
Use of Apple Pomace as Substrate for Production of Lactiplantibacillus plantarum Malolactic Starter Cultures


 , , and
, , and 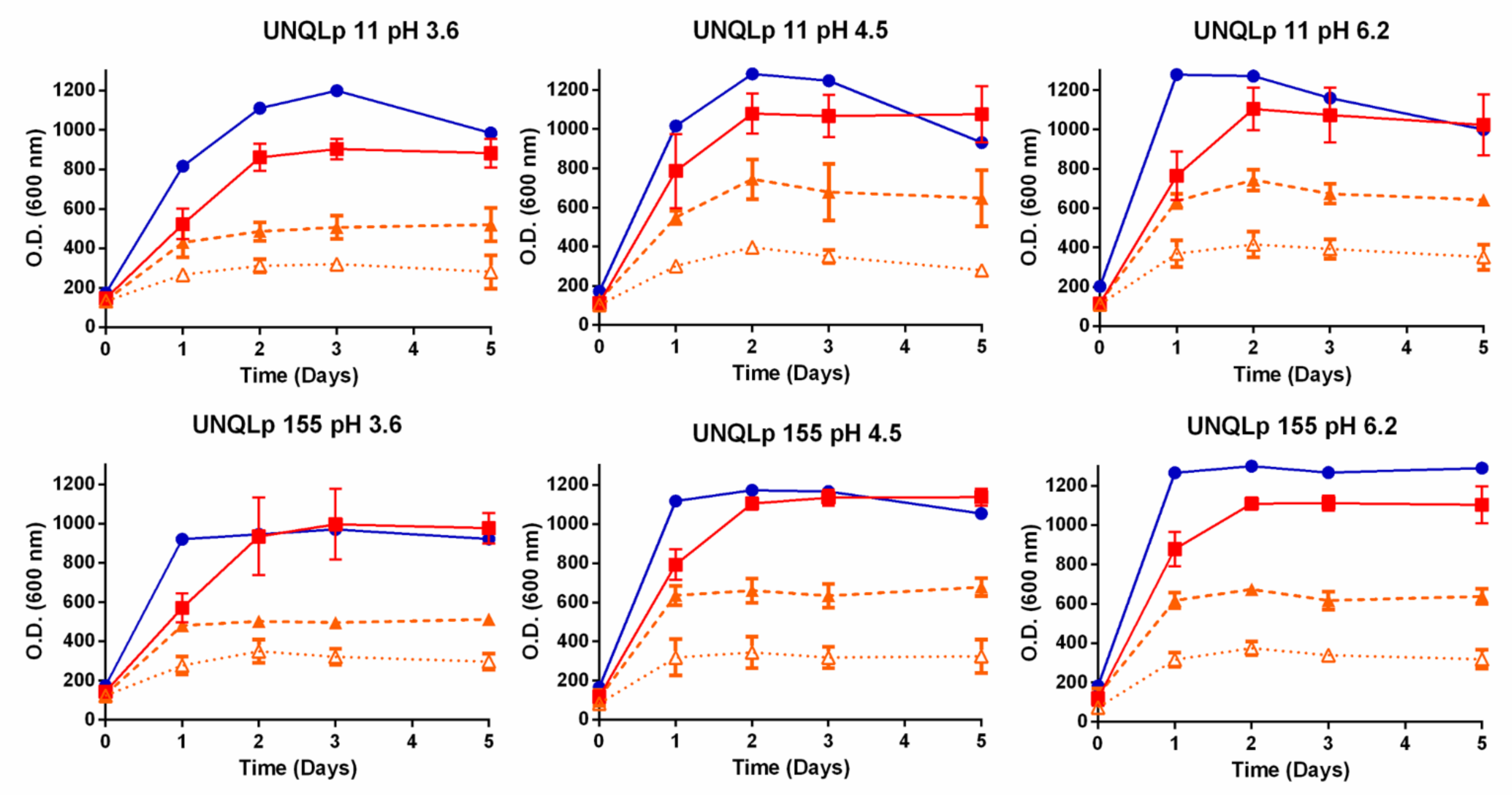{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Selection of Nutritional Supplementation
2.3. Synthetic Wine Incubation
2.4. Fermentation Assays at Laboratory Scale in Sterile Pinot Noir and Merlot Wines
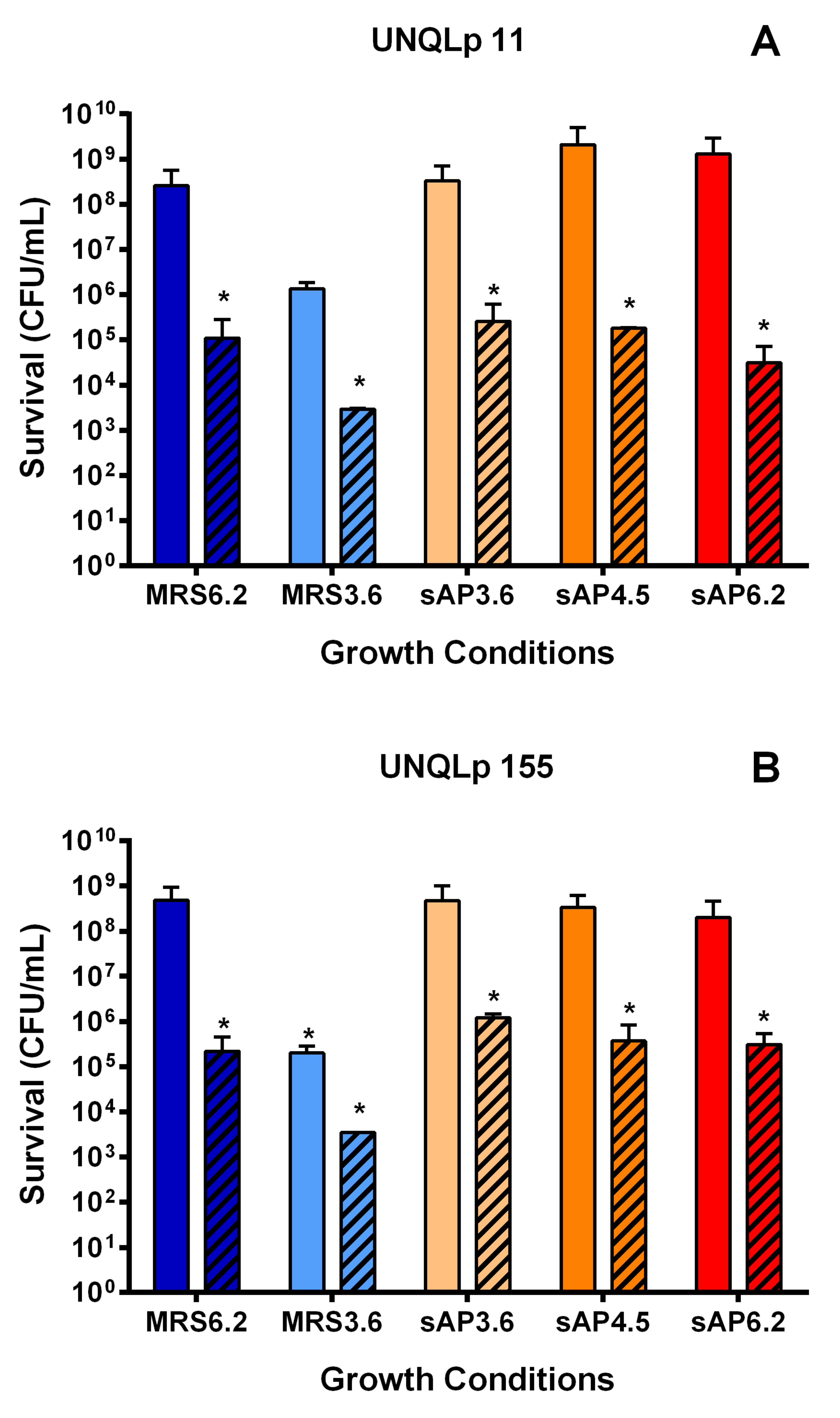
2.5. Cell Survival
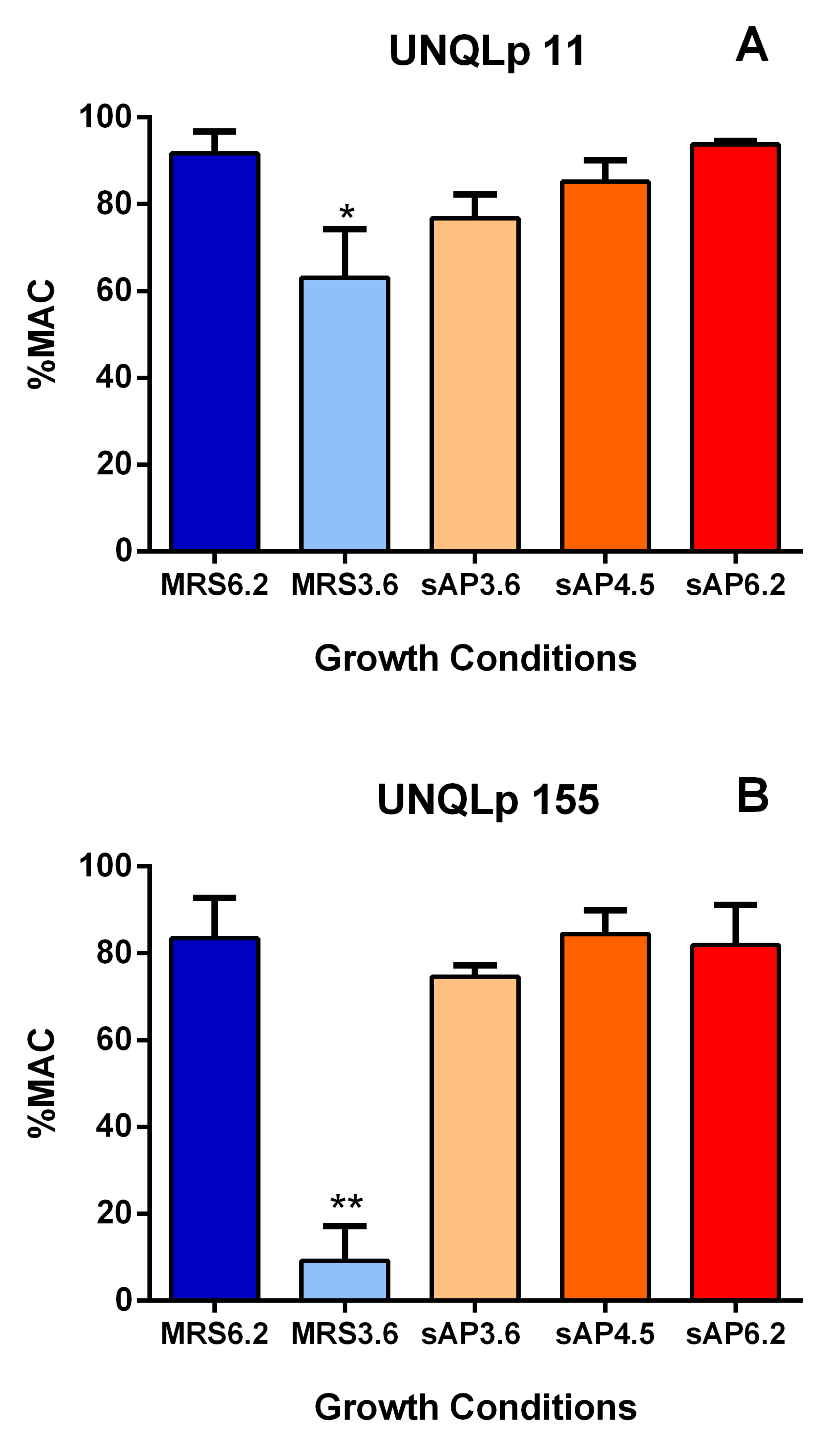
2.6. L-Malic Acid Consumption
2.7. Statistical Analysis
3. Results
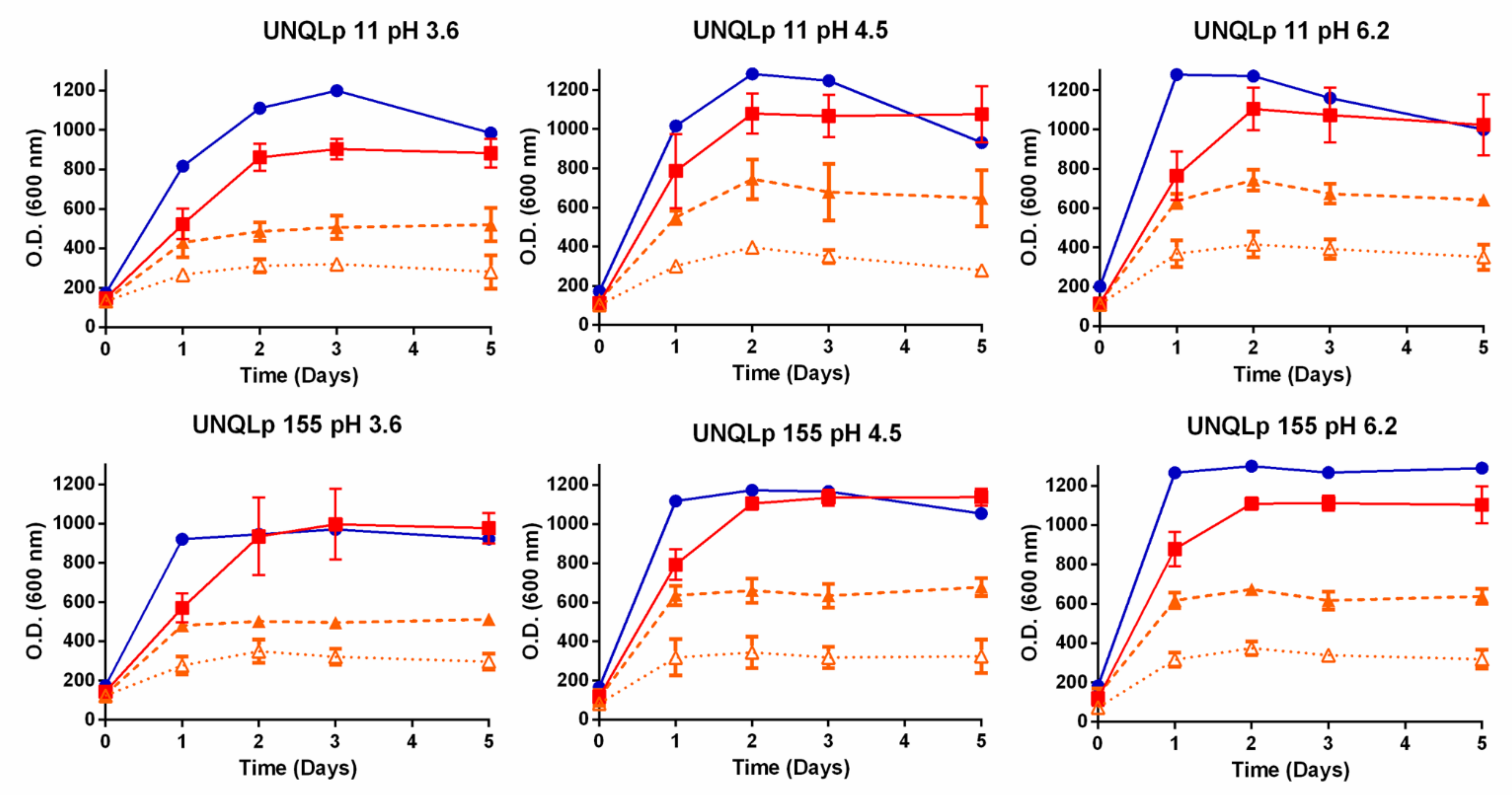
3.1. Growth Kinetics in AP and sAP
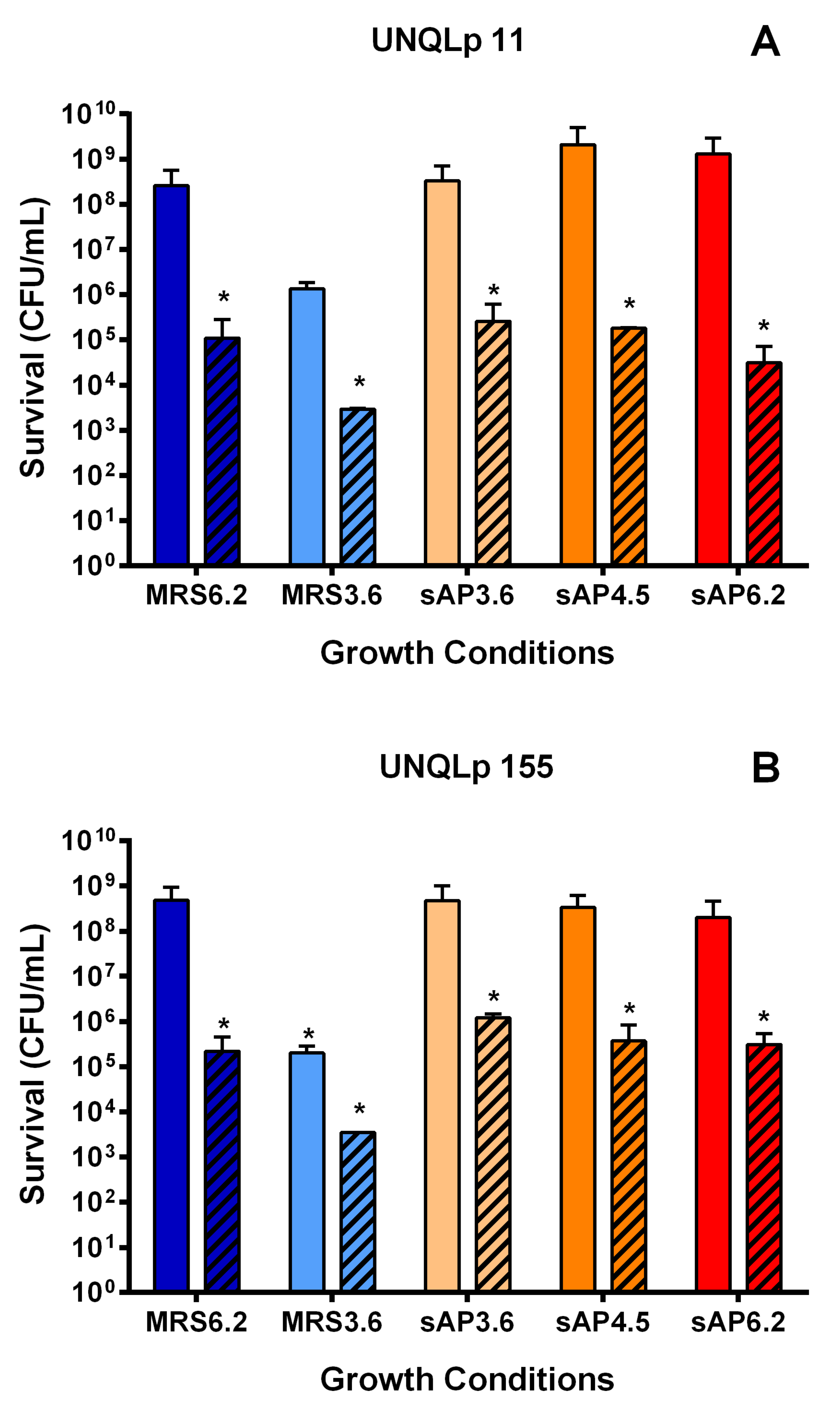
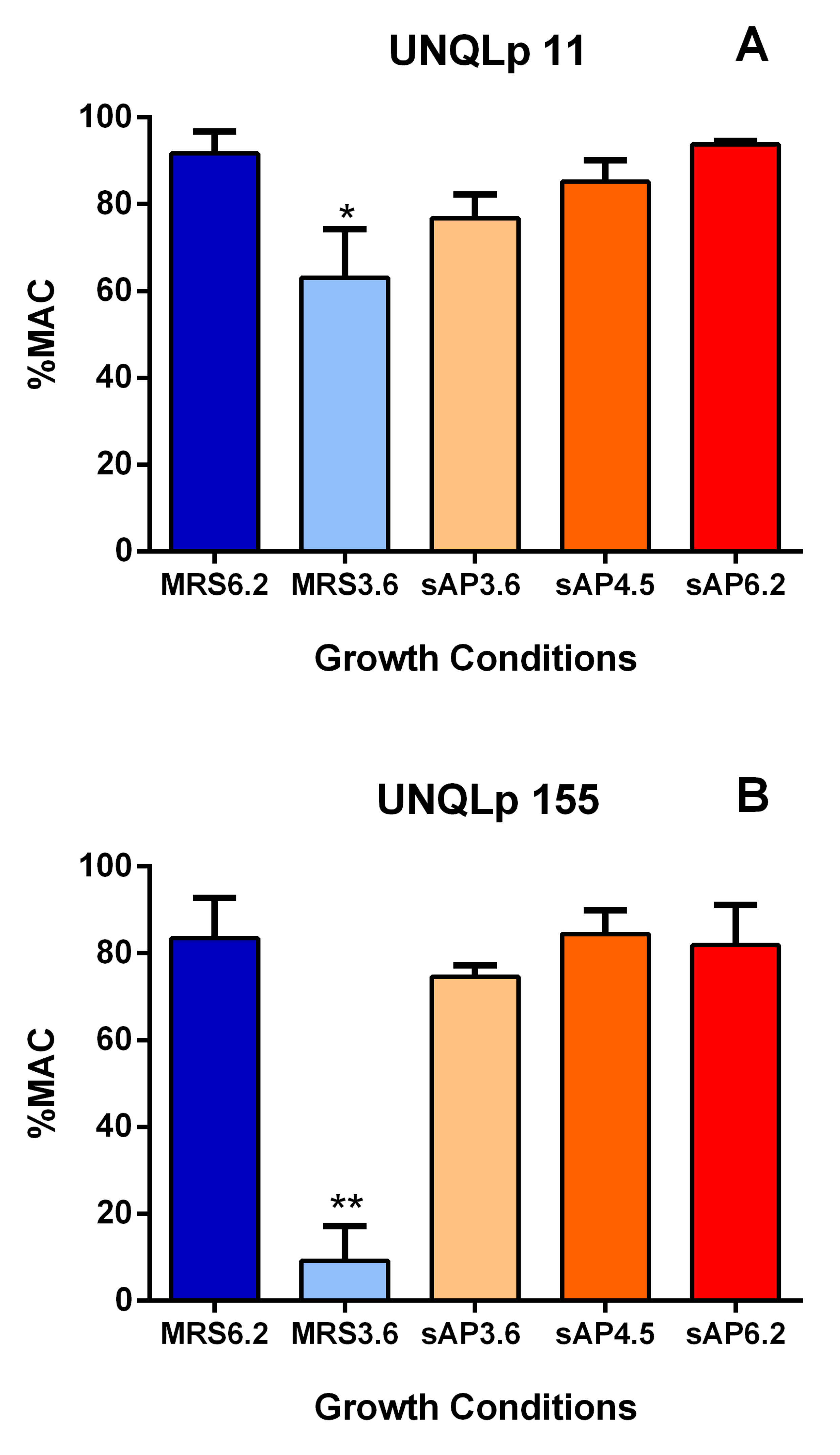
3.2. Behavior of Bacterial Cultures in Synthetic Wine
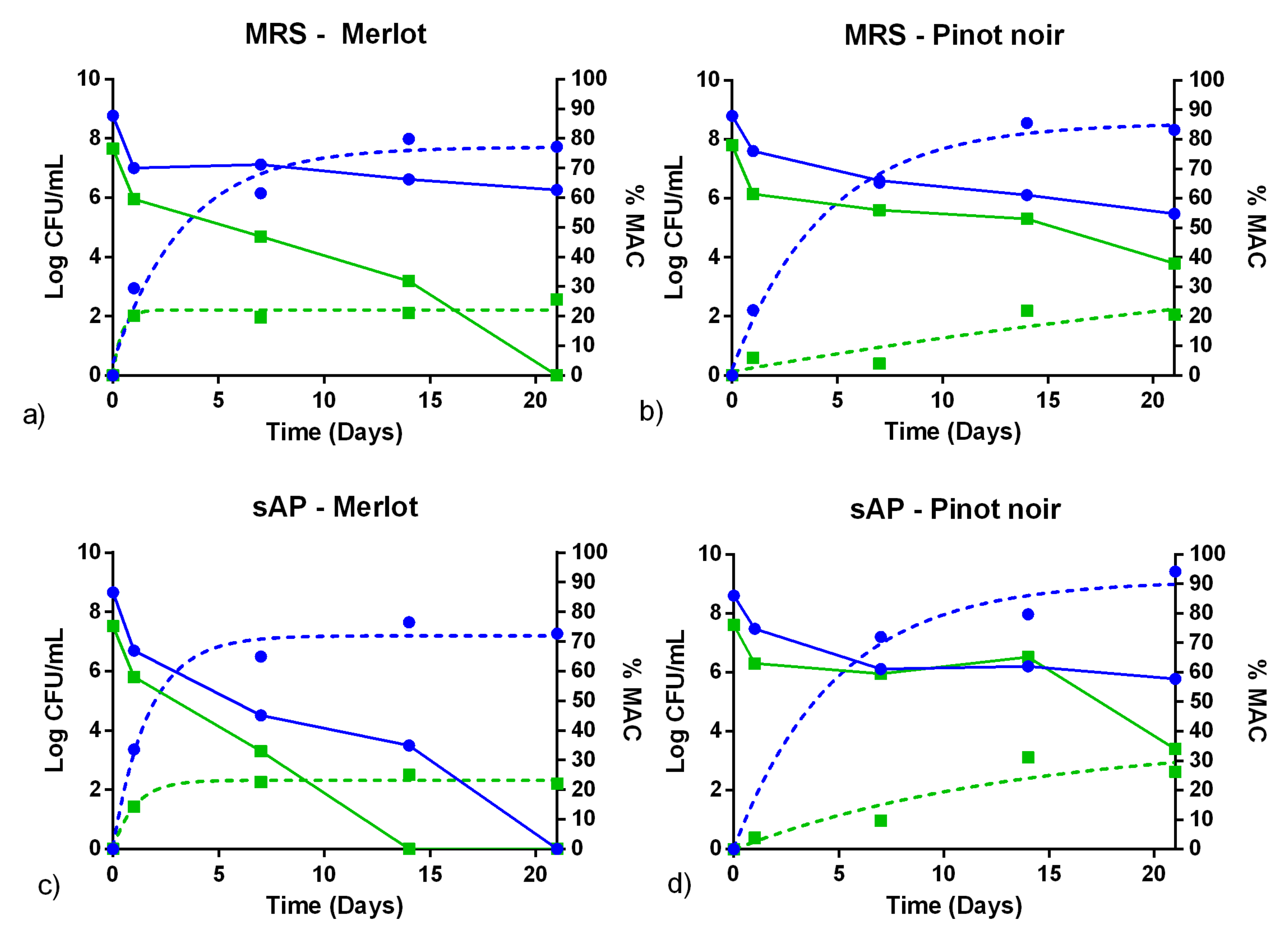
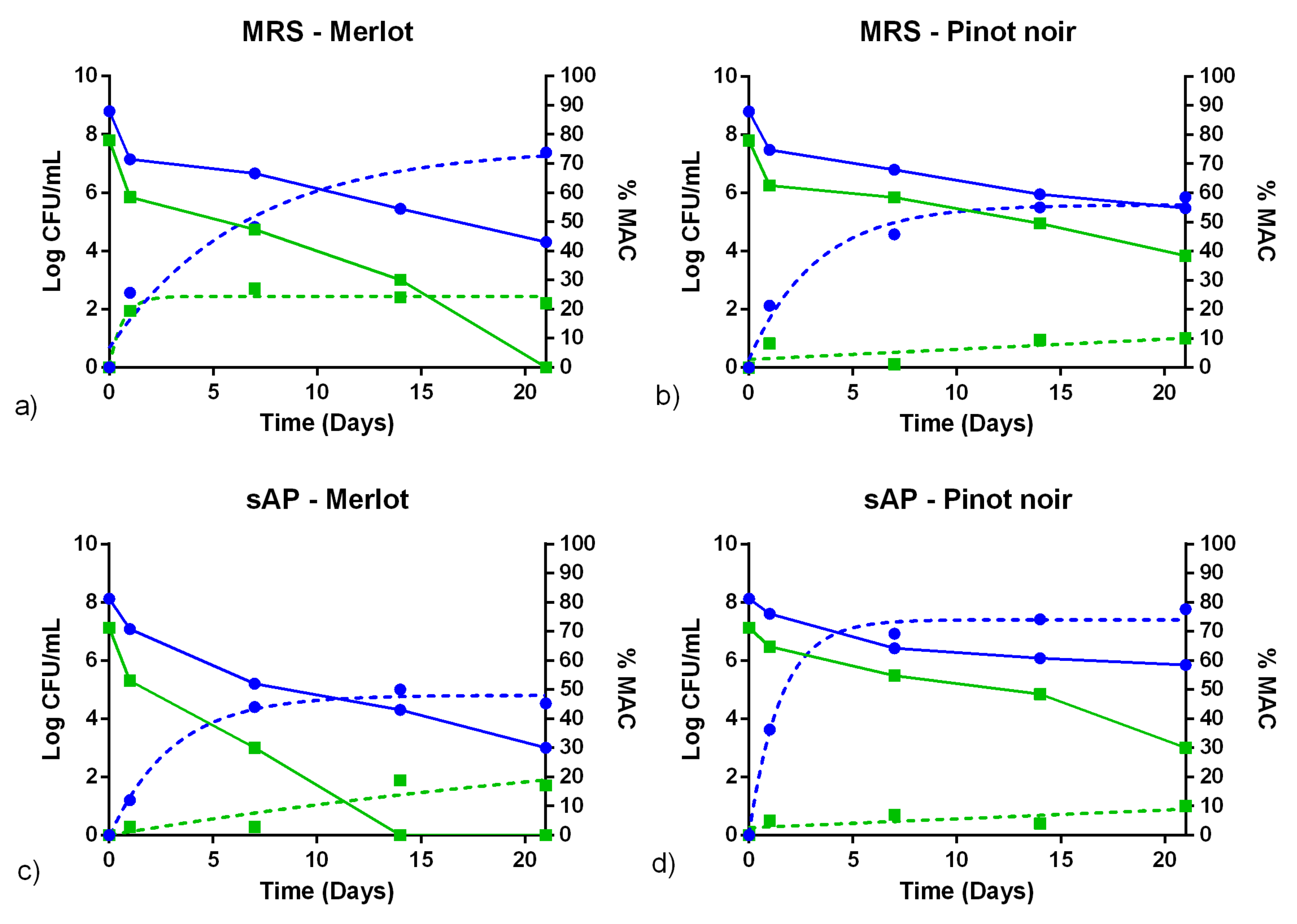
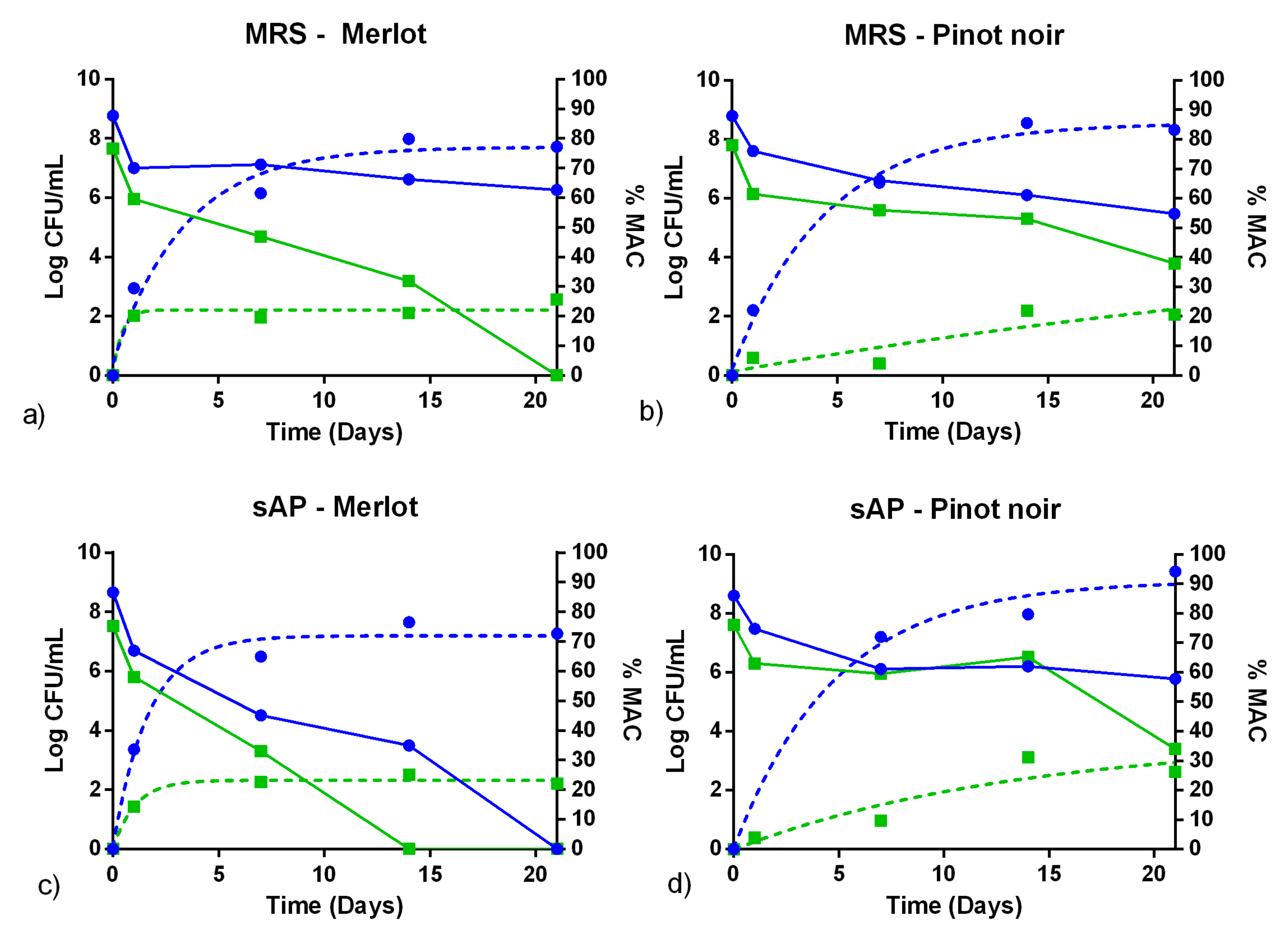
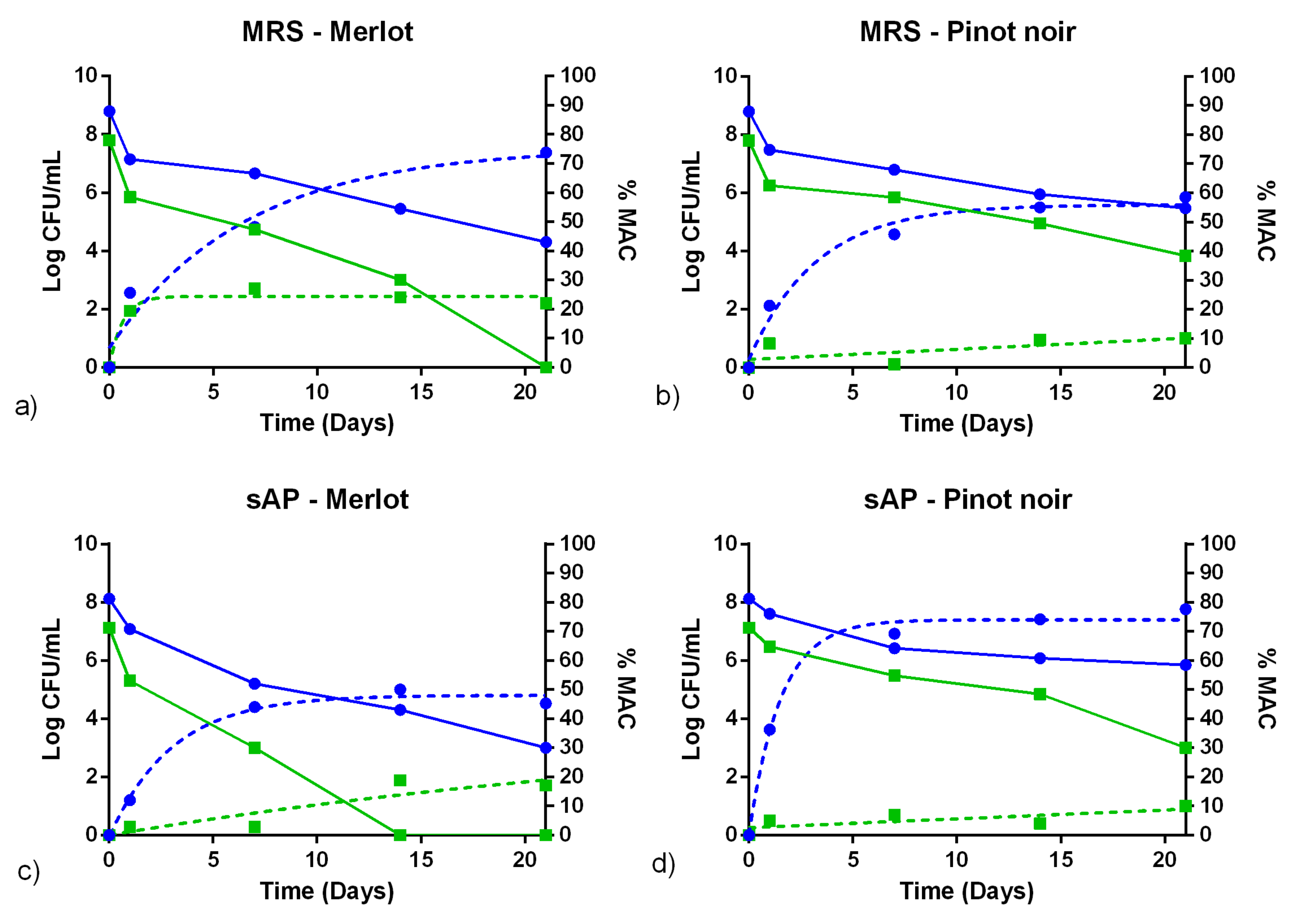
3.3. Fermentation Assays in Sterile Wine
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wibowo, D.; Eschenbruch, R.; Davis, C.R.; Fleet, G.H.; Lee, T. Occurrence and growth of lactic acid bacteria in wine: A review. Am. J. Enol. Vitic. 1985, 36, 302–313. [Google Scholar]
- Lonvaud-Funel, A. Lactic acid bacteria in the quality improvement and depreciation of wine. In Lactic Acid Bacteria: Genetics, Metabolism and Applications; Springer: Dordrecht, The Netherlands, 1999; pp. 317–331. [Google Scholar]
- Bartowsky, E.J.; Henschke, P.A. Use of a polymerase chain reaction for specific detection of the malolactic fermentation bacterium Oenococcus oeni (formerly Leuconostoc oenos) in grape juice and wine samples. Aust. J. Grape Wine Res. 1999, 5, 39–44. [Google Scholar] [CrossRef]
- Pozo-Bayón, M.A.; G-Alegría, E.; Polo, M.C.; Tenorio, C.; Martín-Álvarez, P.J.; Calvo De La Banda, M.T.; Ruiz-Larrea, F.; Moreno-Arribas, M.V. Wine volatile and amino acid composition after malolactic fermentation: Effect of Oenococcus oeni and Lactobacillus plantarum starter cultures. J. Agric. Food Chem. 2005, 53, 8729–8735. [Google Scholar] [CrossRef]
- Valdés La Hens, D.; Bravo-Ferrada, B.M.; Delfederico, L.; Caballero, A.C.; Semorile, L.C. Prevalence of Lactobacillus plantarum and Oenococcus oeni during spontaneous malolactic fermentation in Patagonian red wines revealed by polymerase chain reaction-denaturing gradient gel electrophoresis with two targeted gene. Aust. J. Grape Wine Res. 2015, 21, 49–56. [Google Scholar] [CrossRef]
- Lerm, E.; Engelbrecht, L.; du Toit, M. Selection and characterisation of Oenococcus oeni and lactobacillus plantarum South African wine isolates for use as malolactic fermentation starter cultures. S. Afr. J. Enol. Vitic. 2011, 32, 280–295. [Google Scholar] [CrossRef] [Green Version]
- Brizuela, N.S.; Bravo-Ferrada, B.M.; Pozo-Bayón, M.Á.; Semorile, L.; Tymczyszyn, E.E. Changes in the volatile profile of Pinot noir wines caused by Patagonian Lactobacillus plantarum and Oenococcus oeni strains. Food Res. Int. 2018, 106, 22–28. [Google Scholar] [CrossRef]
- Brizuela, N.S.; Bravo-Ferrada, B.M.; Curilén, Y.; Delfederico, L.; Caballero, A.; Semorile, L.; Pozo-Bayón, M.A.; Tymczyszyn, E.E. Advantages of Using Blend Cultures of Native L. plantarum and O. oeni Strains to Induce Malolactic Fermentation of Patagonian Malbec Wine. Front. Microbiol. 2018, 9, 2109. [Google Scholar] [CrossRef] [Green Version]
- Brizuela, N.S.; Franco-Luesma, E.; Bravo-Ferrada, B.M.; Pérez-Jiménez, M.; Semorile, L.; Tymczyszyn, E.E.; Pozo-Bayon, M.A. Influence of Patagonian Lactiplantibacillus plantarum and Oenococcus oeni strains on sensory perception of Pinot Noir wine after malolactic fermentation. Aust. J. Grape Wine Res. 2021, 27, 118–127. [Google Scholar] [CrossRef]
- Sumby, K.M.; Bartle, L.; Grbin, P.R.; Jiranek, V. Measures to improve wine malolactic fermentation. Appl. Microbiol. Biotechnol. 2019, 103, 2033–2051. [Google Scholar] [CrossRef]
- Lopez, I.; Lòpez, R.; Santamaría, P.; Torres, C.; Ruiz-Larrea, F. Performance of malolactic fermentation by inoculation of selected Lactobacillus plantarum and Oenococcus oeni strains isolated from Rioja red wines. Vitis 2008, 47, 123–129. [Google Scholar]
- Du Toit, M.; Engelbrecht, L.; Lerm, E.; Krieger-Weber, S. Lactobacillus: The Next Generation of Malolactic Fermentation Starter Cultures—An Overview. Food Bioprocess Technol. 2011, 4, 876–906. [Google Scholar] [CrossRef]
- Garofalo, C.; El Khoury, M.; Lucas, P.; Bely, M.; Russo, P.; Spano, G.; Capozzi, V. Autochthonous starter cultures and indigenous grape variety for regional wine production. J. Appl. Microbiol. 2015, 118, 1395–1408. [Google Scholar] [CrossRef]
- Berbegal, C.; Peña, N.; Russo, P.; Grieco, F.; Pardo, I.; Ferrer, S.; Spano, G.; Capozzi, V. Technological properties of Lactobacillus plantarum strains isolated from grape must fermentation. Food Microbiol. 2016, 57, 187–194. [Google Scholar] [CrossRef]
- G-Alegría, E.; López, I.; Ruiz, J.I.; Sáenz, J.; Fernández, E.; Zarazaga, M.; Dizy, M.; Torres, C.; Ruiz-Larrea, F. High tolerance of wild Lactobacillus plantarum and Oenococcus oeni strains to lyophilisation and stress environmental conditions of acid pH and ethanol. FEMS Microbiol. Lett. 2004, 230, 53–61. [Google Scholar] [CrossRef]
- Spano, G.; Massa, S. Environmental Stress Response in Wine Lactic Acid Bacteria: Beyond Bacillus subtilis. Crit. Rev. Microbiol. 2006, 32, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Ferrada, B.M.; Hollmann, A.; Delfederico, L.; Valdés La Hens, D.; Caballero, A.; Semorile, L. Patagonian red wines: Selection of Lactobacillus plantarum isolates as potential starter cultures for malolactic fermentation. World J. Microbiol. Biotechnol. 2013, 29, 1537–1549. [Google Scholar] [CrossRef]
- Brizuela, N.S.; Bravo-Ferrada, B.M.; La Hens, D.V.; Hollmann, A.; Delfederico, L.; Caballero, A.; Tymczyszyn, E.E.; Semorile, L. Comparative vinification assays with selected Patagonian strains of Oenococcus oeni and Lactobacillus plantarum. LWT 2017, 77, 348–355. [Google Scholar] [CrossRef] [Green Version]
- Brizuela, N.S.; Arnez-Arancibia, M.; Semorile, L.; Pozo-Bayón, M.Á.; Bravo-Ferrada, B.M.; Tymczyszyn, E.E. β-Glucosidase Activity of Lactiplantibacillus plantarum UNQLp 11 in Different Malolactic Fermentations Conditions: Effect of pH and Ethanol Content. Fermentation 2021, 7, 22. [Google Scholar] [CrossRef]
- Cerdeira, V.; Bravo-Ferrada, B.M.; Semorile, L.; Tymczyszyn, E. Design of a low-cost culture medium based in whey permeate for biomass production of enological Lactobacillus plantarum strains. Biotechnol. Prog. 2019, 35, e2791. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Ferrada, B.M.; Tymczyszyn, E.E.; Gómez-Zavaglia, A.; Semorile, L. Effect of acclimation medium on cell viability, membrane integrity and ability to consume malic acid in synthetic wine by oenological Lactobacillus plantarum strains. J. Appl. Microbiol. 2014, 116, 360–367. [Google Scholar] [CrossRef]
- Brizuela, N.S.; Arnez-Arancibia, M.; Semorile, L.; Bravo-Ferrada, B.M.; Tymczyszyn, E.E. Whey permeate as a substrate for the production of freeze-dried Lactiplantibacillus plantarum to be used as a malolactic starter culture. World J. Microbiol. Biotechnol. 2021, 37, 115. [Google Scholar] [CrossRef] [PubMed]
- Bravo, S.M.E.; Morales, M.; del Mónaco, S.M.; Caballero, A.C. Apple bagasse as a substrate for the propagation of Patagonian wine yeast biomass. J. Appl. Microbiol. 2019, 126, 1414–1425. [Google Scholar] [CrossRef]
- Kieliszek, M.; Piwowarek, K.; Kot, A.M.; Pobiega, K. The aspects of microbial biomass use in the utilization of selected waste from the agro-food industry. Open Life Sci. 2020, 15, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Lyu, F.; Luiz, S.F.; Azeredo, D.R.P.; Cruz, A.G.; Ajlouni, S.; Ranadheera, C.S. Apple Pomace as a Functional and Healthy Ingredient in Food Products: A Review. Processes 2020, 8, 319. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.; Tymczyszyn, E.; Golowczyc, M.; Mobili, P.; Gomez-Zavaglia, A. Probiotic cell cultivation. In Advances in Probiotic Technology; Foerst, P., Santivarangkna, C., Eds.; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Succi, M.; Pannella, G.; Tremonte, P.; Tipaldi, L.; Coppola, R.; Iorizzo, M.; Lombardi, S.J.; Sorrentino, E. Sub-optimal pH Preadaptation Improves the Survival of Lactobacillus plantarum Strains and the Malic Acid Consumption in Wine-Like Medium. Front. Microbiol. 2017, 0, 470. [Google Scholar] [CrossRef] [PubMed]
- De Man, J.C.; Rogosa, M.; Sharpe, M.E. A Medium for the Cultivation of Lactobacilli. J. Appl. Bacteriol. 1960, 23, 130–135. [Google Scholar] [CrossRef]
- Hayek, S.A.; Ibrahim, S.A. Current Limitations and Challenges with Lactic Acid Bacteria: A Review. Food Nutr. Sci. 2013, 4, 73–87. [Google Scholar] [CrossRef] [Green Version]
- Benaissa, M.; And, H.Z.-K.; Karam, N.-E. Development of a sweet whey-based medium for culture of Lactobacillus. Afr. J. Biotechnol. 2017, 16, 1630–1637. [Google Scholar] [CrossRef] [Green Version]
- Brizuela, N.; Tymczyszyn, E.E.; Semorile, L.C.; Valdes La Hens, D.; Delfederico, L.; Hollmann, A.; Bravo-Ferrada, B. Lactobacillus plantarum as a malolactic starter culture in winemaking: A new (old) player? Electron. J. Biotechnol. 2019, 38, 10–18. [Google Scholar] [CrossRef]
- Sfara, V.; Fonovich, T.M.; Silvia, C.; Coll, P.; Sfara, V.; Fonovich, T.; Pérez-Coll, C. Sustainable pome fruit production in Patagonia, Argentina. Technol. Acta 2016, 9, 57–61. [Google Scholar]
- Champagne, C.P.; Gardner, N.; Doyon, G. Production of Leuconostoc oenos biomass under pH control. Appl. Environ. Microbiol. 1989, 55, 2488–2492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berbegal, C.; Benavent-Gil, Y.; Pardo, I.; Ferrer, S. A novel culture medium for Oenococcus oeni malolactic starter production. LWT-Food Sci. Technol. 2015, 64, 25–31. [Google Scholar] [CrossRef]
- Maicas, S.; Pardo, I.; Ferrer, S. Continuous malolactic fermentation in red wine using free Oenococcus oeni. World J. Microbiol. Biotechnol. 1999, 15, 737–739. [Google Scholar] [CrossRef]
- Thuy, P.T.; Huong, N.L.; Son, C.K. Using Malolactic Fermentation with Acid- and Ethanol-Adapted Oenococcus Oeni Strain to Improve the Quality of Wine from Champs Bourcin Grape in Sapa–Lao Cai. World Acad. Sci. Eng. Technol. 2011, 5, 655–658. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cerdeira, V.; Brizuela, N.S.; Bravo, S.M.E.; Bravo-Ferrada, B.M.; Valdés La Hens, D.; Caballero, A.C.; Semorile, L.C.; Tymczyszyn, E.E. Use of Apple Pomace as Substrate for Production of Lactiplantibacillus plantarum Malolactic Starter Cultures. Fermentation 2021, 7, 244. https://doi.org/10.3390/fermentation7040244
Cerdeira V, Brizuela NS, Bravo SME, Bravo-Ferrada BM, Valdés La Hens D, Caballero AC, Semorile LC, Tymczyszyn EE. Use of Apple Pomace as Substrate for Production of Lactiplantibacillus plantarum Malolactic Starter Cultures. Fermentation. 2021; 7(4):244. https://doi.org/10.3390/fermentation7040244
Chicago/Turabian StyleCerdeira, Victoria, Natalia S. Brizuela, Sebastián M. E. Bravo, Bárbara M. Bravo-Ferrada, Danay Valdés La Hens, Adriana C. Caballero, Liliana C. Semorile, and E. Elizabeth Tymczyszyn. 2021. "Use of Apple Pomace as Substrate for Production of Lactiplantibacillus plantarum Malolactic Starter Cultures" Fermentation 7, no. 4: 244. https://doi.org/10.3390/fermentation7040244
APA StyleCerdeira, V., Brizuela, N. S., Bravo, S. M. E., Bravo-Ferrada, B. M., Valdés La Hens, D., Caballero, A. C., Semorile, L. C., & Tymczyszyn, E. E. (2021). Use of Apple Pomace as Substrate for Production of Lactiplantibacillus plantarum Malolactic Starter Cultures. Fermentation, 7(4), 244. https://doi.org/10.3390/fermentation7040244