
Origin and Fate of Acrolein in Foods

 , ,
, , 
Abstract
1. Introduction
2. Origin of Acrolein in Foods
2.1. Acrolein Content in Foods
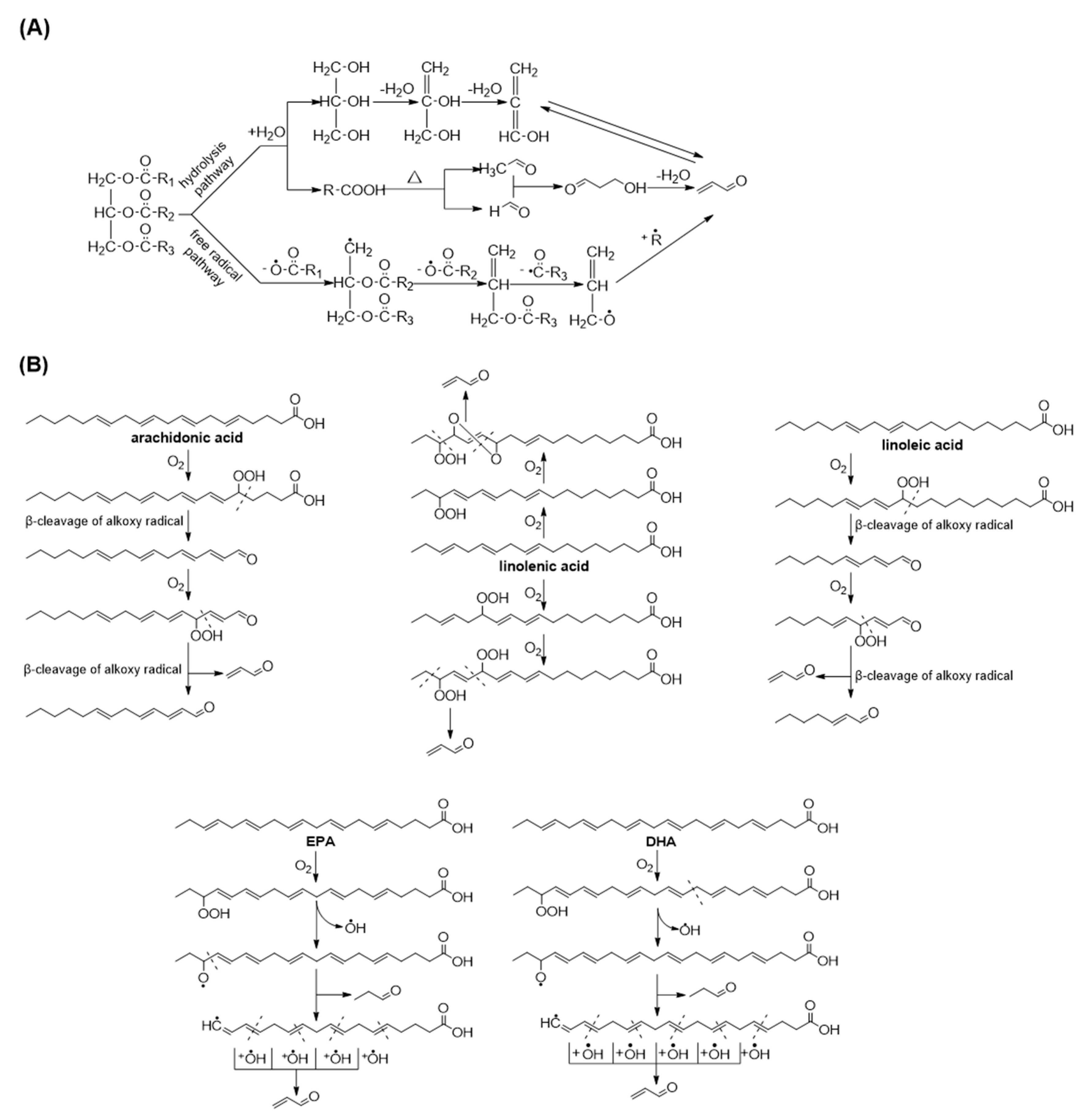
2.2. Acrolein Formation from Edible Oils
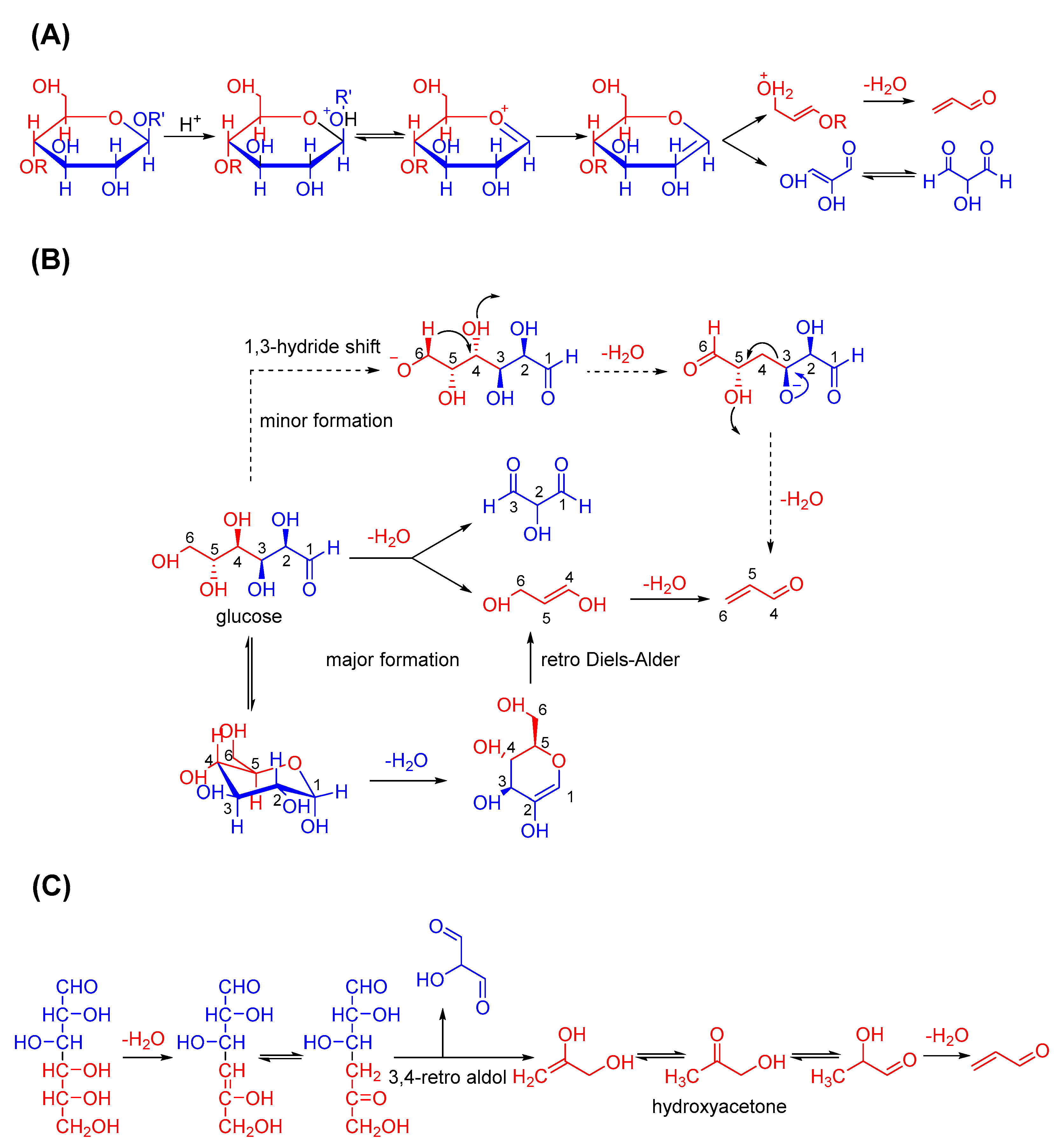
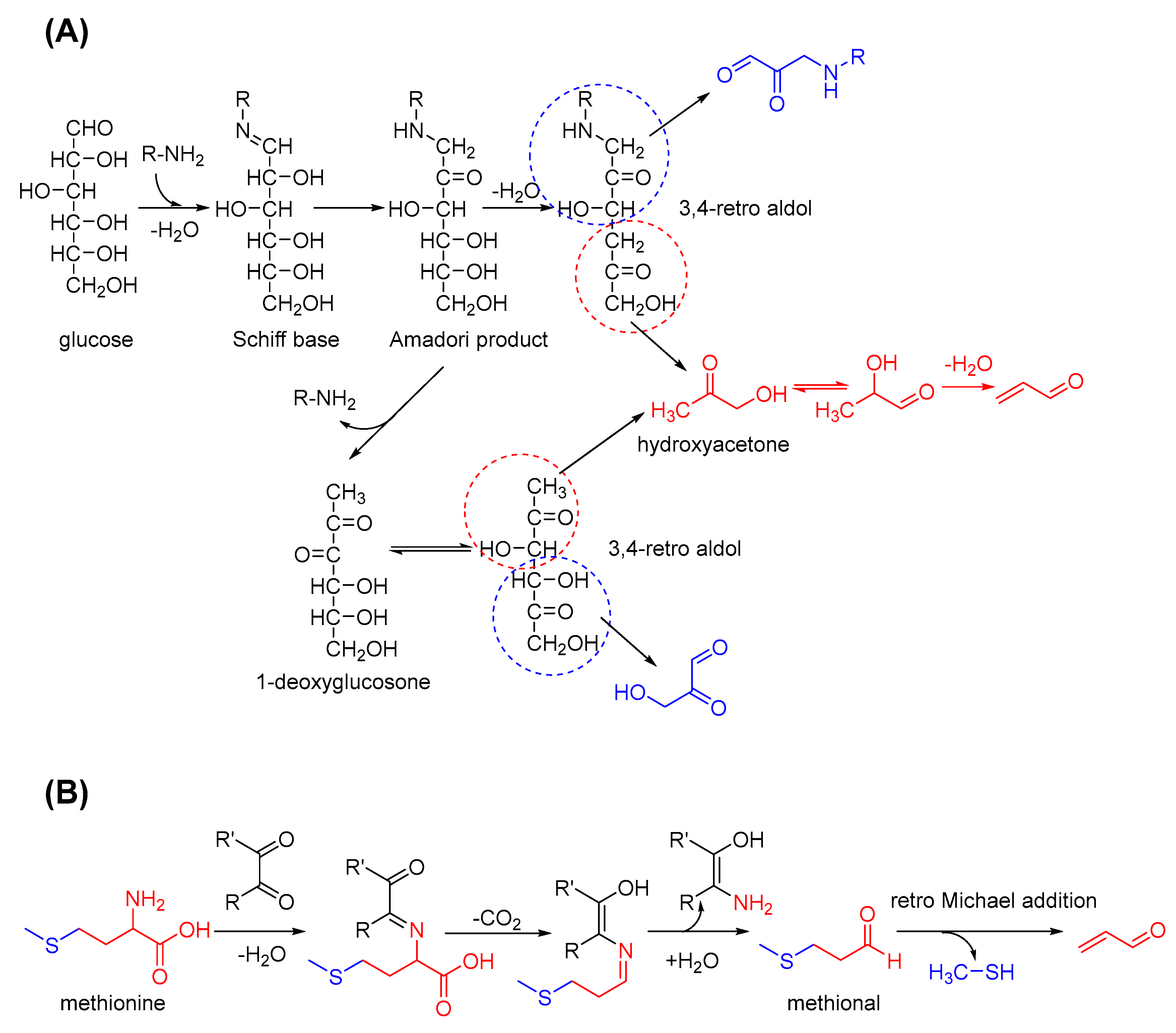
2.3. Acrolein Formation from Carbohydrates
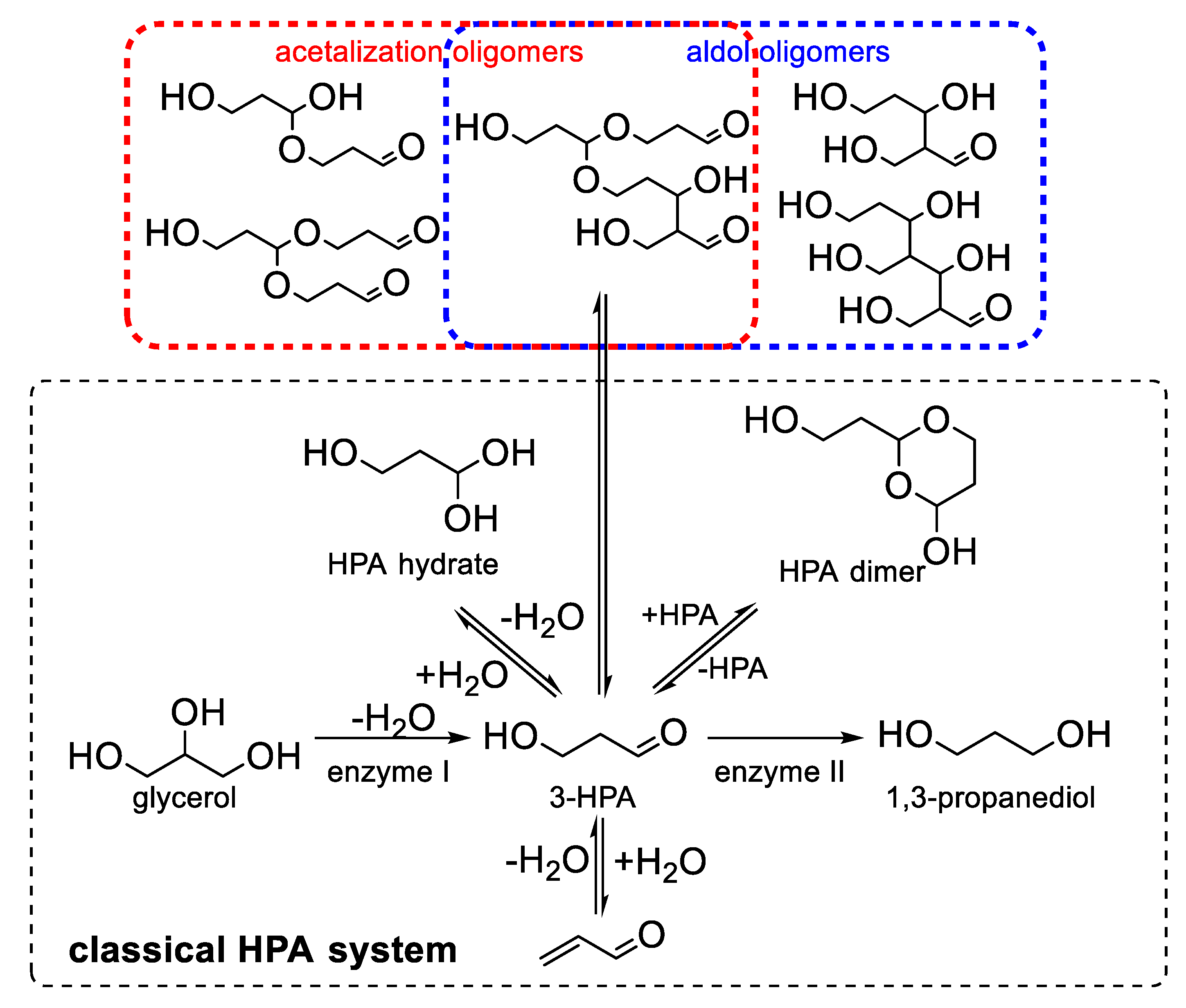
2.4. Acrolein Formation from Alcoholic Beverages
3. Fate of Acrolein in Foods
3.1. Interactions of Acrolein with Small Molecules
3.1.1. Conversion of Acrolein during Food Processing
3.1.2. Interactions of Acrolein with Amino Acids
3.1.3. Interactions of Acrolein with Antioxidant and Natural Products
3.1.4. Interactions of Acrolein with Clinical Drugs
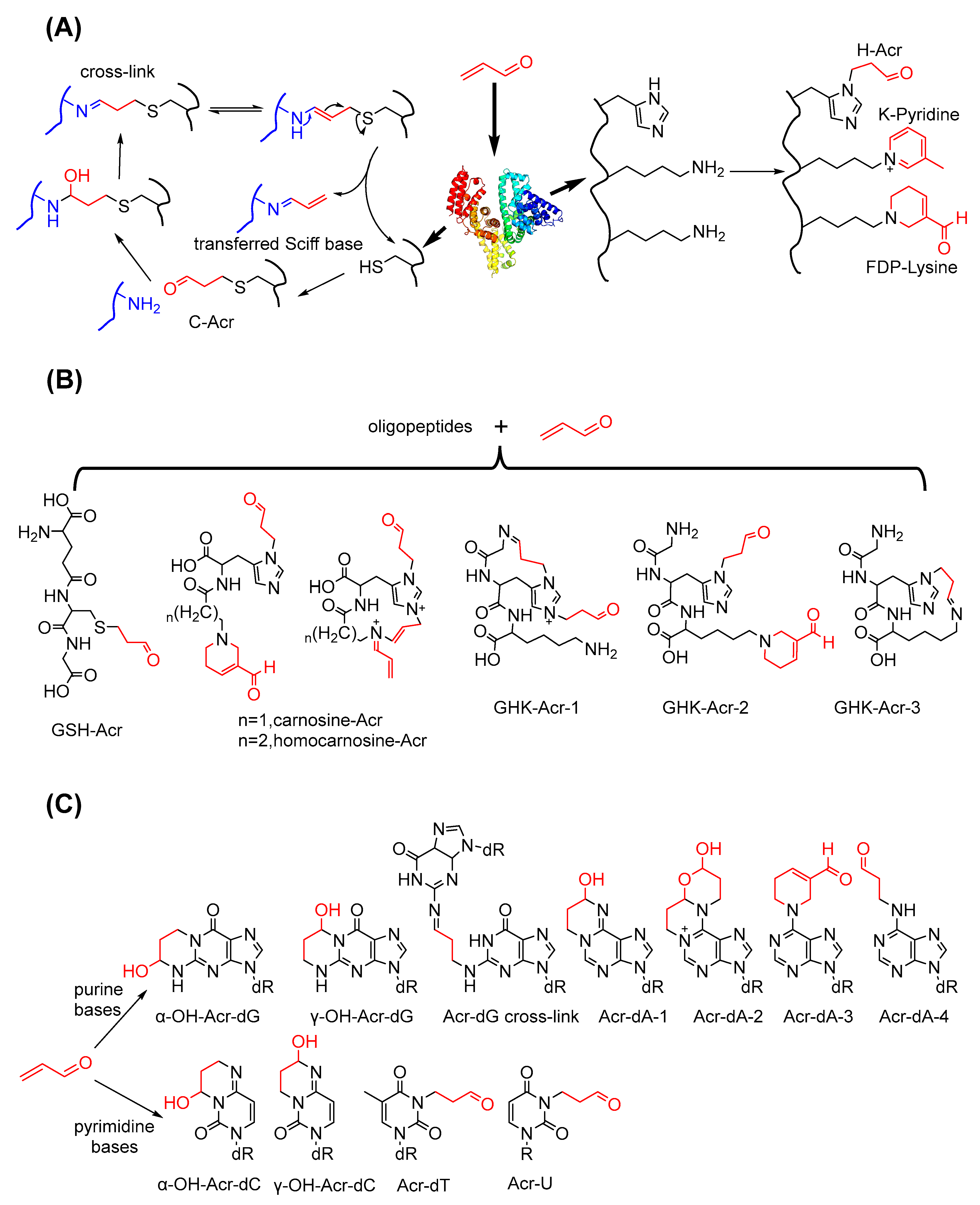
3.2. Conjugations of Acrolein to Biomacromolecules
3.2.1. Conjugations of Acrolein to Peptides and Proteins
3.2.2. Conjugations of Acrolein to Nucleic Acids
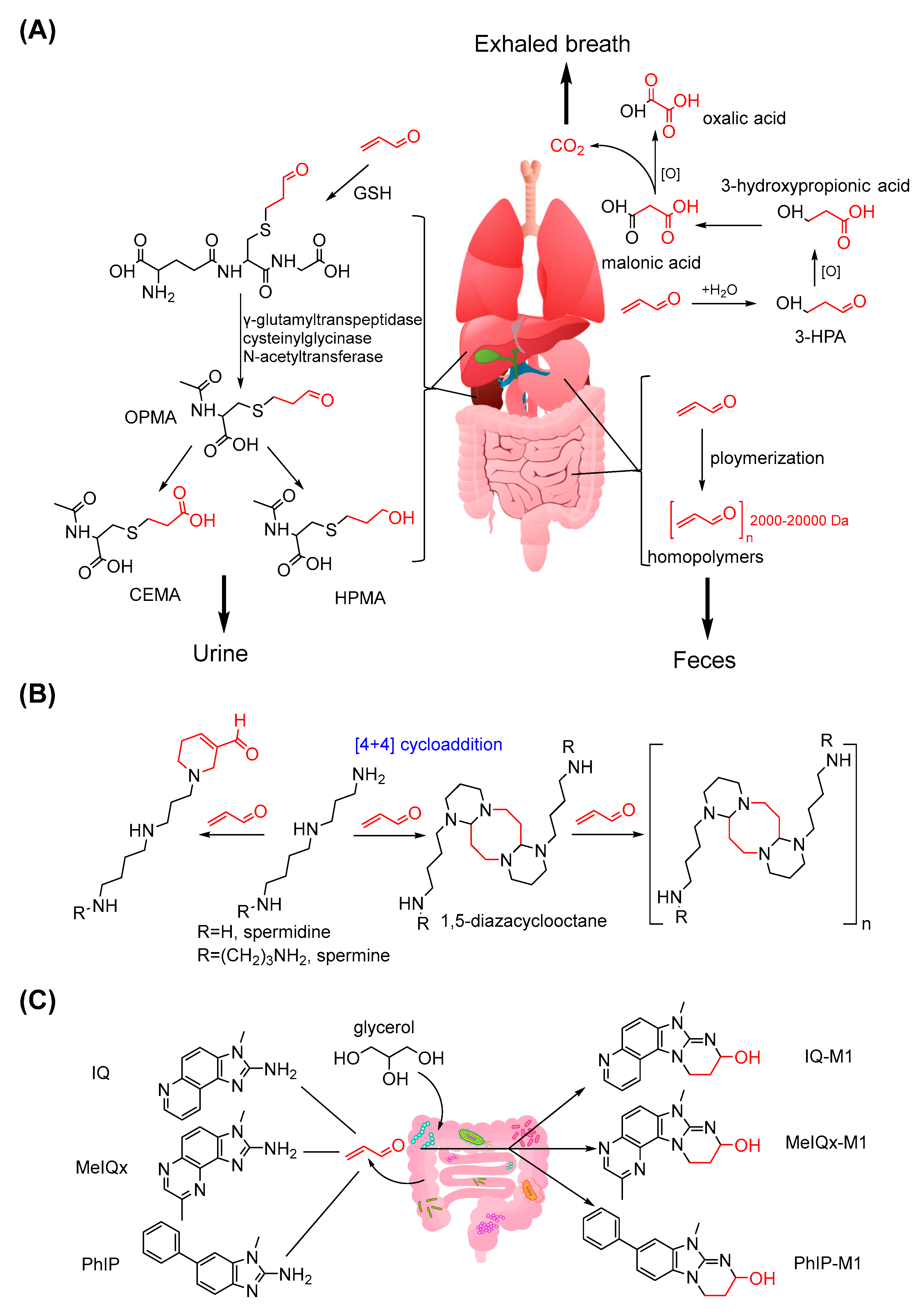
3.3. Metabolism and Biotransformation of Acrolein
4. Concluding Remarks and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chevreul, M.E. Recherches Chimiques sur les Corps Gras d’Origine Animale; F.G. Levrault: Paris, France, 1823. [Google Scholar]
- Berzelius, V. Fernere Nachrichten über das neue Metall. Ann. Phys. 1839, 123, 207–210. [Google Scholar] [CrossRef]
- Liu, L.; Ye, X.P.; Bozell, J.J. A comparative review of petroleum-based and bio-based acrolein production. ChemSusChem 2012, 5, 1162–1180. [Google Scholar] [CrossRef]
- Aizenbud, D.; Aizenbud, I.; Reznick, A.Z.; Avezov, K. Acrolein—An α, β-unsaturated aldehyde: A review of oral cavity exposure and oral pathology effects. Rambam Maimonides Med. J. 2016, 7, e0024. [Google Scholar] [CrossRef] [PubMed]
- Il’yasova, D.; Scarbrough, P.; Spasojevic, I. Urinary biomarkers of oxidative status. Clin. Chim. Acta 2012, 413, 1446–1453. [Google Scholar] [CrossRef]
- Chen, W.Y.; Wang, M.; Zhang, J.; Barve, S.S.; McClain, C.J.; Joshi-Barve, S. Acrolein disrupts tight junction proteins and causes endoplasmic reticulum stress-mediated epithelial cell death leading to intestinal barrier dysfunction and permeability. Am. J. Pathol. 2017, 187, 2686–2697. [Google Scholar] [CrossRef] [PubMed]
- Jeelani, R.; Chatzicharalampous, C.; Kohan-Ghadr, H.R.; Awonuga, A.; Joshi, N.; Morris, R.T.; Abu-Soud, H.M. Acrolein, a commonly found environmental toxin, causes oocyte mitochondrial dysfunction and negatively affects embryo development. Free Radic. Res. 2018, 52, 929–938. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Ito, S.; Nishio, N.; Tanaka, Y.; Chen, N.; Isobe, K. Acrolein induced both pulmonary inflammation and the death of lung epithelial cells. Toxicol. Lett. 2014, 229, 384–392. [Google Scholar] [CrossRef]
- Endo, R.; Uchiyama, K.; Lim, S.Y.; Itakura, M.; Adachi, T.; Uchida, K. Recognition of acrolein-specific epitopes by B cell receptors triggers an innate immune response. J. Biol. Chem. 2021, 296, 100648. [Google Scholar] [CrossRef]
- Henning, R.J.; Johnson, G.T.; Coyle, J.P.; Harbison, R.D. Acrolein can cause cardiovascular disease: A review. Cardiovasc. Toxicol. 2017, 17, 227–236. [Google Scholar] [CrossRef]
- Chen, W.Y.; Zhang, J.; Ghare, S.; Barve, S.; McClain, C.; Joshi-Barve, S. Acrolein is a pathogenic mediator of alcoholic liver disease and the scavenger hydralazine is protective in mice. Cell. Mol. Gastroenterol. Hepatol. 2016, 2, 685–700. [Google Scholar] [CrossRef]
- Huang, Y.J.; Jin, M.H.; Pi, R.B.; Zhang, J.J.; Ouyang, Y.; Chao, X.J.; Chen, M.H.; Liu, P.Q.; Yu, J.C.; Ramassamy, C.; et al. Acrolein induces Alzheimer’s disease-like pathologies in vitro and in vivo. Toxicol. Lett. 2013, 217, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Feroe, A.G.; Attanasio, R.; Scinicariello, F. Acrolein metabolites, diabetes and insulin resistance. Environ. Res. 2016, 148, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Yasuo, M.; Droma, Y.; Kitaguchi, Y.; Ito, M.; Imamura, H.; Kawakubo, M.; Hanaoka, M. The relationship between acrolein and oxidative stress in COPD: In systemic plasma and in local lung tissue. Int. J. Chronic Obstr. Pulm. Dis. 2019, 14, 1527. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.C.; Tsou, H.H.; Lin, C.C.; Chen, S.C.; Cheng, H.W.; Liu, T.S.; Chen, W.S.; Jiang, J.K.; Yang, S.H.; Chang, S.C.; et al. Acrolein contributes to human colorectal tumorigenesis through the activation of RAS-MAPK pathway. Sci. Rep. 2021, 11, 12590. [Google Scholar] [CrossRef]
- De Woskin, R.; Greenberg, M.; Pepelko, W.; Strickland, J. Toxicological Review of Acrolein (Cas No. 107-02-08) in Support of Summary Information on the Integrated Risk Information System (Iris); US Environmental Protection Agency: Washington, DC, USA, 2003.
- Abraham, K.; Andres, S.; Palavinskas, R.; Berg, K.; Appel, K.E.; Lampen, A. Toxicology and risk assessment of acrolein in food. Mol. Nutr. Food Res. 2011, 55, 1277–1290. [Google Scholar] [CrossRef]
- Ewert, A.; Granvogl, M.; Schieberle, P. Isotope-labeling studies on the formation pathway of acrolein during heat processing of oils. J. Agric. Food Chem. 2014, 62, 8524–8529. [Google Scholar] [CrossRef]
- Watzek, N.; Scherbl, D.; Feld, J.; Berger, F.; Doroshyenko, O.; Fuhr, U.; Tomalik-Scharte, D.; Baum, M.; Eisenbrand, G.; Richling, E. Profiling of mercapturic acids of acrolein and acrylamide in human urine after consumption of potato crisps. Mol. Nutr. Food Res. 2012, 56, 1825–1837. [Google Scholar] [CrossRef]
- Zhang, J.; Sturla, S.; Lacroix, C.; Schwab, C. Gut microbial glycerol metabolism as an endogenous acrolein source. MBio 2018, 9, e01947-17. [Google Scholar] [CrossRef]
- Feron, V.J.; Til, H.P.; De Vrijer, F.; Woutersen, R.A.; Cassee, F.R.; Van Bladeren, P.J. Aldehydes: Occurrence, carcinogenic potential, mechanism of action and risk assessment. Mutat. Res./Genet. Toxicol. 1991, 259, 363–385. [Google Scholar] [CrossRef]
- Drakula, S.; Novotni, D.; Mustač, N.Č.; Voučko, B.; Krpan, M.; Hruškar, M.; Ćurić, D. A simple HS-SPME/GC-MS method for determination of acrolein from sourdough to bread. Food Anal. Methods 2019, 12, 2582–2590. [Google Scholar] [CrossRef]
- Osório, V.M.; de Lourdes Cardeal, Z. Using SPME-GC/MS to evaluate acrolein production in cassava and pork sausage fried in different vegetable oils. J. Am. Oil Chem. Soc. 2013, 90, 1795–1800. [Google Scholar] [CrossRef]
- Żyżelewicz, D.; Oracz, J.; Krysiak, W.; Budryn, G.; Nebesny, E. Effects of various roasting conditions on acrylamide, acrolein, and polycyclic aromatic hydrocarbons content in cocoa bean and the derived chocolates. Dry. Technol. 2017, 35, 363–374. [Google Scholar] [CrossRef]
- Suh, J.H.; Ho, C.T.; Wang, Y. Evaluation of carbonyl species in fish oil: An improved LC–MS/MS method. Food Control 2017, 78, 463–468. [Google Scholar] [CrossRef]
- Ewert, A.; Granvogl, M.; Schieberle, P. Development of two stable isotope dilution assays for the quantitation of acrolein in heat-processed fats. J. Agric. Food Chem. 2011, 59, 3582–3589. [Google Scholar] [CrossRef] [PubMed]
- Panosyan, A.G.; Mamikonyan, G.V.; Torosyan, M.; Gabrielyan, E.S.; Mkhitaryan, S.A.; Tirakyan, M.R.; Ovanesyan, A. Determination of the composition of volatiles in Cognac (Brandy) by headspace gas chromatography–mass spectrometry. J. Anal. Chem. 2001, 56, 945–952. [Google Scholar] [CrossRef]
- Miller, B.E.; Danielson, N.D. Derivatization of vinyl aldehydes with anthrone prior to high-performance liquid chromatography with fluorometric detection. Anal. Chem. 1988, 60, 622–626. [Google Scholar] [CrossRef]
- Peterle, G.P.; Hernandes, K.C.; Schmidt, L.; Maciel, J.B.H.; Zini, C.A.; Welke, J.E. Exposure risk to carbonyl compounds and furfuryl alcohol through the consumption of sparkling wines. Ciência Rural 2019, 49, e20180986. [Google Scholar] [CrossRef]
- Ferreira, D.C.; Hernandes, K.C.; Nicolli, K.P.; Souza-Silva, É.A.; Manfroi, V.; Zini, C.A.; Welke, J.E. Development of a method for determination of target toxic carbonyl compounds in must and wine using HS-SPME-GC/MS-SIM after preliminary GC× GC/TOFMS analyses. Food Anal. Methods 2019, 12, 108–120. [Google Scholar] [CrossRef]
- de Las Heras, E.; Zuriarrain-Ocio, A.; Zuriarrain, J.; Bordagaray, A.; Dueñas, M.T.; Berregi, I. Quantitative determination of acrolein in cider by 1H NMR spectrometry. Foods 2020, 9, 1820. [Google Scholar] [CrossRef]
- Hernandes, K.C.; Souza-Silva, É.A.; Assumpção, C.F.; Zini, C.A.; Welke, J.E. Validation of an analytical method using HS-SPME-GC/MS-SIM to assess the exposure risk to carbonyl compounds and furan derivatives through beer consumption. Food Addit. Contam. Part A 2019, 36, 1808–1821. [Google Scholar] [CrossRef]
- Choe, E.; Min, D.B. Chemistry of deep-fat frying oils. J. Food Sci. 2007, 72, R77–R86. [Google Scholar] [CrossRef] [PubMed]
- Umano, K.; Shibamoto, T. Analysis of acrolein from heated cooking oils and beef fat. J. Agric. Food Chem. 1987, 35, 909–912. [Google Scholar] [CrossRef]
- Esterbauer, H.; Schaur, R.J.; Zollner, H. Chemistry and biochemistry of 4-hydroxynonenal, malonaldehyde and related aldehydes. Free Radic. Biol. Med. 1991, 11, 81–128. [Google Scholar] [CrossRef]
- Endo, Y.; Hayashi, C.; Yamanaka, T.; Takayose, K.; Yamaoka, M.; Tsuno, T.; Nakajima, S. Linolenic Acid as the Main Source of Acrolein Formed During Heating of Vegetable Oils. J. Am. Oil Chem. Soc. 2013, 90, 959–964. [Google Scholar] [CrossRef]
- Kato, S.; Shimizu, N.; Otoki, Y.; Ito, J.; Sakaino, M.; Sano, T.; Takeuchi, S.; Imagi, J.; Nakagawa, K. Determination of acrolein generation pathways from linoleic acid and linolenic acid: Increment by photo irradiation. Npj Sci. Food 2022, 6, 21. [Google Scholar] [CrossRef]
- Shibata, A.; Uemura, M.; Hosokawa, M.; Miyashita, K. Acrolein as a Major Volatile in the Early Stages of Fish Oil TAG Oxidation. J. Oleo. Sci. 2018, 67, 515–524. [Google Scholar] [CrossRef]
- Shibata, A.; Uemura, M.; Hosokawa, M.; Miyashita, K. Formation of Acrolein in the Autoxidation of Triacylglycerols with Different Fatty Acid Compositions. J. Am. Oil Chem. Soc. 2015, 92, 1661–1670. [Google Scholar] [CrossRef]
- Byrne, G.A.; Gardiner, D.; Holmes, F.H. The pyrolysis of cellulose and the action of flame-retardants. J. Appl. Chem. 1966, 16, 81–88. [Google Scholar] [CrossRef]
- Paine, J.B.; Pithawalla, Y.B.; Naworal, J.D. Carbohydrate pyrolysis mechanisms from isotopic labeling. J. Anal. Appl. Pyrolysis 2008, 82, 42–69. [Google Scholar] [CrossRef]
- Yaylayan, V.A.; Harty-Majors, S.; Ismail, A.A. Monitoring Carbonyl−Amine Reaction and Enolization of 1-Hydroxy-2-propanone (Acetol) by FTIR Spectroscopy. J. Agric. Food Chem. 1999, 47, 2335–2340. [Google Scholar] [CrossRef]
- Stevens, J.F.; Maier, C.S. Acrolein: Sources, metabolism, and biomolecular interactions relevant to human health and disease. Mol. Nutr. Food Res. 2008, 52, 7–25. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, Y.V.; Kroh, L.W. Investigation of reactive alpha-dicarbonyl compounds generated from the Maillard reactions of L-methionine with reducing sugars via their stable quinoxaline derivatives. J. Agric. Food Chem. 2010, 58, 8293–8299. [Google Scholar] [CrossRef] [PubMed]
- Vollenweider, S.; Lacroix, C. 3-hydroxypropionaldehyde: Applications and perspectives of biotechnological production. Appl. Microbiol. Biotechnol. 2004, 64, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Serjak, W.C.; Day, W.H.; Van Lanen, J.M.; Boruff, C.S. Acrolein production by bacteria found in distillery grain mashes. Appl. Microbiol. 1954, 2, 14–20. [Google Scholar] [CrossRef]
- Smiley, K.L.; Sobolov, M. A Cobamide-requiring glycerol dehydrase from an acrolein-forming Lactobacillus. Arch. Biochem. Biophys. 1962, 97, 538–543. [Google Scholar] [CrossRef]
- Mills, D.E.; Baugh, W.D.; Conner, H.A. Studies on the formation of acrolein in distillery mashes. Appl. Microbiol. 1954, 2, 9–13. [Google Scholar] [CrossRef]
- Luo, Y.; Kong, L.; Xue, R.; Wang, W.; Xia, X. Bitterness in alcoholic beverages: The profiles of perception, constituents, and contributors. Trends Food Sci. Technol. 2020, 96, 222–232. [Google Scholar] [CrossRef]
- Rentschler, H.; Tanner, H. Red wines turning bitter; contribution to the knowledge of presence of acroleine in beverages and its correlation to the turning bitter of red wines. Mitt. Geb. Lebensmittelunters. Hyg. 1951, 42, 463–475. [Google Scholar]
- Hall, R.H.; Stern, E.S. Acid-catalysed hydration of acraldehyde. Kinetics of the reaction and isolation of β-hydroxypropaldehyde. J. Chem. Soc. 1950, 490–498. [Google Scholar]
- Nielsen, A.T.; Moore, D.W.; Schuetze, J.A. 13C and 1H NMR study of formaldehyde reactions with acetaldehyde and acrolein. Synthesis of 2-(hydroxymethyl)-1, 3-propanediol. Pol. J. Chem. 1981, 55, 1393–1403. [Google Scholar]
- Sung, H.W.; Chen, C.N.; Liang, H.F.; Hong, M.H. A natural compound (reuterin) produced by Lactobacillus reuteri for biological-tissue fixation. Biomaterials 2003, 24, 1335–1347. [Google Scholar] [CrossRef]
- Sobolov, M.; Smiley, K.L. Metabolism of glycerol by an acrolein-forming lacto-bacillus. J. Bacteriol. 1960, 79, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Madrera, R.R.; Gomis, D.B.; Alonso, J.J.M. Influence of distillation system, oak wood type, and aging time on volatile compounds of cider brandy. J. Agric. Food Chem. 2003, 51, 5709–5714. [Google Scholar] [CrossRef]
- Williams, P.J.; Strauss, C.R. 3, 3-diethoxybutan-2-one and 1, 1, 3-triethoxypropane: Acetals in spirits distilled from Vitis vinifera grape wines. J. Sci. Food Agric. 1975, 26, 1127–1136. [Google Scholar] [CrossRef]
- Madrera, R.R.; Valles, B.S.; Lobo, A.P. Chemical and sensory changes in fresh cider spirits during maturation in inert containers. J. Sci. Food Agric. 2011, 91, 797–804. [Google Scholar] [CrossRef]
- Bauer, R.; Hiten, F.; Crouch, A.M.; Kossmann, J.; Burger, B.V. Acrolein dimer as a marker for direct detection of acrolein in wine. S. Afr. J. Enol. Vitic. 2012, 33, 72–79. [Google Scholar] [CrossRef][Green Version]
- Weisshaar, R. Acrylamide in heated potato products–analytics and formation routes. Eur. J. Lipid Sci. Technol. 2004, 106, 786–792. [Google Scholar] [CrossRef]
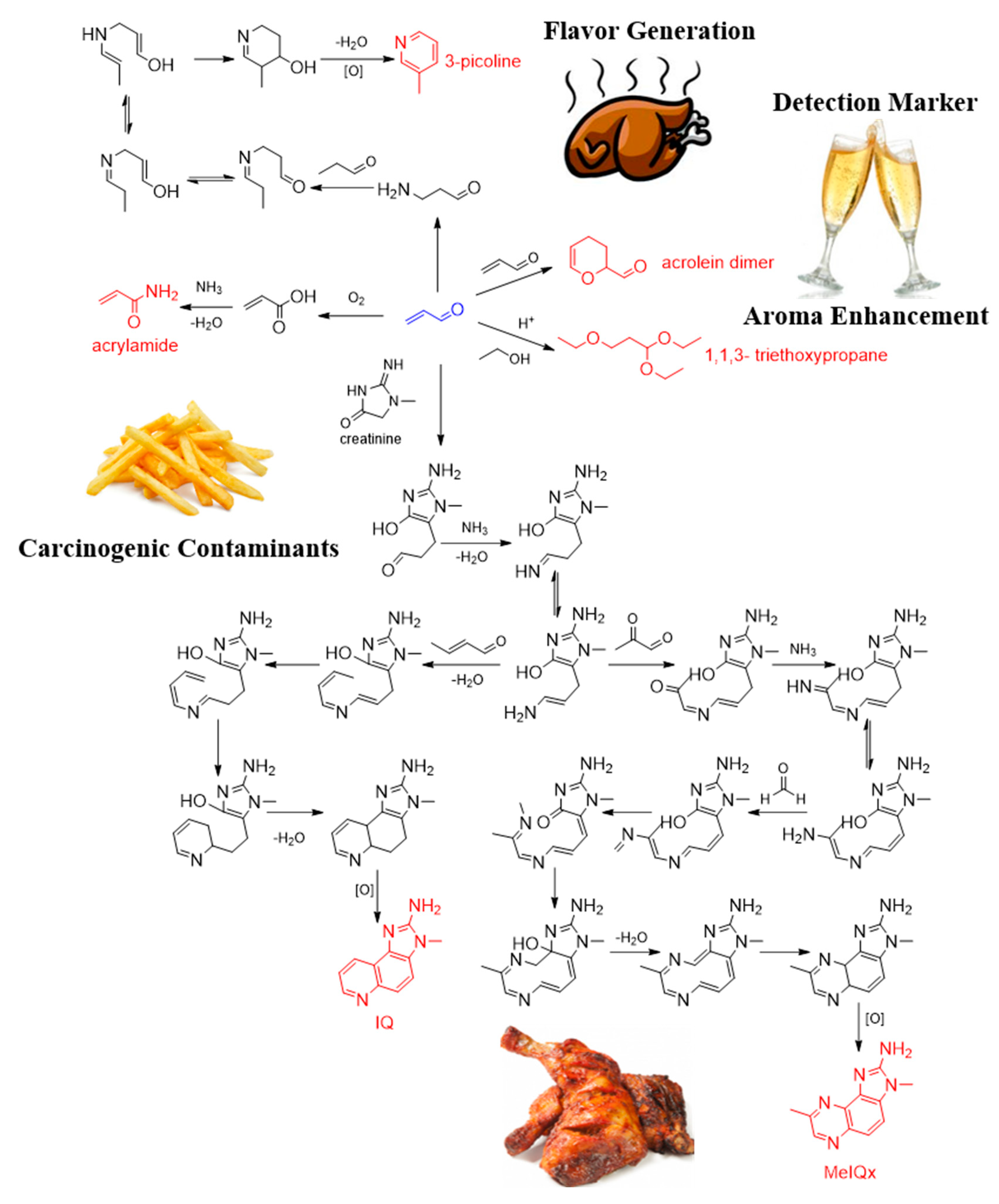
- Zamora, R.; Lavado-Tena, C.M.; Hidalgo, F.J. Identification of Precursors and Formation Pathway for the Heterocyclic Aromatic Amine 2-Amino-3-methylimidazo(4,5-f)quinoline (IQ). J. Agric. Food Chem. 2020, 68, 7474–7481. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, F.J.; Lavado-Tena, C.M.; Zamora, R. Identification of acrolein as the reactive carbonyl responsible for the formation of 2-amino-3,8-dimethylimidazo [4,5-f.quinoxaline] (MeIQx). Food Chem. 2021, 343, 128478. [Google Scholar] [CrossRef] [PubMed]
- Zamora, R.; Lavado-Tena, C.M.; Hidalgo, F.J. Oligomerization of reactive carbonyls in the presence of ammonia-producing compounds: A route for the production of pyridines in foods. Food Chem. 2020, 304, 125284. [Google Scholar]
- Jiang, K.; Huang, C.; Jiao, R.; Bai, W.; Zheng, J.; Ou, S. Adducts formed during protein digestion decreased the toxicity of five carbonyl compounds against Caco-2 cells. J. Hazard. Mater. 2019, 363, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Jiang, K.; Shi, L.; Fei, J.; Zheng, J.; Ou, S.; Ou, J. Formation of di-cysteine acrolein adduct decreases cytotoxicity of acrolein by ROS alleviation and apoptosis intervention. J. Hazard. Mater. 2020, 387, 121686. [Google Scholar] [CrossRef] [PubMed]
- Jiang, K.; Yin, Z.; Zhou, P.; Guo, H.; Huang, C.; Zhang, G.; Hu, W.; Ou, S.; Ou, J. The scavenging capacity of γ-aminobutyric acid for acrolein and the cytotoxicity of the formed adduct. Food Funct. 2020, 11, 7736–7747. [Google Scholar] [CrossRef]
- Zou, Z.; Yin, Z.; Ou, J.; Zheng, J.; Liu, F.; Huang, C.; Ou, S. Identification of adducts formed between acrolein and alanine or serine in fried potato crisps and the cytotoxicity-lowering effect of acrolein in three cell lines. Food Chem. 2021, 361, 130164. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Jiang, K.; Huang, C.; Zheng, J.; Zhou, H.; Ou, J.; Ou, S. Glycine and serine markedly eliminate methylglyoxal in the presence of formaldehyde via the formation of imidazole salts. Food Chem. 2022, 369, 130952. [Google Scholar] [CrossRef]
- Fodor, G.; Arnold, R.; Mohacsi, T.; Karle, I.; Flippen-Anderson, J. A new role for L-ascorbic acid: Michael donor to α, β-unsaturated carbonyl compounds. Tetrahedron 1983, 39, 2137–2145. [Google Scholar] [CrossRef]
- Wang, J.; Lu, Y.; Zheng, T.; Sang, S.; Lv, L. Scavenging of Acrolein by Food-Grade Antioxidant Propyl Gallate in a Model Reaction System and Cakes. J. Agric. Food Chem. 2019, 67, 8520–8526. [Google Scholar] [CrossRef]
- Zhu, Q.; Zheng, Z.P.; Cheng, K.W.; Wu, J.J.; Zhang, S.; Tang, Y.S.; Sze, K.H.; Chen, J.; Chen, F.; Wang, M.F. Natural polyphenols as direct trapping agents of lipid peroxidation-derived acrolein and 4-hydroxy-trans-2-nonenal. Chem. Res. Toxicol. 2009, 22, 1721–1727. [Google Scholar] [CrossRef]
- Zhu, Q.; Zhang, N.Q.; Lau, C.F.; Chao, J.; Sun, Z.; Chang, R.C.; Chen, F.; Wang, M. In vitro attenuation of acrolein-induced toxicity by phloretin, a phenolic compound from apple. Food Chem. 2012, 135, 1762–1768. [Google Scholar] [CrossRef]
- Wang, W.; Qi, Y.; Rocca, J.R.; Sarnoski, P.J.; Jia, A.; Gu, L. Scavenging of Toxic Acrolein by Resveratrol and Hesperetin and Identification of Adducts. J. Agric. Food Chem. 2015, 63, 9488–9495. [Google Scholar] [CrossRef]
- Sugimoto, K.; Matsuoka, Y.; Sakai, K.; Fujiya, N.; Fujii, H.; Mano, J. Catechins in green tea powder (matcha) are heat-stable scavengers of acrolein, a lipid peroxide-derived reactive carbonyl species. Food Chem. 2021, 355, 129403. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Jiang, X.; Xiao, L.; Lu, Y.; Sang, S.; Lv, L.; Dong, W. Mechanistic studies of inhibition on acrolein by myricetin. Food Chem. 2020, 323, 126788. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Zhu, Y.; Lv, L.; Sang, S. Translating In Vitro Acrolein-Trapping Capacities of Tea Polyphenol and Soy Genistein to In Vivo Situation is Mediated by the Bioavailability and Biotransformation of Individual Polyphenols. Mol. Nutr. Food Res. 2020, 64, e1900274. [Google Scholar] [CrossRef]
- Song, X.; Lu, Y.; Si, B.; Lu, Y.; Zhang, Q.; Lv, L. Inhibitory Effect on Acrolein by Cyanidin-3-O-glucoside and Its Acrolein Adducts from the Pigment of Mynica Red. J. Agric. Food Chem. 2021, 69, 11937–11946. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Lv, H.; Lu, Y.; Lu, Y.; Lv, L. Trapping of Acrolein by Curcumin and the Synergistic Inhibition Effect of Curcumin Combined with Quercetin. J. Agric. Food Chem. 2021, 69, 294–301. [Google Scholar] [CrossRef] [PubMed]
- Tao, Z.H.; Li, C.; Xu, X.F.; Pan, Y.J. Scavenging activity and mechanism study of ferulic acid against reactive carbonyl species acrolein. J. Zhejiang Univ.-Sci. B 2019, 20, 868–876. [Google Scholar] [CrossRef]
- Liu, P.; Yin, Z.; Chen, M.; Huang, C.; Wu, Z.; Huang, J.; Ou, S.; Zheng, J. Cytotoxicity of adducts formed between quercetin and methylglyoxal in PC-12 cells. Food Chem. 2021, 352, 129424. [Google Scholar] [CrossRef]
- Davies, S.S.; Zhang, L.S. Reactive Carbonyl Species Scavengers-Novel Therapeutic Approaches for Chronic Diseases. Curr. Pharmacol. Rep. 2017, 3, 51–67. [Google Scholar] [CrossRef]
- Casale, M.; Di Martino, A.; Salvinelli, F.; Trombetta, M.; Denaro, V. MESNA for chemically assisted tissue dissection. Expert Opin. Investig. Drugs 2010, 19, 699–707. [Google Scholar] [CrossRef]
- Stankiewicz, A.; Skrzydlewska, E. Amifostine-antioxidant drug in anticancer therapy. Toxicol. Mech. Methods 2006, 16, 181–188. [Google Scholar] [CrossRef]
- Kaminskas, L.M.; Pyke, S.M.; Burcham, P.C. Reactivity of hydrazinophthalazine drugs with the lipid peroxidation products acrolein and crotonaldehyde. Org. Biomol. Chem. 2004, 2, 2578–2584. [Google Scholar] [CrossRef]
- Hamann, K.; Shi, R. Acrolein scavenging: A potential novel mechanism of attenuating oxidative stress following spinal cord injury. J. Neurochem. 2009, 111, 1348–1356. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, E.M.; Song, M.S.; Dursun, S.M.; Tomlinson, S.; Todd, K.G.; Baker, G.B. Phenelzine: An Old Drug That May Hold Clues to The Development of New Neuroprotective Agents. Klin. Psikofarmakol. Bülteni-Bull. Clin. Psychopharmacol. 2016, 20, 179–186. [Google Scholar] [CrossRef]
- Aldini, G.; Vistoli, G.; Regazzoni, L.; Benfatto, M.C.; Bettinelli, I.; Carini, M. Edaravone inhibits protein carbonylation by a direct carbonyl-scavenging mechanism: Focus on reactivity, selectivity, and reaction mechanisms. Antioxid. Redox Signal. 2010, 12, 381–392. [Google Scholar] [CrossRef]
- Jiang, X.; Zhang, D.; Lu, Y.; Lv, L. Acrolein-Trapping Mechanism of Theophylline in Green Tea, Coffee, and Cocoa: Speedy and Successful. J. Agric. Food Chem. 2020, 68, 9718–9724. [Google Scholar] [CrossRef]
- Jiang, X.; Lu, Y.; Lv, L. Trapping Acrolein by Theophylline/Caffeine and Their Metabolites from Green Tea and Coffee in Mice and Humans. J. Agric. Food Chem. 2020, 68, 14471–14479. [Google Scholar] [CrossRef] [PubMed]
- Burcham, P.C. Potentialities and pitfalls accompanying chemico-pharmacological strategies against endogenous electrophiles and carbonyl stress. Chem. Res. Toxicol. 2008, 21, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K.; Kanematsu, M.; Morimitsu, Y.; Osawa, T.; Noguchi, N.; Niki, E. Acrolein is a product of lipid peroxidation reaction. Formation of free acrolein and its conjugate with lysine residues in oxidized low density lipoproteins. J. Biol. Chem. 1998, 273, 16058–16066. [Google Scholar] [CrossRef]
- Calingasan, N.Y.; Uchida, K.; Gibson, G.E. Protein-bound acrolein: A novel marker of oxidative stress in Alzheimer’s disease. J. Neurochem. 1999, 72, 751–756. [Google Scholar] [CrossRef]
- Furuhata, A.; Ishii, T.; Kumazawa, S.; Yamada, T.; Nakayama, T.; Uchida, K. N(epsilon)-(3-methylpyridinium)lysine, a major antigenic adduct generated in acrolein-modified protein. J. Biol. Chem. 2003, 278, 48658–48665. [Google Scholar] [CrossRef]
- Maeshima, T.; Honda, K.; Chikazawa, M.; Shibata, T.; Kawai, Y.; Akagawa, M.; Uchida, K. Quantitative analysis of acrolein-specific adducts generated during lipid peroxidation-modification of proteins in vitro: Identification of N(tau)-(3-propanal)histidine as the major adduct. Chem. Res. Toxicol. 2012, 25, 1384–1392. [Google Scholar] [CrossRef] [PubMed]
- Seiner, D.R.; LaButti, J.N.; Gates, K.S. Kinetics and mechanism of protein tyrosine phosphatase 1B inactivation by acrolein. Chem. Res. Toxicol. 2007, 20, 1315–1320. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Bhatnagar, A.; Pierce, W.M., Jr. Protein modification by acrolein: Formation and stability of cysteine adducts. Chem. Res. Toxicol. 2009, 22, 708–716. [Google Scholar] [CrossRef]
- Carini, M.; Aldini, G.; Beretta, G.; Arlandini, E.; Facino, R.M. Acrolein-sequestering ability of endogenous dipeptides: Characterization of carnosine and homocarnosine/acrolein adducts by electrospray ionization tandem mass spectrometry. J. Mass Spectrom. 2003, 38, 996–1006. [Google Scholar] [CrossRef] [PubMed]
- Beretta, G.; Arlandini, E.; Artali, R.; Anton, J.M.; Maffei Facino, R. Acrolein sequestering ability of the endogenous tripeptide glycyl-histidyl-lysine (GHK), characterization of conjugation products by ESI-MSn and theoretical calculations. J. Pharm. Biomed. Anal. 2008, 47, 596–602. [Google Scholar] [CrossRef]
- Yoshida, M.; Higashi, K.; Jin, L.; Machi, Y.; Suzuki, T.; Masuda, A.; Dohmae, N.; Suganami, A.; Tamura, Y.; Nishimura, K.; et al. Identification of acrolein-conjugated protein in plasma of patients with brain infarction. Biochem. Biophys. Res. Commun. 2010, 391, 1234–1239. [Google Scholar] [CrossRef]
- Hristova, M.; Spiess, P.C.; Kasahara, D.I.; Randall, M.J.; Deng, B.; van der Vliet, A. The tobacco smoke component, acrolein, suppresses innate macrophage responses by direct alkylation of c-Jun N-terminal kinase. Am. J. Respir. Cell Mol. Biol. 2012, 46, 23–33. [Google Scholar] [CrossRef]
- Tran, T.N.; Kosaraju, M.G.; Tamamizu-Kato, S.; Akintunde, O.; Zheng, Y.; Bielicki, J.K.; Pinkerton, K.; Uchida, K.; Lee, Y.Y.; Narayanaswami, V. Acrolein modification impairs key functional features of rat apolipoprotein E: Identification of modified sites by mass spectrometry. Biochemistry 2014, 53, 361–375. [Google Scholar] [CrossRef]
- Nakamura, M.; Tomitori, H.; Suzuki, T.; Sakamoto, A.; Terui, Y.; Saiki, R.; Dohmae, N.; Igarashi, K.; Kashiwagi, K. Inactivation of GAPDH as one mechanism of acrolein toxicity. Biochem. Biophys. Res. Commun. 2013, 430, 1265–1271. [Google Scholar] [CrossRef]
- Witort, E.; Capaccioli, S.; Becatti, M.; Fiorillo, C.; Batignani, G.; Pavoni, V.; Piccini, M.; Orioli, M.; Carini, M.; Aldini, G.; et al. Albumin Cys34 adducted by acrolein as a marker of oxidative stress in ischemia-reperfusion injury during hepatectomy. Free Radic. Res. 2016, 50, 831–839. [Google Scholar] [CrossRef]
- Raju, S.V.; Lin, V.Y.; Liu, L.; McNicholas, C.M.; Karki, S.; Sloane, P.A.; Tang, L.; Jackson, P.L.; Wang, W.; Wilson, L.; et al. The Cystic Fibrosis Transmembrane Conductance Regulator Potentiator Ivacaftor Augments Mucociliary Clearance Abrogating Cystic Fibrosis Transmembrane Conductance Regulator Inhibition by Cigarette Smoke. Am. J. Respir. Cell Mol. Biol. 2017, 56, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Randall, M.J.; Haenen, G.R.; Bouwman, F.G.; van der Vliet, A.; Bast, A. The tobacco smoke component acrolein induces glucocorticoid resistant gene expression via inhibition of histone deacetylase. Toxicol. Lett. 2016, 240, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Fang, L.; Chen, D.; Yu, C.; Li, H.; Brocato, J.; Huang, L.; Jin, C. Mechanisms Underlying Acrolein-Mediated Inhibition of Chromatin Assembly. Mol. Cell. Biol. 2016, 36, 2995–3008. [Google Scholar] [CrossRef]
- Takamiya, R.; Uchida, K.; Shibata, T.; Maeno, T.; Kato, M.; Yamaguchi, Y.; Ariki, S.; Hasegawa, Y.; Saito, A.; Miwa, S.; et al. Disruption of the structural and functional features of surfactant protein A by acrolein in cigarette smoke. Sci. Rep. 2017, 7, 8304. [Google Scholar] [CrossRef]
- Uemura, T.; Suzuki, T.; Saiki, R.; Dohmae, N.; Ito, S.; Takahashi, H.; Toida, T.; Kashiwagi, K.; Igarashi, K. Activation of MMP-9 activity by acrolein in saliva from patients with primary Sjogren’s syndrome and its mechanism. Int. J. Biochem. Cell Biol. 2017, 88, 84–91. [Google Scholar] [CrossRef]
- Afonso, C.B.; Sousa, B.C.; Pitt, A.R.; Spickett, C.M. A mass spectrometry approach for the identification and localization of small aldehyde modifications of proteins. Arch. Biochem. Biophys. 2018, 646, 38–45. [Google Scholar] [CrossRef]
- Moore, P.J.; Reidel, B.; Ghosh, A.; Sesma, J.; Kesimer, M.; Tarran, R. Cigarette smoke modifies and inactivates SPLUNC1, leading to airway dehydration. FASEB J. 2018, 32, 6559–6574. [Google Scholar] [CrossRef]
- Acosta, G.; Race, N.; Herr, S.; Fernandez, J.; Tang, J.; Rogers, E.; Shi, R. Acrolein-mediated alpha-synuclein pathology involvement in the early post-injury pathogenesis of mild blast-induced Parkinsonian neurodegeneration. Mol. Cell. Neurosci. 2019, 98, 140–154. [Google Scholar] [CrossRef]
- Uemura, T.; Suzuki, T.; Ko, K.; Watanabe, K.; Dohmae, N.; Sakamoto, A.; Terui, Y.; Toida, T.; Kashiwagi, K.; Igarashi, K. Inhibition of dendritic spine extension through acrolein conjugation with alpha-, beta-tubulin proteins. Int. J. Biochem. Cell Biol. 2019, 113, 58–66. [Google Scholar] [CrossRef]
- Cruz, S.; Narayanaswami, V. Cellular Uptake and Clearance of Oxidatively-modified Apolipoprotein E3 by Cerebral Cortex Endothelial Cells. Int. J. Mol. Sci. 2019, 20, 4582. [Google Scholar] [CrossRef] [PubMed]
- Sousa, B.C.; Ahmed, T.; Dann, W.L.; Ashman, J.; Guy, A.; Durand, T.; Pitt, A.R.; Spickett, C.M. Short-chain lipid peroxidation products form covalent adducts with pyruvate kinase and inhibit its activity in vitro and in breast cancer cells. Free Radic. Biol. Med. 2019, 144, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Takamiya, R.; Takahashi, M.; Maeno, T.; Saito, A.; Kato, M.; Shibata, T.; Uchida, K.; Ariki, S.; Nakano, M. Acrolein in cigarette smoke attenuates the innate immune responses mediated by surfactant protein D. Biochim. Biophys. Acta (BBA) Gen. Subj. 2020, 1864, 129699. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, K.; Uemura, T.; Kashiwagi, K. Assessing acrolein for determination of the severity of brain stroke, dementia, renal failure, and Sjogren’s syndrome. Amino Acids 2020, 52, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Bein, K.; Birru, R.L.; Wells, H.; Larkin, T.P.; Cantrell, P.S.; Fagerburg, M.V.; Zeng, X.; Leikauf, G.D. Albumin Protects Lung Cells against Acrolein Cytotoxicity and Acrolein-Adducted Albumin Increases Heme Oxygenase 1 Transcripts. Chem. Res. Toxicol. 2020, 33, 1969–1979. [Google Scholar] [CrossRef] [PubMed]
- Uemura, T.; Suzuki, T.; Ko, K.; Nakamura, M.; Dohmae, N.; Sakamoto, A.; Terui, Y.; Toida, T.; Kashiwagi, K.; Igarashi, K. Structural change and degradation of cytoskeleton due to the acrolein conjugation with vimentin and actin during brain infarction. Cytoskeleton 2020, 77, 414–421. [Google Scholar] [CrossRef]
- Kobayashi, M.; Watanabe, K.; Suzuki, T.; Dohmae, N.; Fujiyoshi, M.; Uchida, M.; Suzuki, T.; Igarashi, K.; Ishii, I. Analysis of the acrolein-modified sites of apolipoprotein B-100 in LDL. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2021, 1866, 158809. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, Y.; Lu, J.; Chen, Z.; Wang, C.; Pi, R. Acrolein-conjugated proteomics in brains of adult C57BL/6 mice chronically exposed to acrolein and aged APP/PS1 transgenic AD mice. Toxicol. Lett. 2021, 344, 11–17. [Google Scholar] [CrossRef]
- Zhu, R.; Zhang, G.; Jing, M.; Han, Y.; Li, J.; Zhao, J.; Li, Y.; Chen, P.R. Genetically encoded formaldehyde sensors inspired by a protein intra-helical crosslinking reaction. Nat. Commun. 2021, 12, 581. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, Y.; Hou, X.; Ye, Z.; Wang, C. Quantitative and Site-Specific Chemoproteomic Profiling of Targets of Acrolein. Chem. Res. Toxicol. 2019, 32, 467–473. [Google Scholar] [CrossRef]
- Chung, F.L.; Young, R.; Hecht, S.S. Formation of cyclic 1, N2-propanodeoxyguanosine adducts in DNA upon reaction with acrolein or crotonaldehyde. Cancer Res. 1984, 44, 990–995. [Google Scholar]
- Smith, R.A.; Williamson, D.S.; Cerny, R.L.; Cohen, S.M. Detection of 1, N6-propanodeoxyadenosine in acrolein-modified polydeoxyadenylic acid and DNA by 32P postlabeling. Cancer Res. 1990, 50, 3005–3012. [Google Scholar] [PubMed]
- Smith, R.A.; Williamson, D.S.; Cohen, S.M. Identification of 3, N4-propanodeoxycytidine 5’-monophosphate formed by the reaction of acrolein with deoxycytidine 5’-monophosphate. Chem. Res. Toxicol. 1989, 2, 267–271. [Google Scholar] [CrossRef] [PubMed]
- Chenna, A.; Rieger, R.A.; Iden, C.R. Characterization of thymidine adducts formed by acrolein and 2-bromoacrolein. Carcinogenesis 1992, 13, 2361–2365. [Google Scholar] [CrossRef] [PubMed]
- Chenna, A.; Iden, C.R. Characterization of 2’-deoxycytidine and 2’-deoxyuridine adducts formed in reactions with acrolein and 2-bromoacrolein. Chem. Res Toxicol. 1993, 6, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Pawłowicz, A.J.; Munter, T.; Zhao, Y.; Kronberg, L. Formation of acrolein adducts with 2′-deoxyadenosine in calf thymus DNA. Chem. Toxicol. 2006, 19, 571–576. [Google Scholar] [CrossRef] [PubMed]
- Kozekov, I.D.; Nechev, L.V.; Moseley, M.S.; Harris, C.M.; Rizzo, C.J.; Stone, M.P.; Harris, T.M. DNA interchain cross-links formed by acrolein and crotonaldehyde. J. Am. Chem. Soc. 2003, 125, 50–61. [Google Scholar] [CrossRef]
- Chung, F.L.; Wu, M.Y.; Basudan, A.; Dyba, M.; Nath, R.G. Regioselective formation of acrolein-derived cyclic 1, N (2)-propanodeoxyguanosine adducts mediated by amino acids, proteins, and cell lysates. Chem. Res. Toxicol. 2012, 25, 1921–1928. [Google Scholar] [CrossRef]
- Tang, M.S.; Wang, H.T.; Hu, Y.; Chen, W.S.; Akao, M.; Feng, Z.; Hu, W. Acrolein induced DNA damage, mutagenicity and effect on DNA repair. Mol. Nutr. Food Res. 2011, 55, 1291–1300. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lovell, M.A.; Lynn, B.C. Development of a method for quantification of acrolein−deoxyguanosine adducts in DNA using isotope dilution-capillary LC/MS/MS and its application to human brain tissue. Anal. Chem. 2005, 77, 5982–5989. [Google Scholar] [CrossRef]
- Daimon, M.; Sugiyama, K.; Kameda, W.; Saitoh, T.; Oizumi, T.; Harata, A.; Yamaguchi, H.; Ohnuma, H.; Igarashi, M.; Kato, T. Increased urinary levels of pentosidine, pyrraline and acrolein adduct in type 2 diabetes. Endocr. J. 2003, 50, 61–67. [Google Scholar] [CrossRef]
- Hong, J.H.; Lee, P.A.H.; Lu, Y.C.; Huang, C.Y.; Chen, C.H.; Chiang, C.H.; Chow, P.M.; Jaw, F.S.; Wang, C.C.; Huang, C.Y.; et al. Acrolein contributes to urothelial carcinomas in patients with chronic kidney disease. Urol. Oncol. Semin. Orig. Investig. 2020, 38, 465–475. [Google Scholar] [CrossRef] [PubMed]
- Feng, Z.; Hu, W.; Hu, Y.; Tang, M.S. Acrolein is a major cigarette related lung cancer agent: Preferential binding at p53 mutational hotspots and inhibition of DNA repair. Proc. Natl. Acad. Sci. USA 2006, 103, 15404–15409. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Wang, H.T.; Weng, M.; Hu, Y.; Chen, W.; Chou, D.; Liu, Y.; Donin, N.; Huang, W.C.; Lepor, H.; et al. Acrolein-and 4-Aminobiphenyl-DNA adducts in human bladder mucosa and tumor tissue and their mutagenicity in human urothelial cells. Oncotarget 2014, 5, 3526–3540. [Google Scholar] [CrossRef] [PubMed]
- Tsou, H.H.; Hu, C.H.; Liu, J.H.; Liu, C.J.; Lee, C.H.; Liu, T.Y.; Wang, H.T. Acrolein Is Involved in the Synergistic Potential of Cigarette Smoking- and Betel Quid Chewing-Related Human Oral Cancer. Cancer Epidemiol. Biomark. Prev. 2019, 28, 954–962. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Balbo, S.; Villalta, P.W.; Hecht, S.S. Analysis of Acrolein-Derived 1, N(2)-Propanodeoxyguanosine Adducts in Human Lung DNA from Smokers and Nonsmokers. Chem. Res. Toxicol. 2019, 32, 318–325. [Google Scholar] [CrossRef]
- Zhang, S.; Balbo, S.; Wang, M.; Hecht, S.S. Analysis of acrolein-derived 1, N 2-propanodeoxyguanosine adducts in human leukocyte DNA from smokers and nonsmokers. Chem. Res. Toxicol. 2011, 24, 119–124. [Google Scholar] [CrossRef]
- Liu, X.Y.; Zhu, M.X.; Xie, J.P. Mutagenicity of acrolein and acrolein-induced DNA adducts. Toxicol. Mech. Methods 2010, 20, 36–44. [Google Scholar] [CrossRef]
- Wei, X.; Yin, H. Covalent modification of DNA by alpha, beta-unsaturated aldehydes derived from lipid peroxidation: Recent progress and challenges. Free Radic. Res. 2015, 49, 905–917. [Google Scholar] [CrossRef]
- Minetti, C.A.; Remeta, D.P.; Johnson, F.; Iden, C.R.; Breslauer, K.J. Impact of alpha-hydroxy-propanodeoxyguanine adducts on DNA duplex energetics: Opposite base modulation and implications for mutagenicity and genotoxicity. Biopolymers 2010, 93, 370–382. [Google Scholar]
- Pan, J.; Sinclair, E.; Xuan, Z.; Dyba, M.; Fu, Y.; Sen, S.; Berry, D.; Creswell, K.; Hu, J.; Roy, R.; et al. Nucleotide excision repair deficiency increases levels of acrolein-derived cyclic DNA adduct and sensitizes cells to apoptosis induced by docosahexaenoic acid and acrolein. Mutat. Res./Fundam. Mol. Mech. Mutagenesis 2016, 789, 33–38. [Google Scholar] [CrossRef]
- Parent, R.A.; Caravello, H.E.; Sharp, D.E. Metabolism and distribution of [2, 3-14C]. acrolein in Sprague-Dawley rats. J. Appl. Toxicol. 1996, 16, 449–457. [Google Scholar] [CrossRef]
- Corradi, M.; Pignatti, P.; Manini, P.; Andreoli, R.; Goldoni, M.; Poppa, M.; Moscato, G.; Balbi, B.; Mutti, A. Comparison between exhaled and sputum oxidative stress biomarkers in chronic airway inflammation. Eur. Respir. J. 2004, 24, 1011–1017. [Google Scholar] [CrossRef] [PubMed]
- Parent, R.A.; Paust, D.E.; Schrimpf, M.K.; Talaat, R.E.; Doane, R.A.; Caravello, H.E.; Lee, S.J.; Sharp, D.E. Metabolism and distribution of [2, 3-14C]. acrolein in Sprague-Dawley rats: II. Identification of urinary and fecal metabolites. Toxicol. Sci. 1998, 43, 110–120. [Google Scholar] [CrossRef]
- Yoshida, M.; Tomitori, H.; Machi, Y.; Hagihara, M.; Higashi, K.; Goda, H.; Ohya, T.; Niitsu, M.; Kashiwagi, K.; Igarashi, K. Acrolein toxicity: Comparison with reactive oxygen species. Biochem. Biophys. Res. Commun. 2009, 378, 313–318. [Google Scholar] [CrossRef] [PubMed]
- Tsutsui, A.; Zako, T.; Bu, T.; Yamaguchi, Y.; Maeda, M.; Tanaka, K. 1,5-Diazacyclooctanes, as Exclusive Oxidative Polyamine Metabolites, Inhibit Amyloid-beta(1-40) Fibrillization. Adv. Sci. 2016, 3, 1600082. [Google Scholar] [CrossRef] [PubMed]
- Tsutsui, A.; Imamaki, R.; Kitazume, S.; Hanashima, S.; Yamaguchi, Y.; Kaneda, M.; Oishi, S.; Fujii, N.; Kurbangalieva, A.; Taniguchi, N.; et al. Polyamine modification by acrolein exclusively produces 1,5-diazacyclooctanes: A previously unrecognized mechanism for acrolein-mediated oxidative stress. Org. Biomol. Chem. 2014, 12, 5151–5157. [Google Scholar] [CrossRef]
- Zhang, J.; Empl, M.T.; Schwab, C.; Fekry, M.I.; Engels, C.; Schneider, M.; Lacroix, C.; Steinberg, P.; Sturla, S.J. Gut Microbial Transformation of the Dietary Imidazoquinoxaline Mutagen MelQx Reduces Its Cytotoxic and Mutagenic Potency. Toxicol. Sci. 2017, 159, 266–276. [Google Scholar] [CrossRef]
- Zhang, J.; Lacroix, C.; Wortmann, E.; Ruscheweyh, H.J.; Sunagawa, S.; Sturla, S.J.; Schwab, C. Gut microbial beta-glucuronidase and glycerol/diol dehydratase activity contribute to dietary heterocyclic amine biotransformation. BMC Microbiol. 2019, 19, 99. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Food | Content (μg/kg or L) | Food | Content (μg/kg or L) |
|---|---|---|---|
| Fruits [21] | 10–50 | Roasted Cocoa Beans [24] | 0.25–0.45 |
| Vegetables [21] | 590 | Fish Oil [25] | 200–1600 |
| Cheese [10] | 1000 | Frying Oils [26] | 7400–198,100 |
| Doughnuts [18] | 14.1–16.9 | Frying Fats [26] | 56,500 |
| Codfish Fillet [10] | 100 | Cognacs [27] | 1420–1500 |
| Sour Dough [22] | 14.72 | Scotch Whiskey [28] | 670–11,100 |
| Bread [22] | 161 | Sparkling Wine [29] | 20.3–33.4 |
| French Fries [18] | 14.8–19.9 | Red Wine [30] | 1.0–1.5 |
| Potato Chips [18] | 16.3–23.3 | Cider [31] | 2600–31,800 |
| Frying Cassava [23] | 1.7–10.2 | Beer [32] | <2.5–5.4 |
| Frying Pork Sausage [23] | ≈2–6 |
| Sample | Target Protein | Modified Site | Interfered Function | Related Disease | Ref. |
|---|---|---|---|---|---|
| Plasma, patients | albumin | K557, K560 | - | Silent brain infarction | [98] |
| In vitro reaction | recombinant JNK2α2 | C41, C177 | Weaken interations with MKK4/7 | - | [99] |
| In vitro reaction | rat apolipoprotein E | M60, K64, K68, K135, K138, K149, K155, K254 | Impair plasma cholesterol homeostasis | Dysregulation in lipid metabolism | [100] |
| Neuro2a cells | GAPDH | C150, C282 | Inactivation of GAPDH | - | [101] |
| Plasma, patients | albumin | C34 | Oxidative stress | Ischemia-reperfusion injury | [102] |
| HBE cells, human lung explants | cystic fibrosis transmembrane conductance regulator | C524, C647, C1395, K464, K1334, K1177, K532 | Instability at the cell suface | Chronic obstructive pulmonary disease | [103] |
| U-937 monocytes | hinder histone deacetylases 2 | C274 | - | Chronic obstructive pulmonary disease | [104] |
| BEAS-2B cells | histone H4 | K5, K8, K12, K16 | Chromatin assembly | - | [105] |
| Lung tissues, C57BL/6 mice | surfactant protein A | H39, H116, C155, K180, K221, C224 | Immunity dysfunction | Chronic obstructive pulmonary disease | [106] |
| Saliva, patients | MMP-9 | C99, C230, C244, C302, C314, C329, C347, C361, C388, K384, H405, H411, C516, K535 | Activation of MMP-9 | Primary Sjögren’s syndrome | [107] |
| In vitro reaction | lysozyme | C6, C30, C64, C76, C80, K96, C155, K116 | - | - | [108] |
| In vitro reaction | human serum albumin | C34, H67, K137, H146, K262, K276, H288, H338, K414, K525, K574 | - | - | [108] |
| In vitro reaction | short palate lung and nasal epithelial clone 1 | C180, C224 | Disruption of the disulfide bond | Chronic obstructive pulmonary disease | [109] |
| Brain tissue, SD rat | α-synuclein | - | Oligomerisation and aggregation | Parkinson’s disease | [110] |
| In vitro reaction | α- and β-tubulins | C25, C295, C347, C376 in α-tubulin; C12, C129, C211, C239, C303, C354 in β-tubulin | Microtubule formation inhibition | Brain infarction | [111] |
| bEnd.3 cells | apolipoprotein E3 | K1, K69, K72, K75, K95, K157, K233, K242, K282 | Impair plasma cholesterol homeostasis | Lipid disorders | [112] |
| MCF-7 cells | pyruvate kinase | C49, C152, K166, K207, C358, H391, K393, C423, C474, K475 | Activity inhibition | Cancer | [113] |
| Lung tissues, C57BL/6 mice | surfactant protein D | K243, K246, K287, K299, K303 | Immunity dysfunction | Smoking-associated respiratory diseases | [114] |
| Saliva, patients | immunoglobulins | K43(λ); K75, K80, H81, K82, C86(κ); C300(α-2); C27, K30(γ-1); C297, K300 (γ-3) | Autoantibody functionalisation | Primary Sjögren’s syndrome | [115] |
| Epithelial lining fluid, C57BL/6J mice | albumin | C34 | HMOX1 transcript increase | - | [116] |
| FM3A cells | vimentin | C328 | Dysfunction of the cytoskeleton | Brain infarction | [117] |
| FM3A cells | actin | C217, C257, C285, K118 | Dysfunction of the cytoskeleton | Brain infarction | [117] |
| Low density lipoprotein | apolipoprotein B-100 | C212, K327, K742, K949, K1087, H1923, K2634, K3237, K3846 | Uptake of LDL | - | [118] |
| Hippocampus tissues, C57BL/6 mice | 14-3-3 protein | - | Aggregation of tau | Alzheimer’s disease | [119] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, K.; Huang, C.; Liu, F.; Zheng, J.; Ou, J.; Zhao, D.; Ou, S. Origin and Fate of Acrolein in Foods. Foods 2022, 11, 1976. https://doi.org/10.3390/foods11131976
Jiang K, Huang C, Liu F, Zheng J, Ou J, Zhao D, Ou S. Origin and Fate of Acrolein in Foods. Foods. 2022; 11(13):1976. https://doi.org/10.3390/foods11131976
Chicago/Turabian StyleJiang, Kaiyu, Caihuan Huang, Fu Liu, Jie Zheng, Juanying Ou, Danyue Zhao, and Shiyi Ou. 2022. "Origin and Fate of Acrolein in Foods" Foods 11, no. 13: 1976. https://doi.org/10.3390/foods11131976
APA StyleJiang, K., Huang, C., Liu, F., Zheng, J., Ou, J., Zhao, D., & Ou, S. (2022). Origin and Fate of Acrolein in Foods. Foods, 11(13), 1976. https://doi.org/10.3390/foods11131976