
Combined Cytotoxic Effects of the Fungicide Azoxystrobin and Common Food-Contaminating Mycotoxins




Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture
2.3. In Vitro Cytotoxicity
2.4. Experimental Design and Analysis of the Interaction of Chemical Mixtures
2.5. Statistical Analysis
3. Results
3.1. Cytotoxicity of Individual Components
3.2. Cytotoxicity of AZX and Mycotoxins Combinations
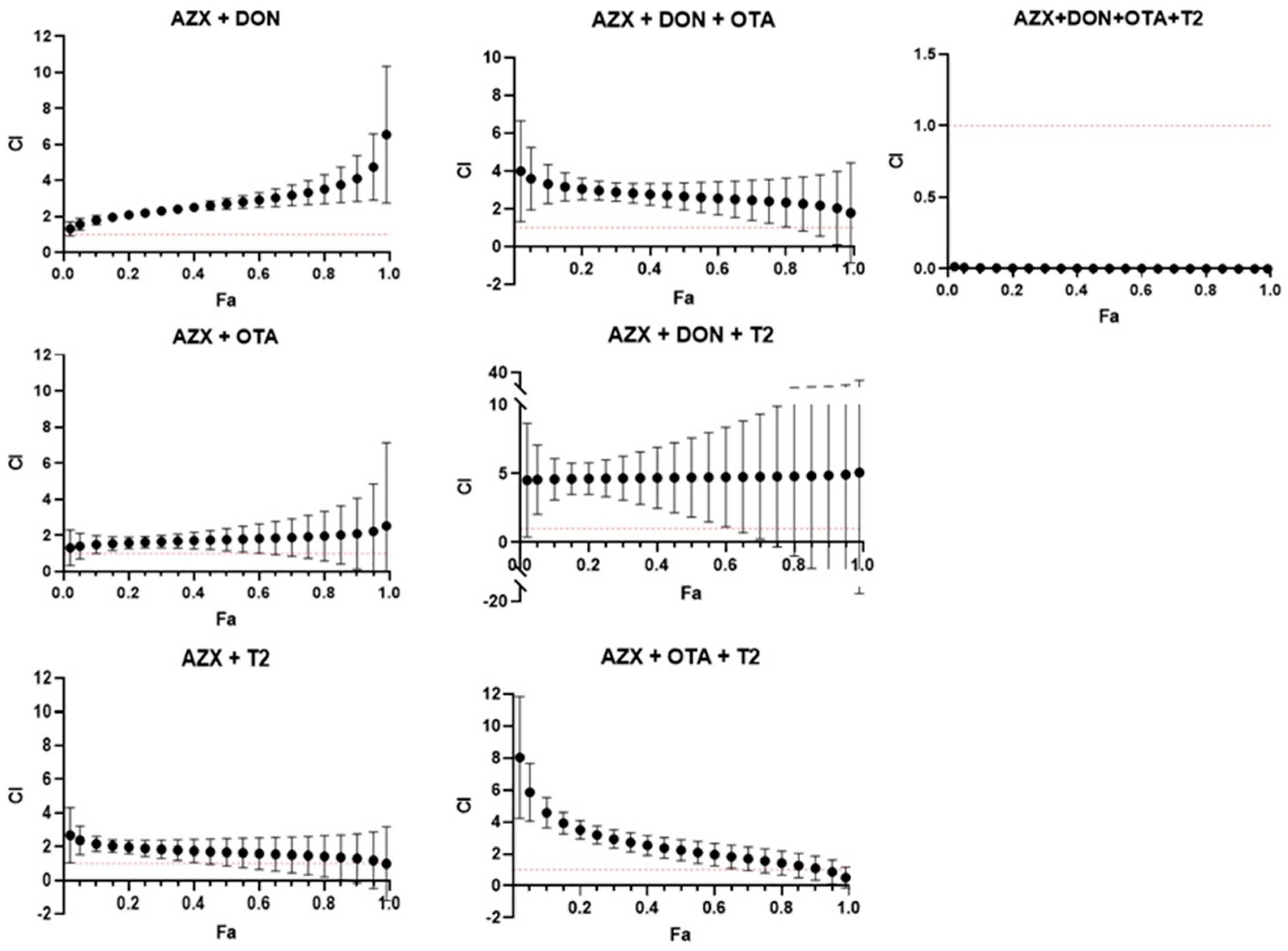
3.3. Toxicological Interactions Between AZX and Mycotoxins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AZX | azoxystrobin |
| CI | combination index |
| Dm | median–effect concentration |
| DMEM | Dulbecco’s Modified Eagle Medium |
| DMSO | dimethyl sulfoxide |
| DON | deoxynivalenol |
| EFSA | European Food Safety Authority |
| IC50 | mean inhibition concentration |
| MRL | maximum residue level |
| MTT | thiazolyl blue tetrazolium bromide |
| NBCS | Newborn Calf Serum |
| OTA | ochratoxin A |
| PBS | Phosphate-Buffered Saline |
| RASFF | Rapid Alert System for Food and Feed |
| SEM | standard error of the mean |
| T2 | T-2 toxin |
References
- Gupta, P.K. Chapter 24—Herbicides and Fungicides BT—Biomarkers in Toxicology. In Biomarkers in Toxicology; Academic Press: New York, NY, USA, 2014; pp. 409–431. ISBN 978-0-12-404630-6. [Google Scholar]
- FAOSTAT. Available online: https://www.fao.org/faostat/en/#data/RP (accessed on 17 September 2024).
- MarketsandMarkets Fungicides Market Size & Share, Trends and Forecast 2027. Available online: https://www.marketsandmarkets.com/Market-Reports/fungicides-356.html?gad_source=1&gclid=Cj0KCQjwrp-3BhDgARIsAEWJ6SyiQ9hRjFkJL4wcX4aIwmyTx1lyUH7MqkvLvLS0faGYjBYvQ0IWOlgaArnnEALw_wcB (accessed on 16 September 2024).
- Zingales, V.; Taroncher, M.; Martino, P.A.; Ruiz, M.J.; Caloni, F. Climate Change and Effects on Molds and Mycotoxins. Toxins 2022, 14, 445. [Google Scholar] [CrossRef] [PubMed]
- European Parliament and Council of the European Union. Regulation (EC) No 396/2005 of 23 February 2005 on maximum residue levels of pesticides in or on food and feed of plant and animal origin and amending Council Directive 91/414/EEC. Off. J. Eur. Union 2005, L70, 1–16. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32005R0396 (accessed on 30 March 2024).
- Carrasco Cabrera, L.; Di Piazza, G.; Dujardin, B.; Marchese, E.; Medina Pastor, P. The 2022 European Union Report on Pesticide Residues in Food; John Wiley and Sons: Hoboken, NJ, USA, 2024; Volume 22. [Google Scholar]
- Kopp, B.; Sanders, P.; Alassane-Kpembi, I.; Fessard, V.; Zalko, D.; Le Hégarat, L.; Audebert, M. Synergic Toxic Effects of Food Contaminant Mixtures in Human Cells. Mutagenesis 2020, 35, 415–424. [Google Scholar] [CrossRef]
- Silins, I.; Högberg, J. Combined Toxic Exposures and Human Health: Biomarkers of Exposure and Effect. Int. J. Environ. Res. Public Health 2011, 8, 629–647. [Google Scholar] [CrossRef]
- Hernández, A.F.; Gil, F.; Lacasaña, M. Toxicological Interactions of Pesticide Mixtures: An Update. Arch. Toxicol. 2017, 91, 3211–3223. [Google Scholar] [CrossRef]
- Thompson, P.A.; Khatami, M.; Baglole, C.J.; Sun, J.; Harris, S.A.; Moon, E.Y.; Al-Mulla, F.; Al-Temaimi, R.; Brown, D.G.; Colacci, A.M.; et al. Environmental Immune Disruptors, Inflammation and Cancer Risk. Carcinogenesis 2015, 36, S232–S253. [Google Scholar] [CrossRef]
- EC EUR-Lex-52012DC0252-EN; OPOCE: Brussels, Belgium, 2012.
- Drakvik, E.; Altenburger, R.; Aoki, Y.; Backhaus, T.; Bahadori, T.; Barouki, R.; Brack, W.; Cronin, M.T.D.; Demeneix, B.; Hougaard Bennekou, S.; et al. Statement on Advancing the Assessment of Chemical Mixtures and Their Risks for Human Health and the Environment. Environ. Int. 2020, 134, 105267. [Google Scholar] [CrossRef]
- Maino, J.L.; Thia, J.; Hoffmann, A.A.; Umina, P.A. Estimating Rates of Pesticide Usage from Trends in Herbicide, Insecticide, and Fungicide Product Registrations. Crop Prot. 2023, 163, 106125. [Google Scholar] [CrossRef]
- FAO/WHO. Joint Meeting of the FAO Panel of Experts on Pesticide Residues in Food and the Environment and the WHO Core Assessment Group on Pesticide Residues. Evaluation of Data for Acceptable Daily Intake and Acute Dietary Intake for Humans, Maximum Residue Levels, and Supervised Trial Median Residue Values: Azoxystrobin; FAO Plant Production and Protection Paper 232; FAO and WHO: Rome, Italy, 2017; Available online: https://www.fao.org/3/a-i7851e.pdf (accessed on 30 March 2024).
- Serra, R.S.; Martínez, L.C.; Cossolin, J.F.S.; de Resende, M.T.C.S.; Carneiro, L.S.; Fiaz, M.; Serrão, J.E. The Fungicide Azoxystrobin Causes Histopathological and Cytotoxic Changes in the Midgut of the Honey Bee Apis Mellifera (Hymenoptera: Apidae). Ecotoxicology 2023, 32, 234–242. [Google Scholar] [CrossRef]
- Pohanish, R.P. Sittig’s Handbook of Pesticides and Agricultural Chemicals, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar]
- EFSA. Conclusion on the Peer Review of the Pesticide Risk Assessment of the Active Substance Azoxystrobin. EFSA J. 2010, 8, 1542. [Google Scholar] [CrossRef]
- El-Sayed, R.A.; Jebur, A.B.; Kang, W.; El-Demerdash, F.M. An Overview on the Major Mycotoxins in Food Products: Characteristics, Toxicity, and Analysis. J. Future Foods 2022, 2, 91–102. [Google Scholar] [CrossRef]
- Awuchi, C.G.; Ondari, E.N.; Nwozo, S.; Odongo, G.A.; Eseoghene, I.J.; Twinomuhwezi, H.; Ogbonna, C.U.; Upadhyay, A.K.; Adeleye, A.O.; Okpala, C.O.R. Mycotoxins’ Toxicological Mechanisms Involving Humans, Livestock and Their Associated Health Concerns: A Review. Toxins 2022, 14, 167. [Google Scholar] [CrossRef] [PubMed]
- D’Mello, J.P.F.; Macdonald, A.M.C.; Rinna, R. Effects of Azoxystrobin on Mycotoxin Production in a Carbendazim-Resistant Strain of Fusarium Sporotrichioides. Phytoparasitica 2001, 29, 431–440. [Google Scholar] [CrossRef]
- Dorigan, A.F.; Moreira, S.I.; da Silva Costa Guimarães, S.; Cruz-Magalhães, V.; Alves, E. Target and Non-Target Site Mechanisms of Fungicide Resistance and Their Implications for the Management of Crop Pathogens. Pest Manag. Sci. 2023, 79, 4731–4753. [Google Scholar] [CrossRef] [PubMed]
- Lucas, J.A.; Hawkins, N.J.; Fraaije, B.A. The Evolution of Fungicide Resistance. Adv. Appl. Microbiol. 2015, 90, 29–92. [Google Scholar] [CrossRef]
- Giorni, P.; Rolla, U.; Romani, M.; Mulazzi, A.; Bertuzzi, T. Efficacy of Azoxystrobin on Mycotoxins and Related Fungi in Italian Paddy Rice. Toxins 2019, 11, 310. [Google Scholar] [CrossRef]
- Palladino, C.; Puigvert, F.; Muela, A.; Taborda, B.; Pérez, C.A.; Pérez-Parada, A.; Pareja, L. Evaluation of Fusarium Mycotoxins and Fungicide Residues in Barley Grain Produced in Uruguay. J. Agric. Food Res. 2021, 3, 100092. [Google Scholar] [CrossRef]
- Wang, R.; Huang, N.; Ji, J.; Chen, C. An Integrated Approach for Evaluating the Interactive Effects between Azoxystrobin and Ochratoxin A: Pathway-Based In Vivo Analyses. Pestic. Biochem. Physiol. 2023, 195, 105556. [Google Scholar] [CrossRef]
- Kortenkamp, A.; Faust, M. Regulate to Reduce Chemical Mixture Risk. Science 2018, 361, 224–226. [Google Scholar]
- Efeoglu, E.; Maher, M.A.; Casey, A.; Byrne, H.J. Label-Free, High Content Screening Using Raman Microspectroscopy: The Toxicological Response of Different Cell Lines to Amine-Modified Polystyrene Nanoparticles (PS-NH2). Analyst 2017, 142, 3500–3513. [Google Scholar] [CrossRef]
- Chou, T.C.; Talalay, P. Quantitative Analysis of Dose-Effect Relationships: The Combined Effects of Multiple Drugs or Enzyme Inhibitors. Adv. Enzym. Regul. 1984, 22, 27–55. [Google Scholar] [CrossRef]
- Chou, T.C. Frequently Asked Questions in Drug Combinations and the Mass-Action Law-Based Answers. Synergy 2014, 1, 3–21. [Google Scholar] [CrossRef]
- Kleber, A.; Gruber-Dorninger, C.; Platzer, A.; Payet, C.; Novak, B. Effect of Fungicide Treatment on Multi-Mycotoxin Occurrence in French Wheat during a 4-Year Period. Toxins 2023, 15, 443. [Google Scholar] [CrossRef] [PubMed]
- Groten, J.P.; Feron, V.J.; Sühnel, J. Toxicology of Simple and Complex Mixtures. Trends Pharmacol. Sci. 2001, 22, 316–322. [Google Scholar] [PubMed]
- Kamiloglu, S.; Sari, G.; Ozdal, T.; Capanoglu, E. Guidelines for Cell Viability Assays. Food Front. 2020, 1, 332–349. [Google Scholar] [CrossRef]
- Riss, T.L.; Niles, A.L.; Minor, L.; Moravec, R.A.; Benink, H.A.; Worzella, T.J. Assay Guidance Manual: Cell Viability Assays. In Assay Guidance Manual; Eli Lilly & Company and the National Center for Advancing Translational Sciences: Bethesda, MD, USA, 2013. [Google Scholar]
- Ghasemi, M.; Turnbull, T.; Sebastian, S.; Kempson, I. The Mtt Assay: Utility, Limitations, Pitfalls, and Interpretation in Bulk and Single-Cell Analysis. Int. J. Mol. Sci. 2021, 22, 2827. [Google Scholar] [CrossRef]
- Taroncher, M.; Rodríguez-Carrasco, Y.; Ruiz, M.J. T-2 Toxin and Its Metabolites: Characterization, Cytotoxic Mechanisms and Adaptive Cellular Response in Human Hepatocarcinoma (HepG2) Cells. Food Chem. Toxicol. 2020, 145, 111654. [Google Scholar] [CrossRef] [PubMed]
- Gayathri, L.; Dhivya, R.; Dhanasekaran, D.; Periasamy, V.S.; Alshatwi, A.A.; Akbarsha, M.A. Hepatotoxic Effect of Ochratoxin A and Citrinin, Alone and in Combination, and Protective Effect of Vitamin E: Invitro Study in HepG2 Cell. Food Chem. Toxicol. 2015, 83, 151–163. [Google Scholar] [CrossRef]
- Fernández-Blanco, C.; Elmo, L.; Waldner, T.; Ruiz, M.J. Cytotoxic Effects Induced by Patulin, Deoxynivalenol and Toxin T2 Individually and in Combination in Hepatic Cells (HepG2). Food Chem. Toxicol. 2018, 120, 12–23. [Google Scholar] [CrossRef]
- Skrzydlewski, P.; Twarużek, M.; Grajewski, J. Cytotoxicity of Mycotoxins and Their Combinations on Different Cell Lines: A Review. Toxins 2022, 14, 244. [Google Scholar] [CrossRef]
- Janik, E.; Niemcewicz, M.; Podogrocki, M.; Ceremuga, M.; Stela, M.; Bijak, M. T-2 Toxin—The Most Toxic Trichothecene Mycotoxin: Metabolism, Toxicity, and Decontamination Strategies. Molecules 2021, 26, 6868. [Google Scholar] [CrossRef]
- Almutairi, M.; Alsaleem, T.; Jeperel, H.; Alsamti, M.; Alowaifeer, A.M. Determination of Inorganic Arsenic, Heavy Metals, Pesticides and Mycotoxins in Indian Rice (Oryza sativa) and a Probabilistic Dietary Risk Assessment for the Population of Saudi Arabia. Regul. Toxicol. Pharmacol. 2021, 125, 104986. [Google Scholar] [CrossRef]
- Smith, M.C.; Madec, S.; Coton, E.; Hymery, N. Natural Co-Occurrence of Mycotoxins in Foods and Feeds and Their in Vitro Combined Toxicological Effects. Toxins 2016, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Bart, S.; Short, S.; Jager, T.; Eagles, E.J.; Robinson, A.; Badder, C.; Lahive, E.; Spurgeon, D.J.; Ashauer, R. How to Analyse and Account for Interactions in Mixture Toxicity with Toxicokinetic-Toxicodynamic Models. Sci. Total Environ. 2022, 843, 157048. [Google Scholar] [CrossRef] [PubMed]
- Altenburger, R.; Scholz, S.; Schmitt-Jansen, M.; Busch, W.; Escher, B.I. Mixture Toxicity Revisited from a Toxicogenomic Perspective. Environ. Sci. Technol. 2012, 46, 2508–2522. [Google Scholar] [PubMed]
- Cano-Sancho, G.; González-Arias, C.A.; Ramos, A.J.; Sanchis, V.; Fernández-Cruz, M.L. Cytotoxicity of the Mycotoxins Deoxynivalenol and Ochratoxin A on Caco-2 Cell Line in Presence of Resveratrol. Toxicol. Vitr. 2015, 29, 1639–1646. [Google Scholar] [CrossRef]
- Sobral, M.M.C.; Faria, M.A.; Cunha, S.C.; Ferreira, I.M.P.L.V.O. Toxicological Interactions between Mycotoxins from Ubiquitous Fungi: Impact on Hepatic and Intestinal Human Epithelial Cells. Chemosphere 2018, 202, 538–548. [Google Scholar] [CrossRef]
- Lei, Y.; Guanghui, Z.; Xi, W.; Yingting, W.; Xialu, L.; Fangfang, Y.; Goldring, M.B.; Xiong, G.; Lammi, M.J. Cellular Responses to T-2 Toxin and/or Deoxynivalenol That Induce Cartilage Damage Are Not Specific to Chondrocytes. Sci. Rep. 2017, 7, 2231. [Google Scholar] [CrossRef]
- Lin, X.; Shao, W.; Yu, F.; Xing, K.; Liu, H.; Zhang, F.; Goldring, M.B.; Lammi, M.J.; Guo, X. Individual and Combined Toxicity of T-2 Toxin and Deoxynivalenol on Human C-28/I2 and Rat Primary Chondrocytes. J. Appl. Toxicol. 2019, 39, 343–353. [Google Scholar] [CrossRef]
- Ruiz, M.J.; Macáková, P.; Juan-García, A.; Font, G. Cytotoxic Effects of Mycotoxin Combinations in Mammalian Kidney Cells. Food Chem. Toxicol. 2011, 49, 2718–2724. [Google Scholar] [CrossRef]
- Eze, U.A.; Huntriss, J.; Routledge, M.N.; Gong, Y.Y. Toxicological Effects of Regulated Mycotoxins and Persistent Organochloride Pesticides: In Vitro Cytotoxic Assessment of Single and Defined Mixtures on MA-10 Murine Leydig Cell Line. Toxicol. Vitr. 2018, 48, 93–103. [Google Scholar] [CrossRef]
- Lu, S.; Liu, S.; Cui, J.; Liu, X.; Zhao, C.; Fan, L.; Yin, S.; Hu, H. Combination of Patulin and Chlorpyrifos Synergistically Induces Hepatotoxicity via Inhibition of Catalase Activity and Generation of Reactive Oxygen Species. J. Agric. Food Chem. 2019, 67, 11474–11480. [Google Scholar] [CrossRef] [PubMed]
- Speijers, G.J.A.; Speijers, M.H.M. Combined Toxic Effects of Mycotoxins. Toxicol. Lett. 2004, 153, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Pescatore, T.; Di Nica, V.; Finizio, A.; Ademollo, N.; Spataro, F.; Rauseo, J.; Patrolecco, L. Sub-Lethal Effects of Soil Multiple Contamination on the Avoidance Behaviour of Eisenia Fetida. Ecotoxicol. Environ. Saf. 2021, 226, 112861. [Google Scholar] [CrossRef]
- Chentouf, A. Toxicological Evaluation of Complex Mixtures: Prediction and Interactions—A Review. Pharm. Pharmacol. Res. 2022, 5, 1–44. [Google Scholar] [CrossRef]
- Shi, X.K.; Bian, X.B.; Huang, T.; Wen, B.; Zhao, L.; Mu, H.X.; Fatima, S.; Fan, B.M.; Bian, Z.X.; Huang, L.F.; et al. Azoxystrobin Induces Apoptosis of Human Esophageal Squamous Cell Carcinoma KYSE-150 Cells through Triggering of the Mitochondrial Pathway. Front. Pharmacol. 2017, 8, 277. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Ma, J.; Cheng, Y.; Wang, Z.; Wang, G.; Jia, A.; Wang, H.; Sun, J.; Yan, Y. DON Induced DNA Damage Triggers Absence of P53-Mediated G2 Arrest and Apoptosis in IPEC-1 Cells. Toxicology 2024, 501, 153707. [Google Scholar] [CrossRef]
- Chu, C.S.; Chen, Y.T.; Liang, W.Z. Investigation of the Mechanisms behind Ochratoxin A-Induced Cytotoxicity in Human Astrocytes and the Protective Effects of N-Acetylcysteine. J. Appl. Toxicol. 2024, 44, 1454–1465. [Google Scholar] [CrossRef]
- Wang, R.; Liu, N.; Huang, N.; Shu, F.; Lou, Y.; Zhang, S.; Zhu, H.; Chen, C. Combined Toxicity Assessment of a Naturally Occurring Toxin and a Triazole Fungicide on Different Biological Processes through Toxicogenomic Data Mining with Mixtures. Pestic. Biochem. Physiol. 2023, 193, 105440. [Google Scholar] [CrossRef]
- Qin, L.T.; Lei, Y.X.; Liu, M.; Zeng, H.H.; Liang, Y.P.; Mo, L.Y. Toxic Interactions at the Physiological and Biochemical Levels of Green Algae under Stress of Mixtures of Three Azole Fungicides. Sci. Total Environ. 2024, 926, 171771. [Google Scholar] [CrossRef]
- Nian, M.; Braun, G.; Escher, B.I.; Fang, M. Toxicological Study of Human Exposure to Mixtures of Chemicals: Challenges and Approaches. Environ. Sci. Technol. Lett. 2024, 11, 773–782. [Google Scholar] [CrossRef]
- Autrup, H.; Bridges, J.; Greim, H.; Gard, A.; Janssen, C.; Rousselle, C.; Sanner, T.; Tarazona, J.; Testai, E.; Vermeire, T.; et al. Toxicity and Assessment of Chemical Mixtures; European Commission: Brussels, Belgium, 2011; Available online: https://ec.europa.eu/health/scientific_committees/environmental_risks/docs/scher_o_155.pdf (accessed on 27 March 2025).
- Noureddine, A.; Brinker, C.J. Pendant/Bridged/Mesoporous Silsesquioxane Nanoparticles: Versatile and Biocompatible Platforms for Smart Delivery of Therapeutics. Chem. Eng. J. 2018, 340, 125–147. [Google Scholar] [CrossRef]
- Fu, Y.; Yin, S.; Zhao, C.; Fan, L.; Hu, H. Combined Toxicity of Food-Borne Mycotoxins and Heavy Metals or Pesticides. Toxicon 2022, 217, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Onyeaka, H.; Ghosh, S.; Obileke, K.C.; Miri, T.; Odeyemi, O.A.; Nwaiwu, O.; Tamasiga, P. Preventing Chemical Contaminants in Food: Challenges and Prospects for Safe and Sustainable Food Production. Food Control 2024, 155, 110040. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration (µM) | |||||
|---|---|---|---|---|---|
| IC50/8 | IC50/4 | IC50/2 | IC50 | Ratio | |
| AZX | 25.76 | 51.53 | 103.05 | 206.10 | 1 |
| DON | 1.17 | 2.35 | 4.70 | 9.39 | 0.05 |
| OTA | 24.41 | 48.83 | 97.65 | 195.30 | 0.95 |
| T2 | 0.007 | 0.015 | 0.030 | 0.059 | 0.0003 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fuentes, C.; Zingales, V.; Barat, J.M.; Ruiz, M.-J. Combined Cytotoxic Effects of the Fungicide Azoxystrobin and Common Food-Contaminating Mycotoxins. Foods 2025, 14, 1226. https://doi.org/10.3390/foods14071226
Fuentes C, Zingales V, Barat JM, Ruiz M-J. Combined Cytotoxic Effects of the Fungicide Azoxystrobin and Common Food-Contaminating Mycotoxins. Foods. 2025; 14(7):1226. https://doi.org/10.3390/foods14071226
Chicago/Turabian StyleFuentes, Cristina, Veronica Zingales, José Manuel Barat, and María-José Ruiz. 2025. "Combined Cytotoxic Effects of the Fungicide Azoxystrobin and Common Food-Contaminating Mycotoxins" Foods 14, no. 7: 1226. https://doi.org/10.3390/foods14071226
APA StyleFuentes, C., Zingales, V., Barat, J. M., & Ruiz, M.-J. (2025). Combined Cytotoxic Effects of the Fungicide Azoxystrobin and Common Food-Contaminating Mycotoxins. Foods, 14(7), 1226. https://doi.org/10.3390/foods14071226