
Proximate, Physicochemical, Techno-Functional and Antioxidant Properties of Three Edible Insect (Gonimbrasia belina, Hermetia illucens and Macrotermes subhylanus) Flours




Abstract
:1. Introduction
2. Materials and Methods
2.1. Source of Materials
2.2. Preparation of Insect Flours
2.3. Proximate Composition Analysis
2.4. Determination of Physicochemical Properties
2.4.1. Evaluation of Colour Properties of Edible Insect Flours
2.4.2. Determination of Bulk Density
2.4.3. Determination of Water Activity
2.5. Determination of Techno-Functional Properties
2.5.1. Determination of Water Binding Capacity and Oil Binding Capacity
2.5.2. Determination of Emulsion Capacity and Emulsion Stability
2.5.3. Determination of Foam Capacity and Foam Stability
2.6. Determination of Antioxidant Activity
2.6.1. Preparation of Edible Insect Extract
2.6.2. Determination of DPPH Radical Scavenging Activity
2.6.3. Determination of ABTS+ Radical Scavenging Activity
2.6.4. Determination of Fe2+ Chelating Activity
2.6.5. Determination of Reducing Power
2.7. Statistical Analysis
3. Results and Discussion
3.1. Proximate Composition of Edible Insect Flours
3.2. Physicochemical Properties
3.2.1. Colour Properties of Edible Insect Flours
3.2.2. Bulk Density
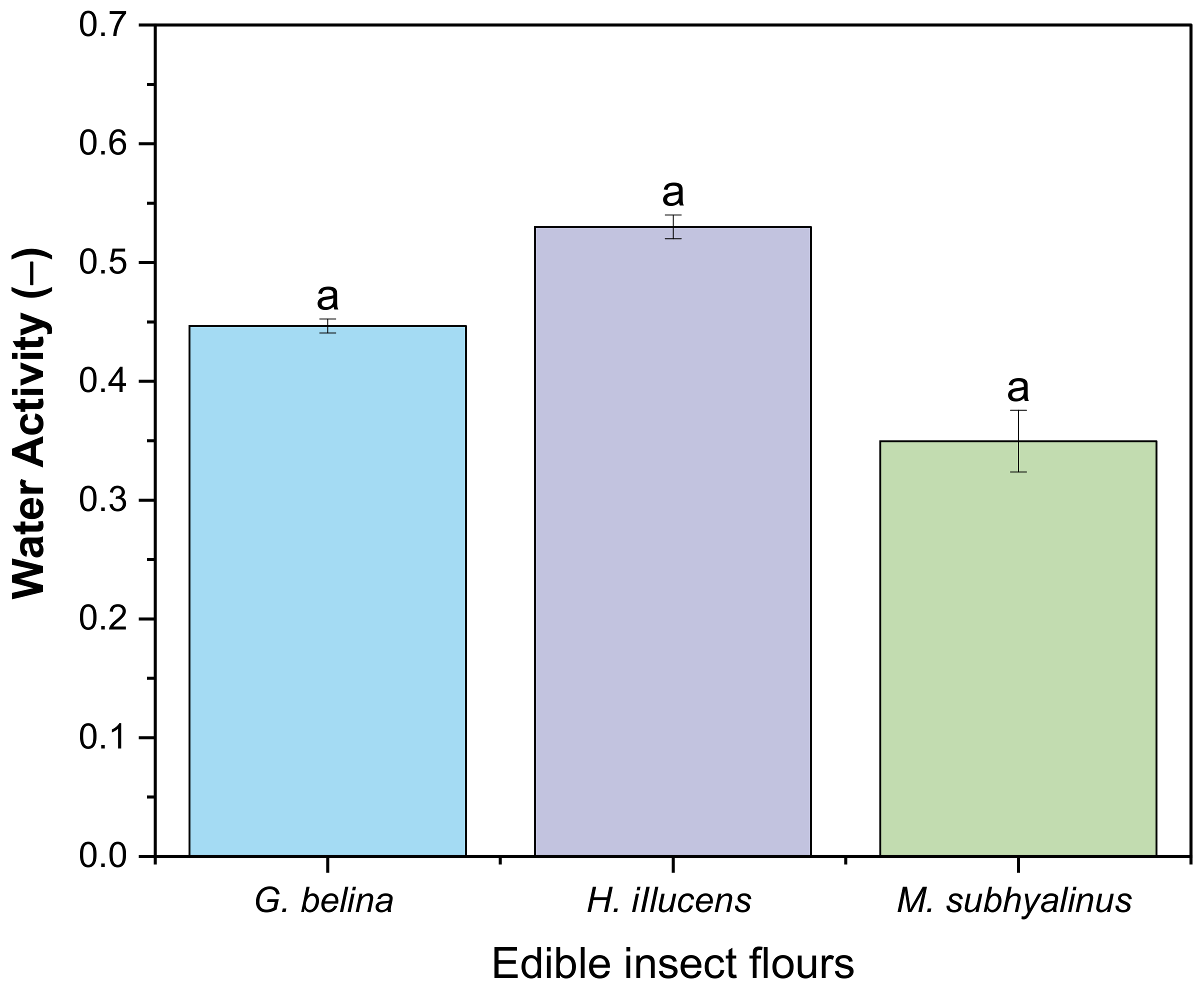
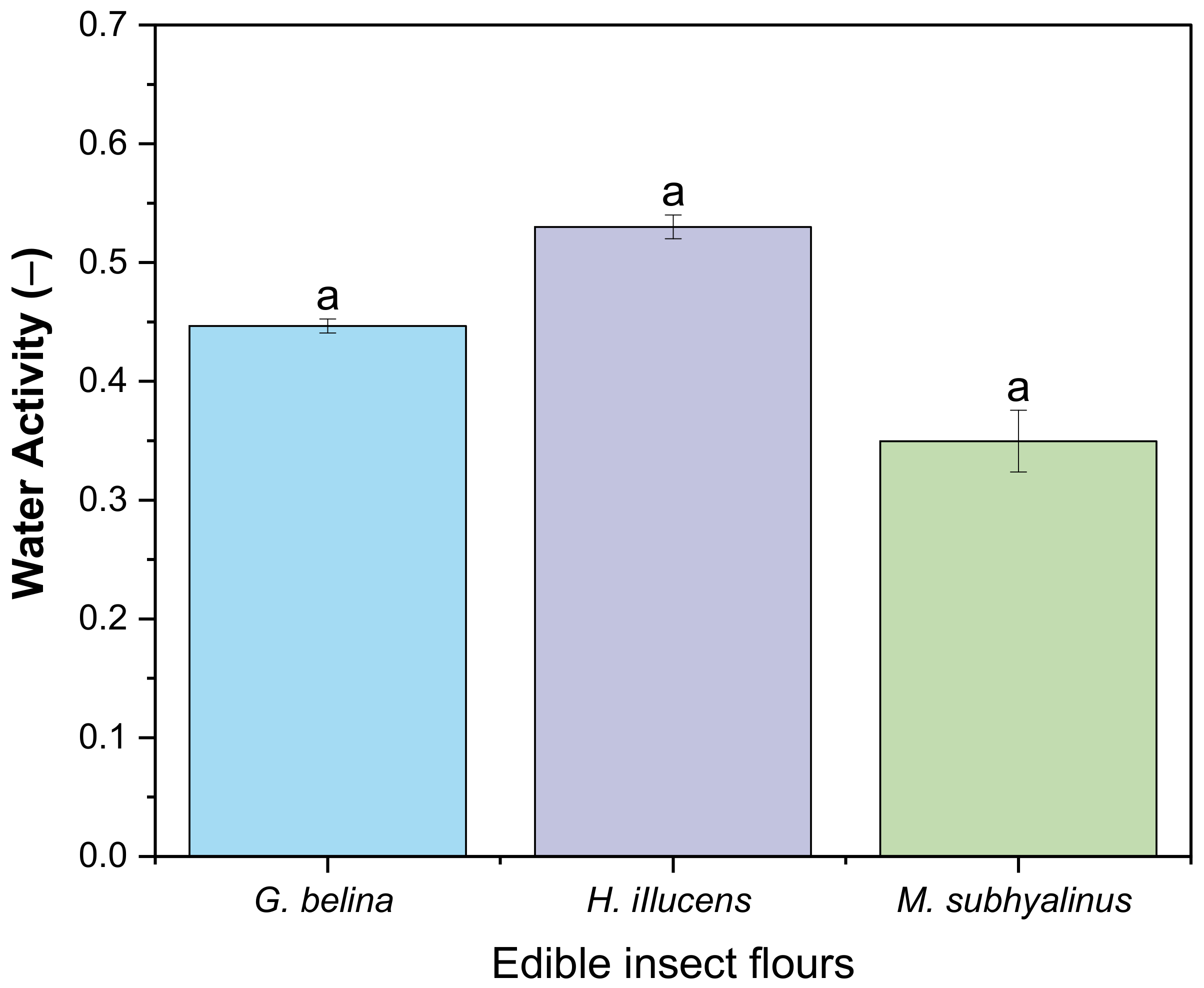
3.2.3. Water Activity and pH of Edible Insects
3.3. Techno-Functional Properties
3.3.1. Water Binding Capacity and Oil Binding Capacity
3.3.2. Emulsion Capacity and Emulsion Stability
3.3.3. Foam Capacity and Foam Stability
3.4. Antioxidant Properties
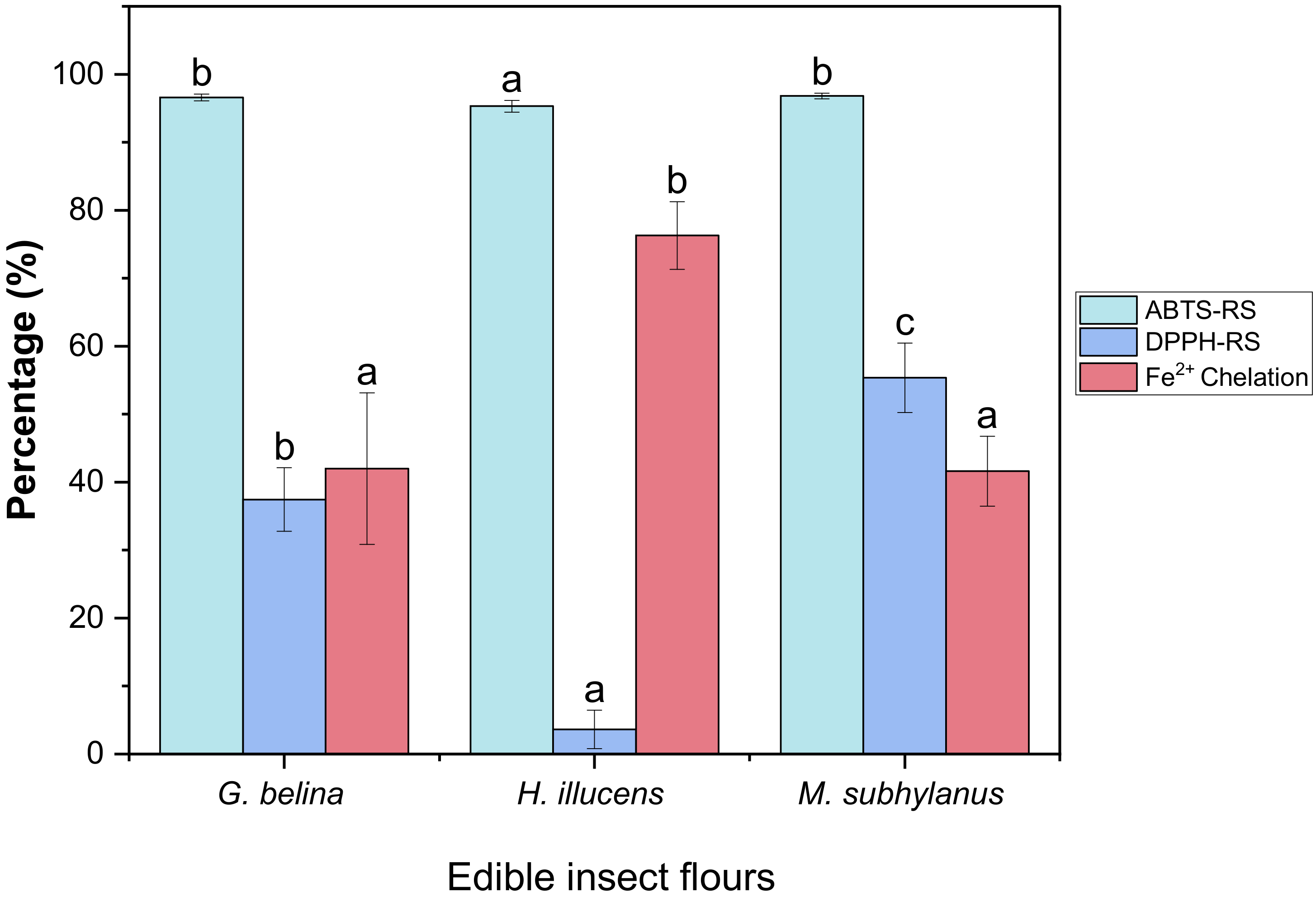
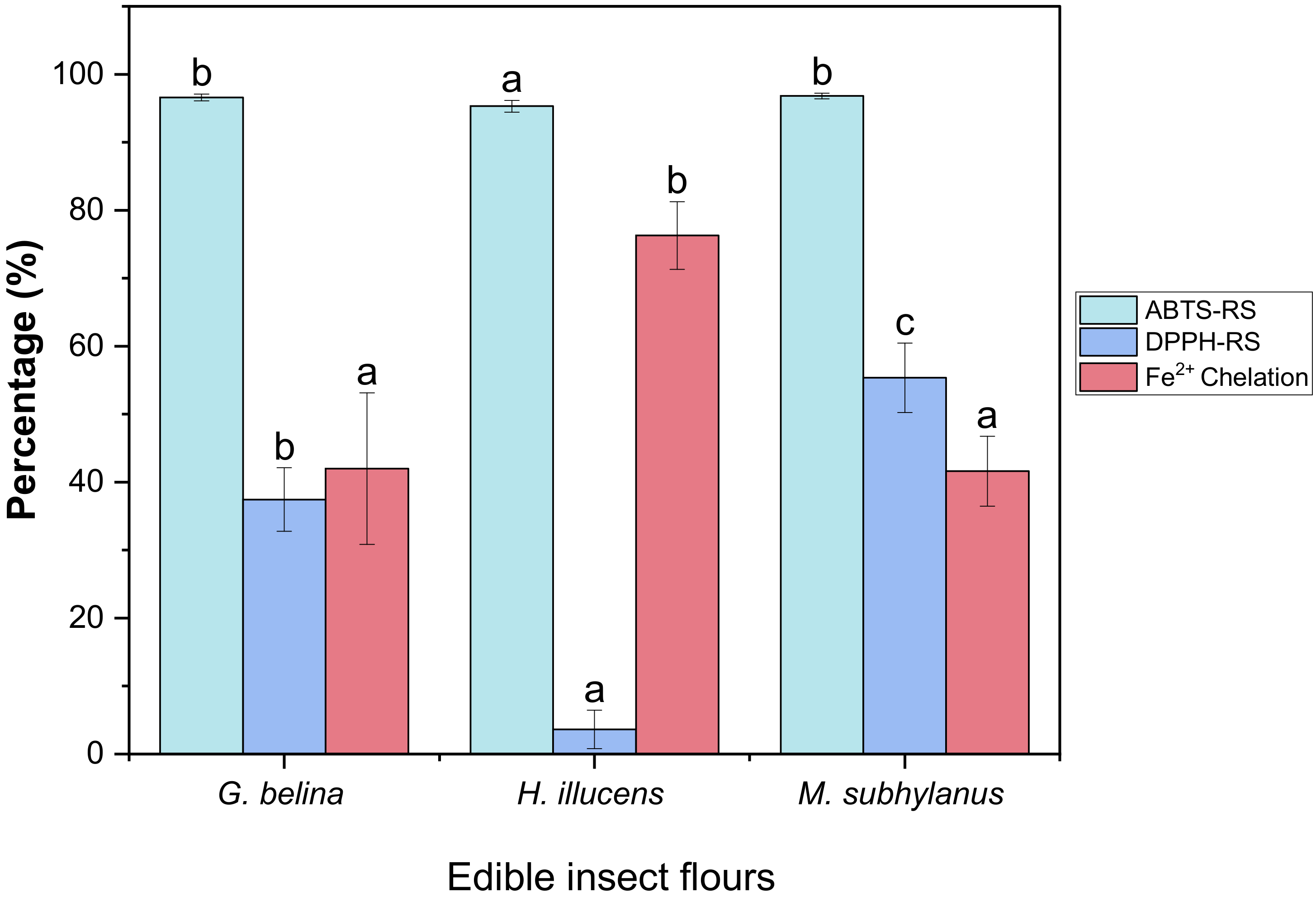
3.4.1. DPPH-RS of Edible Insect Flours
3.4.2. ABTS-RS of Edible Insect Flours
3.4.3. Metal Chelation of Edible Insect Flours
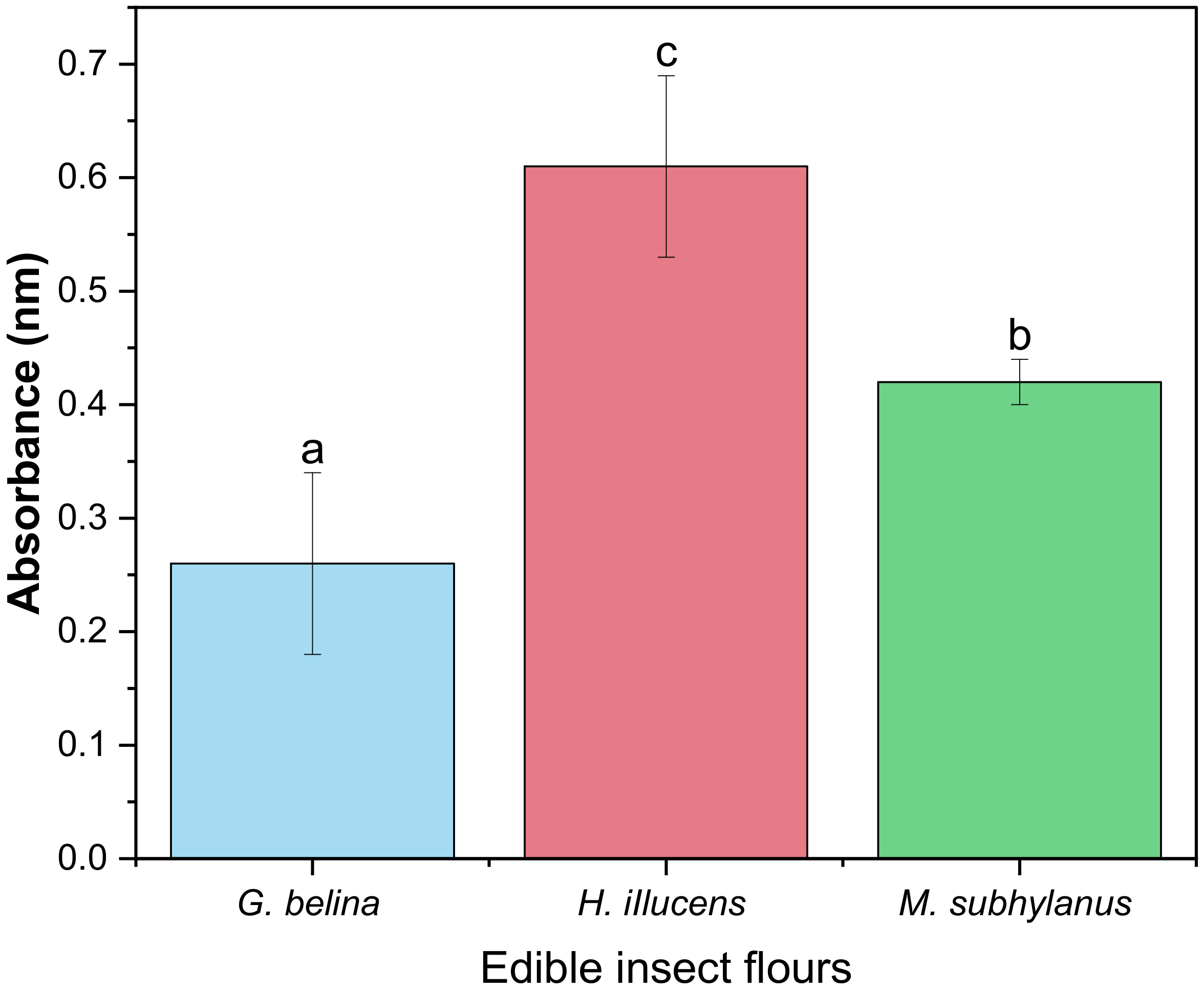
3.4.4. Reducing Power of Edible Insect Flours
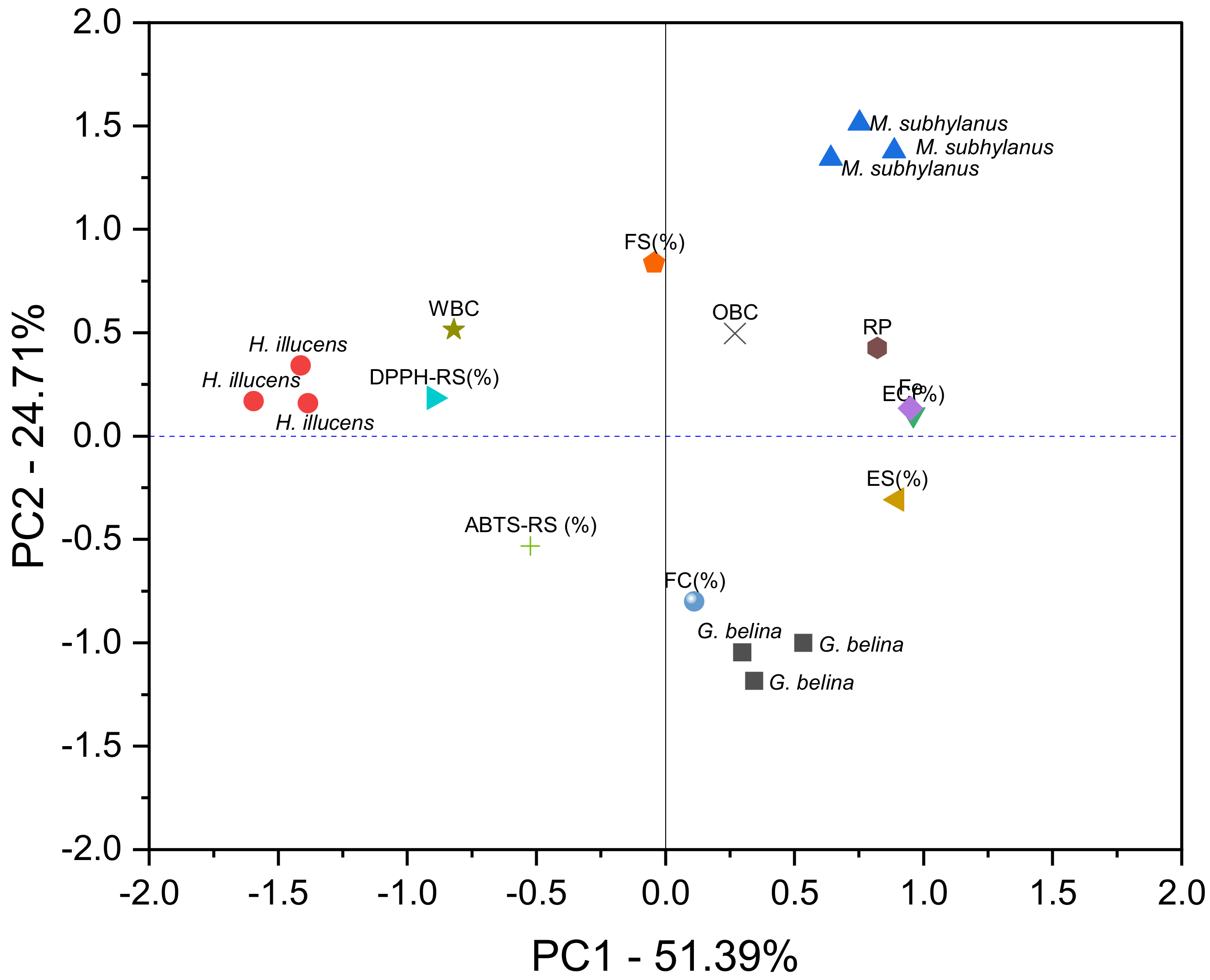
3.5. Principal Component Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gahukar, R.T. Entomophagy and human food security. Int. J. Trop. Insect Sci. 2011, 31, 129–144. [Google Scholar] [CrossRef] [Green Version]
- Imathiu, S. Benefits and food safety concerns associated with consumption of edible insects. NFS J. 2020, 18, 1–11. [Google Scholar] [CrossRef]
- Meyer-Rochow, V.B. Meyer-Rochow (1975) Can insects help to ease the problem of world food shortage. Search 1975, 6, 261–262. [Google Scholar]
- Hlongwane, Z.T.; Slotow, R.; Munyai, T.C. Indigenous knowledge about consumption of edible insects in South Africa. Insects 2021, 12, 22. [Google Scholar] [CrossRef] [PubMed]
- Kinyuru, J.N.; Konyole, S.O.; Roos, N.; Onyango, C.A.; Owino, V.O.; Owuor, B.O.; Estambale, B.B.; Friis, H.; Aagaard-Hansen, J.; Kenji, G.M. Nutrient composition of four species of winged termites consumed in western kenya. J. Food Compos. Anal. 2013, 30, 120–124. [Google Scholar] [CrossRef]
- Van-Huis, A.; Van-Itterbeeck, J.; Harmke, K.; Esther, M.; Afton, H.; Muir, M.; Paul, V. Edible Insects: Future Prospects for Food and Feed Security; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013; Available online: http://edepot.wur.nl/258042 (accessed on 8 January 2021).
- Kim, T.K.; Yong, H.I.; Kim, Y.B.; Kim, H.W.; Choi, Y.S. Edible insects as a protein source: A review of public perception, processing technology, and research trends. Food Sci. Anim. Resour. 2019, 39, 521–540. [Google Scholar] [CrossRef] [Green Version]
- Toti, E.; Massaro, L.; Kais, A.; Aiello, P.; Palmery, M.; Peluso, I. Entomophagy: A Narrative Review on Nutritional Value, Safety, Cultural Acceptance and a Focus on the Role of Food Neophobia in Italy. Eur. J. Investig. Health Psychol. Educ. 2020, 10, 46. [Google Scholar] [CrossRef]
- Orsi, L.; Voege, L.L.; Stranieri, S. Eating edible insects as sustainable food? Exploring the determinants of consumer acceptance in Germany. Food Res. Int. 2019, 125, 108573. [Google Scholar] [CrossRef]
- Raheem, D.; Raposo, A.; Oluwole, O.B.; Nieuwland, M.; Saraiva, A.; Carrascosa, C. Entomophagy: Nutritional, ecological, safety and legislation aspects. Food Res. Int. 2019, 126, 108672. [Google Scholar] [CrossRef]
- Bußler, S.; Rumpold, B.A.; Jander, E.; Rawel, H.M.; Schlüter, O.K. Recovery and techno-functionality of flours and proteins from two edible insect species: Meal worm (Tenebrio molitor) and black soldier fly (Hermetia illucens) larvae. Heliyon 2016, 2, e00218. [Google Scholar] [CrossRef]
- Van-Huis, A.; van Itterbeeck, J.; Klunder, H.; Esther, M.; Halloran, A.; Muir, G.; Vantomme, P. Edible Insects Future Prospects for Food and Feed Security; Food and agriculture organization of the United Nations: Rome, Italy, 2013. [Google Scholar]
- González, C.M.; Garzón, R.; Rosell, C.M. Insects as ingredients for bakery goods. A comparison study of H. illucens, A. domestica and T. molitor flours. Innov. Food Sci. Emerg. Technol. 2019, 51, 205–210. [Google Scholar] [CrossRef]
- Hlongwane, Z.T.; Slotow, R.; Munyai, T.C. Nutritional composition of edible insects consumed in africa: A systematic review. Nutrients 2020, 12, 2786. [Google Scholar] [CrossRef]
- Netshifhefhe, S.R.; Kunjeku, E.C.; Duncan, F.D. Human uses and indigenous knowledge of edible termites in Vhembe District, Limpopo Province, South Africa. South Afr. J. Sci. 2018, 114, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hall, F.G.; Jones, O.G.; O’Haire, M.E.; Liceaga, A.M. Functional properties of tropical banded cricket (Gryllodes sigillatus) protein hydrolysates. Food Chem. 2017, 224, 414–422. [Google Scholar] [CrossRef]
- Schösler, H.; de Boer, J.; Boersema, J.J. Can we cut out the meat of the dish? Constructing consumer-oriented pathways towards meat substitution. Appetite 2012, 58, 39–47. [Google Scholar] [CrossRef]
- Lucas-González, R.; Fernández-López, J.; Pérez-Álvarez, J.A.; Viuda-Martos, M. Effect of drying processes in the chemical, physico-chemical, techno-functional and antioxidant properties of flours obtained from house cricket (Acheta domesticus). Eur. Food Res. Technol. 2019, 245, 1451–1458. [Google Scholar] [CrossRef]
- Zozo, B.; Wicht, M.M.; Mshayisa, V.V.; Van-Wyk, J. Characterisation of black soldier fly larva protein before and after conjugation by the Maillard reaction. J. Insects Food Feed 2021, 8, 169–183. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC, 15th ed.; Association of Official Agricultural Chemists: Arlington, VA, USA, 2015. [Google Scholar] [CrossRef]
- Janssen, R.H.; Vincken, J.P.; van den Broek, L.A.M.; Fogliano, V.; Lakemond, C.M.M. Nitrogen-to-Protein Conversion Factors for Three Edible Insects: Tenebrio molitor, Alphitobius diaperinus, and Hermetia illucens. J. Agric. Food Chem. 2017, 65, 2275–2278. [Google Scholar] [CrossRef]
- Farzana, T.; Mohajan, S. Effect of incorporation of soy flour to wheat flour on nutritional and sensory quality of biscuits fortified with mushroom. Food Sci. Nutr. 2015, 3, 363–369. [Google Scholar] [CrossRef]
- Larouche, J.; Deschamps, M.H.; Saucier, L.; Lebeuf, Y.; Doyen, A.; Vandenberg, G.W. Effects of killing methods on lipid oxidation, colour and microbial load of black soldier fly (Hermetia illucens) larvae. Animals 2019, 9, 182. [Google Scholar] [CrossRef] [Green Version]
- Mintah, B.K.; He, R.; Agyekum, A.A.; Dabbour, M.; Golly, M.K.; Ma, H. Edible insect protein for food applications: Extraction, composition, and functional properties. J. Food Process Eng. 2020, 43, 1–12. [Google Scholar] [CrossRef]
- Benamara, R.N.; Gemelas, L.; Ibri, K.; Moussa-Boudjemaa, B.; Demarigny, Y. Sensory, microbiological and physico-chemical characterization of Klila, a traditional cheese made in the south-west of Algeria. Afr. J. Microbiol. Res. 2016, 10, 1728–1738. [Google Scholar] [CrossRef]
- Mshayisa, V.V.; Van-Wyk, J. Hermetia illucens Protein Conjugated with Glucose via Maillard Reaction: Antioxidant and Techno-Functional Properties. Int. J. Food Sci. 2021, 2021, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zielińska, E.; Karaś, M.; Baraniak, B. Comparison of functional properties of edible insects and protein preparations thereof. LWT-Food Sci. Technol. 2018, 91, 168–174. [Google Scholar] [CrossRef]
- Vhangani, L.N.; Van-Wyk, J. Antioxidant activity of Maillard reaction products (MRPs) derived from fructose-lysine and ribose-lysine model systems. Food Chem. 2013, 137, 92–98. [Google Scholar] [CrossRef]
- Chatsuwan, N.; Nalinanon, S.; Puechkamut, Y.; Lamsal, B.P.; Pinsirodom, P. Characteristics, Functional Properties, and Antioxidant Activities of Water-Soluble Proteins Extracted from Grasshoppers, Patanga succincta and Chondracris roseapbrunner. J. Chem. 2018, 2018, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Sudan, R.; Bhagat, M.; Gupta, S.; Singh, J.; Koul, A. Iron (FeII) Chelation, Ferric Reducing Antioxidant Power, and Immune Modulating Potential of Arisaema jacquemontii (Himalayan Cobra Lily). BioMed Res. Int. 2014, 2014, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Athukorala, Y.; Kim, K.N.; Jeon, Y.J. Antiproliferative and antioxidant properties of an enzymatic hydrolysate from brown alga, Ecklonia cava. Food Chem. Toxicol. 2006, 44, 1065–1074. [Google Scholar] [CrossRef]
- Churchward-Venne, T.A.; Pinckaers, P.J.M.; van Loon, J.J.A.; van Loon, L.J.C. Consideration of insects as a source of dietary protein for human consumption. Nutr. Rev. 2017, 75, 1035–1045. [Google Scholar] [CrossRef]
- Meyer-Rochow, V.B.; Gahukar, R.T.; Ghosh, S.; Jung, C. Chemical composition, nutrient quality and acceptability of edible insects are affected by species, developmental stage, gender, diet, and processing method. Foods 2021, 10, 1036. [Google Scholar] [CrossRef]
- Kwiri, R.; Winini, C.; Muredzi, P.; Tongonya, J.; Gwala, W.; Mujuru, F.; Gwala, S.T. Mopane Worm (Gonimbrasia belina) Utilisation, a Potential Source of Protein in Fortified Blended Foods in Zimbabwe: A Review. Glob. J. Sci. Front. Res. D Agric. Vet. 2014, 14, 55–66. [Google Scholar]
- Montowska, M.; Kowalczewski, P.Ł.; Rybicka, I.; Fornal, E. Nutritional value, protein and peptide composition of edible cricket powders. Food Chem. 2019, 289, 130–138. [Google Scholar] [CrossRef]
- Anaduaka, E.G.; Uchendu, N.O.; Osuji, D.O.; Ene, L.N.; Amoke, O.P. Nutritional compositions of two edible insects: Oryctes rhinoceros larva and Zonocerus variegatus. Heliyon 2021, 7, e06531. [Google Scholar] [CrossRef]
- Omotoso, O. Nutrient Composition, Mineral Analysis and Anti-nutrient Factors of Oryctes rhinoceros L. (Scarabaeidae: Coleoptera) and Winged Termites, Marcrotermes nigeriensis Sjostedt. (Termitidae: Isoptera). Br. J. Appl. Sci. Technol. 2015, 8, 97–106. [Google Scholar] [CrossRef]
- Adepoju, O.; Omotayo, O. Nutrient Composition and Potential Contribution of Winged Termites (Marcrotermes bellicosus Smeathman) to Micronutrient Intake of Consumers in Nigeria. Br. J. Appl. Sci. Technol. 2014, 4, 1149–1158. [Google Scholar] [CrossRef]
- Torruco-Uco, J.G.; Hernández-Santos, B.; Herman-Lara, E.; Martínez-Sánchez, C.E.; Juárez-Barrientos, J.M.; Rodríguez-Miranda, J. Chemical, functional and thermal characterization, and fatty acid profile of the edible grasshopper (Sphenarium purpurascens Ch.). Eur. Food Res. Technol. 2018, 245, 285–292. [Google Scholar] [CrossRef]
- Nyakeri, E.M.; Ogola, H.J.; Ayieko, M.A.; Amimo, F.A. An open system for farming black soldier fly larvae as a source of proteins for smallscale poultry and fish production. J. Insects Food Feed. 2017, 3, 51–56. [Google Scholar] [CrossRef]
- Clarkson, C.; Mirosa, M.; Birch, J. Potential of extracted Locusta migratoria protein fractions as value-added ingredients. Insects 2018, 9, 20. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, S.; Jung, C.; Meyer-Rochow, V.B. Nutritional value and chemical composition of larvae, pupae, and adults of worker honey bee, Apis mellifera ligustica as a sustainable food source. J. Asia-Pac. Entomol. 2016, 19, 487–495. [Google Scholar] [CrossRef]
- Siulapwa, N.; Mwambungu, A.; Lungu, E.; Sichilima, W. Nutritional Value of Four Common Edible Insects in Zambia. Int. J. Sci. Res. 2012, 3, 2319–7064. [Google Scholar]
- Payne, C.L.R.; Scarborough, P.; Rayner, M.; Nonaka, K. A systematic review of nutrient composition data available for twelve commercially available edible insects, and comparison with reference values. Trends Food Sci. Technol. 2016, 47, 69–77. [Google Scholar] [CrossRef]
- Ganguly, A.; Chakravorty, R.; Das, M.; Gupta, M.; Mandal, D.K.; Haldar, P.; Ramos-Elorduy, J.; Moreno, J.M.P. A preliminary study on the estimation of nutrients and anti-nutrients in Oedaleus abruptus (Thunberg) (Orthoptera-Acridida). Int. J. Nutr. Metab. 2013, 5, 50–65. [Google Scholar] [CrossRef]
- Melo, V.; Garcia, M.; Sandoval, H.; Jiménez, H.D.; Calvo, C. Quality proteins from edible indigenous insect food of latin America and Asia. Emir. J. Food Agric. 2011, 23, 283–289. [Google Scholar]
- Sogbesan, A.O.; Ugwumba, A.A.A. Nutritional Evaluation of Termite (Macrotermes subhyalinus) Meal as Animal Protein Supplements in the Diets of Heterobranchus longifilis (Valenciennes, 1840) Fingerlings. Turk. J. Fish. Aquat. Sci. 2008, 8, 149–157. [Google Scholar]
- Rodríguez-Miranda, J.; Alcántar-Vázquez, J.P.; Zúñiga-Marroquín, T.; Juárez-Barrientos, J.M. Insects as an alternative source of protein: A review of the potential use of grasshopper (Sphenarium purpurascens Ch.) as a food ingredient. Eur. Food Res. Technol. 2019, 245, 2613–2620. [Google Scholar] [CrossRef]
- Mishyna, M.; Martinez, J.J.I.; Chen, J.; Benjamin, O. Extraction, characterization and functional properties of soluble proteins from edible grasshopper (Schistocerca gregaria) and honey bee (Apis mellifera). Food Res. Int. 2019, 116, 697–706. [Google Scholar] [CrossRef]
- Manera, F.J.; Legua, P.; Melgarejo, P.; Martínez, R.; Martínez, J.J.; Hernández, F. Effect of air temperature on rind colour development in pomegranates. Sci. Hortic. 2012, 134, 245–247. [Google Scholar] [CrossRef]
- Piornos, J.A.; Burgos-Díaz, C.; Ogura, T.; Morales, E.; Rubilar, M.; Maureira-Butler, I.; Salvo-Garrido, H. Functional and physicochemical properties of a protein isolate from AluProt-CGNA: A novel protein-rich lupin variety (Lupinus luteus). Food Res. Int. 2015, 76, 719–724. [Google Scholar] [CrossRef]
- Sharma, A.; Jana, A.H.; Chavan, R.S. Functionality of Milk Powders and Milk-Based Powders for End Use Applications-A Review. Compr. Rev. Food Sci. Food Saf. 2012, 11, 518–528. [Google Scholar] [CrossRef]
- Akpossan, R.; Digbeu, Y.; Koffi, M.; Kouadio, J.; Dué, E.; Kouamé, P. Protein fractions and functional properties of dried Imbrasia oyemensis larvae full-fat and defatted flours. Int. J. Biochem. Res. Rev. 2015, 5, 116–126. [Google Scholar] [CrossRef]
- Ekpo, K.E.E.; Ugbenyen, M.A.A.; Azeke, M.A.; Ugbenyen, A.M.; Ugbenyen, M.A.A.; Azeke, M.A.; Ugbenyen, A.M.; Ugbenyen, M.A.A.; Azeke, M.A.; Ugbenyen, A.M. Functional Properties of dried Imbrasia Belina larvae flour as affected by mesh size and pH. Niger. Ann. Nat. Sci. 2008, 8, 6–9. [Google Scholar]
- Gupta, A.; Sharama, S.; Singh, B. Influence of Germination Conditions on the Techno-functional Properties of Amaranth flour. In Proceedings of the 2018 International Conference on Food Properties, Sharjah, United Arab Emirates, 22–24 January 2018. [Google Scholar]
- Sandulachi, E.; Tatarov, P. Water Activity Concept and Its Role in Strawberries Food. Chem. J. Mold. 2012, 7, 103–115. [Google Scholar] [CrossRef]
- Tapia, M.S.; Alzamora, S.M.; Chirife, J. Effects of Water Activity (aw) on Microbial Stability: As a Hurdle in Food Preservation. Water Act. Foods Fundam. Appl. 2008, 239–271. [Google Scholar] [CrossRef]
- Wang, J.; Jousse, M.; Jayakumar, J.; Fernández-Arteaga, A.; de Lamo-Castellví, S.; Ferrando, M.; Güell, C. Black soldier fly (Hermetia illucens) protein concentrates as a sustainable source to stabilize o/w emulsions produced by a low-energy high-throughput emulsification technology. Foods 2021, 10, 1048. [Google Scholar] [CrossRef]
- Chandra, S. Assessment of functional properties of different flours. Afr. J. Agric. Res. 2013, 8, 4849–4852. [Google Scholar] [CrossRef]
- Assielou, B.; Due, E.; Koffi, M.; Dabonne, S.; Kouame, P. Oryctes owariensis Larvae as Good Alternative Protein Source: Nutritional and Functional Properties. Annu. Res. Rev. Biol. 2015, 8, 1–9. [Google Scholar] [CrossRef]
- Akubor, P.I.; Ike, E.Z.E.J. Quality evaluation and cake making potential of sun and oven dried carrot fruit. Int. J. Biosci. 2012, 2, 19–27. [Google Scholar]
- Lucas, A.J.d.; de Oliveira, L.M.; da Rocha, M.; Prentice, C. Edible insects: An alternative of nutritional, functional and bioactive compounds. Food Chem. 2020, 311, 126022. [Google Scholar] [CrossRef]
- Adebowale, Y.A.; Adeyemi, I.A.; Oshodi, A.A. Functional and physicochemical properties of flours of six Mucuna species. Afr. J. Biotechnol. 2005, 4, 1461–1468. [Google Scholar] [CrossRef]
- Ndiritu, A.K.; Kinyuru, J.N.; Kenji, G.M.; Gichuhi, P.N. Extraction technique influences the physico-chemical characteristics and functional properties of edible crickets (Acheta domesticus) protein concentrate. J. Food Meas. Charact. 2017, 11, 2013–2021. [Google Scholar] [CrossRef]
- Omotoso, O.T. Nutritional quality, functional properties and anti-nutrient compositions of the larva of Cirina forda (Westwood) (Lepidoptera: Saturniidae). J. Zhejiang Univ. Sci. B 2005, 7, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Gravel, A.; Doyen, A. The use of edible insect proteins in food: Challenges and issues related to their functional properties. Innov. Food Sci. Emerg. Technol. 2020, 59, 2–11. [Google Scholar] [CrossRef]
- Liang, N.; Kitts, D.D. Antioxidant property of coffee components: Assessment of methods that define mechanism of action. Molecules 2014, 19, 19180–19208. [Google Scholar] [CrossRef] [Green Version]
- Khaled, H.B.; Ktari, N.; Ghorbel-Bellaaj, O.; Jridi, M.; Lassoued, I.; Nasri, M. Composition, functional properties and in vitro antioxidant activity of protein hydrolysates prepared from sardinelle (Sardinella aurita) muscle. J. Food Sci. Technol. 2014, 51, 622–633. [Google Scholar] [CrossRef] [Green Version]
- Del Hierro, J.N.; Gutiérrez-Docio, A.; Otero, P.; Reglero, G.; Martin, D. Characterization, antioxidant activity, and inhibitory effect on pancreatic lipase of extracts from the edible insects Acheta domesticus and Tenebrio molitor. Food Chem. 2020, 309, 125742. [Google Scholar] [CrossRef]
- Nabil, B.; Ouaabou, R.; Ouhammou, M.; Essaadouni, L.; Mahrouz, M. Functional Properties, Antioxidant Activity, and Organoleptic Quality of Novel Biscuit Produced by Moroccan Cladode Flour “Opuntia ficus-indica”. J. Food Qual. 2020, 2020, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Chalamaiah, M.; Kumar, B.D.; Hemalatha, R.; Jyothirmayi, T. Fish protein hydrolysates: Proximate composition, amino acid composition, antioxidant activities and applications: A review. Food Chem. 2012, 135, 3020–3038. [Google Scholar] [CrossRef]
- Khantaphant, S.; Benjakul, S.; Ghomi, M.R. The effects of pretreatments on antioxidative activities of protein hydrolysate from the muscle of brownstripe red snapper (Lutjanus vitta). LWT-Food Sci. Technol. 2011, 44, 1139–1148. [Google Scholar] [CrossRef]
- Nooshkam, M.; Varidi, M.; Bashash, M. The Maillard reaction products as food-born antioxidant and antibrowning agents in model and real food systems. Food Chem. 2019, 275, 644–660. [Google Scholar] [CrossRef]
- Khantaphant, S.; Benjakul, S. Comparative study on the proteases from fish pyloric caeca and the use for production of gelatin hydrolysate with antioxidative activity. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2008, 151, 410–419. [Google Scholar] [CrossRef]
- Zielińska, E.; Pankiewicz, U. Characteristics of Shortcake Biscuits Enriched with Tenebrio molitor Flour. Molecules 2020, 25, 5629. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Edible Insects | Crude Protein (%) | Ash (%) | Moisture (%) | Crude Fat (%) | Carbohydrates (%) | Energy (%) |
|---|---|---|---|---|---|---|
| G. belina | 46.70 ± 0.82 b | 11.38 ± 2.20 b | 5.68 ± 0.25 a | 14.04 ± 0.12 b | 22.10 ± 1.45 a | 399.38 ± 6.03 a |
| H. illucens | 34.90 ± 0.47 a | 7.50 ± 1.65 a | 5.76 ± 0.01 ab | 27.93 ± 6.13 c | 23.66 ± 7.84 a | 485.58 ± 26.69 b |
| M. subhylanus | 52.74 ± 1.47 c | 6.41 ± 0.07 a | 6.40 ± 0.06 b | 6.36 ± 0.05 a | 27.27 ± 1.19 a | 379.91 ± 1.06 a |
| Edible Insects | L* | a* | b* | Bulk Density (g/mL) | pH |
|---|---|---|---|---|---|
| G. belina | 57.95 ± 0.31 c | 3.92 ± 1.49 a | 20.02 ± 1.97 b | 0.65 ± 0.01 b | 6.12 ± 0.03 a |
| H. illucens | 53.69 ± 0.54 b | 4.46 ± 0.36 a | 13.08 ± 2.68 a | 0.51 ± 0.01 a | 8.93 ± 0.05 b |
| M. subhylanus | 43.52 ± 0.56 a | 5.72 ± 3.90 a | 12.00 ± 2.70 a | 0.64 ± 0.00 b | 6.14 ± 0.02 a |
| Edible Insects | WBC (g/g) | OBC (g/g) | EC (%) | ES (%) | FC (%) | FS (%) |
|---|---|---|---|---|---|---|
| G. belina | 1.30 ± 0.12 ab | 0.89 ± 0.12 a | 41.76 ± 2.84 a | 33.75 ± 2.29 a | 5.81 ± 3.69 a | 95.32 ± 2.37 a |
| H. illucens | 0.11 ± 0.02 a | 1.35 ± 0.09 b | 67.33 ± 8.49 b | 42.45 ± 5.07 b | 5.69 ± 1.41 a | 97.38 ± 1.70 a |
| M. subhylanus | 1.46 ± 0.06 b | 1.48 ± 0.07 b | 45.44± 4.28 a | 32.80 ± 0.47 a | 4.71 ± 2.46 a | 97.51 ± 1.22 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vanqa, N.; Mshayisa, V.V.; Basitere, M. Proximate, Physicochemical, Techno-Functional and Antioxidant Properties of Three Edible Insect (Gonimbrasia belina, Hermetia illucens and Macrotermes subhylanus) Flours. Foods 2022, 11, 976. https://doi.org/10.3390/foods11070976
Vanqa N, Mshayisa VV, Basitere M. Proximate, Physicochemical, Techno-Functional and Antioxidant Properties of Three Edible Insect (Gonimbrasia belina, Hermetia illucens and Macrotermes subhylanus) Flours. Foods. 2022; 11(7):976. https://doi.org/10.3390/foods11070976
Chicago/Turabian StyleVanqa, Nthabeleng, Vusi Vincent Mshayisa, and Moses Basitere. 2022. "Proximate, Physicochemical, Techno-Functional and Antioxidant Properties of Three Edible Insect (Gonimbrasia belina, Hermetia illucens and Macrotermes subhylanus) Flours" Foods 11, no. 7: 976. https://doi.org/10.3390/foods11070976
APA StyleVanqa, N., Mshayisa, V. V., & Basitere, M. (2022). Proximate, Physicochemical, Techno-Functional and Antioxidant Properties of Three Edible Insect (Gonimbrasia belina, Hermetia illucens and Macrotermes subhylanus) Flours. Foods, 11(7), 976. https://doi.org/10.3390/foods11070976