 453, 117154. [Google Scholar] [CrossRef]
- Willard, S.J.; Liang, G.P.; Adkins, S.; Foley, K.; Murray, J.; Waring, B. Land use drives the distribution of free, physically protected, and chemically protected soil organic carbon storage at a global scale. Glob. Change Biol. 2024, 30, e17507. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 2006, 440, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Dong, L.; Zhang, H.R.; Zheng, C.H.; Zhou, J.X.; Peng, X.W. Microbial effect on soil organic carbon accumulation and stabilization is lithology-depend in subtropical coniferous forest. Appl. Soil Ecol. 2025, 210, 106103. [Google Scholar] [CrossRef]
- Yan, X.; Li, M.; Guo, D.F.; Yang, D.Y.; Zhan, D.Q. Spatial-temporal evolution and prediction of carbon storage in Mohe city by linking the logistic-CA-Markov and InVEST models. Front. Earth Sci. 2024, 12, 1383237. [Google Scholar] [CrossRef]
- Obu, J.; Westermann, S.; Bartsch, A.; Berdnikov, N.; Christiansen, H.H.; Dashtseren, A.; Delaloye, R.; Elberling, B.; Etzelmüller, B.; Kholodov, A.; et al. Northern Hemisphere permafrost map based on TTOP modelling for 2000–2016 at 1 km2 scale. Earth-Sci. Rev. 2019, 193, 299–316. [Google Scholar] [CrossRef]
- Liu, L.; Wang, Z.L.; Ma, D.L.; Zhang, M.; Fu, L.Y. Diversity and distribution characteristics of soil microbes across forest–peatland ecotones in the permafrost regions. Int. J. Environ. Res. Public Health 2022, 19, 14782. [Google Scholar] [CrossRef]
- Gao, F.; Jiang, H.; Cui, X.Y. Soil organic carbon pools and their turnover under two different types of forest in Xiao-xing’an Mountains, Northeast China. Chin. J. Appl. Ecol. 2015, 26, 1913–1920. (In Chinese) [Google Scholar] [CrossRef]
- Pei, J.M.; Fang, C.M.; Li, B.; Nie, M.; Li, J.Q. Direct evidence for microbial regulation of the temperature sensitivity of soil carbon decomposition. Glob. Change Biol. 2024, 30, e17523. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.J.; Zhou, Z.H.; Delgado-Baquerizo, M.; Bastida, F.; Zhao, F.Z.; Yang, Y.H.; Zhang, S.H.; Wang, J.Y.; Zhang, C.; Han, X.H.; et al. Thermal sensitivity of soil microbial carbon use efficiency across forest biomes. Nat. Commun. 2024, 15, 6269. [Google Scholar] [CrossRef]
- Dou, Y.X. Characteristics of Soil Organic Carbon Pool, Stability and Influencing Factors under Different Small Watersheds on the Loess Hilly Region, China. Ph.D. Thesis, University of Chinese Academy of Sciences, Xianyang, China, June 2020. (In Chinese) [Google Scholar] [CrossRef]
- Dong, X.F.; Liu, C.; Wu, X.D.; Man, H.R.; Wu, X.W.; Ma, D.L.; Li, M.; Zang, S.Y. Linking soil organic carbon mineralization with soil variables and bacterial communities in a permafrost-affected tussock wetland during laboratory incubation. Catena 2023, 221, 106783. [Google Scholar] [CrossRef]
- Dong, X.F.; Chen, Q.; Zang, S.Y.; Zhao, G.Y.; Liu, C.; Wu, X.W. Effect of temperature and moisture on soil organic carbon mineralization of predominantly permafrost forest in the Great Hing’an Mountains. Acta Sci. Circumstantiae 2019, 39, 4269–4275. (In Chinese) [Google Scholar] [CrossRef]
- Rovira, P.; Vallejo, V.R. Labile and recalcitrant pools of carbon and nitrogen in organic matter decomposing at different depths in soil: An acid hydrolysis approach. Geoderma 2002, 107, 109–141. [Google Scholar] [CrossRef]
- Zuo, Y.J.; Song, Y.Y.; Jiang, L.; Li, Y.; Wang, Y.S.; Chen, N.; Jiang, P.; Zheng, S.J.; Song, C.C.; Xu, X.F.; et al. Long-term carbon release of peatland soil after permafrost thaw in northmost China. Catena 2025, 261, 109551. [Google Scholar] [CrossRef]
- Bai, X.J.; Zhai, G.Q.; Wang, B.R.; An, S.S.; Liu, J.Z.; Xue, Z.J.; Dippold, M.A. Litter quality controls the contribution of microbial carbon to main microbial groups and soil organic carbon during its decomposition. Biol. Fertil. Soils 2024, 60, 167–181. [Google Scholar] [CrossRef]
- Chen, X.F.; Liu, M.; Jiang, C.Y.; Wu, M.; Jia, Z.J.; Li, Z.P. Mineralization of soil organic carbon and its sensitivity to temperature in soil aggregates, relative to particle size in red paddy soil. Acta Pedol. Sin. 2019, 56, 1118–1127. (In Chinese) [Google Scholar] [CrossRef]
- Wang, X.Y.; Helgason, B.; Westbrook, C.; Bedard-Haughn, A. Effect of mineral sediments on carbon mineralization, organic matter composition and microbial community dynamics in a mountain peatland. Soil Biol. Biochem. 2016, 103, 16–27. [Google Scholar] [CrossRef]
- Sun, B.Y.; Sun, R.F.; Xu, J.J.; Gao, W.J.; Chu, X.J.; Yuan, H.L.; Wan, F.X.; Yan, L.M.; Han, G.G.; Xia, J.Y.; et al. Warming-Induced Plant Species Shifts Lead to Substantial Losses of Wetland Soil Carbon. Ecol. Lett. 2025, 28, e70129. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, M.M.; Xiao, L.J.; Guo, X.W.; Zheng, J.Y.; Zhu, B.; Luo, Z.K. Reconciling carbon quality with availability predicts temperature sensitivity of global soil carbon mineralization. Proc. Natl. Acad. Sci. USA 2024, 121, e2313842121. [Google Scholar] [CrossRef]
- Xu, X.; Zhou, Y.; Ruan, H.H.; Luo, Y.Q.; Wang, J.S. Temperature sensitivity increases with soil organic carbon recalcitrance along an elevational gradient in the Wuyi Mountains, China. Soil Biol. Biochem. 2010, 42, 1811–1815. [Google Scholar] [CrossRef]
- Frøseth, R.B.; Bleken, M.A. Effect of low temperature and soil type on the decomposition rate of soil organic carbon and clover leaves, and related priming effect. Soil Biol. Biochem. 2015, 80, 156–166. [Google Scholar] [CrossRef]
- Jiang, L.; Ma, X.Y.; Song, Y.Y.; Gao, S.Q.; Ren, J.H.; Zhang, H.; Wang, X.W. Warming-induced labile carbon change soil organic carbon mineralization and microbial abundance in a northern peatland. Microorganisms 2022, 10, 1329. [Google Scholar] [CrossRef]
- Chen, J.C.; Bai, E.; Liang, Y.T.; Liu, Z.P.; Ji, Y.X.; Sun, T.T.; Guo, Z.X.; Huo, Y.D.; Liu, S.S.; Berg, B. The origin and succession of the microbial community in decomposing litter. ISME Commun. 2025, 5, ycaf155. [Google Scholar] [CrossRef]
- Keenor, S.G.; Lee, R.; Reid, B.J. Physical protection of soil carbon stocks under regenerative agriculture. Soil 2025, 11, 957–973. [Google Scholar] [CrossRef]
- Acevedo-Sandoval, O.A.; Romero-Natale, A.; Romo-Gómez, C.; Camacho-López, C.; Leyva-Morales, J.B.; Salas-Martínez, F.; González-Ramírez, C.A. Carbon Fraction Distribution in Forest Soils and Leaf Litter Across Vegetation Types in El Chico National Park, Mexico. Sustainability 2025, 17, 11028. [Google Scholar] [CrossRef]
- Sae-Tun, O.; Bodner, G.; Rosinger, C.; Zechmeister-Boltenstern, S.; Mentler, A.; Keiblinger, K. Fungal biomass and microbial necromass facilitate soil carbon sequestration and aggregate stability under different soil tillage intensities. Appl. Soil Ecol. 2022, 179, 104599. [Google Scholar] [CrossRef]
- Zhang, J.; Song, M.Y.; Zheng, S.L.; Zhou, P.; Guo, J.; He, J.N.; Yue, H.J. Distribution characteristics of top SOC in different forest types of Genhe in the Greater Khingan Range of Inner Mongolia. Sci. Rep. 2025, 15, 6736. [Google Scholar] [CrossRef]
- Qiu, Y.F.; Ying, M.S.; Tang, R.G.; Trakić, T.J.; Liu, Y.H.; Chen, Y.C.; Wang, X.; Cai, Y.J.; Chang, S.X. Temperature-dependent modulation of soil organic carbon by earthworm (Pheretima guillelmi) bioturbation in a subtropical forest: Topsoil versus Subsoil. Agric. Ecosyst. Environ. 2026, 400, 110240. [Google Scholar] [CrossRef]
- Li, Q.C.; Wang, L.L.; Fu, Y.; Lin, D.G.; Hou, M.R.; Li, X.D.; Hu, D.D.; Wang, Z.H. Transformation of soil organic matter subjected to environmental disturbance and preservation of organic matter bound to soil minerals: A review. J. Soils Sediments 2023, 23, 1485–1500. [Google Scholar] [CrossRef]
- Xu, Z.B.; Tsang, D.C.W. Mineral-mediated stability of organic carbon in soil and relevant interaction mechanisms. Eco-Environ. Health 2024, 3, 59–76. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Huang, X.Y.; Zhao, B.Y.; Yan, C.Y.; Zhao, H.Y.; Zhang, H.B.; Halamka, T.A.; Peel, R.H.; Vreeken, M.; Gallego-Sala, A.V.; et al. Microbial responses to changing plant community protect peatland carbon stores during Holocene drying. Nat. Commun. 2025, 16, 6912. [Google Scholar] [CrossRef]
- Heckman, K.A.; Possinger, A.R.; Badgley, B.D.; Bowman, M.M.; Gallo, A.C.; Hatten, J.A.; Nave, L.E.; SanClements, M.D.; Swanston, C.W.; Weiglein, T.L.; et al. Moisture-driven divergence in mineral-associated soil carbon persistence. Proc. Natl. Acad. Sci. USA 2023, 120, e2210044120. [Google Scholar] [CrossRef]
- Sun, X.L.; Liu, J.T.; Liu, S.T.; Gao, W.L. Soil organic carbon fractions comparison after 40-year long-term fertilisation in a wheat-corn rotation field. Soil Water Res. 2022, 17, 149–157. [Google Scholar] [CrossRef]
- Liu, J.; Huang, N.; Wang, L.; Lin, X.Y.; Zhu, L.Y.; Niu, Z.; Zhang, Y.L.; Duan, W.S. Analysing the spatial variation of soil respiration during the early growing season of different grasslands in China. PeerJ 2024, 12, e18480. [Google Scholar] [CrossRef]
- Li, J.Q.; Yan, D.; Pendall, E.; Pei, J.M.; Noh, N.J.; He, J.S.; Li, B.; Nie, M.; Fang, C.M. Depth dependence of soil carbon temperature sensitivity across Tibetan permafrost regions. Soil Biol. Biochem. 2018, 126, 82–90. [Google Scholar] [CrossRef]
- Yi, Y.H.; Kimball, J.S.; Jones, L.A.; Reichle, R.H.; Nemani, R.; Margolis, H.A. Recent climate and fire disturbance impacts on boreal and arctic ecosystem productivity estimated using a satellite-based terrestrial carbon flux model. J. Geophys. Res. Biogeosci. 2013, 118, 606–622. [Google Scholar] [CrossRef]
- Witzgall, K.; Vidal, A.; Schubert, D.I.; Höschen, C.; Schweizer, S.A.; Buegger, F.; Pouteau, V.; Chenu, C.; Mueller, C.W. Particulate organic matter as a functional soil component for persistent soil organic carbon. Nat. Commun. 2021, 12, 4115. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.Y.; Cheng, Y.; Anenkhonov, O.A.; Sandanov, D.V.; Wang, H.Y.; Zhou, M.; Wei, J.S.; Korolyuk, A.Y. Dynamics of the climate-permafrost-vegetation coupling system at its southernmost zone in Eurasia under climate warming. Fundam. Res. 2025, 5, 1077–1083. [Google Scholar] [CrossRef]
- Pan, Y.D.; Birdsey, R.A.; Fang, J.Y.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef]
- Bailey, V.L.; Bond-Lamberty, B.; DeAngelis, K.; Grandy, A.S.; Hawkes, C.V.; Heckman, K.; Lajtha, K.; Phillips, R.P.; Sulman, B.N.; Todd-Brown, K.E.O.; et al. Soil carbon cycling proxies: Understanding their critical role in predicting climate change feedbacks. Glob. Change Biol. 2018, 24, 895–905. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.