Trichuris spp. in Animals, with Specific Reference to Neo-Tropical Rodents


Abstract
1. Introduction
2. Methodology
3. Trichuris spp. of Veterinary and Public Health Importance
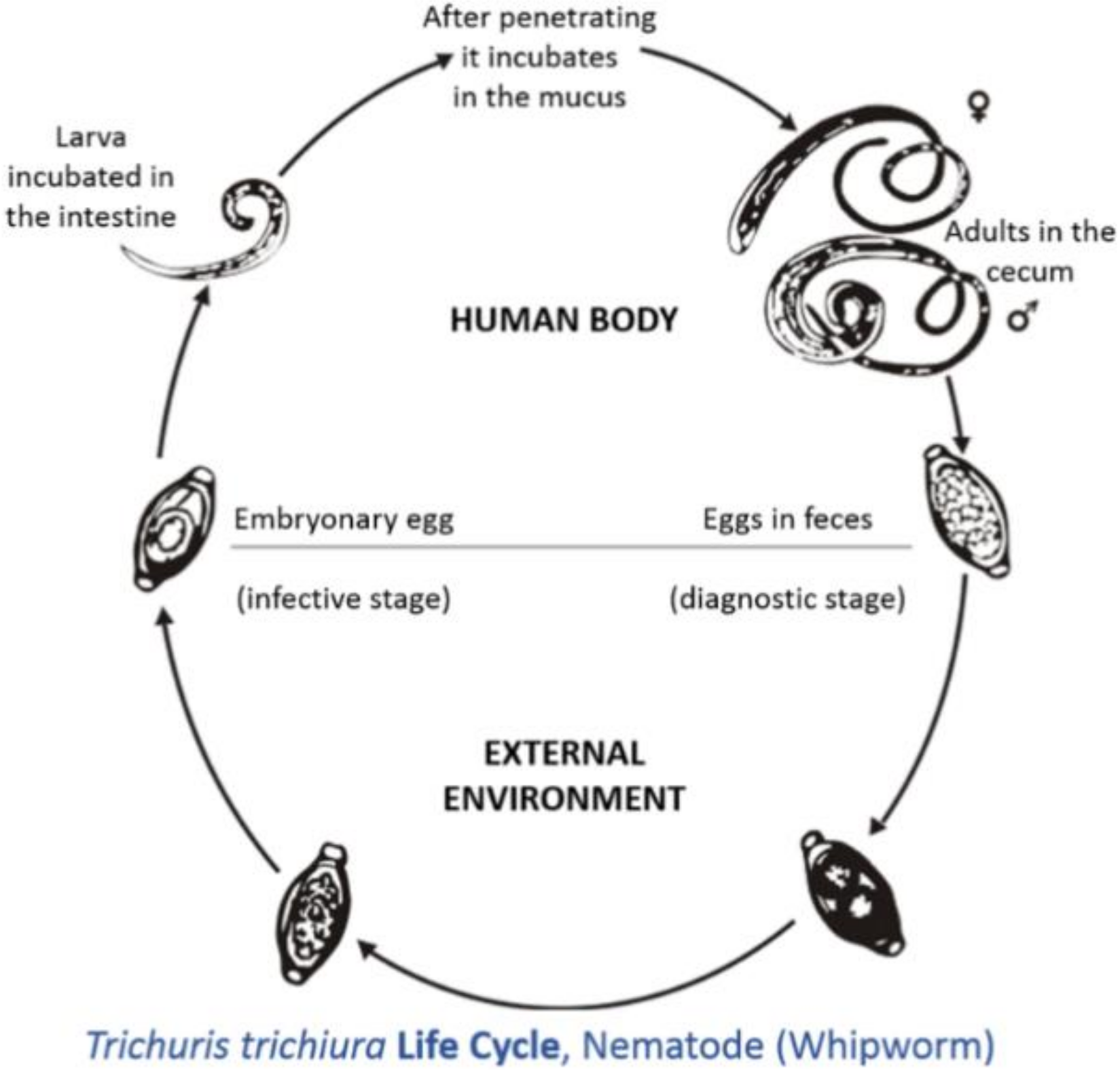
3.1. Trichuriasis of Man
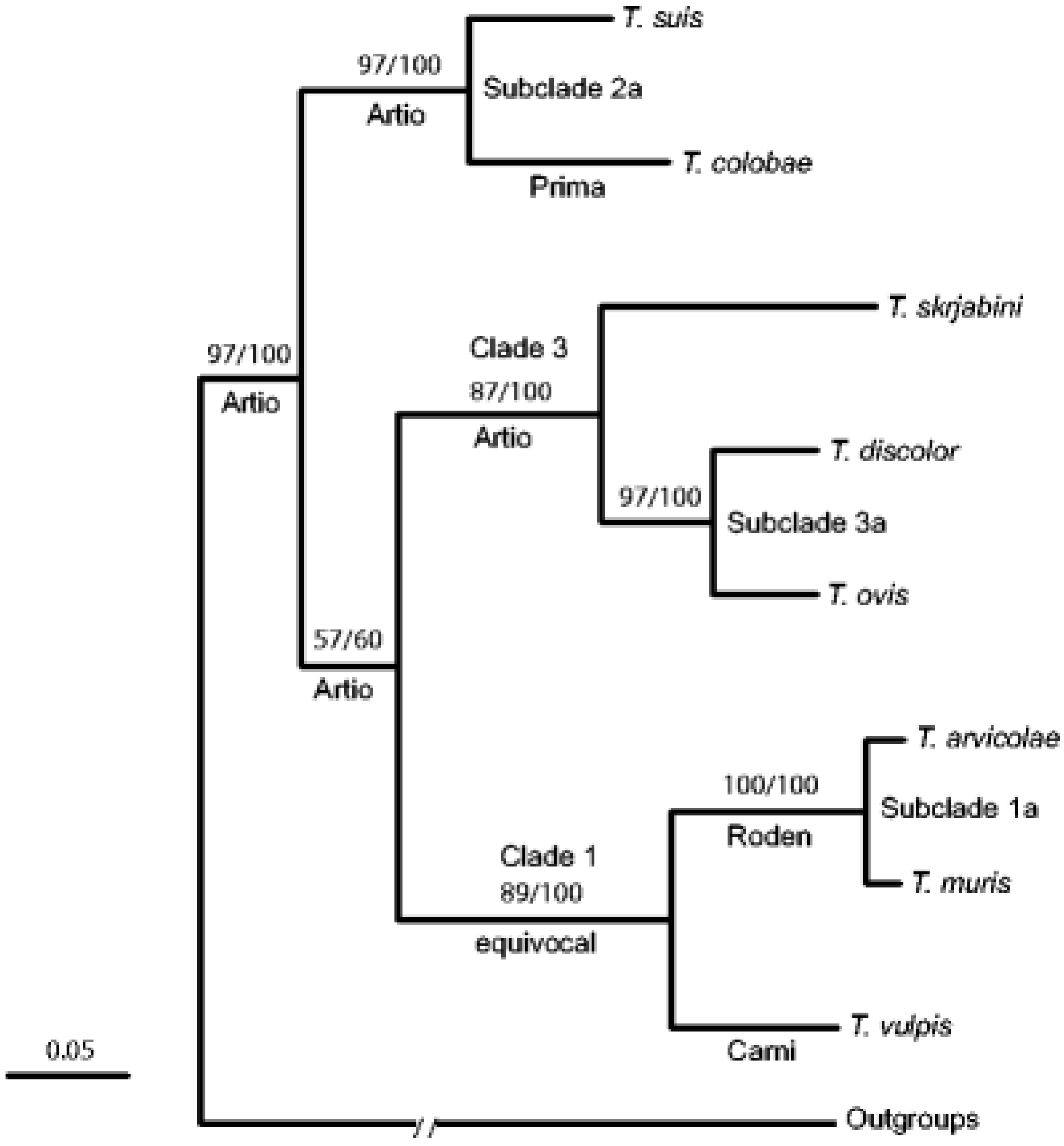
3.2. Morphological and Molecular Identifications of Trichuris spp.
3.2.1. Morphological Identification of Trichuris spp. in Pigs, Dogs, Cats, Humans, and Non-Human Primates
3.2.2. Molecular Identification of Trichuris spp. in Domestic and Non-Domestic Ruminants
3.2.3. Molecular Identification of Trichuris spp. in Cats, Dogs, Pigs, Humans, and Non-Human Primates
3.2.4. Molecular Identification Trichuris spp. in Rodents
3.3. Immunomodulatory Effect of Trichuris spp.
4. Domesticated Neo-Tropical Rodent
4.1. Guinea Pig (Cavia Porcellus)
4.2. Semi-Domesticated Neo-Tropical Rodents
4.2.1. Agouti (Dasyprocta leporina)
4.2.2. Lappe (Agouti paca/Cuniculus paca)
4.2.3. Capybara (Hydrochoerus hydrochaeris)
5. Conclusions
6. Recommendations
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown-Uddenberg, R.C.; Garcia, G.W.; Baptiste, Q.S.; Counand, T.; Adogwa, A.O.; Sampson, T. The Agouti (Dasyprocta leporina, D. aguti) Booklet and Producers’ Manual; GWG Publications: Champs Fleurs, Trinidad and Tobago, 2014; Available online: http://ostasp.brinkster.net/ (accessed on 17 June 2020).
- Jones, K.R.; Garcia, G.W. Gastrointestinal parasites of domesticated animals introduced into the Neo-tropics (New World Tropics). Concepts Dairy Vet. Sci. 2018, 1, 56–78. [Google Scholar]
- Jones, K.R.; Garcia, G.W. Endoparasites of domesticated animals that originated in the neo-tropics (new world tropics). Vet. Sci. 2019, 6, 24. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.R.; Lall, K.R.; Garcia, G.W. Endoparasites of selective native non-domesticated mammals in the neo-tropics (new world tropics). Vet. Sci. 2019, 6, 87. [Google Scholar] [CrossRef] [PubMed]
- Paterson, R.T.; Joaquin, N.; Chamon, K.; Palomino, E. The productivity of small animal species in small-scale mixed farming systems in subtropical Bolivia. Trop. Anim. Health Prod. 2001, 33, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Manjeli, Y.; Tchoumboue, J.; Njwe, R.M.; Teguia, A. Guinea-pig productivity under traditional management. Trop. Anim. Health Prod. 1998, 30, 115–122. [Google Scholar] [CrossRef]
- Jones, K.R.; Garcia, G.W. Anthelmintic usage on the reproductive parameter in captive reared agoutis (Dasyprocta leporina) in Trindad and Tobago, West Indies. Trop. Agric. 2020, 97, in Press. [Google Scholar]
- Govoni, G.; Fielding, D. Paca (Agouti paca) and Agouti (Dasyprocta spp.)-Minilivestock Production in the Amazonas State of Venezuela: 1. Biology. Tropicultura 2001, 19, 56–60. [Google Scholar]
- Chapman, C.A. Biology of Capybaras. J. Mammal. 1991, 72, 206–208. [Google Scholar] [CrossRef]
- Alvarez, M.R.; Kravetz, F.O. Reproductive performance of capybaras (Hyrochoerus hydrochaeris) in captivity under different management systems in Argentina. Anim. Res. 2006, 55, 153–164. [Google Scholar] [CrossRef]
- Ali, A.J.; Jones, K.R. Nutritive value and physical properties of Neo-tropical rodent meat-with emphasis on the Capybara (Hydrochoerus hydrochaeris). Animals 2020, 10, 2134. [Google Scholar] [CrossRef]
- Dalle Zotte, A. Perception of rabbit meat quality and major factors influencing the rabbit carcass and meat quality. Livest. Prod. Sci. 2002, 75, 11–32. [Google Scholar] [CrossRef]
- Pla, M.; Pascual, M.; Arino, B. Protein, Fat and Moisture Content of Retail Cuts of Rabbit Meat Evaluated with the NIRS Methodology. World Rabbit. Sci. 2004, 12, 149–158. [Google Scholar] [CrossRef][Green Version]
- Nogueira-Filho, S.L.G.; Nogueira, S.S.C. Capybara meat: An extraordinary resource for food security in South America. Meat Sci. 2018, 145, 329–333. [Google Scholar] [CrossRef]
- Soulsby, E.J. Helminths, Arthropods and Protozoa of Domesticated Animals, 6th ed.; Baillière Tindall & Cassell; The American Society of Tropical Medicine and Hygiene: Arlington, VA, USA, 1968. [Google Scholar]
- Zajac, A.M.; Conboy, G.A. Veterinary Clinical Parasitology; John Wiley & Sons: Hoboken, NJ, USA, 2012. [Google Scholar]
- Life Cycle of Trichuris Found inside the Body. Available online: https://en.wikipedia.org/wiki/Trichuris_trichiura#/media/File:Trichuris_trichiura_Life_Cycle.tif (accessed on 7 January 2021).
- Albonico, M.; Allen, H.; Chitsulo, L.; Engels, D.; Gabrielli, A.F.; Savioli, L. Controlling soil-transmitted helminthiasis in pre-school-age children through preventive chemotherapy. PLoS Negl. Trop. Dis. 2008, 2, e126. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.E.; Sonnenberg, B. An apparent case of human infection with whip-worm of dogs, Trichuris vulpis (Froelich, 1789). J. Parasitol. 1956, 42, 197–199. [Google Scholar] [CrossRef]
- Phosuk, I.; Sanpool, O.; Thanchomnang, T.; Sadaow, L.; Rodpai, R.; Anamnart, W.; Janwan, P.; Wijit, A.; Laymanivong, S.; Aung, W.P.P.; et al. Molecular identification of Trichuris suis and Trichuris trichiura eggs in human populations from Thailand, Lao PDR, and Myanmar. Am. J. Trop. Med. Hyg. 2018, 98, 39–44. [Google Scholar] [CrossRef]
- Beer, R.J. Experimental infection of man with pig whipworm. Br. Med. J. 1971, 2, 44. [Google Scholar] [CrossRef][Green Version]
- Kradin, R.L.; Badizadegan, K.; Auluck, P.; Korzenik, J.; Lauwers, G.Y. Iatrogenic Trichuris suis infection in a patient with Crohn disease. Arch. Pathol. Lab. Med. 2006, 130, 718–720. [Google Scholar]
- Sunkara, T.; Sharma, S.R.; Ofosu, A. Trichuris trichiura—An Unwelcome Surprise during Colonoscopy. Am. J. Trop. Med. Hyg. 2018, 99, 555. [Google Scholar] [CrossRef]
- Vásquez, O.T.; Martínez, I.B.; Romero, R.C.; Valencia, S.R.; Tay, J.Z. Mixed infection by Trichuris trichiura and Trichuris vulpis. Rev. Gastroenterol. Peru Organo Soc. Gastroenterol. Peru 1997, 17, 255–258. [Google Scholar]
- Areekul, P.; Putaporntip, C.; Pattanawong, U.; Sitthicharoenchai, P.; Jongwutiwes, S. Trichuris vulpis and T. trichiura infections among schoolchildren of a rural community in north-western Thailand: The possible role of dogs in disease transmission. Asian Biomed. 2010, 4, 49–60. [Google Scholar] [CrossRef]
- Dunn, J.J.; Columbus, S.T.; Aldeen, W.E.; Davis, M.; Carroll, K.C. Trichuris vulpis recovered from a patient with chronic diarrhea and five dogs. J. Clin. Microbiol. 2002, 40, 2703–2704. [Google Scholar] [CrossRef] [PubMed]
- Márquez-Navarro, A.; García-Bracamontes, G.; Álvarez-Fernández, B.E.; Ávila-Caballero, L.P.; Santos-Aranda, I.; Díaz-Chiguer, D.L.; Sánchez-Manzano, R.M.; Rodríguez-Bataz, E.; Nogueda-Torres, B. Trichuris vulpis (Froelich, 1789) infection in a child: A case report. Korean J. Parasitol. 2012, 50, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Olsen, A.; Namwanje, H.; Nejsum, P.; Roepstorff, A.; Thamsborg, S.M. Albendazole and mebendazole have low efficacy against Trichuris trichiura in school-age children in Kabale District, Uganda. Trans. R. Soc. Trop. Med. Hyg. 2009, 103, 443–446. [Google Scholar] [CrossRef] [PubMed]
- Ooi, H.K.; Tenora, F.; Itoh, K.; Kamiya, M. Comparative study of Trichuris trichiura from non-human primates and from man, and their difference with T. suis. J. Vet. Med. Sci. 1993, 55, 363–366. [Google Scholar] [CrossRef]
- Kuchai, J.A.; Ahmad, F.; Chishti, M.Z.; Dar, J.A.; Tak, H. On morphology and morphometry of Trichuris ovis Abildgaard, 1795 recovered from ruminants of Ladakh, India. J. Buffalo Sci. 2013, 2, 49–52. [Google Scholar] [CrossRef]
- Ketzis, J.K. Trichuris spp. infecting domestic cats on St. Kitts: Identification based on size or vulvar structure? SpringerPlus 2015, 4, 115. [Google Scholar] [CrossRef]
- Yevstafieva, V.A.; Kravchenko, S.O.; Gutyj, B.V.; Melnychuk, V.V.; Kovalenko, P.N.; Volovyk, L.B. Morphobiological analysis of Trichuris vulpis (Nematoda, Trichuridae), obtained from domestic dogs. Regul. Mech. Biosyst. 2019, 10, 165–171. [Google Scholar] [CrossRef]
- García-Sánchez, A.M.; Rivero, J.; Callejón, R.; Zurita, A.; Reguera-Gomez, M.; Valero, M.A.; Cutillas, C. Differentiation of Trichuris species using a morphometric approach. Int. J. Parasitol. Parasites Wildl. 2019, 9, 218–223. [Google Scholar] [CrossRef]
- García-Sánchez, A.M.; Reguera-Gomez, M.; Valero, M.A.; Cutillas, C. Differentiation of Trichuris species eggs from non-human primates by geometric morphometric analysis. Int. J. Parasitol. Parasites Wildl. 2020, 12, 214–219. [Google Scholar] [CrossRef]
- Cutillas, C.; German, P.; Arias, P.; Guevara, D. Trichuris ovis and Trichuris globulosa: Morphological, biometrical, and genetic studies. Exp. Parasitol. 1995, 81, 621–625. [Google Scholar] [CrossRef] [PubMed]
- Cutillas, C.; German, P.; Arias, P.; Guevara, D. Characterization of Trichuris skrjabini by isoenzyme gel electrophoresis: Comparative study with Trichuris ovis. Acta Trop. 1996, 62, 63–69. [Google Scholar] [CrossRef]
- Cutillas, C.; Oliveros, R.; De Rojas, M.; Guevara, D.C. Determination of Trichuris skrjabini by sequencing of the ITS1–5.8 S–ITS2 segment of the ribosomal DNA: Comparative molecular study of different species of trichurids. J. Parasitol. 2004, 90, 648–652. [Google Scholar] [CrossRef]
- Oliveros, R.; Cutillas, C.; De Rojas, M.; Arias, P. Characterization of four species of Trichuris (Nematoda: Enoplida) by their second internal transcribed spacer ribosomal DNA sequence. Parasitol. Res. 2000, 86, 1008–1013. [Google Scholar] [CrossRef]
- Callejón, R.; De Rojas, M.; Ariza, C.; Ubeda, J.M.; Guevara, D.C.; Cutillas, C. Cytochrome oxidase subunit 1 and mitochondrial 16S rDNA sequences of Trichuris skrjabini (Tricocephalida: Trichuridae). Parasitol. Res. 2009, 104, 715–716. [Google Scholar] [CrossRef] [PubMed]
- Callejón, R.; Halajian, A.; De Rojas, M.; Marrugal, A.; Guevara, D.; Cutillas, C. 16S partial gene mitochondrial DNA and internal transcribed spacers ribosomal DNA as differential markers of Trichuris discolor populations. Vet. Parasitol. 2012, 186, 350–363. [Google Scholar] [CrossRef] [PubMed]
- Callejón, R.; Nadler, S.; De Rojas, M.; Zurita, A.; Petrášová, J.; Cutillas, C. Molecular characterization and phylogeny of whipworm nematodes inferred from DNA sequences of cox1 mtDNA and 18S rDNA. Parasitol. Res. 2013, 112, 3933–3949. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.H.; Wang, Y.; Xu, M.J.; Zhou, D.H.; Ye, Y.G.; Li, J.Y.; Song, H.Q.; Lin, R.Q.; Zhu, X.Q. Characterization of the complete mitochondrial genomes of two whipworms Trichuris ovis and Trichuris discolor (Nematoda: Trichuridae). Infect. Genet. Evol. 2012, 12, 1635–1641. [Google Scholar] [CrossRef]
- Salaba, O.; Rylková, K.; Vadlejch, J.; Petrtýl, M.; Scháňková, Š.; Brožová, A.; Jankovská, I.; Jebavý, L.; Langrová, I. The first determination of Trichuris sp. from roe deer by amplification and sequenation of the ITS1-5.8 S-ITS2 segment of ribosomal DNA. Parasitol. Res. 2013, 112, 955–960. [Google Scholar] [CrossRef]
- Vejl, P.; Nechybová, S.; Peřinková, P.; Melounová, M.; Sedláková, V.; Vašek, J.; Čílová, D.; Rylková, K.; Jankovská, I.; Vadlejch, J.; et al. Reliable molecular differentiation of Trichuris ovis and Trichuris discolor from sheep (Ovis orientalis aries) and roe deer (Capreolus capreolus) and morphological characterisation of their females: Morphology does not work sufficiently. Parasitol. Res. 2017, 116, 2199–2210. [Google Scholar] [CrossRef]
- Nechybová, S.; Vejl, P.; Hart, V.; Melounová, M.; Čílová, D.; Vašek, J.; Jankovská, I.; Vadlejch, J.; Langrová, I. Long-term occurrence of Trichuris species in wild ruminants in the Czech Republic. Parasitol. Res. 2018, 117, 1699–1708. [Google Scholar] [CrossRef] [PubMed]
- Wulcan, J.M.; Ketzis, J.K.; Dennis, M.M. Typhlitis Associated with Natural Trichuris sp. Infection in Cats. Vet. Pathol. 2020, 57, 266–271. [Google Scholar] [CrossRef] [PubMed]
- Ketzis, J.K.; Verma, A.; Burgess, G. Molecular characterization of Trichuris serrata. Parasitol. Res. 2015, 114, 1993–1995. [Google Scholar] [CrossRef] [PubMed]
- Cutillas, C.; de Rojas, M.; Ariza, C.; Ubeda, J.M.; Guevara, D. Molecular identification of Trichuris vulpis and Trichuris suis isolated from different hosts. Parasitol. Res. 2007, 100, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Elsemore, D.A.; Geng, J.; Flynn, L.; Cruthers, L.; Lucio-Forster, A.; Bowman, D.D. Enzyme-linked immunosorbent assay for coproantigen detection of Trichuris vulpis in dogs. J. Vet. Diagn. Investig. 2014, 26, 404–411. [Google Scholar] [CrossRef]
- Oliveros, R.; Cutillas, C.; Arias, P.; Guevara, D. Morphologic, biometric, and isoenzyme characterization of Trichuris suis. Parasitol. Res. 1998, 84, 513–515. [Google Scholar] [CrossRef]
- Liu, G.H.; Zhou, W.; Nisbet, A.J.; Xu, M.J.; Zhou, D.H.; Zhao, G.H.; Wang, S.K.; Song, H.Q.; Lin, R.Q.; Zhu, X.Q. Characterization of Trichuris trichiura from humans and T. suis from pigs in China using internal transcribed spacers of nuclear ribosomal DNA. J. Helminthol. 2014, 88, 64–68. [Google Scholar] [CrossRef]
- Muramatsu, R.; Sato, R.; Onuma, N.; Sasai, K.; Shibahara, T.; Matsubayashi, M. Molecular Identification of Trichuris suis Worms and Eggs in Pig Feces, Infected Intestines, and Farm Environments in Japan. Jpn. Agric. Res. Q. JARQ 2020, 54, 271–275. [Google Scholar] [CrossRef]
- Cutillas, C.; Callejon, R.; De Rojas, M.; Tewes, B.; Ubeda, J.M.; Ariza, C.; Guevara, D.C. Trichuris suis and Trichuris trichiura are different nematode species. Acta Trop. 2009, 111, 299–307. [Google Scholar] [CrossRef]
- Nissen, S.; Al-Juburry, A.; Hansen, V.A.; Olsen, A.; Christensen, H.; Thamsborg, S.M.; Nejsum, P. Genetic analysis of Trichuris suis and Trichuris trichiura recovered from humans and pigs in a sympatric setting in Uganda. Vet. Parasitol. 2012, 188, 68–77. [Google Scholar] [CrossRef]
- Liu, G.H.; Gasser, R.B.; Su, A.; Nejsum, P.; Peng, L.; Lin, R.Q.; Li, M.W.; Xu, M.J.; Zhu, X.Q. Clear genetic distinctiveness between human-and pig-derived Trichuris based on analyses of mitochondrial datasets. PLoS Negl. Trop. Dis. 2012, 6, e1539. [Google Scholar] [CrossRef] [PubMed]
- Arizono, N.; Yamada, M.; Tegoshi, T.; Onishi, K. Molecular identification of Oesophagostomum and Trichuris eggs isolated from wild Japanese macaques. Korean J. Parasitol. 2012, 50, 253–257. [Google Scholar] [CrossRef] [PubMed]
- Ravasi, D.F.; O’Riain, M.J.; Davids, F.; Illing, N. Phylogenetic evidence that two distinct Trichuris genotypes infect both humans and non-human primates. PLoS ONE 2012, 7, e44187. [Google Scholar] [CrossRef] [PubMed]
- Ghai, R.R.; Simons, N.D.; Chapman, C.A.; Omeja, P.A.; Davies, T.J.; Ting, N.; Goldberg, T.L. Hidden population structure and cross-species transmission of whipworms (Trichuris sp.) in humans and non-human primates in Uganda. PLoS Negl. Trop. Dis. 2014, 8, e3256. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.H.; Gasser, R.B.; Nejsum, P.; Wang, Y.; Chen, Q.; Song, H.Q.; Zhu, X.Q. Mitochondrial and nuclear ribosomal DNA evidence supports the existence of a new Trichuris species in the endangered françois’ leaf-monkey. PLoS ONE 2013, 8, e 66249. [Google Scholar] [CrossRef]
- Cutillas, C.; de Rojas, M.; Zurita, A.; Oliveros, R.; Callejón, R. Trichuris colobae n. sp. (Nematoda: Trichuridae), a new species of Trichuris from Colobus guereza kikuyensis. Parasitol. Res. 2014, 113, 2725–2732. [Google Scholar] [CrossRef]
- Callejón, R.; de Rojas, M.; Nieberding, C.; Foronda, P.; Feliú, C.; Guevara, D.; Cutillas, C. Molecular evolution of Trichuris muris isolated from different Muridae hosts in Europe. Parasitol. Res. 2010, 107, 631–641. [Google Scholar] [CrossRef]
- Feliu, C.; Spakulová, M.; Casanova, J.C.; Renaud, F.; Morand, S.; Hugot, J.P.; Santalla, F.; Durand, P. Genetic and morphological heterogeneity in small rodent whipworms in southwestern Europe: Characterization of Trichuris muris and description of Trichuris arvicolae n. sp. (Nematoda: Trichuridae). J. Parasitol. 2000, 86, 442–449. [Google Scholar] [CrossRef]
- Cutillas, C.; Oliveros, R.; De Rojas, M.; Guevara, D.C. Determination of Trichuris muris from murid hosts and T. arvicolae (Nematoda) from arvicolid rodents by amplification and sequentiation of the ITS1–5.8 S-ITS2 segment of the ribosomal DNA. Parasitol. Res. 2002, 88, 574–582. [Google Scholar] [CrossRef]
- Callejon, R.; de Rojas, M.; Feliu, C.; Balao, F.; Marrugal, A.; Henttonen, H.; Guevara, D.; Cutillas, C. Phylogeography of Trichuris populations isolated from different Cricetidae rodents. Parasitology 2012, 139, 1795–1813. [Google Scholar] [CrossRef]
- Robles, M.D.R. New species of Trichuris (Nematoda: Trichuridae) from Akodon montensis Thomas, 1913, of the Paranaense forest in Argentina. J. Parasitol. 2011, 97, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Robles, M.D.R.; Cutillas, C.; Panei, C.J.; Callejón, R. Morphological and molecular characterization of a new Trichuris species (Nematoda-Trichuridae), and phylogenetic relationships of Trichuris species of cricetid rodents from Argentina. PLoS ONE 2014, 9, e112069. [Google Scholar] [CrossRef] [PubMed]
- Callejón, R.; Robles, M.D.R.; Panei, C.J.; Cutillas, C. Molecular diversification of Trichuris spp. from Sigmodontinae (Cricetidae) rodents from Argentina based on mitochondrial DNA sequences. Parasitol. Res. 2016, 115, 2933–2945. [Google Scholar] [CrossRef] [PubMed]
- Robles, M.D.R.; Cutillas, C.; Callejón, R. Morphological-molecular characterization and phylogenetic relationships of a new Trichuris species (Nematoda: Trichuridae) parasitic on Holochilus chacarius (Cricetidae: Sigmodontinae) from the Chaco ecoregion (Argentina). Infect. Genet. Evol. 2018, 58, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Callejón, R.; Cutillas, C.; Nadler, S.A. Nuclear and mitochondrial genes for inferring Trichuris phylogeny. Parasitol. Res. 2015, 114, 4591–4599. [Google Scholar] [CrossRef] [PubMed]
- Summers, R.W.; Elliott, D.E.; Qadir, K.; Urban, J.F., Jr.; Thompson, R.; Weinstock, J.V. Trichuris suis seems to be safe and possibly effective in the treatment of inflammatory bowel disease. Am. J. Gastroenterol. 2003, 98, 2034–2041. [Google Scholar] [CrossRef]
- Summers, R.W.; Elliott, D.E.; Urban, J.F.; Thompson, R.; Weinstock, J.V. Trichuris suis therapy in Crohn’s disease. Gut 2005, 54, 87–90. [Google Scholar] [CrossRef]
- Summers, R.W.; Elliott, D.E.; Urban, J.F., Jr.; Thompson, R.A.; Weinstock, J.V. Trichuris suis therapy for active ulcerative colitis: A randomized controlled trial. Gastroenterology 2005, 128, 825–832. [Google Scholar] [CrossRef]
- Parthasarathy, G.; Mansfield, L.S. Trichuris suis excretory secretory products (ESP) elicit interleukin-6 (IL-6) and IL-10 secretion from intestinal epithelial cells (IPEC-1). Vet. Parasitol. 2005, 131, 317–324. [Google Scholar] [CrossRef]
- Santos, L.N.; Gallo, M.B.; Silva, E.S.; Figueiredo, C.A.V.; Cooper, P.J.; Barreto, M.L.; Loureiro, S.; Pontes-de-Carvalho, L.C.; Alcantara-Neves, N.M. A proteomic approach to identify proteins from Trichuris trichiura extract with immunomodulatory effects. Parasite Immunol. 2013, 35, 188–193. [Google Scholar] [CrossRef]
- Liu, Z.; Kelleher, A.; Tabb, S.; Wei, J.; Pollet, J.; Hotez, P.J.; Bottazzi, M.E.; Zhan, B.; Asojo, O.A. Identification, Characterization, and Structure of Tm16 from Trichuris muris. J. Parasitol. Res. 2017, 2017, 4342789. [Google Scholar] [CrossRef] [PubMed]
- Shears, R.K.; Bancroft, A.J.; Sharpe, C.; Grencis, R.K.; Thornton, D. Vaccination against whipworm: Identification of potential immunogenic proteins in Trichuris muris excretory/secretory material. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Umba, J.M.; Kashala, J.C.K.; Ilaka, A.N.; Ngulu, A.N.; Tshikung, K.M.D.; Atangana, A.; Khasa, D. Mortality of young guinea pigs (Cavia porcellus) crossed and its main causes around Kinshasa, DR Congo. Livest. Res. Rural Dev. 2016, 7, 1–8. [Google Scholar]
- Kouman, M.K.; Meutchieye, F.; Nguafack, T.T.; Miegoue, E.; Tchoumboue, J.; Theodoropoulos, G. Parasitic fauna of domestic cavies in the western highland of Cameroon (Central Africa). BMC Vet. Res. 2015, 11, 288. [Google Scholar]
- d’ Ovidio, D.; Noviello, E.; Ianniello, D.; Cringoli, G.; Rinaldi, L. Survey of endoparasites in pet guinea pigs in Italy. Parasitol. Res. 2015, 114, 1213–1216. [Google Scholar] [CrossRef]
- Matamedi, G.; Moharami, M.; Paykari, H.; Eslampanah, M.; Omraninava, A. A survey of the gastrointestinal parasites of rabbits and guinea pigs in a laboratory animal house. Arch. Razi Inst. 2014, 69, 77–81. [Google Scholar]
- Gressler, L.T.; Silva, A.S.D.; Silva, M.K.D.; Tonin, A.A.; Monteiro, S.G. Gastrointestinal parasites of cavy (Cavia aperea aperea) in southern Brazil. Res. Vet. Sci. 2010, 89, 206–208. [Google Scholar] [CrossRef]
- Pinto, R.M.; Gomes, D.C.; Muniz-Pereira, L.C.; Noronha, D. Helminths of guinea pigs, Cavia porcellus (Linnaeus), in Brazil. Rev. Bras. Zool. 2002, 19, 261–269. [Google Scholar] [CrossRef]
- Dittmar, K. Arthropod and helminths parasites of the wild guinea pig, Cavia aperea, from the Andes and the Cordillera in Peru, South America. J. Parasitol. 2002, 88, 409–411. [Google Scholar] [CrossRef]
- Vargas, M.R.; Chavez, A.V.; Pinedo, R.S.; Morales, S.C.; Suarez, F.A. Gastrointestinal parasitism in two seasons in guinea pigs (Cavia porcellus) of Oxapampa, Pasco. Rev. Int. Vet. Peru 2014, 25, 276–283. [Google Scholar]
- Garcia, C.J.; Chavez, A.V.; Pinedo, R.S.; Suarez, F.A. Gastrointestinal helminthiasis in guinea pig (Cavia porcellus) family-commercial breeding farms in Ancash, Peru. Rev. Int. Vet. Peru 2013, 24, 473–479. [Google Scholar]
- Meutchieye, F.; Kouman, M.K.; Miegoue, E.; Nguafack, T.T.; Tchoumboue, J.; Teguia, A.; Theodoropoulos, G. A survey of potentially zoonotic gastrointestinal parasites in domestic cavies in Cameroon (Central Africa). BMC Vet. Res. 2017, 13, 196. [Google Scholar] [CrossRef] [PubMed]
- Payne, V.K.; Germaine, N.; Ngwa, F.; Yamssi, C.; Megwi, L.; Florence, F.A.; Mbida, M.; Felix, B.B.C. Prevalence and intensity of infection of gastrointestinal parasites in cavies from Menoua division-west region of Cameroon. J. Agri. Ecol. Res. 2016, 5, 1–12. [Google Scholar] [CrossRef]
- Faihun, A.M.L.; Zoffoun, G.A.; Adenile, A.D.; Anago, D.E.; Hounzangbe-Adote, M.S. Gastrointestinal parasites of guinea pigs (Cavia porcellus) reared in different breeding systems in Benin. Livest. Res. Rural Dev. 2019, 31, 11. [Google Scholar]
- Suepaul, R.; Charles, C.; Dziva, F. Aerobic microflora and endoparasites of freshly shot wild agouti (Dasyprocta leporina) in Trinidad, West Indies. J. Zoo Wild. Med. 2016, 47, 1044–1048. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.R.; Garcia, G.W. A survey of the gastrointestinal parasites present in the Agouti (Dasyprocta leporina) reared intensively in Trinidad. Livest. Res. Rural Dev. 2017, 29, 1–7. [Google Scholar]
- Matamoro, Y.; Velazquez, J.; Pashov, B. Parasitos intestinalis del tepezcuinte, Agouti paca (Rodentia: Dasyproctidae) en Costa Rica. Rev. Biol. Trop. 1991, 39, 173–176. [Google Scholar]
- Ramirez-Hererra, O.; Rodriguez-Vivas, R.I.; Montes-Perez, R.; Torres-Acosta, J.F. Seguimiento annual de la parasitosis gastrointestinal del tepezcuintle, Agouti paca (Rodentia: Agoutidae) en cautiverio en la tropic mexicano. Rev. Biol. Trop. 2001, 49, 1171–1176. [Google Scholar]
- Eberhardt, A.T.; Robles, M.D.S.; Monje, L.D.; Beldomenico, P.M.; Callejon, R. A new Trichuris species (Nematoda: Trichuridae) from capybaras: Morphological-molecular characterization and polygenetic relationships. Acta Trop. 2019, 190, 244–252. [Google Scholar] [CrossRef]
- Jones, K.R.; Lall, K.R.; Garcia, G.W. Omnivorous Behaviour of the Agouti (Dasyprocta leporina): A Neotropical Rodent with the Potential for Domestication. Scientifica 2019, 2019, 3759783. [Google Scholar] [CrossRef]
- Garcia, G.W.; Baptiste, Q.S.; Adogwa, A.O.; Kakuni, M.; Arishima, K.; Makita, T. The digestive system of the agouti (Dasyprocta leporina)-gross anatomy and histology. Jpn. J. Zoo Wild. Med. 2000, 5, 55–66. [Google Scholar] [CrossRef]
- Lall, K.R.; Jones, K.R.; Garcia, G.W. Nutrition of six selected neo-tropical mammals in Trinidad and Tobago with the potential for domestication. Vet. Sci. 2018, 5, 52. [Google Scholar] [CrossRef] [PubMed]
- Brown-Uddenberg, R.C. Conceptualisation of an Intensive Production Model for the Agouti (Dasyprocta leporina) a Neotropical Rodent in Trinidad, West Indies. Ph.D. Thesis, University of the West Indies, Mona, Jamaica, 2001. [Google Scholar]
- Cameron, T.W.M.; Reesal, M.R. Studies on the endoparasitic fauna of Trinidad mammals. Can. J. Zool. 1951, 29, 276–289. [Google Scholar] [CrossRef]
- Goncalves, A.Q.; Bola, M.N.; Coura, J.R.; Pinto, R.M. New records of helminths if hystricomorphic rodent from the middle and high Rio Negro microregion, State of Amazonas, Brazil. Rev. Bras. Zool. 2006, 23, 716–726. [Google Scholar] [CrossRef]
- Noronha, D.; Vincente, J.J.; Pinto, R.M. A survey of new host records of nematodes form mammals deposited in the Helminthological Collection of the Oswaldo Cruz Institute (CHIOC). Rev. Bras. Zool. 2002, 19, 945–949. [Google Scholar] [CrossRef]
- Jones, K.R.; Garcia, G.W. Observations on the endoparasitic load in captive reared agoutis (Dasyprocta leporina) without anthelminthic exposure in Trinidad, Republic of Trinidad and Tobago. Livest. Res. Rural Dev. 2018, 30, 1–8. Available online: http://www.lrrd.org/lrrd30/10/kegan30181.html (accessed on 2 August 2020).
- Jones, K.R.; Lall, K.R.; Garcia, G.W. Haematological and Serum biochemical values of the agouti (Dasyprocta leporina) reared intensively in Trinidad, Republic of Trinidad and Tobago. Livest. Res. Rural Dev. 2019, 31, 1–8. [Google Scholar]
- Morales, G.A.; Guzman, V.H.; Wells, E.A.; Angel, D. Polycystic echinococcus in Columbia: The larval cestodes in infected rodents. J. Wild. Dis. 1979, 15, 421–428. [Google Scholar] [CrossRef]
- Gardner, S.L.; Rausch, R.L.; Camacho, O.C.J. Echinococcus vogeli Rausch and Bernstein, 1972, from the paca, Cuniculus paca L. (Rodentia: Dasyproctidae), in the Departmento de Santa Cruz, Bolivia. J. Parasitol. 1988, 74, 399–402. [Google Scholar] [CrossRef]
- Meneghelli, U.G.; Martinelli, A.L.C.; Velludo, M.A.S.L. Cisto de Echinococcus vogeli em figado de paca (Cuniculus paca) origniaria do estado do Acre, Brazil. Rev. Soc. Bras. Med. Trop. 1990, 23, 153–155. [Google Scholar] [CrossRef]
- Tantalean, M.V.; Angulo, J.V.; Martinez, R.R.; Diaz, S.M. First record of the Echinococcus vogeli (Cesotda, Taeniidae) metacestod in finding in Iquitos, Peru. Peruv. J. Parasitol. 2012, 20, 74–76. [Google Scholar]
- Almeida, F.; Caldas, R.; Corre, C.; Rodriguez- Silva, R.; Siqueira, N.; Machado-Silva, J.R. Co-infection of the cestode Echinoccocus vogeli and the nematode Calodium hepaticum in the hystricomorphic rodent Agouti paca from a forest reserve in Acre, Brazil. J. Helminthol. 2013, 87, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Ribiero, V.M.F.; de Souza, S.F.; Pinto, N.N.M.; Alves, A.L.F.; de Araujo Santos, F.G. Monitoring of the intestinal tract parasites load and of the sanitary management at the pacific breeding farm. Braz. Anim. Sci. 2015, 16, 608–614. [Google Scholar]
- Kiani, A.; Clauss, M.; Ortmann, S.; Vendl, C.; Congdon, E.R.; Herrera, E.A.; Kreuzer, M.; Schwarm, A. Digestive physiology of captive capybara (Hydrochoerus hydrochaeris). Zoo Biol. 2019, 38, 167–179. [Google Scholar] [CrossRef]
- Santarem, V.A.; Tostes, R.A.; Alberti, H.; Sanches, O.C. Fasciola hepatica in capybara. Acta Trop. 2006, 98, 311–313. [Google Scholar] [CrossRef]
- Bellato, V.; de Souza, A.P.; Sartor, A.P.; Veiga, L.P.H.N.; Centenaro, F. Fasciola hepatica occurrence in capybaras (Hydrochaeris hydrochaeris) and bovines (Bos taurus) in Timbo, SC, Brazil. Rev. Cienc. Agrovet. 2009, 8, 66–70. [Google Scholar]
- Dracz, R.M.; Ribeiro, V.M.A.; Pereira, C.A.J.; Lima, W.D.S. Occurrence of Fasciola hepatica (Linnaeus, 1758) in capybara (Hydrochoerus hydrochaeris) (Linnaeus, 1766) in Minas Gerais, Brazil. Braz. J. Vet. Parasitol. 2016, 25, 364–367. [Google Scholar] [CrossRef]
- Meireles, M.V.; Soares, R.M.; Bonello, F.; Gennari, S.M. Natural infection with zoonotic subtype of Cryptosporidium parvum in Capybara (Hydrochoerus hydrochaeris) from Brazil. Vet. Parasitol. 2007, 147, 166–170. [Google Scholar] [CrossRef]
- Casas, M.C.; Duszynski, D.W.; Zalles, L.M. Three new eimerians in capybara (Hydrochaeris hydrochaeris) populations form eastern Bolivia and southern Venezuela. J. Parasitol. 1995, 81, 247–251. [Google Scholar] [CrossRef]
- Silva, M.K.D.; da Silva, A.S.; Oliveira, C.B.; Soares, J.F.; Monteiro, S.G. Occorrencia de Eimeria ichiloensis em capivara (Hydrochaeris hydrochaeris) de criatorio. Arq. Cienc. Vet. Zool. 2007, 10, 129–131. [Google Scholar]
- Gurgel, A.C.F.; Sartori, A.D.S.; de Araujo, F.A.P. Eimeriosis in Capybara (Hydrochaeris hydrochaeris) in the state of Rio Grande do Sul, Brazil. Parasitol. Lat. 2007, 62, 76–78. [Google Scholar]
- Albuquerque, G.R.; Berto, B.P.; Catenacci, L.S.; Nogueira, S.S.C.; Nogueira-Filho, S.L.G.; Lopes, C.W.G. Eimerid coccidian from capybaras (Hydrochoerus hydrochaeris) in southern Bahia, Brazil. Presq. Vet. Bras. 2008, 26, 323–328. [Google Scholar] [CrossRef]
- Rodriguez-Duran, A.; Palma, L.C.B.; Florez, R.P. Main gastrointestinal protozoa in wild capybara (Hydrochoerus hydrochaeris) in a village in the municipality of Arauca, Columbia. Zootec. Trop. 2015, 33, 261–268. [Google Scholar]
- Sinkoc, A.L.; Brum, J.G.W.; Muller, G. Gastrointestinal Helminths of Capybara (Hydrochoerus hydrochaeris, Linnaeus, 1766) in Cattle Breeding Farms in the Area of the Ecological Reserve of Taim, Rio Grande. Braz. Arch. Biol. Technol. 2009, 52, 327–333. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | Geographical Location | Environment | Sample Size (n) | Prevalence (%) | Reference |
|---|---|---|---|---|---|
| Cavia porcellus | Benin | Captive reared | 18 | 2/18 (11.11) | [88] |
| Cavia porcellus | Peru | Captive reared | 400 | 20/400 (5.00) | [84] |
| Cavia porcellus | Peru | Captive reared | 100 | 31/100 (31.00) | [85] |
| Cavia porcellus | Cameroon | Captive reared | 397 | 4/397 (1.00) | [86] |
| Cavia porcellus | Cameroon | Captive reared | 300 | 10 (3.30) | [87] |
| Cavia aperera | Peru | Free range | 143 | 5/143 (3.50) | [83] |
| Dasyprocta leporina | Trinidad | Free range | 13 | 11/13 (53.85) | [89] |
| Dasyprocta leporina | Trinidad | Captive reared | 65 | 3/65 (4.62) | [90] |
| Agouti paca | Costa Rica | Captive reared | 140 | 3/140 (2.41) | [91] |
| Agouti paca | Mexico | Captive reared | 10 | 1/10 (10.00) | [92] |
| Hyrdochoerus hydrochaeris | Brazil | Captive reared | 24 | 12/24 (50.00) | [93] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jones, K.R. Trichuris spp. in Animals, with Specific Reference to Neo-Tropical Rodents. Vet. Sci. 2021, 8, 15. https://doi.org/10.3390/vetsci8020015
Jones KR. Trichuris spp. in Animals, with Specific Reference to Neo-Tropical Rodents. Veterinary Sciences. 2021; 8(2):15. https://doi.org/10.3390/vetsci8020015
Chicago/Turabian StyleJones, Kegan Romelle. 2021. "Trichuris spp. in Animals, with Specific Reference to Neo-Tropical Rodents" Veterinary Sciences 8, no. 2: 15. https://doi.org/10.3390/vetsci8020015
APA StyleJones, K. R. (2021). Trichuris spp. in Animals, with Specific Reference to Neo-Tropical Rodents. Veterinary Sciences, 8(2), 15. https://doi.org/10.3390/vetsci8020015