


Bacillus Strains with Catalase Enzyme Improve the Physiology and Growth of Rice (Oryza sativa L.)

 , ,
, ,
Abstract
:1. Introduction
2. Results
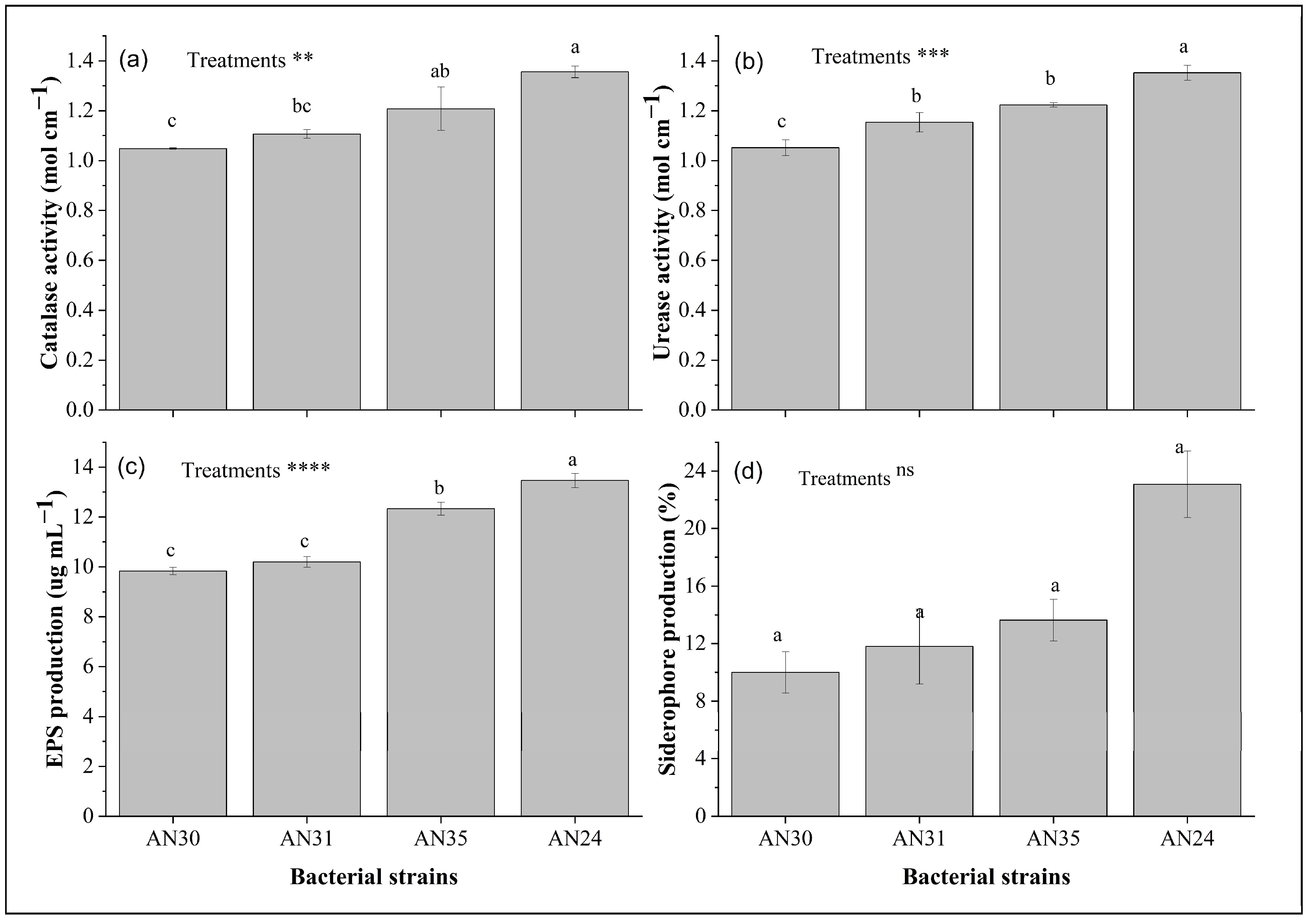
2.1. Biochemical Characterization of Bacillus Strains
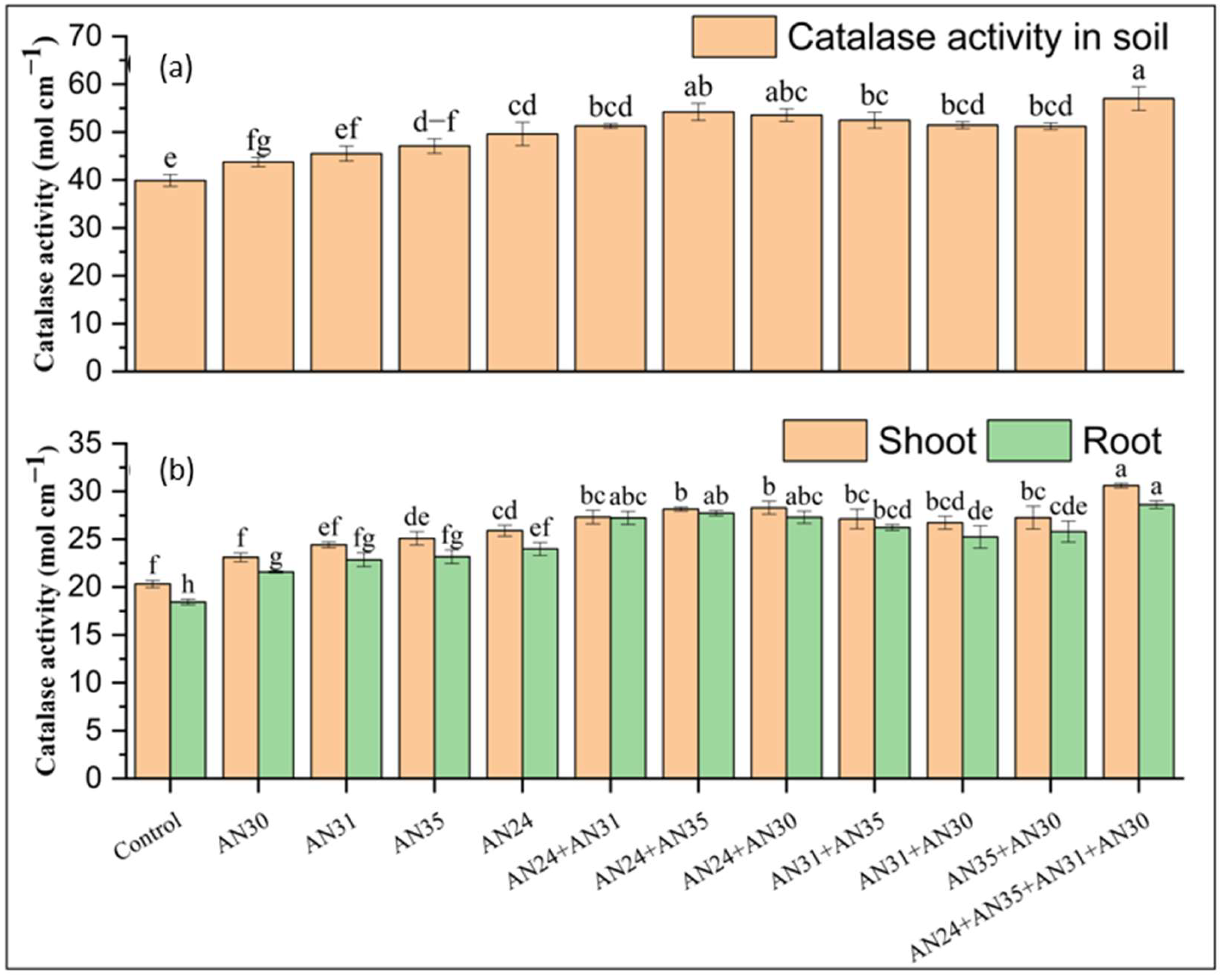
2.2. Effect of Catalase-Producing Bacillus Strains to Improve Catalase Activity in Soil and Plant
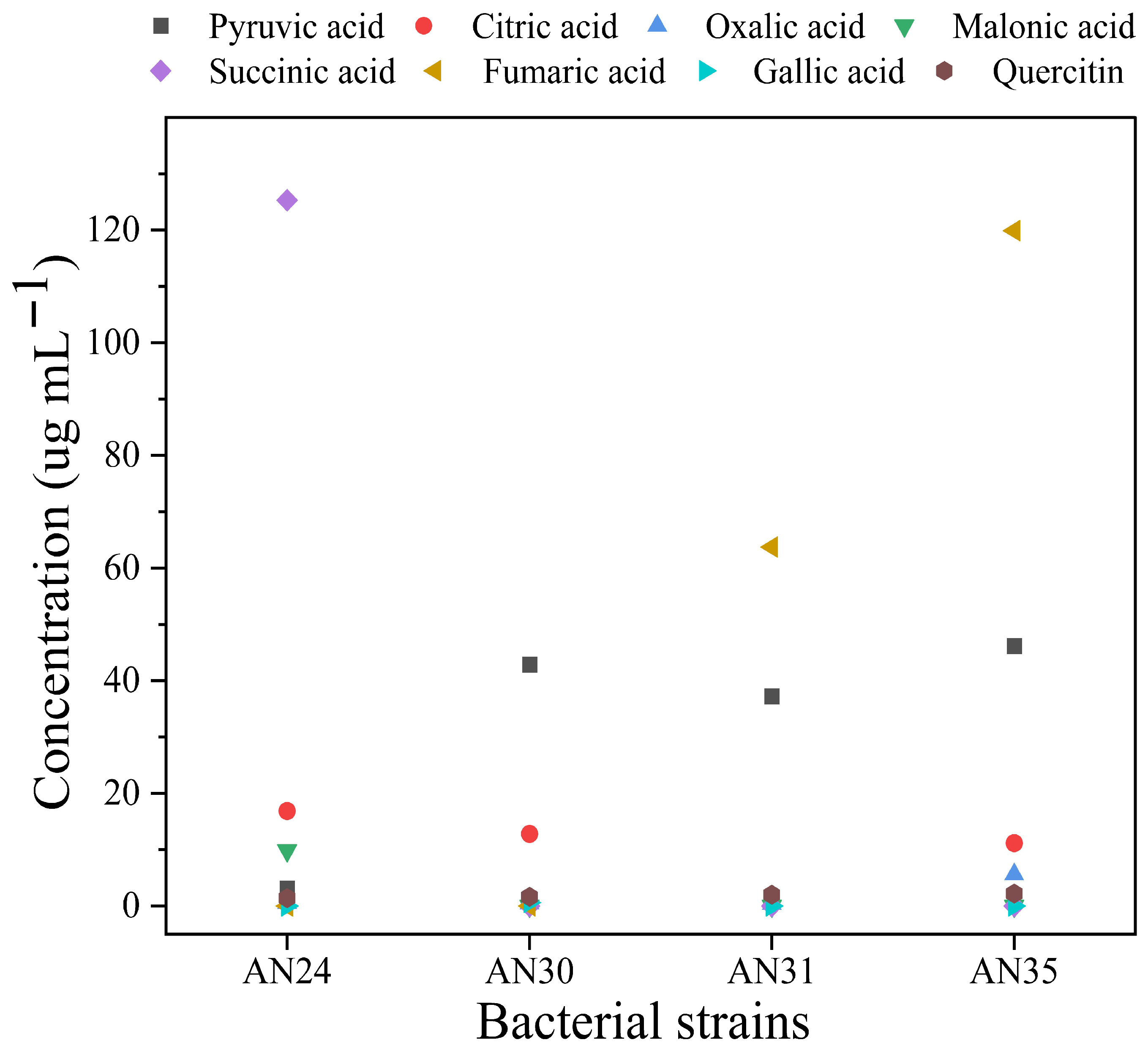
2.3. Organic and Phenolics Production in Bacterial Culture
2.4. Plant Growth Promotion by the Application of Catalase-Producing Bacillus Strains
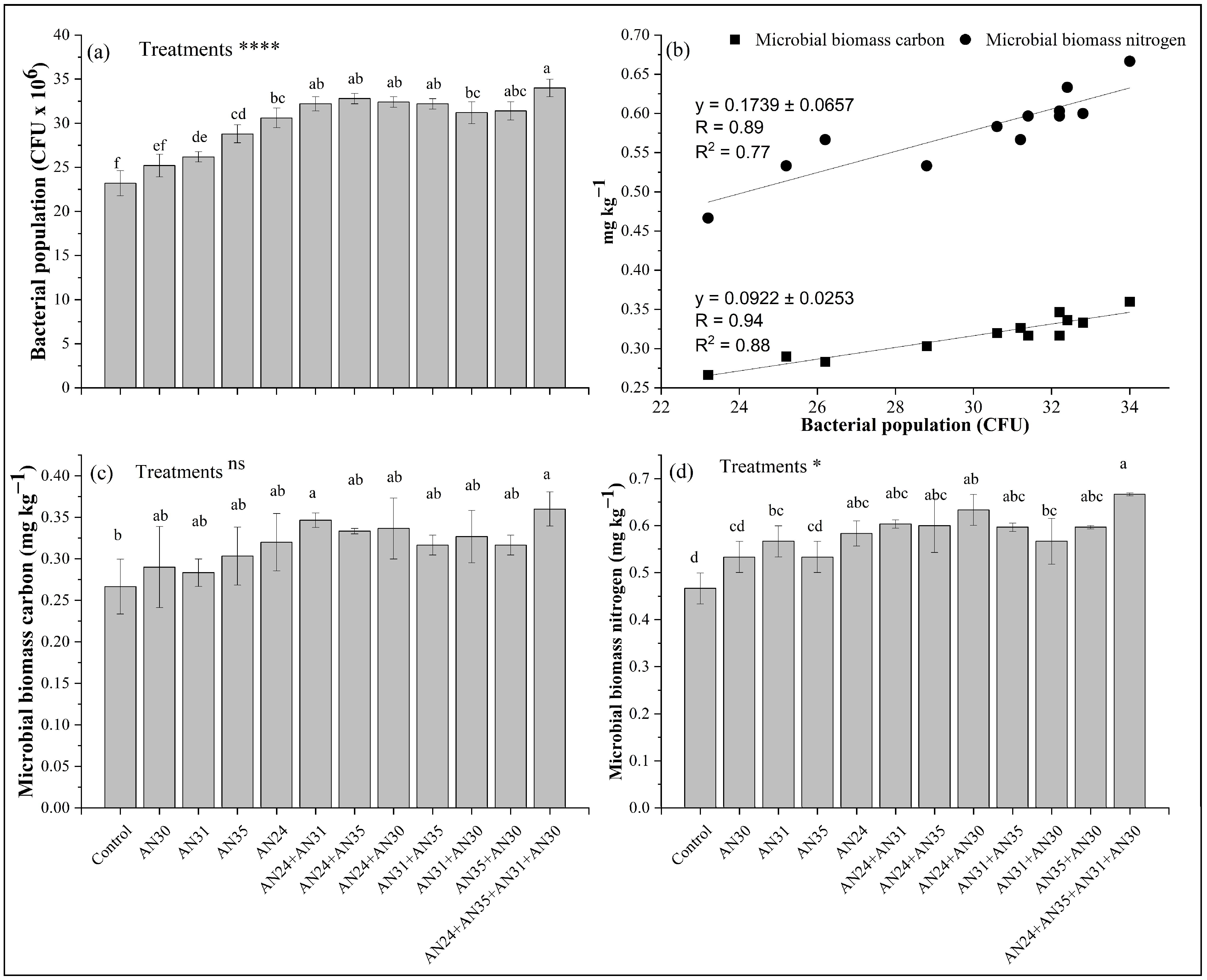
2.5. Effect of Bacillus Strains with Catalase Activity on Soil Biological Properties
3. Discussion
4. Materials and Methods
4.1. Quantitative Characterization of Bacillus Strains
4.2. Biochemical Quantification of Strains
4.3. Determination of Organic Acids
4.4. In Vitro Rice Growth Promotion Characterization of Catalase-Producing Bacillus Strains
4.5. Postharvest Soil Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thangaraj, B.; Solomon, P.R. Immobilization of lipases–A review. Part I. Enzyme Immobilization. ChemBioEng. Rev. 2019, 6, 157–166. [Google Scholar] [CrossRef]
- Nandi, A.; Yan, L.J.; Jana, C.K.; Das, N. Role of Catalase in Oxidative Stress- and Age- Associated Degenerative Diseases. Oxid. Med. Cell. Longev. 2019, 2019, 9613090. [Google Scholar] [CrossRef]
- Wang, X.; Li, S.; Liu, Y.; Ma, C. Redox regulated peroxisome homeostasis. Redox Biol. 2015, 4, 104–108. [Google Scholar] [CrossRef] [PubMed]
- Simanjuntak, E.; Zulham. Superoxide dismutase (SOD) and free radical. J. Keperawatan Dan Fisioter. JKF 2020, 2, 124–129. [Google Scholar] [CrossRef]
- Afzal, S.; Chaudhary, N.; Singh, N.K. Role of soluble sugars in metabolism and sensing under abiotic stress. In Plant Growth Regulators; Springer: Cham, Switzerland, 2021; pp. 305–334. [Google Scholar]
- Johnson, L.A.; Hug, L.A. Distribution of reactive oxygen species defense mechanisms across domain bacteria. Free Radic. Biol. Med. 2019, 140, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Noctor, G. Stress-triggered redox signaling: What’s in pROSpect? Plant Cell Environ. 2016, 39, 951–964. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Tiwari, A.; Ghosh, P.; Arora, K.; Sharma, S. Enhanced lignin degradation of paddy straw and pine needle biomass by combinatorial approach of chemical treatment and fungal enzymes for pulp making. Bioresour. Technol. 2023, 368, 128314. [Google Scholar] [CrossRef]
- Chen, C.H. Xenobiotic Metabolic Enzymes: Bioactivation and Antioxidant Defense; Springer: Cham, Switzerland, 2020; pp. 221–234. [Google Scholar]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Falade, A.O.; Mabinya, L.V.; Okoh, A.I.; Nwdo, U.U. Studies on peroxidase production and detection of Sporotrichum thermophile-like catalase-peroxidase gene in a Bacillus species isolated from Hogsback Forest reserve, South Africa. Heliyon 2019, 5, e03012. [Google Scholar] [CrossRef]
- Schlüter, U.; Bouvier, J.W.; Guerreiro, R.; Malisic, M.; Kontny, C.; Westhoff, P.; Stich, B.; Weber, A.P.M. Brassicaceae display diverse photorespiratory carbon recapturing mechanisms. bioRxiv 2022. [Google Scholar] [CrossRef]
- Eisenhut, M.; Roell, M.S.; Weber, A.P. Mechanistic understanding of photorespiration paves the way to a new green revolution. New Phytol. 2019, 223, 1762–1769. [Google Scholar] [CrossRef] [PubMed]
- Fernie, A.R.; Bauwe, H. Wasteful, essential, evolutionary stepping stone? The multiple personalities of the photorespiratory pathway. Plant J. 2020, 102, 666–677. [Google Scholar] [CrossRef] [PubMed]
- Garcia, C.P.; Filippis, D.L.; Gul, A.; Hasanuzzaman, M.M.; Ozturk, V.; Altay, M.; Lao, T. Oxidative stress and antioxidant metabolism under adverse environmental conditions: A review. Bot. Rev. 2021, 87, 421–466. [Google Scholar] [CrossRef]
- Busch, F.A. Photorespiration in the context of Rubisco biochemistry, CO2 diffusion and metabolism. Plant J. 2020, 101, 919–939. [Google Scholar] [CrossRef]
- Nosheen, A.; Bano, A. Potential of plant growth promoting rhizobacteria and chemical fertilizers on soil enzymes and plant growth. Pak. J. Bot. 2014, 46, 1521–1530. [Google Scholar]
- Khatun, M.R.; Mukta, R.H.; Islam, M.A.; Hud, A.N. Insight into Citric Acid-Induced Chromium Detoxification in Rice (Oryza sativa L). Int. J. Phytoremediation 2019, 21, 1234–1240. [Google Scholar] [CrossRef] [PubMed]
- Tahjib, U.A.M.; Zahan, M.I.; Karim, M.M.; Imran, S.; Hunter, C.T.; Islam, M.S.; Murata, Y. Citric acid-mediated abiotic stress tolerance in plants. Int. J. Mol. Sci. 2021, 22, 7235. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, D.K.; Kasotia, A.; Jain, S.; Vaishany, A.; Kumari, S.; Sharma, K.P.; Varma, A. Bacterial-mediated tolerance and resistance to plants under abiotic and biotic stresses. J. Plant Grow Regul. 2016, 35, 276–300. [Google Scholar] [CrossRef]
- Hussain, S.; Khan, F.; Cao, W.; Wu, L.; Geng, M. Seed priming alters the production and detoxifcation of reactive oxygen intermediates in rice seedlings grown under sub-optimal temperature and nutrient supply. Front. Plant Sci. 2016, 7, 439. [Google Scholar] [CrossRef]
- Bibián, M.E.; Pérez-Sánchez, A.; Mejía, A.; Barrios, G.J. Penicillin and cephalosporin biosyntheses are also regulated by reactive oxygen species. Appl. Microbiol. Biotechnol. 2020, 104, 1773–1783. [Google Scholar] [CrossRef]
- Taniguchi, I.; Yoshida, S.; Hiraga, K.; Miyamoto, K.; Kimura, Y.; Oda, K. Biodegradation of PET. Current status and application aspects. Acs Catal. 2019, 9, 4089–4105. [Google Scholar] [CrossRef]
- Chauhan, A.; Siani, R.; Sharma, J.C. Plant growth promoting rhizobacteria and their biological properties for soil enrichment and growth promotion. J. Plant Nutr. 2021, 45, 273–299. [Google Scholar] [CrossRef]
- Harman, G.; Khadka, R.; Doni, F.; Uphoff, N. Benefits to plant health and productivity from enhancing plant microbial symbionts. Front. Plant Sci. 2021, 11, 610065. [Google Scholar] [CrossRef]
- Babiker, B.M.; Ahmed, A.E.; Ibrahim, H.M. Isolation & identification of catalase producing Bacillus spp: A comparative study. Int. J. Adv. Res. 2017, 4, 1206–1211. [Google Scholar]
- Philibert, T.; Rao, Z.; Yang, T.; Zhou, J.; Huang, G.; Irene, K.; Samuel, N. Heterologous expression and characterization of a new heme-catalase in Bacillus subtilis 168. J. Ind. Microbiol. Biotechnol. 2016, 43, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.O.; Bai, Z.; Bao, L.; Xue, L.; Zhang, S.; Wei, Y. Bacillus subtilis biofertilizer mitigating agricultural ammonia emission and shifting soil nitrogen cycling microbiomes. Environ. Int. 2020, 144, 105989. [Google Scholar] [CrossRef]
- Anbu, P.; Kang, C.H.; Shin, Y.J.; So, J.S. Formations of calcium carbonate minerals by bacteria and its multiple applications. Springer Plus 2016, 5, 1–26. [Google Scholar] [CrossRef]
- Dar, A.A.; Pan, B.; Qin, J.; Zhu, Q.; Lichtfouse, E.; Usman, M.; Wang, C. A review on sustainable ferrate oxidation: Reaction chemistry, mechanisms and applications to eliminate micro pollutant (pharmaceuticals) in wastewater. Environ. Pollut. 2021, 290, 117957. [Google Scholar] [CrossRef]
- Morcillo, R.; Manzanera, M. The Effects of Plant-Associated Bacterial Exopolysaccharides on Plant Abiotic Stress Tolerance. Metabolites 2021, 11, 337. [Google Scholar] [CrossRef] [PubMed]
- Naseer, I.; Ahmad, M.; Hussain, A.; Jamil, M. Potential of zinc solubilizing Bacillus strains to improve rice growth under axenic conditions. Pak. J. Agric. Sci. 2020, 57, 1057–1071. [Google Scholar]
- Mumtaz, M.Z.; Barrya, K.M.; Bakera, A.L.; Nichols, D.S.; Ahmad, M.; Zahir, Z.A.; Britza, M.L. Production of lactic and acetic acids by Bacillus sp. ZM20 and Bacillus cereus following exposure to zinc oxide: A possible mechanism for Zn solubilization. Rhizosphere 2019, 12, 100170. [Google Scholar] [CrossRef]
- Timmusk, S.; Behers, L.; Muthoni, J.; Muraya, A.; Aronsson, A.C. Perspectives and challenges of microbial application for crop improvement. Front. Plant. Sci. 2017, 8, 49. [Google Scholar]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Singh, B.K. Plant-microbiome interactions: From community to plant health. Nat. Rev. Microbial. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.Q.; Lu, X.B.; Li, Z.H.; Tian, C.Y.; Song, J. The role of root-associated microbes in growth stimulation of plants under saline conditions. Land Degrad. Dev. 2021, 32, 3471–3486. [Google Scholar] [CrossRef]
- Jian, S.; Li, J.; Chen, J.I.; Wang, G.; Mayes, M.A.; Dzantor, K.E.; Hui, D.; Luo, Y. Soil extracellular enzyme activities, soil carbon and nitrogen storage under nitrogen fertilization: A meta-analysis. Soil Biol. Biochem. 2016, 101, 32–43. [Google Scholar] [CrossRef]
- Hussain, A.; Zahir, Z.A.; Ditta, A.; Tahir, M.U.; Ahmad, M.; Mumtaz, M.Z.; Hayat, K.; Hussain, S. Production and implication of bio-activated organic fertilizer enriched with zinc-solubilizing bacteria to boost up maize (Zea mays L.) production and biofortification under two cropping seasons. Agronomy 2020, 10, 39. [Google Scholar] [CrossRef]
- Lukashe, N.S.; Mupambwa, H.A.; Green, E.; Mnkeni, P.N.S. Inoculation of fy ash amended vermicompost with phosphate solubilizing bacteria (Pseudomonas fuorescens) and its influence on vermi-degradation, nutrient release and biological activity. Waste Manag. 2019, 84, 14–22. [Google Scholar] [CrossRef]
- Iqbal, Z.; Bushra; Hussain, A.; Dar, A.; Ahmad, M.; Wang, X.; Brtnicky, M.; Mustafa, A. Combined Use of Novel Endophytic and Rhizobacterial Strains Upregulates Antioxidant Enzyme Systems and Mineral Accumulation in Wheat. Agronomy 2022, 12, 551. [Google Scholar] [CrossRef]
- Beers. R., F.; Sizer, I.W. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J. Biol. Chem. 1952, 195, 133–140. [Google Scholar] [CrossRef]
- Hildebrandt, G.; Roots, I. Reduced nicotinamide adenine dinucleotide phosphate (NADPH)-dependent formation and breakdown of hydrogen peroxide during mixed function oxidation reactions in liver microsomes. Arch. Biochem. Biophy. 1975, 171, 385–397. [Google Scholar] [CrossRef]
- Burbank, M.B.; Weaver, T.J.; Williams, B.C.; Crawford, R.L. Urease activity of ureolytic bacteria isolation from six soils in which calcite was precipitated by indigenous bacteria. Geomicrobiol. J. 2012, 29, 389–395. [Google Scholar] [CrossRef]
- Cappuccino, J.G.; Sherman, N. Microbiology. In Laboratory Manual, 8th ed.; Pearson: London, UK, 2002; Volume 13, ISBN 978-0805325782. [Google Scholar]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Payne, S.M. Iron acquisition in microbial pathogenesis. Trends Microbiol. 1993, 1, 66–69. [Google Scholar] [CrossRef]
- Ashraf, M.; Hasnain, S.; Berge, O.; Mahmood, T. Inoculating wheat seedlings with exopolysaccharide-producing bacteria restricts sodium uptake and stimulates plant growth under salt stress. Biol. Fertil. Soils 2004, 40, 157–162. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Butsat, N.; Weerapreeyakul, N.; Siriamornpun, S. Change in the phenolic acids and antioxidant activity in Thai rice husk at fie growth stages during grain development. J. Agric. Food Chem. 2009, 57, 4566–4571. [Google Scholar] [CrossRef] [PubMed]
- Bertani, G. Studies on Lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. J. Bacteriol. 1952, 62, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Wollum, A.G., II. Cultural methods for soil microorganisms. In Methods of Soil Analysis: Chemical and Microbial Properties; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; ASA and SSSA Publication: Madison, WI, USA, 1982; pp. 718–802. [Google Scholar]
- Cao, J.; Ji, D.; Wang, C. Interaction between earthworms and arbuscular mycorrhizal fungi on the degradation of oxytetracycline in soils. Soil Biol. Biochem. 2015, 90, 283–292. [Google Scholar] [CrossRef]
- Li, Q.; Liang, J.H.; He, Y.Y.; Hu, Q.J.; Yu, S. Effect of land use on soil enzyme activities at karst area in Nanchuan, Chongqing Southwest China. Plant Soil Environ. 2014, 60, 15–20. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Shoot Length (cm) | Root Length (cm) | Dry Weight (g Seedling−1) | Root Surface Area (cm2) | Root Diameter (mm) | Root Volume (cm3) |
|---|---|---|---|---|---|---|
| Control | 6.5 ± 0.08 f | 4.3 ± 0.12 f | 0.51 ± 0.01 f | 9.8 ± 0.06 d | 0.53 ± 0.03 e | 0.37 ± 0.09 c |
| AN30 | 7.1 ± 0.18 e | 4.9 ± 0.11 e | 0.59 ± 0.02 de | 10.1 ± 0.15 d | 0.57 ± 0.03 de | 0.39 ± 0.05 ac |
| AN31 | 7.9 ± 0.16 de | 5.2 ± 0.16 de | 0.61 ± 0.01 de | 10.9 ± 0.06 cd | 0.60 ± 0.06 de | 0.40 ± 0.06 bc |
| AN35 | 8.0 ± 0.32 de | 5.5 ± 0.10 de | 0.58 ± 0.01 e | 11.3 ± 0.25 cd | 0.63 ± 0.03 cde | 0.43 ± 0.03 ac |
| AN24 | 8.3 ± 0.23 cd | 5.7 ± 0.05 cd | 0.64 ± 0.01 cd | 11.8 ± 0.09 bc | 0.63 ± 0.03 cde | 0.44 ± 0.04 ac |
| AN24 + AN31 | 8.8 ± 0.09 b | 6.1 ± 0.10 b | 0.67 ± 0.03 bc | 12.5 ± 0.83 b | 0.77 ± 0.03 abc | 0.48 ± 0.02 ac |
| AN24 + AN35 | 8.8 ± 0.16 b | 6.1 ± 0.16 b | 0.71 ± 0.01 b | 12.4 ± 0.27 b | 0.67 ± 0.03 cde | 0.47 ± 0.03 ac |
| AN24 + AN30 | 8.7 ± 0.05 bc | 5.8 ± 0.09 bc | 0.68 ± 0.00 bc | 12.3 ± 0.24 b | 0.80 ± 0.06 ab | 0.47 ± 0.03 ac |
| AN31 + AN35 | 8.5 ± 0.09 bc | 5.8 ± 0.09 bc | 0.68 ± 0.01 bc | 12.0 ± 0.94 bc | 0.63 ± 0.03 de | 0.47 ± 0.03 ac |
| AN31 + AN30 | 8.8 ± 0.08 bc | 5.9 ± 0.08 bc | 0.66 ± 0.03 bc | 11.9 ± 0.73 bc | 0.70 ± 0.06 bcd | 0.51 ± 0.01 ab |
| AN35 + AN30 | 8.4 ± 0.16 bc | 5.8 ± 0.09 bc | 0.67 ± 0.02 bc | 12.0 ± 0.58 bc | 0.70 ± 0.06 bcd | 0.47 ± 0.03 ac |
| AN24 + AN35 + AN31 + AN30 | 9.4 ± 0.26 a | 6.6 ± 0.10 a | 0.80 ± 0.02 a | 13.0 ± 0.25 a | 0.87 ± 0.12 a | 0.54 ± 0.07 a |
| LSD (p ≤ 0.05) | 0.5161 | 0.3056 | 0.0562 | 1.3950 | 0.1589 | 0.1317 |
| Analysis | Unit | Values |
|---|---|---|
| Textural class | Sandy loam | |
| ECe | dS m−1 | 1.59 ± 0.0233 |
| pH | 7.91 ± 0.0581 | |
| Saturation percentage | % | 31.73 ± 0.2333 |
| Organic matter | % | 0.58 ± 0.0088 |
| Phosphorus | mg kg−1 | 3.02 ± 0.0722 |
| Potassium | mg kg−1 | 75.50 ± 1.0408 |
| Nitrogen | % | 0.03 ± 0.0006 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saeed, S.W.Z.; Naseer, I.; Zahir, Z.A.; Hilger, T.; Shahid, S.; Iqbal, Z.; Ahmad, M. Bacillus Strains with Catalase Enzyme Improve the Physiology and Growth of Rice (Oryza sativa L.). Stresses 2023, 3, 736-748. https://doi.org/10.3390/stresses3040050
Saeed SWZ, Naseer I, Zahir ZA, Hilger T, Shahid S, Iqbal Z, Ahmad M. Bacillus Strains with Catalase Enzyme Improve the Physiology and Growth of Rice (Oryza sativa L.). Stresses. 2023; 3(4):736-748. https://doi.org/10.3390/stresses3040050
Chicago/Turabian StyleSaeed, Syeda Wadia Zainab, Iqra Naseer, Zahir Ahmad Zahir, Thomas Hilger, Shumila Shahid, Zafar Iqbal, and Maqshoof Ahmad. 2023. "Bacillus Strains with Catalase Enzyme Improve the Physiology and Growth of Rice (Oryza sativa L.)" Stresses 3, no. 4: 736-748. https://doi.org/10.3390/stresses3040050
APA StyleSaeed, S. W. Z., Naseer, I., Zahir, Z. A., Hilger, T., Shahid, S., Iqbal, Z., & Ahmad, M. (2023). Bacillus Strains with Catalase Enzyme Improve the Physiology and Growth of Rice (Oryza sativa L.). Stresses, 3(4), 736-748. https://doi.org/10.3390/stresses3040050