
Different Gene Expression Patterns of Hexose Transporter Genes Modulate Fermentation Performance of Four Saccharomyces cerevisiae Strains



Abstract
1. Introduction
2. Materials and Methods
2.1. Yeast Strains
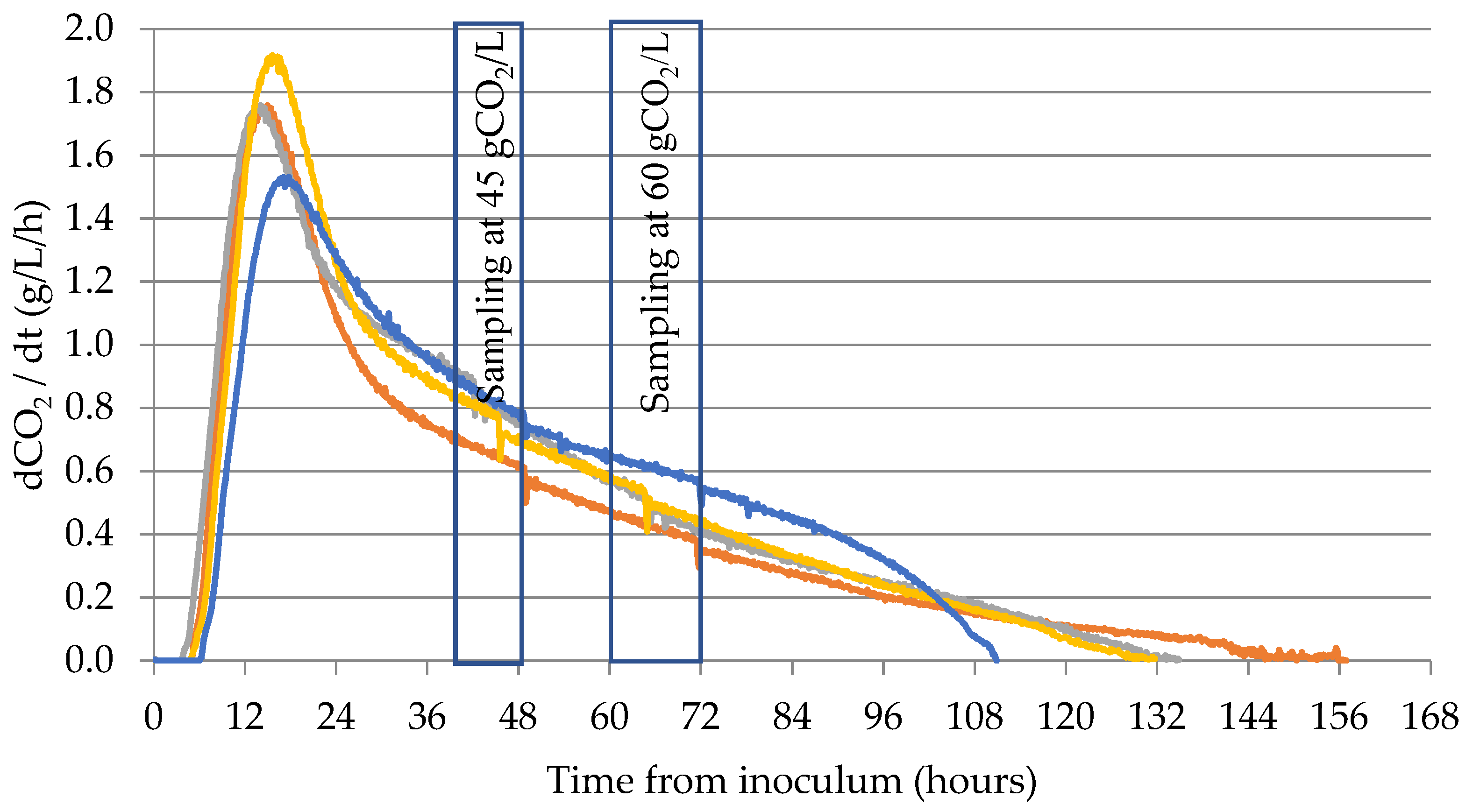
2.2. Fermentation Trial and Samplings
2.3. Chemical Analysis
2.4. RNA Extraction and Reverse Transcription
2.5. Primer Design
2.6. Real-Time PCR
2.7. Statistical Analysis
3. Results
3.1. Yeasts Fermentation Process
3.2. Expression of Hexose Transporters Genes during Late Fermentation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guillaume, C.; Delobel, P.; Sablayrolles, J.-M.; Blondin, B. Molecular Basis of Fructose Utilization by the Wine Yeast Saccharomyces cerevisiae: A Mutated Hxt3 Allele Enhances Fructose Fermentation. Appl. Environ. Microbiol. 2007, 73, 2432–2439. [Google Scholar] [CrossRef]
- Berthels, N.; Corderootero, R.; Bauer, F.; Thevelein, J.; Pretorius, I. Discrepancy in Glucose and Fructose Utilisation during Fermentation by Wine Yeast Strains. FEMS Yeast Res. 2004, 4, 683–689. [Google Scholar] [CrossRef]
- Galeote, V.; Novo, M.; Salema-Oom, M.; Brion, C.; Valério, E.; Gonçalves, P.; Dequin, S. FSY1, a Horizontally Transferred Gene in the Saccharomyces cerevisiae EC1118 Wine Yeast Strain, Encodes a High-Affinity Fructose/H+ Symporter. Microbiology 2010, 156, 3754–3761. [Google Scholar] [CrossRef]
- Blateyron, L.; Sablayrolles, J.M. Stuck and Slow Fermentations in Enology: Statistical Study of Causes and Effectiveness of Combined Additions of Oxygen and Diammonium Phosphate. J. Biosci. Bioeng. 2001, 91, 184–189. [Google Scholar] [CrossRef]
- Anjos, J.; Rodrigues de Sousa, H.; Roca, C.; Cássio, F.; Luttik, M.; Pronk, J.T.; Salema-Oom, M.; Gonçalves, P. Fsy1, the Sole Hexose-Proton Transporter Characterized in Saccharomyces Yeasts, Exhibits a Variable Fructose:H+ Stoichiometry. Biochim. Biophys. Acta Biomembr. 2013, 1828, 201–207. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, H.R.; Spencer-Martins, I.; Gonçalves, P. Differential Regulation by Glucose and Fructose of a Gene Encoding a Specific Fructose/H+ Symporter In Saccharomyces Sensu Stricto Yeasts. Yeast 2004, 21, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Luyten, K.; Riou, C.; Blondin, B. The Hexose Transporters of Saccharomyces cerevisiae Play Different Roles during Enological Fermentation. Yeast 2002, 19, 713–726. [Google Scholar] [CrossRef]
- Boles, E.; Hollenberg, C.P. The Molecular Genetics of Hexose Transport in Yeasts. FEMS Microbiol. Rev. 1997, 21, 85–111. [Google Scholar] [CrossRef]
- Kim, D.; Song, J.-Y.; Hahn, J.-S. Improvement of Glucose Uptake Rate and Production of Target Chemicals by Overexpressing Hexose Transporters and Transcriptional Activator Gcr1 in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2015, 81, 8392–8401. [Google Scholar] [CrossRef]
- Novo, M.; Bigey, F.; Beyne, E.; Galeote, V.; Gavory, F.; Mallet, S.; Cambon, B.; Legras, J.-L.; Wincker, P.; Casaregola, S.; et al. Eukaryote-to-Eukaryote Gene Transfer Events Revealed by the Genome Sequence of the Wine Yeast Saccharomyces cerevisiae EC1118. Proc. Natl. Acad. Sci. USA 2009, 106, 16333–16338. [Google Scholar] [CrossRef]
- Treu, L.; Toniolo, C.; Nadai, C.; Sardu, A.; Giacomini, A.; Corich, V.; Campanaro, S. The Impact of Genomic Variability on Gene Expression in Environmental Saccharomyces cerevisiae Strains: Genome Analysis of Environmental Yeast Strains. Environ. Microbiol. 2014, 16, 1378–1397. [Google Scholar] [CrossRef]
- Marsit, S.; Mena, A.; Bigey, F.; Sauvage, F.X.; Couloux, A.; Guy, J.; Legras, J.L.; Barrio, E.; Dequin, S.; Galeote, V. Evolutionary Advantage Conferred by an Eukaryote-to-Eukaryote Gene Transfer Event in Wine Yeasts. Mol. Biol. Evol. 2015, 32, 1695–1707. [Google Scholar] [CrossRef]
- Devia, J.; Bastías, C.; Kessi-Pérez, E.I.; Villarroel, C.A.; De Chiara, M.; Cubillos, F.A.; Liti, G.; Martínez, C.; Salinas, F. Transcriptional Activity and Protein Levels of Horizontally Acquired Genes in Yeast Reveal Hallmarks of Adaptation to Fermentative Environments. Front. Genet. 2020, 11, 293. [Google Scholar] [CrossRef] [PubMed]
- Alexandre, H.; Ansanay-Galeote, V.; Dequin, S.; Blondin, B. Global Gene Expression during Short-Term Ethanol Stress in Saccharomyces cerevisiae. FEBS Lett. 2001, 498, 98–103. [Google Scholar] [CrossRef]
- Boer, V.M.; de Winde, J.H.; Pronk, J.T.; Piper, M.D.W. The Genome-Wide Transcriptional Responses of Saccharomyces cerevisiae Grown on Glucose in Aerobic Chemostat Cultures Limited for Carbon, Nitrogen, Phosphorus, or Sulfur. J. Biol. Chem. 2003, 278, 3265–3274. [Google Scholar] [CrossRef] [PubMed]
- Gibson, B.R.; Boulton, C.A.; Box, W.G.; Graham, N.S.; Lawrence, S.J.; Linforth, R.S.T.; Smart, K.A. Carbohydrate Utilization and the Lager Yeast Transcriptome during Brewery Fermentation. Yeast 2008, 25, 549–562. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, N.; Hayes, A.; Panoutsopoulou, K.; Oliver, S.G. Global Analysis of Nutrient Control of Gene Expression in Saccharomyces cerevisiae during Growth and Starvation. Proc. Nat. Acad. Sci. USA 2004, 101, 3148–3153. [Google Scholar] [CrossRef] [PubMed]
- Bely, M.; Sablayrolles, J.; Barre, P. Description of Alcoholic Fermentation Kinetics: Its Variability and Significance. Am. J. Enol. Viticult. 1990, 40, 319–324. [Google Scholar]
- Lemos Junior, W.J.F.; Nadai, C.; Crepalde, L.T.; de Oliveira, V.S.; de Matos, A.D.; Giacomini, A.; Corich, V. Potential Use of Starmerella bacillaris as Fermentation Starter for the Production of Low-Alcohol Beverages Obtained from Unripe Grapes. Int. J. Food Microbiol. 2019, 303, 1–8. [Google Scholar] [CrossRef]
- Teste, M.-A.; Duquenne, M.; François, J.M.; Parrou, J.-L. Validation of Reference Genes for Quantitative Expression Analysis by Real-Time RT-PCR in Saccharomyces cerevisiae. BMC Mol. Biol. 2009, 10, 99. [Google Scholar] [CrossRef]
- Cankorur-Cetinkaya, A.; Dereli, E.; Eraslan, S.; Karabekmez, E.; Dikicioglu, D.; Kirdar, B. A Novel Strategy for Selection and Validation of Reference Genes in Dynamic Multidimensional Experimental Design in Yeast. PLoS ONE 2012, 7, e38351. [Google Scholar] [CrossRef]
- Nadai, C.; Campanaro, S.; Giacomini, A.; Corich, V. Selection and Validation of Reference Genes for Quantitative Real-Time PCR Studies during Saccharomyces cerevisiae Alcoholic Fermentation in the Presence of Sulfite. Int. J. Food Microbiol. 2015, 215, 49–56. [Google Scholar] [CrossRef]
- Rossignol, T.; Dulau, L.; Julien, A.; Blondin, B. Genome-Wide Monitoring of Wine Yeast Gene Expression during Alcoholic Fermentation. Yeast 2003, 20, 1369–1385. [Google Scholar] [CrossRef] [PubMed]
- Vendramini, C.; Beltran, G.; Nadai, C.; Giacomini, A.; Mas, A.; Corich, V. The Role of Nitrogen Uptake on the Competition Ability of Three Vineyard Saccharomyces cerevisiae Strains. Int. J. Food Microbiol. 2017, 258, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zaman, S.; Lippman, S.I.; Schneper, L.; Slonim, N.; Broach, J.R. Glucose Regulates Transcription in Yeast through a Network of Signaling Pathways. Mol. Syst. Biol. 2009, 5, 245. [Google Scholar] [CrossRef] [PubMed]
- Perez, M.; Luyten, K.; Michel, R.; Riou, C.; Blondin, B. Analysis of Hexose Carrier Expression during Wine Fermentation: Both Low- and High-Affinity Hxt Transporters Are Expressed. FEMS Yeast Res. 2005, 5, 351–361. [Google Scholar] [CrossRef] [PubMed]
- Reifenberger, E.; Boles, E.; Ciriacy, M. Kinetic Characterization of Individual Hexose Transporters of Saccharomyces cerevisiae and Their Relation to the Triggering Mechanisms of Glucose Repression. Eur. J. Biochem. 1997, 245, 324–333. [Google Scholar] [CrossRef]
- Horák, J. Regulations of Sugar Transporters: Insights from Yeast. Curr. Genet. 2013, 59, 1–31. [Google Scholar] [CrossRef]
- Leandro, M.J.; Sychrová, H.; Prista, C.; Loureiro-Dias, M.C. ZrFsy1, a High-Affinity Fructose/H+ Symporter from Fructophilic Yeast Zygosaccharomyces rouxii. PLoS ONE 2013, 8, e68165. [Google Scholar] [CrossRef]
- Leandro, M.J.; Fonseca, C.; Gonçalves, P. Hexose and Pentose Transport in Ascomycetous Yeasts: An Overview. FEMS Yeast Res. 2009, 9, 511–525. [Google Scholar] [CrossRef] [PubMed]
- Wieczorke, R.; Krampe, S.; Weierstall, T.; Freidel, K.; Hollenberg, C.P.; Boles, E. Concurrent Knock-out of at Least 20 Transporter Genes Is Required to Block Uptake of Hexoses in Saccharomyces cerevisiae. FEBS Lett. 1999, 464, 123–128. [Google Scholar] [CrossRef]
- Özcan, S.; Johnston, M. Function and Regulation of Yeast Hexose Transporters. Microbiol. Mol. Biol. Rev. 1999, 63, 554–569. [Google Scholar] [CrossRef] [PubMed]
- Diderich, J.A.; Merijn Schuurmans, J.; Van Gaalen, M.C.; Kruckeberg, A.L.; Van Dam, K. Functional Analysis of the Hexose Transporter Homologue HXT5 in Saccharomyces cerevisiae. Yeast 2001, 18, 1515–1524. [Google Scholar] [CrossRef] [PubMed]
- Verwaal, R.; Paalman, J.W.G.; Hogenkamp, A.; Verkleij, A.J.; Verrips, C.T.; Boonstra, J. HXT5 Expression is Determined by Growth Rates in Saccharomyces cerevisiae. Yeast 2002, 19, 1029–1038. [Google Scholar] [CrossRef] [PubMed]
- Buziol, S.; Becker, J.; Baumeister, A.; Jung, S.; Mauch, K.; Reuss, M.; Boles, E. Determination of in Vivo Kinetics of the Starvation-Induced Hxt5 Glucose Transporter Of. FEMS Yeast Res. 2002, 2, 283–291. [Google Scholar] [CrossRef][Green Version]
- Maier, A.; Völker, B.; Boles, E.; Fuhrmann, G.F. Characterisation of Glucose Transport in Saccharomyces cerevisiae with Plasma Membrane Vesicles (Countertransport) and Intact Cells (Initial Uptake) with Single Hxt1, Hxt2, Hxt3, Hxt4, Hxt6, Hxt7 or Gal2 Transporters. FEMS Yeast Res. 2002, 2, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Ozcan, S.; Johnston, M. Two Different Repressors Collaborate to Restrict Expression of the Yeast Glucose Transporter Genes HXT2 and HXT4 to Low Levels of Glucose. Mol. Cell Biol. 1996, 16, 5536–5545. [Google Scholar] [CrossRef]
- Ozcan, S.; Johnston, M. Three Different Regulatory Mechanisms Enable Yeast Hexose Transporter (HXT) Genes to Be Induced by Different Levels of Glucose. Mol. Cell Biol. 1995, 15, 1564–1572. [Google Scholar] [CrossRef]
- Gonçalves, P.; Rodrigues de Sousa, H.; Spencer-Martins, I. FSY1, a Novel Gene Encoding a Specific Fructose/H+ Symporter in the Type Strain of Saccharomyces carlsbergensis. J. Bacteriol. 2000, 182, 5628–5630. [Google Scholar] [CrossRef]
- Varela, C.; Cárdenas, J.; Melo, F.; Agosin, E. Quantitative Analysis of Wine Yeast Gene Expression Profiles under Winemaking Conditions: Gene Expression ‘in’ Wine Fermentation. Yeast 2005, 22, 369–383. [Google Scholar] [CrossRef]
- Bisson, L. Stuck and Sluggish Fermentations. Am. J. Enol. Vitic. 1999, 50, 107–119. [Google Scholar]
- Pritchard, L.; Kell, D.B. Schemes of Flux Control in a Model of Saccharomyces cerevisiae Glycolysis: Flux Control in Yeast Glycolysis. Europ. J. Biochem. 2002, 269, 3894–3904. [Google Scholar] [CrossRef] [PubMed]
- Kayikci, Ö.; Nielsen, J. Glucose Repression in Saccharomyces Cerevisiae. FEMS Yeast Res. 2015, 15, fov068. [Google Scholar] [CrossRef]
- Treu, L.; Campanaro, S.; Nadai, C.; Toniolo, C.; Nardi, T.; Giacomini, A.; Valle, G.; Blondin, B.; Corich, V. Oxidative Stress Response and Nitrogen Utilization Are Strongly Variable in Saccharomyces cerevisiae Wine Strains with Different Fermentation Performances. Appl. Microbiol. Biotechnol. 2014, 98, 4119–4135. [Google Scholar] [CrossRef]
- Jordan, P.; Choe, J.-Y.; Boles, E.; Oreb, M. Hxt13, Hxt15, Hxt16 and Hxt17 from Saccharomyces cerevisiae Represent a Novel Type of Polyol Transporters. Sci. Rep. 2016, 6, 23502. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Molecular Function (SGD Curated) | Primer Sequence [5′ → 3′] | Amplicon Length | Efficiency % | R2 |
|---|---|---|---|---|---|
| HXT1 | HeXose Transporter (low-affinity glucose transporter) | F: 5′-GAA GCT GGC AGA ATC GAC GA-3′ | 71 bp | 102.9 | 0.996 |
| R: 5′-TAT GGA TGG TCA GGT GGG CA-3′ | |||||
| HXT2 | HeXose Transporter (high-affinity glucose transporter) | F: 5′-TGA ACT CCC AGC AAA GCC AA-3′ | 90 bp | 104.9 | 0.992 |
| R: 5′-TCC CAA CCA AAG ACA AAC CCA-3′ | |||||
| HXT3 | HeXose Transporter (low affinity glucose transporter) | F: 5′-CAC GTT ATT TGG TTG AAG CTG GT-3′ | 93 bp | 101.3 | 0.998 |
| R: 5′-GTT GAA TGA ATG GAT GGT CTG GG-3′ | |||||
| HXT4 | HeXose Transporter (high-affinity glucose transporter) | F: 5′-TGG TGG TAT GAC ATT CGT TCC-3′ | 101 bp | 104.7 | 0.989 |
| R: 5′-CGC TGA CCT TAT TTG AAA GAG CA-3′ | |||||
| HXT5 | HeXose Transporter (moderate affinity for glucose) | F: 5′-TCC AAA TCG CCT CCA TTG ACA-3′ | 77 bp | 102.5 | 0.983 |
| R: 5′-AAT ACC ACC AAC GCC CAG TC-3′ | |||||
| HXT6/7 | HeXose Transporter (high-affinity glucose transporter) | F: 5′-GAC TTT GGA AGA AGT CAA CAC CA-3′ | 106 bp | 100.0 | 0.998 |
| R: 5′-TTC TTC AGC GTC GTA GTT GGC-3′ | |||||
| HXT8 | Protein of unknown function with similarity to hexose transporters | F: 5′-AAT TCT GTC CAG TGG CGT GT-3′ | 81 bp | 95.2 | 0.991 |
| R: 5′-CGG AAC AAA CGT CAT ACC ACC-3′ | |||||
| FSY1 | Fructose/H+ symporter (Galeote et al., 2010) | F: 5′-CGA TGT TAA AGG CGG GTG GA-3′ | 98 bp | 95.1 | 0.989 |
| R: 5′-AAC GTG GTG ACT CGG GTA AG-3′ | |||||
| GAL2 | GALactose metabolism (also able to transport glucose) | F: 5′-GGG TCT GAA GGC TCC CAA AG-3′ | 85 bp | 104.7 | 0.981 |
| R: 5′-ACA AAC AAA GCA AGG AAA CGG T-3′ |
| Gene | Reference | Primer Sequence [5′ → 3′] | Amplicon Length | Efficiency % | R2 |
|---|---|---|---|---|---|
| ALG9 | [20] | F: 5′-CAC GGA TAG TGG CTT TGG TGA ACA ATT AC-3′ | 156 bp | 95.7 | 0.996 |
| R: 5′-TAT GAT TAT CTG GCA GCA GGA AAG AAC TTG GG-3′ | |||||
| UBC6 | [20] | F: 5′-GAT ACT TGG AAT CCT GGC TGG TCT GTC TC-3′ | 272 bp | 99.0 | 0.985 |
| R: 5′-AAA GGG TCT TCT GTT TCA TCA CCT GTA TTT GC-3′ | |||||
| FBA1 | [21] | F: 5′-GGT TTG TAC GCT GGT GAC ATC GC-3′ | 125 bp | 102.4 | 0.998 |
| R: 5′-CCG GAA CCA CCG TGG AAG ACC A-3′ | |||||
| PFK1 | [22] | F: 5′-GAG GTT GAT GCT TCT GGG TTC CGT-3′ | 138 bp | 97.7 | 0.998 |
| R: 5′-TGT GGC GGT TTC GTT GGT GTC G-3′ |
| Strain | CO2 Production Peak (g/L/hours) | Peak Time (hours) | Sampling Time 45 g/L (hours) | Sampling Time 60 g/L (hours) | Fermentation Time (hours) |
|---|---|---|---|---|---|
| EC1118 | 1.53 ± 0.05 A | 16.75 ± 0.85 B | 50.3 ± 2.7 A | 73.9 ± 3.7 B | 110.2 ± 1.7 A |
| R008 | 1.76 ± 0.00 B | 14.91 ± 0.33 A | 46.3 ± 4.9 A | 67.5 ± 3.1 AB | 156.9 ± 1.0 C |
| R31.3 | 1.76 ± 0.05 B | 14.08 ± 1.08 A | 41.8 ± 1.2 A | 64.6 ± 1.2 A | 134.9 ± 4.2 B |
| P301.9 | 1.93 ± 0.05 C | 15.58 ± 0.84 AB | 43.2 ± 4.1 A | 63.5 ± 1.5 A | 131.7 ± 4.2 B |
| Ethanol (% vol.) | Glucose (g/L) | Fructose (g/L) | |
|---|---|---|---|
| EC1118 | 11.49 ± 0.04 B | nd | nd |
| P301.9 | 11.45 ± 0.04 B | nd | nd |
| R31.3 | 11.38 ± 0.26 B | nd | nd |
| R008 | 10.64 ± 0.07 A | nd | 7.62 ± 0.04 |
| Fructose/Total Sugars (%) | NH2 (mg/L) | |||||
|---|---|---|---|---|---|---|
| 45 g/L | 60 g/L | p | 45 g/L | 60 g/L | ||
| EC1118 | 60.30 ± 0.43 A | 66.87 ± 0.70 A | <0.0001 *** | 61.17 ± 2.35 | 59.87 ± 3.10 | Ns |
| R008 | 66.05 ± 0.29 B | 74.97 ± 1.65 B | 0.012 * | 55.75 ± 1.95 | 63.10 ± 1.70 | * |
| R31.3 | 67.01 ± 1.08 B | 79.27 ± 1.84 B | 0.016 * | 51.93 ± 7.35 | 56.57 ± 6.33 | * |
| P301.9 | 66.56 ± 0.74 B | 76.67 ± 1.72 B | 0.002 ** | 38.03 ± 4.29 | 39.20 ± 3.21 | Ns |
| Gene | Strain | 45 g/L of CO2 Produced | 60 g/L of CO2 Produced | ||
|---|---|---|---|---|---|
| GAL2 | EC1118 | 2.660 ± 0.655 | C | 1.533 ± 0.434 | B |
| R008 | 0.775 ± 0.184 | A | 0.465 ± 0.082 | A | |
| R31.3 | 1.801 ± 0.519 | B | 0.851 ± 0.521 | A | |
| P301.9 | 0.862 ± 0.236 | A | 0.914 ± 0.131 | AB | |
| HXT1 | EC1118 | 0.925 ± 0.239 | B | 0.771 ± 0.130 | B |
| R008 | 0.996 ± 0.125 | B | 1.072 ± 0.116 | BC | |
| R31.3 | 0.475 ± 0.045 | A | 0.356 ± 0.075 | A | |
| P301.9 | 0.849 ± 0.111 | B | 1.300 ± 0.254 | C | |
| HXT2 | EC1118 | 1.364 ± 0.565 | B | 0.536 ± 0.385 | B |
| R008 | 0.304 ± 0.198 | A | 0.089 ± 0.017 | A | |
| R31.3 | 0.399 ± 0.044 | A | 0.084 ± 0.045 | A | |
| P301.9 | 0.181 ± 0.052 | A | 0.167 ± 0.035 | A | |
| HXT3 | EC1118 | 3.004 ± 0.977 | B | 1.385 ± 0.134 | BC |
| R008 | 1.257 ± 0.106 | A | 1.031 ± 0.123 | AB | |
| R31.3 | 1.454 ± 0.395 | A | 0.648 ± 0.101 | A | |
| P301.9 | 1.117 ± 0.183 | A | 1.662 ± 0.399 | C | |
| HXT4 | EC1118 | 1.642 ± 0.455 | C | 1.090 ± 0.224 | C |
| R008 | 0.578 ± 0.234 | A | 0.640 ± 0.126 | B | |
| R31.3 | 1.065 ± 0.230 | B | 0.240 ± 0.128 | A | |
| P301.9 | 0.686 ± 0.194 | AB | 1.209 ± 0.474 | C | |
| HXT5 | EC1118 | 1.028 ± 0.466 | B | 0.859 ± 0.257 | A |
| R008 | 0.806 ± 0.281 | AB | 1.673 ± 0.754 | A | |
| R31.3 | 0.731 ± 0.158 | AB | 3.341 ± 1.546 | B | |
| P301.9 | 0.505 ± 0.116 | B | 1.301 ± 0.338 | A | |
| HXT6/7 | EC1118 | 2.252 ± 0.181 | C | 1.439 ± 0.050 | A |
| R008 | 1.045 ± 0.129 | A | 1.502 ± 0.499 | A | |
| R31.3 | 1.511 ± 0.447 | B | 2.782 ± 0.938 | B | |
| P301.9 | 1.267 ± 0.277 | AB | 1.931 ± 0.613 | AB | |
| HXT8 | EC1118 | 1.354 ± 0.496 | B | 2.430 ± 0.870 | B |
| R008 | 0.593 ± 0.097 | A | 0.862 ± 0.314 | A | |
| R31.3 | 1.613 ± 0.390 | B | 2.782 ± 1.106 | B | |
| P301.9 | 0.606 ± 0.213 | A | 1.289 ± 0.692 | A | |
| P301_O30021 | EC1118 | 1.254 ± 0.496 | A | 0.712 ± 0.256 | A |
| R31.3 | 1.709 ± 0.308 | A | 3.164 ± 1.018 | B | |
| P301.9 | 1.617 ± 0.203 | A | 3.301 ± 0.727 | B | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nadai, C.; Crosato, G.; Giacomini, A.; Corich, V. Different Gene Expression Patterns of Hexose Transporter Genes Modulate Fermentation Performance of Four Saccharomyces cerevisiae Strains. Fermentation 2021, 7, 164. https://doi.org/10.3390/fermentation7030164
Nadai C, Crosato G, Giacomini A, Corich V. Different Gene Expression Patterns of Hexose Transporter Genes Modulate Fermentation Performance of Four Saccharomyces cerevisiae Strains. Fermentation. 2021; 7(3):164. https://doi.org/10.3390/fermentation7030164
Chicago/Turabian StyleNadai, Chiara, Giulia Crosato, Alessio Giacomini, and Viviana Corich. 2021. "Different Gene Expression Patterns of Hexose Transporter Genes Modulate Fermentation Performance of Four Saccharomyces cerevisiae Strains" Fermentation 7, no. 3: 164. https://doi.org/10.3390/fermentation7030164
APA StyleNadai, C., Crosato, G., Giacomini, A., & Corich, V. (2021). Different Gene Expression Patterns of Hexose Transporter Genes Modulate Fermentation Performance of Four Saccharomyces cerevisiae Strains. Fermentation, 7(3), 164. https://doi.org/10.3390/fermentation7030164