
Isolation and Phenotypic Microarray Profiling of Different Pseudomonas Strains Isolated from the Rhizosphere of Curcuma longa L.

 ,
, 
Abstract
1. Introduction
2. Result and Discussion
2.1. Plant Growth Promotion (PGP) Activity
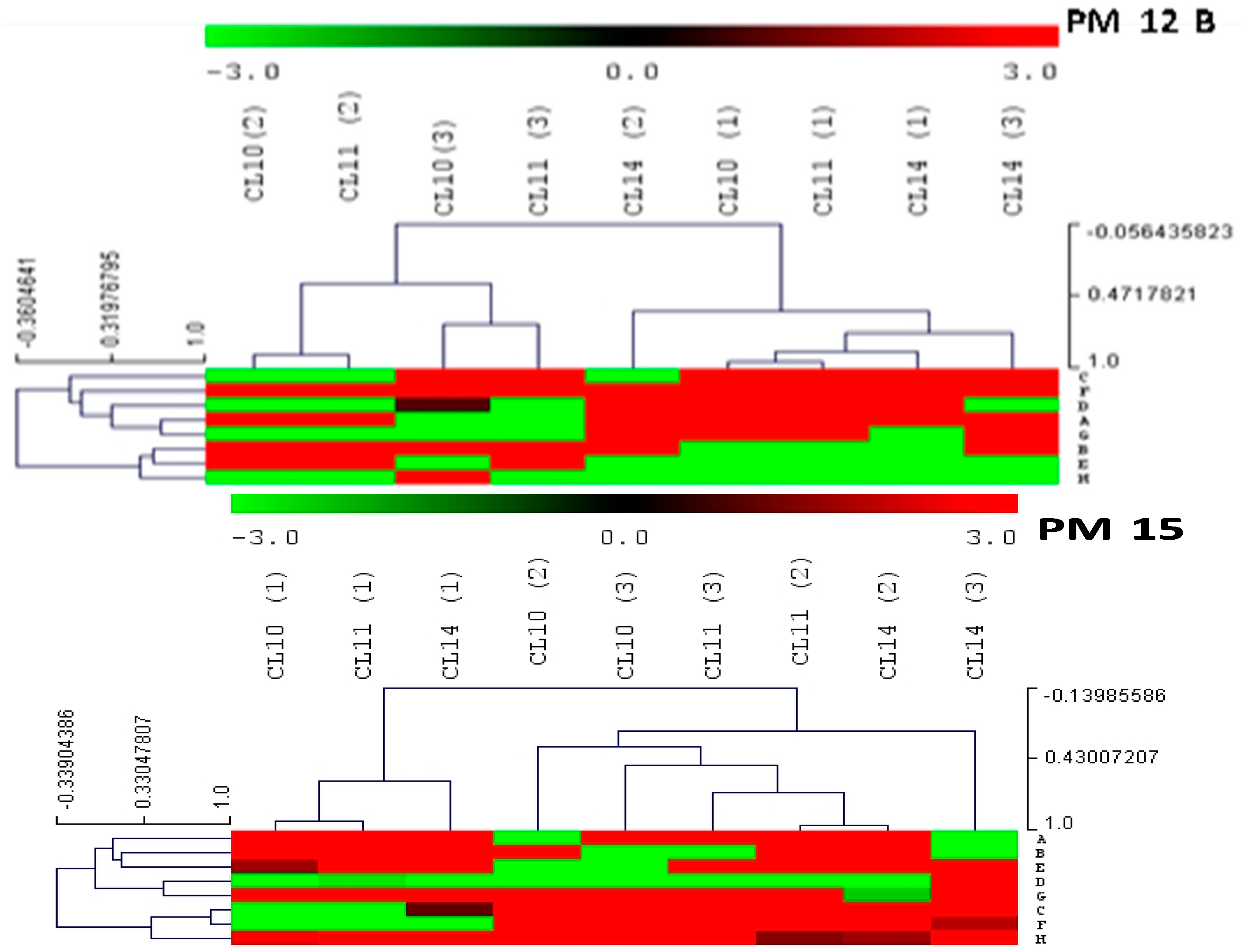
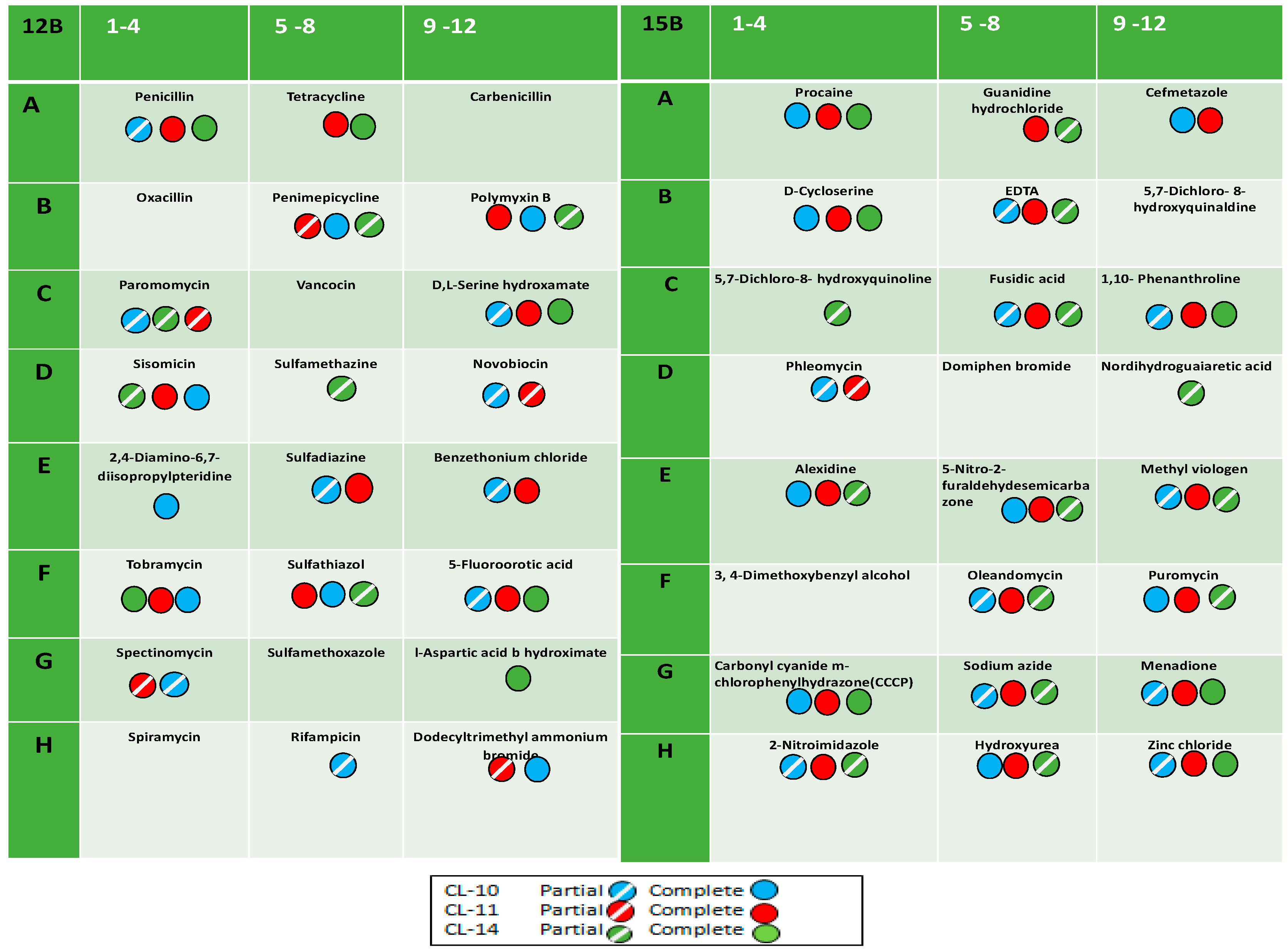
2.2. Antibiotic and Chemical Sensitivity Using Phenotypic Microarray Technology
3. Materials and Methods
3.1. Isolation of Rhizospheric Bacteria
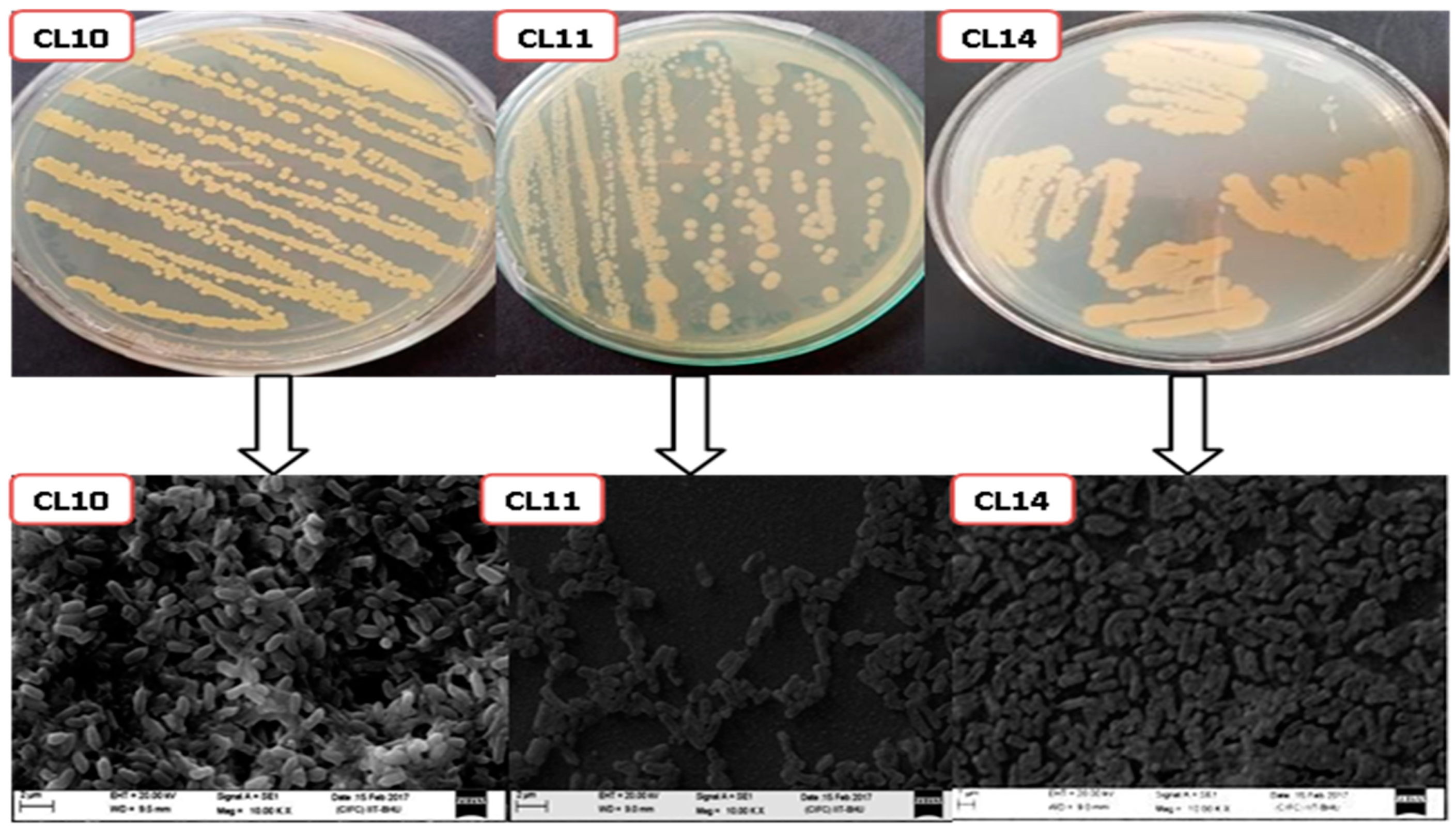
3.2. Characterization of the Bacterial Isolates
3.3. Scanning Electron Microscopy Analysis of Bacterial Strains
3.4. Plant Growth Promoting (PGP) Traits Analysis of Bacterial Isolates
3.5. Salt Tolerance
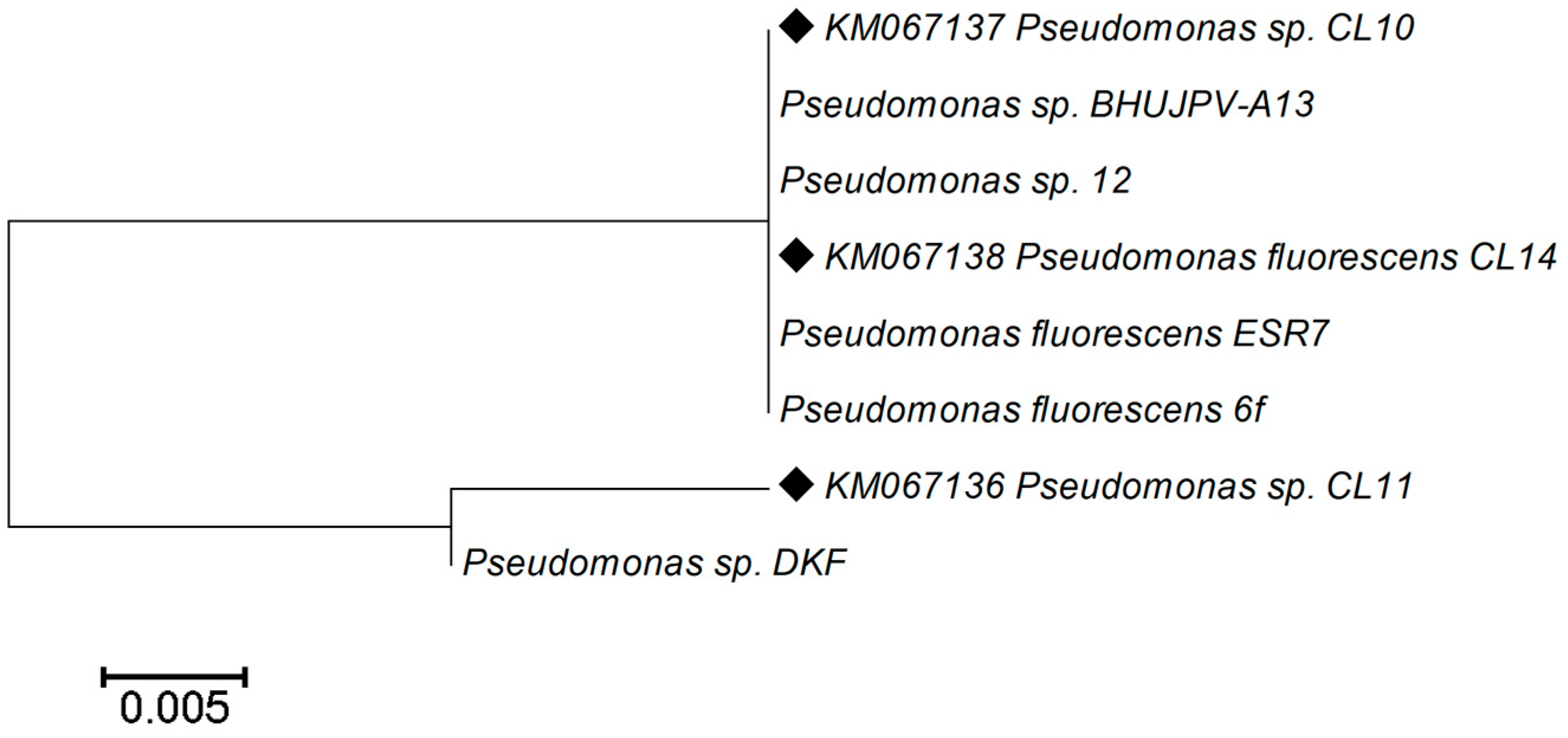
3.6. 16S rRNA Gene Amplification, Sequencing, and Their Analysis
3.7. Evaluation of Antibiotic Sensitivity by Using PM Plates of Biolog Microstation System
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kumar, A. Microbial Biocontrol: Food Security and Post Harvest Management; Springer Nature: Cham, Switzerland, 2022; Volume 2, pp. 1–334. [Google Scholar]
- Souza, R.D.; Ambrosini, A.; Passaglia, L.M. Plant growth-promoting bacteria as inoculants in agricultural soils. Genet. Mol. Biol. 2015, 38, 401–419. [Google Scholar] [CrossRef] [PubMed]
- Glick, B. Plant growth-promoting bacteria: Mechanisms and applications. Australas. Sci. 2012, 2012, 963401. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishna, W.; Yadav, R.; Li, K. Plant growth promoting bacteria in agriculture: Two sides of a coin. Appl. Soil Ecol. 2019, 138, 10–18. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moënne-Loccoz, T. The rhizosphere: A play ground and battlefield for soil-borne pathogens and beneficial microorganisms. Plant Soil 2009, 321, 341–361. [Google Scholar] [CrossRef]
- Choudhary, D.K.; Sharma, K.P.; Gaur, R.K. Biotechnological perspectives of microbes in agroecosystems. Biotechnol. Lett. 2011, 33, 1905–1910. [Google Scholar] [CrossRef]
- Garcia de Salamone, I.E.; Hynes, R.K.; Nelson, L.M. Cytokinin production by plant growth promoting rhizobacteria and selected mutants. Can. J. Microbiol. 2001, 47, 404–411. [Google Scholar] [CrossRef]
- Mavrodi, O.V.; McWilliams, J.R.; Peter, J.O.; Berim, A.; Hassan, K.A.; Elbourne, L.D.H.; LeTourneau, M.K.; Gang, D.R.; Paulsen, I.T.; Weller, D.M.; et al. Root Exudates Alter the Expression of Diverse Metabolic, Transport, Regulatory, and Stress Response Genes in Rhizosphere Pseudomonas. Front. Microbiol. 2021, 12, 651282. [Google Scholar] [CrossRef]
- Yu, K.; Pieterse, C.M.J.; Bakker, P.A.H.M.; Berendsen, R.L. Beneficial microbes going underground of root immunity. Plant Cell Environ. 2019, 42, 2860–2870. [Google Scholar] [CrossRef]
- Bais, H.P.; Tiffany, L.W.; Laura, G.P.; Gilroy, S.; Vivanco, J.M. The Role of Root Exudates in Rhizosphere Interactions with Plants and Other Organisms. Annu Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef]
- Kumar, A.; Bahadur, I.; Maurya, B.; Raghuwanshi, R.; Meena, V.; Singh, D. Does a plant growth promoting rhizobacteria enhance agricultural sustainability. J. Pure Appl. Microbiol. 2015, 9, 715–724. [Google Scholar]
- Yang, D.; Wang, L.; Wang, T.; Zhang, Y.; Zhang, S.; Luo, Y. Plant Growth-Promoting Rhizobacteria HN6 Induced the Change and Reorganization of Fusarium Microflora in the Rhizosphere of Banana Seedlings to Construct a Healthy Banana Microflora. Front. Microbiol. 2021, 12, 685408. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Kumar, A.; Singh, M.; Maurya, S.K.; Pandey, K.D. Microbial diversity in the rhizosphere of Momordica charaentia L. Bitter Gourd. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 67–76. [Google Scholar] [CrossRef]
- Ordookhani, K.; Zare, S.M. Influence of PGPR ongrowth, essential oil and nutrients uptake of sweet basil. Adv. Environ. Biol. 2011, 5, 672–677. [Google Scholar]
- Ostadi, A.; Arab-Zozani, M.; Zarei, E.; Ferns, G.A.; Bahrami, A. Therapeutic effect of turmeric on radiodermatitis: A systematic review. Physiol. Rep. 2023, 11, e15624. [Google Scholar] [CrossRef]
- Panknin, T.M.; Howe, C.L.; Hauer, M.; Bucchireddigari, B.; Rossi, A.M.; Funk, J.L. Curcumin Supplementation and Human Disease: A Scoping Review of Clinical Trials. Int. J. Mol. Sci. 2023, 24, 4476. [Google Scholar] [CrossRef]
- Rattis, B.A.C.; Ramos, S.G.; Celes, M.R.N. Curcumin as a Potential Treatment for COVID-19. Front. Pharmacol. 2021, 12, 675287. [Google Scholar] [CrossRef]
- Rizvi, A.; Ahmed, B.; Khan, M.S.; El-Beltagi, H.S.; Umar, S.; Lee, J. Bioprospecting Plant Growth Promoting Rhizobacteria for Enhancing the Biological Properties and Phytochemical Composition of Medicinally Important Crops. Molecules 2022, 27, 1407. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, A.K.; Kaushik, M.S.; Mishra, S.K.; Raj, P.; Singh, P.K.; Pandey, K.D. Interaction of Turmeric (Curcuma longa L.) with Beneficial Microbes: A Review. 3 Biotech. 2017, 7, 357. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, M.; Singh, P.P.; Singh, S.K.; Singh, P.K.; Pandey, K.D. Isolation of Plant Growth Promoting Rhizobacteria and Their Impact on Growth and Curcumin Content in Curcuma longa L. Biocatal. Agric. Biotechnol. 2016, 8, 1–7. [Google Scholar] [CrossRef]
- Kumar, A.; Singh, R.; Yadav, A.; Giri, D.D.; Singh, P.K.; Pandey, K.D. Isolation and characterization of bacterial endophytes of Curcuma longa L. 3 Biotech. 2016, 6, 60. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Schroth, M.N.; Miller, T.D. Effects of rhizosphere colonization by plant growth-promoting rhizobacteria on potato plant development and yield. Phytopathology 1980, 70, 1078–1082. [Google Scholar] [CrossRef]
- Chandran, H.; Meena, M.; Swapnil, P. Plant Growth-Promoting Rhizobacteria as a Green Alternative for Sustainable Agriculture. Sustainability 2021, 13, 10986. [Google Scholar] [CrossRef]
- Basu, A.; Prasad, P.; Das, S.N.; Kalam, S.; Sayyed, R.Z.; Reddy, M.S.; El Enshasy, H. Plant Growth Promoting Rhizobacteria (PGPR) as Green Bioinoculants: Recent Developments, Constraints, and Prospects. Sustainability 2021, 13, 1140. [Google Scholar] [CrossRef]
- Shaharoona, B.; Naveed, M.; Arshad, M.; Zahir, Z.A. Fertilizer-dependent efficiency of Pseudomonads for improving growth, yield, and nutrient use efficiency of wheat (Triticum aestivum L.). Appl. Microbiol. Biotechnol. 2008, 79, 147–155. [Google Scholar] [CrossRef]
- Arshad, M.; Shaharoona, B.; Mahmood, T. Inoculation with Pseudomonas spp. containing ACC-deaminase partially eliminates the effects of drought stress on growth, yield, and ripening of pea (Pisum sativum L.). Pedosphere 2008, 18, 611–620. [Google Scholar] [CrossRef]
- Mahdi, I.; Fahsi, N.; Hijri, M.; Sobeh, M. Antibiotic resistance in plant growth promoting bacteria: A comprehensive review and future perspectives to mitigate potential gene invasion risks. Front. Microbiol. 2022, 13, 999988. [Google Scholar] [CrossRef]
- Kang, Y.; Shen, M.; Xia, D.; Ye, K.; Zhao, Q.; Hu, J. Caution of intensified spread of antibiotic resistance genes by inadvertent introduction of beneficial bacteria into soil. Acta Agric. Scand. Sect. B Soil Plant Sci. 2017, 67, 576–582. [Google Scholar] [CrossRef]
- Blumenstein, K.; Macaya-Sanz, D.; Martín, J.A.; Albrectsen, B.R.; Witzell, J. Phenotype MicroArrays as a complementary tool to next generation sequencing for characterization of tree endophytes. Front. Microbiol. 2015, 6, 1033. [Google Scholar] [CrossRef]
- Kumar, B.; Kumar, M.S.; Annapurna, K.; Maheshwari, D.K. Genetic diversity of plant growth-promoting rhizobia isolated from a medicinal legume, Mucuna pruriens Linn. Curr. Sci. 2006, 91, 1524–1529. [Google Scholar]
- Bochner, B.R.; Gadzinski, P.; Panomitros, E. Phenotype MicroArrays for high throughput phenotypic testing and assay of gene function. Genome Res. 2001, 11, 1246–1255. [Google Scholar] [CrossRef]
- Singh, M.; Kumar, A.; Pandey, K.D. Biochemical and molecular identification of Solanum lycopersicum L. temperature tolerant bacterial endophytes. Biocat. Agric. Biotechnol. 2019, 22, 101409. [Google Scholar] [CrossRef]
- Galardini, M.; Mengoni, A.; Biondi, E.G.; Semeraro, R.; Florio, A.; Bazzicalupo, M.; Benedetti, A.; Mocali, S. Duct Ape: A suite for the analysis and correlation of genomic and Omni Log ™ Phenotype Microarray data. Genomics 2014, 103, 1–10. [Google Scholar] [CrossRef]
- Chojniak, J.; Wasilkowski, D.; Płaza, G.; Mrozik, A.; Brigmon, R. Application of Biolog Microarrays Techniques for Characterization of Functional Diversity of Microbial Community in Phenolic-contaminated Water. Int. J. Environ. Res. 2015, 9, 785–794. [Google Scholar]
- Jałowiecki, L.; Chojniak, J.; Dorgelo, E.; Hegedusova, B.; Ejhed, H.; Magnér, J.; Płaza, G. Using phenotype microarrays in the assessment of the antibiotic susceptibility profile of bacteria isolated from wastewater in on-site treatment facilities. Folia Microbiol. 2017, 62, 453–461. [Google Scholar] [CrossRef]
- Decorosi, F.; Santopolo, L.; Mora, D.; Viti, C.; Giovannetti, L. The improvement of a phenotype microarray protocol for the chemical sensitivity analysis of Streptococcus thermophilus. J. Microbiol. Meth. 2011, 86, 258–261. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Maringoni, A.C.; Theodoro, G.F.; Ming, L.C.; Cardoso, J.C.; Kurozawa, C. First report of Pseudomonas cichorii on Turmeric (Curcuma longa) in Brazil. New Dis. Rep. 2003, 7, 16. [Google Scholar] [CrossRef]
- Sah, S.; Krishnani, S.; Singh, R. Pseudomonas mediated nutritional and growth promotional activities for sustainable food security. Curr. Res. Microb. Sci. 2021, 4, 100084. [Google Scholar] [CrossRef]
- Ansari, F.A.; Jabeen, M.; Ahmad, I. Pseudomonas azotoformans FAP5, a novel biofilm forming PGPR strain, alleviates drought stress in wheat plant. Int. J. Environ. Sci. Technol. 2021, 18, 3855–3870. [Google Scholar] [CrossRef]
- Zaidi, A.; Khan, M.S.; Ahemad, M.; Oves, M. Plant growth promotion by phosphate solubilizing bacteria. Acta Microbiol. Immunol. Hung. 2009, 56, 263–284. [Google Scholar] [CrossRef]
- Taghavi, S.; Garafola, C.; Monchy, S.; Newma, L.; Hoffman, A.; Weyens, N. Genome survey and characterization of endophytic bacteria exhibiting a beneficial effect on growth and development of poplar. Appl. Environ. Microbiol. 2009, 75, 748–757. [Google Scholar] [CrossRef]
- Shi, Y.; Lou, K.; Li, C. Isolation quantity distribution and characterization of endophytic microorganisms within sugarbeet. Afr. J. Biotechnol. 2009, 8, 835–840. [Google Scholar]
- Khan, A.; Singh, P.; Shrivastava, A. Synthesis, nature and utility of universal iron chelator—Siderophore: A review. Microbiol. Res. 2018, 7, 103–111. [Google Scholar] [CrossRef]
- Laslo, E.; Gyorgy, E.; Mara, G.; Tamas, E.; Abraham, B.; Lanyi, S. Screening of plant growth promoting rhizobacteria as potential microbial inoculants. Crop Prot. 2012, 40, 43–48. [Google Scholar] [CrossRef]
- Zaho, K.; Penttinen, P.; Zhang, X.; Ao, X.; Liu, M.; Yu, X.; Chen, Q. Maize rhizosphere in Sichuan, China, host plant growth promoting Burkholderia cepacia with phosphate solubilizing and antifungal abilities. Microb. Res. 2014, 169, 76–82. [Google Scholar] [CrossRef]
- Ilangumaran, G.; Smith, D.L. Plant Growth Promoting Rhizobacteria in Amelioration of Salinity Stress: A Systems Biology Perspective. Front. Plant Sci. 2017, 8, 1768. [Google Scholar] [CrossRef] [PubMed]
- Chhabra, R. Nutrient requirement for sodic soil. Fertil. News 1995, 40, 13–21. [Google Scholar]
- Rashid, R.; Charles, T.C.; Glick, B.R. Isolation and characterization of new plant growth promoting bacterial endophyte. Appl. Soil. Ecol. 2012, 61, 217–224. [Google Scholar] [CrossRef]
- Cai, D.; Xu, Y.; Zhao, F.; Zhang, Y.; Duan, H.; Guo, X. Improved salt tolerance of Chenopodium quinoa Willd. contributed by Pseudomonas sp. strain M3035. Peer J. 2021, 9, e10702. [Google Scholar] [CrossRef] [PubMed]
- Borglin, S.; Joyner, D.; DeAngelis, K.M.; Khudyakov, J.; D’haeseleer, P.; Joachimiak, M.P.; Hazen, T. Application of phenotypic microarrays to environmental microbiology. Curr. Opin. Biotechnol. 2012, 23, 41–48. [Google Scholar] [CrossRef]
- Vaas, L.A.; Sikorski, J.; Hofner, B.; Fiebig, A.; Buddruhs, N.; Klenk, H.P.; Göker, M. opm: An R package for analysing OmniLog® phenotype microarray data. Bioinformatics 2013, 29, 1823–1824. [Google Scholar] [CrossRef] [PubMed]
- Bochner, B. New technologies to assess genotype-phenotype relationships. Nat. Rev. Genet. 2003, 4, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Greetham, D. Phenotype microarray technology and its application in industrial biotechnology. Biotechnol. Lett. 2014, 36, 1153–1160. [Google Scholar] [CrossRef]
- Santopolo, L.; Marchi, E.; Frediani, L.; Decorosi, F.; Viti, C.; Giovannetti, L. A novel approach combining the Calgary Biofilm Device and Phenotype MicroArray for the characterization of the chemical sensitivity of bacterial biofilms. Biofouling 2012, 28, 1023–1032. [Google Scholar] [CrossRef] [PubMed]
- Cray, J.A.; Bell, A.N.; Bhaganna, P.; Mswaka, A.Y.; Timson, D.J.; Hallsworth, J.E. The biology of habitat dominance; can microbes behave as weeds? Microb. Biotechnol. 2013, 6, 453–492. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Pandey, A.; Palni, L.M.S.; Bag, N.; Tamang, M. Colonization of Rhizosphere of Tea by Growth Promoting Bacteria. 2004. Available online: https://repository.up.ac.za/handle/2263/8370 (accessed on 20 June 2023).
- Wani, P.A.; Khan, M.S. Nickel detoxification and plant growth promotion by multi metal resistant plant growth promoting rhizobium species RL9. Bull. Environ. Contam. Toxicol. 2013, 91, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Wellington, E.M.H.; Boxall, A.B.A.; Cross, P.; Feil, E.J.; Gaze, W.H.; Hawkey, P.M. The role of the natural environment in the emergence of antibiotic resistance in gram-negative bacteria. Lancet Infect. Dis. 2013, 13, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Holt, J.G.; Krieg, N.R.; Sneath, P.H.A.; Staley, J.T.; Williams, S.T. Bergey’s Manual of Determinative Bacteriology; Williams and Wilkins: Philadelphia, PA, USA, 1994. [Google Scholar]
- Golding, C.G.; Lamboo, L.L.; Beniac, D.R.; Booth, T.F. The scanning electron microscope in microbiology and diagnosis of infectious disease. Sci. Rep. 2016, 6, 26516. [Google Scholar] [CrossRef]
- Shehadat, S.A.; Gorduysus, M.O.; Hamid, S.S.A.; Abdullah, N.A.; Samsudin, A.R.; Ahmad, A. Optimization of scanning electron microscope technique for amniotic membrane investigation: A preliminary study. Eur. J. Dent. 2018, 12, 574–578. [Google Scholar] [CrossRef]
- Brick, J.M.; Bostock, R. Silverstone Rapid in situ assay for indole acetic acid production by bacteria immobilized on nitrocellulose membrane. Appl. Environ. Microbiol. 1991, 57, 535–538. [Google Scholar] [CrossRef]
- Marques, A.P.; Pires, C.; Moreira, H.; Rangel, A.O.; Castro, P.M. Assessment of the plant growth promotion abilities of six bacterial isolates using Zea mays as indicator plant. Soil Biol. Biochem. 2010, 42, 1229–1235. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Gram Staining | Shape | Catalase | Oxidase | Glucose | Lactose | Maltose | Mannitol | D-Mannose | Sucrose | Nitrate Reduction | H2S Production | Starch Hydrolysis |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CL10 | − | Rod | + | − | + | − | + | + | − | + | − | − | + |
| C L11 | − | Rod | + | + | + | − | − | + | − | + | − | − | + |
| CL14 | − | Rod | + | + | + | − | − | − | + | − | − | − | + |
| Identified Isolate | Accession Number | Nearest Phylogenetic Neighbor Strain | Percent of Similarity |
|---|---|---|---|
| CL10 | KM067137 | Pseudomonas sp. strain BHUJPV-A13 | 100% |
| CL11 | KM067136 | Pseudomonas sp. strain DKF | 100% |
| CL14 | KM067138 | Pseudomonas fluorescens strain ESR7 | 100% |
| Identified Isolate | Indole-Acetic Acid Production | Phosphate Solubilization | Ammonia (NH3) Production | Siderophore Production | Salinity Tolerance |
|---|---|---|---|---|---|
| Pseudomonas sp. CL10 | + | + | + | + | 5% |
| Pseudomonas sp. CL11 | + | + | + | + | 5% |
| Pseudomonas fluorescence CL12 | + | + | + | + | 6% |
| Well No. in PM 12 B Plate | Compounds and Their Mode of Action | Well No. in PM 15B Plate | Compounds and Their Mode of Action | ||
|---|---|---|---|---|---|
| A1 (1–4) | Penicillin | Inhibiting cell wall | A1 (1–4) | Procaine | Inhibiting sodium influx through voltage gated sodium channels |
| A2 (5–8) | Tetracycline | Inhibit protein synthesis, prevent the association of aminoacyl-tRNA with ribosomes | A2 (5–8) | Guanidine hydrochloride | Chaotropic agent |
| A3 (9–12) | Carbenicillin | Inhibit final cell wall synthesis of susceptible bacteria. | A3 (9–12) | Cefmetazole | Inhibition of cell wall synthesis |
| B1 (1–4) | Oxacillin | Block the peptidoglycan synthesis | B1 (1–4) | D-Cycloserine | Inhibiting cell-wall biosynthesis |
| B2 (5–8) | Penimepicycline | A bactericidal action on the streptococci, and a bacteriostatic action on various gram-positive and gram-negative bacilli | B2 (5–8) | EDTA | Chelating agent |
| B3 (9–12) | Polymyxin B | Interact with lipopolysaccharide (LPS) of the outer membrane of Gram -ve bacteria | B3 (9–12) | 5,7-Dichloro-8-hydroxyquinaldine | Induces SOS-DNA repair |
| C1 (1–4) | Paromomycin | Inhibit protein synthesis via ribosomal subunits interaction | C1 (1–4) | 5,7-Dichloro-8-hydroxyquinoline | Inhibits the growth of Gram-positive and some Gram-negative bacteria |
| C2 (5–8) | Vancocin | Wall, lactam | C2 (5–8) | Fusidic acid | Interfering with bacterial protein synthesis, |
| C3 (9–12) | D,LSerin hydroxamate | tRNA synthetase | C3 (9–12) | 1,10-Phenanthroline | Inhibition of carboxypeptidase |
| D1 (1–4) | Sisomicin | Binding to small ribosomal subunit, inhibit the protein synthesis | D1 (1–4) | Phleomycin | DNA breakage |
| D2 (5–8) | Sulfamethazine | Folate antagonist, PABA analog | D2 (5–8) | Domiphen bromide | Cationic surfactant |
| D3 (9–12) | Novobiocin | DNA topoisomerase | D3 (9–12) | Nordihydroguaia retic acid | Inhibit arachidonic acid 5-lipoxygenase activity |
| E1 (1–4) | 2,4-Diamino-6,7-diisopropylpteridine | Antifolate (disruption of the metabolic pathways) | E1 (1–4) | Alexidine | Binds to lipopolysaccharide and lipoteichoic acid |
| E2 (5–8) | Sulfadiazine | Antifolate | E2 (5–8) | 5-Nitro-2- furaldehydesemicarbazone | Inhibits several bacterial enzymes |
| E3 (9–12) | Benzethonium chloride | Detergent | E3 (9–12) | Methyl viologen | Electron relay in photochemical systems |
| F1 (1–4) | Tobramycin | Protein synthesis, 30S ribosomal subunit, aminoglycoside | F1 (1–4) | 3, 4-Dimethoxybenzyl alcohol | Used as the fuel of the microbial fuel cell |
| F2 (5–8) | Sulfathiazole | Antifolate, PABA analog | F2 (5–8) | Oleandomycin | Binds to the 50s subunit of bacterial ribosomes |
| F3 (9–12) | 5-Fluoroorotic acid | Nucleic acid analog, pyrimidine | F3 (9–12) | Puromycin | Analogue of the 3′ end of aminoacyl-transfer RNA |
| G1 (1–4) | Spectinomycin | Inhibit protein synthesis with the binding of 30S ribosomal subunit, | G1 (1–4) | Carbonyl cyanide m- chlorophenylhydrazone(CCCP) | Inhibitor of oxidative phosphorylation |
| G2 (5–8) | Sulfamethoxazole | Antifolate, PABA analog | G2 (5–8) | Sodium azide | Inhibits activities of peroxidases, catalase and few more enzymes |
| G3 (9–12) | l-Aspartic acid b- hydroxamate | tRNAsynthetase | G3 (9–12) | Menadione | As a cofactor in the posttranslational gamma- carboxylation |
| H1 (1–4) | Spiramycin | Inhibits translocation by binding to bacterial 50S ribosomal subunits | H1 (1–4) | 2-Nitroimidazole | Inhibits protein synthesis and RNA synthesis |
| H2 (5–8) | Rifampicin | Inhibit bacterial DNA-dependent RNA polymerase | H2 (5–8) | Hydroxyurea | Inhibition of the enzyme ribonucleotide reductase |
| H3 (9–12) | Dodecyl trimethyl ammonium bromide | Surfactant | H3 (9–12) | Zinc chloride | Catalyst in chemical metal |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pathak, P.; Singh, M.; Naskar, A.; Singh, S.K.; Bhardwaj, N.; Kumar, A. Isolation and Phenotypic Microarray Profiling of Different Pseudomonas Strains Isolated from the Rhizosphere of Curcuma longa L. Stresses 2023, 3, 749-761. https://doi.org/10.3390/stresses3040051
Pathak P, Singh M, Naskar A, Singh SK, Bhardwaj N, Kumar A. Isolation and Phenotypic Microarray Profiling of Different Pseudomonas Strains Isolated from the Rhizosphere of Curcuma longa L. Stresses. 2023; 3(4):749-761. https://doi.org/10.3390/stresses3040051
Chicago/Turabian StylePathak, Parul, Monika Singh, Ananya Naskar, Sandeep Kumar Singh, Nikunj Bhardwaj, and Ajay Kumar. 2023. "Isolation and Phenotypic Microarray Profiling of Different Pseudomonas Strains Isolated from the Rhizosphere of Curcuma longa L." Stresses 3, no. 4: 749-761. https://doi.org/10.3390/stresses3040051
APA StylePathak, P., Singh, M., Naskar, A., Singh, S. K., Bhardwaj, N., & Kumar, A. (2023). Isolation and Phenotypic Microarray Profiling of Different Pseudomonas Strains Isolated from the Rhizosphere of Curcuma longa L. Stresses, 3(4), 749-761. https://doi.org/10.3390/stresses3040051