
The UHPLC-QTOF-MS Phenolic Profiling and Activity of Cydonia oblonga Mill. Reveals a Promising Nutraceutical Potential

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Extraction Procedure
2.3. Profiling of C. oblonga Parts by UHPLC-QTOF Mass Spectrometry
2.4. In Vitro Antioxidant Potential and Enzymes-Inhibitory Activities
2.5. Statistics and Chemometrics
3. Results and Discussion
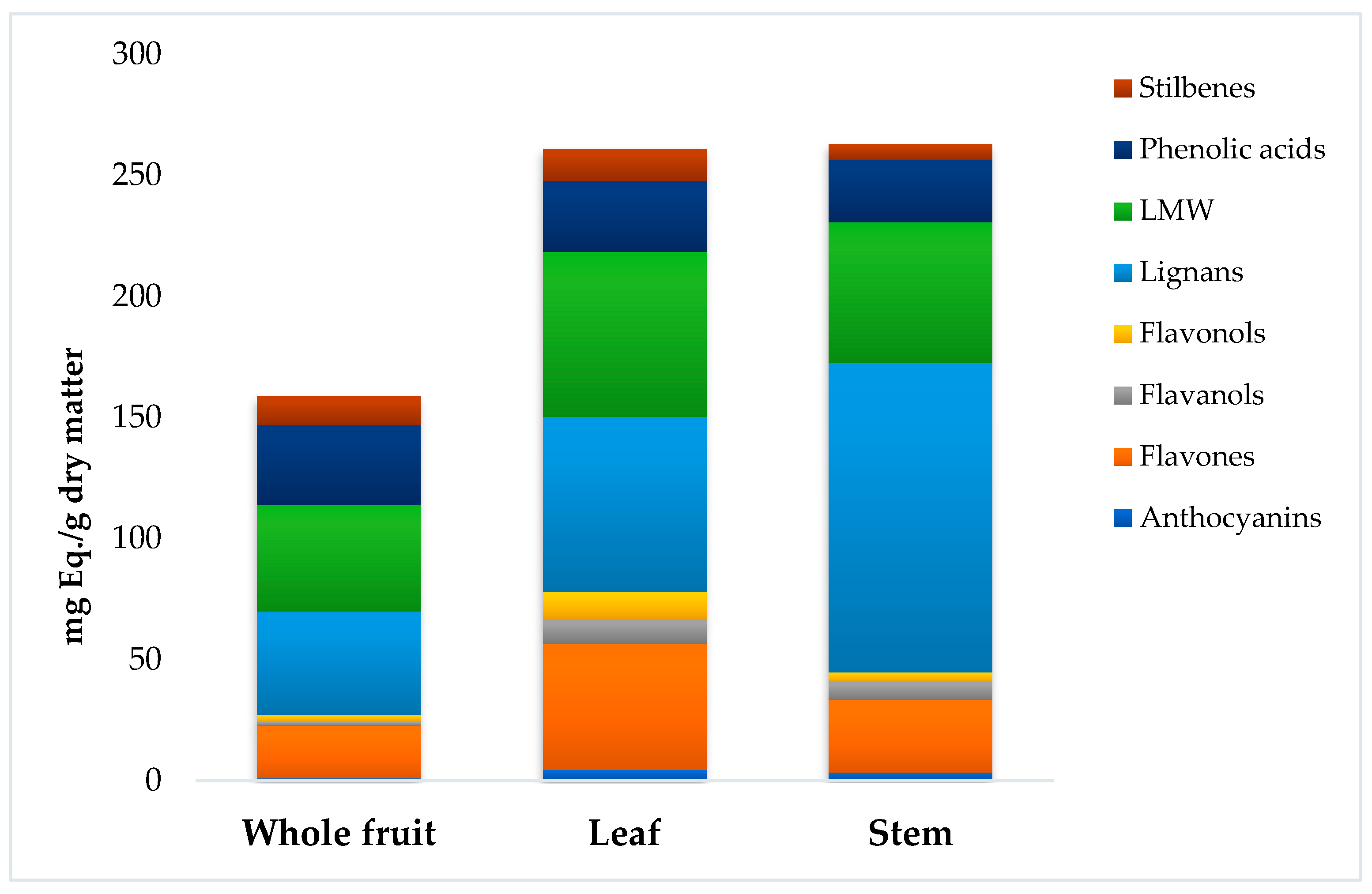
3.1. Phenolic Profiling of C. oblonga by UHPLC-QTOF Mass Spectrometry
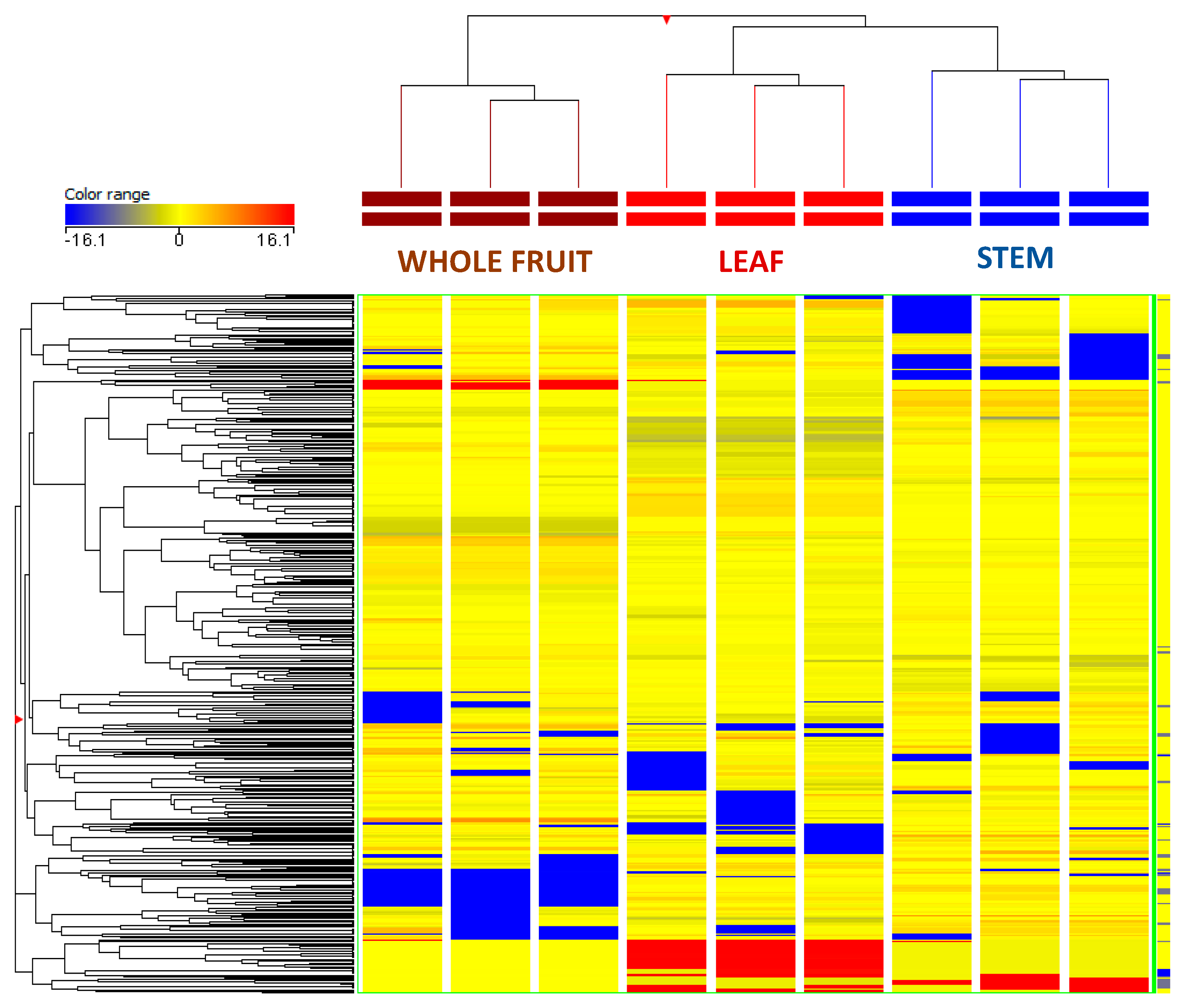
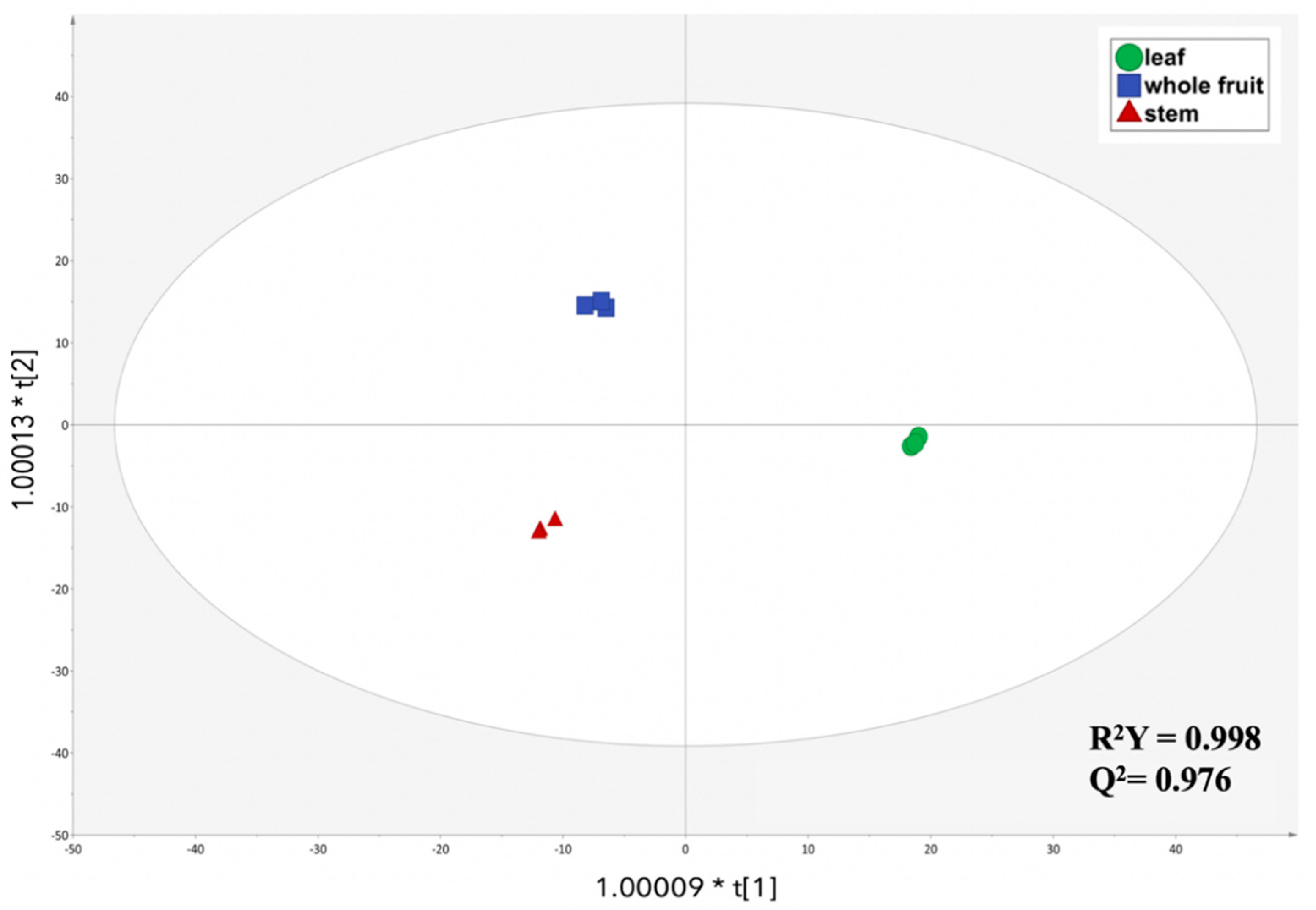
3.2. Multivariate Discrimination of the Different Plant Parts
3.3. In Vitro Antioxidant and Enzyme Inhibitory Activities
3.4. Correlations between Phytochemical Profiles and Biological Activities
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ashraf, M.U.; Muhammad, G.; Hussain, M.A.; Bukhari, S.N. Cydonia oblonga M., a medicinal plant rich in phytonutrients for pharmaceuticals. Front. Pharmacol. 2016, 7, 163. [Google Scholar] [CrossRef]
- Yildirim, A.; Oktay, M.; Bİlaloğlu, V. The antioxidant activity of the leaves of Cydonia vulgaris. Turk. J. Med. Sci. 2001, 31, 23–27. [Google Scholar]
- Lopes, M.M.D.A.; Sanches, A.G.; De Souza, K.O.; Silva, E.D.O. Quince—Cydonia oblonga. In Exotic Fruits; Elsevier: Amsterdam, The Netherlands, 2018; pp. 363–368. [Google Scholar]
- Griñán, I.; Galindo, A.; Rodríguez, P.; Morales, D.; Corell, M.; Centeno, A.; González, M.C.; Torrecillas, A.; Carbonell-Barrachina, A.A.; Hernández, F. Volatile composition and sensory and quality attributes of quince (Cydonia oblonga Mill.) fruits as affected by water stress. Sci. Hortic. 2019, 244, 68–74. [Google Scholar] [CrossRef]
- Rop, O.; Balík, J.; Řezníček, V.; Juríková, T.; Škardová, P.; Salaš, P.; Sochor, J.; Mlček, J.; Kramářová, D. Chemical characteristics of fruits of some selected quince (Cydonia oblonga Mill.) cultivars. Czech J. Food Sci. 2011, 29, 65–73. [Google Scholar] [CrossRef]
- Zapata, P.J.; Martínez-Esplá, A.; Gironés-Vilaplana, A.; Santos-Lax, D.; Noguera-Artiaga, L.; Carbonell-Barrachina, Á.A. Phenolic, volatile, and sensory profiles of beer enriched by macerating quince fruits. LWT 2019, 103, 139–146. [Google Scholar] [CrossRef]
- Mustafa, B.; Hajdari, A.; Pulaj, B.; Quave, C.L.; Pieroni, A. Medical and food ethnobotany among Albanians and Serbs living in the Shtërpcë/Štrpce area, South Kosovo. J. Herb. Med. 2020, 22, 100344. [Google Scholar] [CrossRef]
- Sabir, S.; Qureshi, R.; Arshad, M.; Amjad, M.S.; Fatima, S.; Masood, M.; Saboon; Chaudhari, S.K. Pharmacognostic and clinical aspects of Cydonia oblonga: A review. Asian Pac. J. Trop. Dis. 2015, 5, 850–855. [Google Scholar] [CrossRef]
- Costa, R.M.; Magalhães, A.S.; Pereira, J.A.; Andrade, P.B.; Valentão, P.; Carvalho, M.; Silva, B.M. Evaluation of free radical-scavenging and antihemolytic activities of quince (Cydonia oblonga) leaf: A comparative study with green tea (Camellia sinensis). Food Chem. Toxicol. 2009, 47, 860–865. [Google Scholar] [CrossRef]
- Oliveira, A.P.; Costa, R.M.; Magalhães, A.S.; Pereira, J.A.; Carvalho, M.; Valentão, P.; Andrade, P.B.; Silva, B.M. Targeted metabolites and biological activities of Cydonia oblonga Miller leaves. Food Res. Int. 2012, 46, 496–504. [Google Scholar] [CrossRef]
- Umar, A.; Iskandar, G.; Aikemu, A.; Yiming, W.; Zhou, W.; Berke, B.; Begaud, B.; Moore, N. Effects of Cydonia oblonga Miller leaf and fruit flavonoids on blood lipids and anti-oxydant potential in hyperlipidemia rats. J. Ethnopharmacol. 2015, 169, 239–243. [Google Scholar] [CrossRef]
- Sut, S.; Poloniato, G.; Maggi, F.; Malagoli, M.; Dall’Acqua, S. Preliminary evaluation of quince (Cydonia oblonga Mill.) fruit as extraction source of antioxidant phytoconstituents for nutraceutical and functional food applications. J. Sci. Food Agric. 2019, 99, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Rocchetti, G.; Zengin, G.; Ak, G.; Yıldıztugay, E.; Mahomoodally, M.F.; Picot-Allain, M.C.N.; Lucini, L. Profiling of polyphenols and sesquiterpenoids using different extraction methods in Muscari turcicum, an endemic plant from Turkey. Ind. Crop. Prod. 2020, 154, 112626. [Google Scholar] [CrossRef]
- Tsugawa, H.; Cajka, T.; Kind, T.; Ma, Y.; Higgins, B.T.; Ikeda, K.; Kanazawa, M.; Vander Gheynst, J.S.; Fiehn, O.; Arita, M. MS-DIAL: Data-independent MS/MS deconvolution for comprehensive metabolome analysis. Nat. Methods 2015, 12, 523–526. [Google Scholar] [CrossRef] [PubMed]
- Tsugawa, H.; Kind, T.; Nakabayashi, R.; Yukihira, D.; Tanaka, W.; Cajka, T.; Saito, K.; Fiehn, O.; Arita, M. Hydrogen Rearrangement Rules: Computational MS/MS Fragmentation and Structure Elucidation Using MS-FINDER Software. Anal. Chem. 2016, 88, 7946–7958. [Google Scholar] [CrossRef]
- Rocchetti, G.; Gallo, A.; Nocetti, M.; Lucini, L.; Masoero, F. Milk metabolomics based on ultra-high-performance liquid chromatography coupled with quadrupole time-of-flight mass spectrometry to discriminate different cows feeding regimens. Food Res. Int. 2020, 134, 109279. [Google Scholar] [CrossRef]
- Zengin, G.; Locatelli, M.; Carradori, S.; Mocan, A.M.; Aktumsek, A. Total Phenolics, Flavonoids, Condensed Tannins Content of Eight Centaurea Species and Their Broad Inhibitory Activities against Cholinesterase, Tyrosinase, α-Amylase and α-Glucosidase. Not. Bot. Horti Agrobot. Cluj. 2016, 44, 195–200. [Google Scholar] [CrossRef]
- Rocchetti, G.; Tomas, M.; Zhang, L.; Zengin, G.; Lucini, L.; Capanoglu, E. Red beet (Beta vulgaris) and amaranth (Amaranthus sp.) microgreens: Effect of storage and in vitro gastrointestinal digestion on the untargeted metabolomic profile. Food Chem. 2020, 332, 127415. [Google Scholar] [CrossRef]
- Ferreres, F.; Silva, B.M.; Andrade, P.B.; Seabra, R.M.; Ferreira, M.A. Approach to the study of C-glycosyl flavones by ion trap HPLC-PAD-ESI/MS/MS: Application to seeds of quince (Cydonia oblonga). Phytochem. Anal. 2003, 14, 352–359. [Google Scholar] [CrossRef]
- Silva, B.M.; Andrade, P.B.; Martins, R.C.; Seabra, R.M.; Ferreira, M.A. Principal component analysis as tool of characterization of quince (Cydonia oblonga Miller) jam. Food Chem. 2006, 94, 504–512. [Google Scholar] [CrossRef]
- Wojdyło, A.; Oszmiański, J.; Bielicki, P. Polyphenolic Composition, Antioxidant Activity, and Polyphenol Oxidase (PPO) Activity of Quince (Cydonia oblonga Miller) Varieties. J. Agric. Food Chem. 2013, 61, 2762–2772. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Feng, J.-Q.; Zhao, W.-M. A new lignan and anti-inflammatory flavonoids from Kerria japonica. J. Asian Nat. Prod. Res. 2008, 10, 435–438. [Google Scholar] [CrossRef]
- Yeo, H.; Chin, Y.-W.; Park, S.-Y.; Kim, J. Lignans of Rosa multiflora roots. Arch. Pharmacal Res. 2004, 27, 287–290. [Google Scholar] [CrossRef]
- El-Seedi, H.R.; El-Said, A.M.A.; Khalifa, S.A.M.; Göransson, U.; Bohlin, L.; Borg-Karlson, A.-K.; Verpoorte, R. Biosynthesis, Natural Sources, Dietary Intake, Pharmacokinetic Properties, and Biological Activities of Hydroxycinnamic Acids. J. Agric. Food Chem. 2012, 60, 10877–10895. [Google Scholar] [CrossRef]
- Stojanović, B.T.; Mitić, S.S.; Stojanović, G.S.; Mitić, M.N.; Kostić, D.A.; Paunović, D.Ð.; Arsić, B.B.; Pavlović, A.N. Phenolic profiles and metal ions analyses of pulp and peel of fruits and seeds of quince (Cydonia oblonga Mill.). Food Chem. 2017, 232, 466–475. [Google Scholar] [CrossRef]
- Magalhães, A.S.; Silva, B.M.; Pereira, J.A.; Andrade, P.B.; Valentão, P.; Carvalho, M. Protective effect of quince (Cydonia oblonga Miller) fruit against oxidative hemolysis of human erythrocytes. Food Chem. Toxicol. 2009, 47, 1372–1377. [Google Scholar] [CrossRef] [PubMed]
- Silva, B.M.; Andrade, P.B.; Valentão, P.; Ferreres, F.; Seabra, R.M.; Ferreira, M.A. Quince (Cydonia oblonga Miller) Fruit (Pulp, Peel, and Seed) and Jam: Antioxidant Activity. J. Agric. Food Chem. 2004, 52, 4705–4712. [Google Scholar] [CrossRef]
- Dos Santos, T.C.; Gomes, T.M.; Pinto, B.A.S.; Camara, A.L.; Paes, A.M.D.A. Naturally Occurring Acetylcholinesterase Inhibitors and Their Potential Use for Alzheimer’s Disease Therapy. Front. Pharmacol. 2018, 9, 1192. [Google Scholar] [CrossRef]
- Saxena, M. Target Enzyme in Alzheimer’s Disease: Acetylcholinesterase Inhibitors. Curr. Top. Med. Chem. 2019, 19, 264–275. [Google Scholar] [CrossRef]
- Cuong, T.D.; Hung, T.M.; Han, H.Y.; Roh, H.S.; Seok, J.-H.; Lee, J.K.; Jeong, J.Y.; Choi, J.S.; Kim, J.A.; Min, B.S. Potent Acetylcholinesterase Inhibitory Compounds from Myristica fragrans. Nat. Prod. Commun. 2014, 9, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Hung, T.M.; Na, M.; Min, B.S.; Ngoc, T.M.; Lee, I.; Zhang, X.; Bae, K. Acetylcholinesterase inhibitory effect of lignans isolated from Schizandra chinensis. Arch. Pharmacal Res. 2007, 30, 685–690. [Google Scholar] [CrossRef] [PubMed]
- Johnson, G. The Peripheral Anionic Site of Acetylcholinesterase: Structure, Functions and Potential Role in Rational Drug Design. Curr. Pharm. Des. 2006, 12, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Aslan, M.; Orhan, N.; Orhan, D.D.; Ergun, F. Hypoglycemic activity and antioxidant potential of some medicinal plants traditionally used in Turkey for diabetes. J. Ethnopharmacol. 2010, 128, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Nyambe-Silavwe, H.; Williamson, G. Chlorogenic and phenolic acids are only very weak inhibitors of human salivary α-amylase and rat intestinal maltase activities. Food Res. Int. 2018, 113, 452–455. [Google Scholar] [CrossRef] [PubMed]
- Pillaiyar, T.; Manickam, M.; Namasivayam, V. Skin whitening agents: Medicinal chemistry perspective of tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 403–425. [Google Scholar] [CrossRef]
- Zolghadri, S.; Bahrami, A.; Khan, M.T.H.; Munoz-Munoz, J.; Garcia-Molina, F.; Garcia-Canovas, F.; Saboury, A.A. A comprehensive review on tyrosinase inhibitors. J. Enzym. Inhib. Med. Chem. 2019, 34, 279–309. [Google Scholar] [CrossRef]
- Panzella, L.; Napolitano, A. Natural and Bioinspired Phenolic Compounds as Tyrosinase Inhibitors for the Treatment of Skin Hyperpigmentation: Recent Advances. Cosmetics 2019, 6, 57. [Google Scholar] [CrossRef]
- Uesugi, D.; Hamada, H.; Shimoda, K.; Kubota, N.; Ozaki, S.-I.; Nagatani, N. Synthesis, oxygen radical absorbance capacity, and tyrosinase inhibitory activity of glycosides of resveratrol, pterostilbene, and pinostilbene. Biosci. Biotechnol. Biochem. 2017, 81, 226–230. [Google Scholar] [CrossRef]
- Srisayam, M.; Weerapreeyakul, N.; Kanokmedhakul, K. Inhibition of two stages of melanin synthesis by sesamol, sesamin and sesamolin. Asian Pac. J. Trop. Biomed. 2017, 7, 886–895. [Google Scholar] [CrossRef]
- Wu, B.; Zhang, X.; Wu, X. New lignan glucosides with tyrosinase inhibitory activities from exocarp of Castanea henryi. Carbohydr. Res. 2012, 355, 45–49. [Google Scholar] [CrossRef]
- Aramwit, P.; Bang, N.; Srichana, T. The properties and stability of anthocyanins in mulberry fruits. Food Res. Int. 2010, 43, 1093–1097. [Google Scholar] [CrossRef]
- Koyu, H.; Kazan, A.; Demir, S.; Haznedaroglu, M.Z.; Yesil-Celiktas, O. Optimization of microwave assisted extraction of Morus nigra L. fruits maximizing tyrosinase inhibitory activity with isolation of bioactive constituents. Food Chem. 2018, 248, 183–191. [Google Scholar] [CrossRef]
- Hwang, J.-M.; Kuo, H.-C.; Lin, C.-T.; Kao, E.-S. Inhibitory effect of liposome-encapsulated anthocyanin on melanogenesis in human melanocytes. Pharm. Biol. 2013, 51, 941–947. [Google Scholar] [CrossRef] [PubMed]
- Jhan, J.; Chung, Y.; Chen, G.; Chang, C.; Lu, Y.; Hsu, C. Anthocyanin contents in the seed coat of black soya bean and their anti-human tyrosinase activity and antioxidative activity. Int. J. Cosmet. Sci. 2016, 38, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Liu, K.; Liang, Y.; Liu, G.; Sang, J.; Li, C. Extraction optimization and purification of anthocyanins from Lycium ruthenicum Murr. and evaluation of tyrosinase inhibitory activity of the anthocyanins. J. Food Sci. 2020, 85, 696–706. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.C.; Sathisha, U.V.; Dharmesh, S.M.; Rao, A.A.; Singh, S.A. Effect of sesamol, a potent tyrosinase inhibitor, on melanin and melanin synthesizing melanoma cells. FASEB J. 2008, 22, 1062.1. [Google Scholar] [CrossRef]
- Fernandes, I.; Marques, C.; Évora, A.; Faria, A.; Calhau, C.; Mateus, N.; De Freitas, V. Anthocyanins: Nutrition and Health; Springer: Cham, Switzerland, 2018; pp. 1–37. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Class | Sub-Class | Discriminant Compound (OPLS-DA) | VIP Score ± SE | Log FC [Leaf] vs. [Stem] | Log FC [Leaf] vs. [Whole Fruit] | Log FC [Stem] vs. [Whole Fruit] |
|---|---|---|---|---|---|---|
| Flavonoids | Anthocyanins | Petunidin 3-O-glucoside | 1.23 ± 0.31 | −2.63 | 1.98 | 4.61 |
| Delphinidin 3-O-arabinoside/Delphinidin 3-O-xyloside | 1.23 ± 0.30 | −2.45 | −0.08 | 2.37 | ||
| Cyanidin 3-O-xyloside/Cyanidin 3-O-arabinoside | 1.22 ± 0.33 | −2.55 | −0.22 | 2.33 | ||
| Cyanidin 3-O-(6″-malonyl−3″-glucosyl-glucoside) | 1.22 ± 0.32 | 0.68 | −1.43 | −2.11 | ||
| Cyanidin 3-O-sambubioside 5-O-glucoside | 1.21 ± 0.35 | −5.03 | −2.14 | 2.89 | ||
| Pelargonidin 3-O-sambubioside | 1.21 ± 0.33 | −7.07 | 5.10 | 12.16 | ||
| Peonidin 3-O-(6″-acetyl-glucoside) | 1.21 ± 0.33 | −3.58 | 0.22 | 3.81 | ||
| Cyanidin 3-O-xylosyl-rutinoside | 1.21 ± 0.27 | −1.18 | 16.11 | 17.29 | ||
| Dihydrochalcones | 3-Hydroxyphloretin 2′-O-xylosyl-glucoside | 1.20 ± 0.32 | −4.04 | −2.99 | 1.05 | |
| Dihydroquercetin 3-O-rhamnoside | 1.21 ± 0.37 | −3.72 | −1.56 | 2.16 | ||
| Flavanols | Theaflavin | 1.21 ± 0.34 | −21.79 | −18.84 | 2.96 | |
| Flavanones | Eriodictyol | 1.22 ± 0.34 | −1.60 | 7.36 | 8.97 | |
| Eriodictyol 7-O-glucoside | 1.21 ± 0.37 | −3.72 | −1.56 | 2.16 | ||
| Flavones | Cannflavin A | 1.21 ± 0.34 | 1.13 | −1.36 | −2.49 | |
| Chrysoeriol 7-O-(6″-malonyl-glucoside) | 1.21 ± 0.33 | −7.01 | −0.43 | 6.58 | ||
| Luteolin 7-O-glucuronide | 1.21 ± 0.33 | −21.43 | −1.39 | 20.04 | ||
| Cirsimaritin | 1.20 ± 0.27 | −0.16 | −2.97 | −2.80 | ||
| Flavonols | Quercetin 3-O-xylosyl-glucuronide | 1.23 ± 0.28 | −1.92 | 16.55 | 18.47 | |
| Kaempferol 3-O-(2″-rhamnosyl−6″-acetyl-galactoside) 7-O-rhamnoside | 1.22 ± 0.31 | −2.21 | 16.25 | 18.45 | ||
| Quercetin 3-O-xylosyl-rutinoside | 1.21 ± 0.35 | −3.42 | −0.97 | 2.45 | ||
| Kaempferol 3-O-glucuronide | 1.21 ± 0.33 | −21.41 | −1.39 | 20.02 | ||
| Kaempferide | 1.21 ± 0.33 | −10.61 | −7.96 | 2.65 | ||
| Myricetin | 1.20 ± 0.27 | 17.61 | −4.19 | −21.80 | ||
| Isoflavonoids | Glycitin | 1.22 ± 0.32 | −3.02 | 7.17 | 10.19 | |
| 6″-O-Malonylgenistin | 1.20 ± 0.32 | −5.45 | −4.09 | 1.36 | ||
| Phenolic acids | Hydroxybenzoic acids | Protocatechuic acid / 3,5-Dihydroxybenzoic acid | 1.22 ± 0.34 | 19.83 | −1.75 | −21.58 |
| Hydroxycinnamic acids | p-Coumaroylquinic acid/3-p-Coumaroylquinic acid/4-p-Coumaroylquinic acid/5-p-Coumaroylquinic acid | 1.22 ± 0.33 | −3.97 | −0.78 | 3.18 | |
| Stigmastanol ferulate | 1.20 ± 0.44 | −2.85 | 1.85 | 19.70 | ||
| 3,5-Dicaffeoylquinic acid/4,5-Dicaffeoylquinic acid/3,4-Dicaffeoylquinic acid | 1.20 ± 0.33 | −4.09 | −1.86 | 2.23 |
| DPPH (mg TE/g) | ABTS (mg TE/g) | CUPRAC (mg TE/g) | FRAP (mg TE/g) | Metal Chelating mg (EDTAE/g) | Phosphomolybdenum (mmol TE/g) | |
|---|---|---|---|---|---|---|
| Leaf | 189.53 ± 0.39 a | 285.65 ± 4.90 a | 348.84 ± 1.87 a | 158.86 ± 2.90 a | 10.80 ± 0.15 a | 2.31 ± 0.17 a |
| Whole fruits | 6.53 ± 0.31 c | 9.15 ± 1.07 c | 24.84 ± 0.29 c | 13.66 ± 0.03 c | 11.15 ± 0.58 a | 0.56 ± 0.03 a |
| Stem | 129.68 ± 2.31 b | 174.32 ± 5.66 b | 215.60 ± 3.38 b | 120.86 ± 2.09 b | 11.66 ± 0.14 a | 1.85 ± 0.06 a |
| AChE (mg GALAE/g) | BChE (mg GALAE/g) | Tyrosinase (mg KAE/g) | α-Amylase (mmol ACAE/g) | α-Glucosidase (mmol ACAE/g) | |
|---|---|---|---|---|---|
| Leaf | 2.37 ± 0.04 a | 2.35 ± 0.07 b | 68.32 ± 0.30 b | 0.48 ± 0.00 a | n.d |
| Whole fruits | 2.36 ± 0.02 a | 2.94 ± 0.13 a | 56.64 ± 0.12 c | 0.29 ± 0.03 a | 1.36 ± 0.09 |
| Stem | 2.54 ± 0.02 a | 2.95 ± 0.07 a | 72.11 ± 0.26 a | 0.43 ± 0.00 a | n.d. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Rocchetti, G.; Zengin, G.; Ak, G.; Saber, F.R.; Montesano, D.; Lucini, L. The UHPLC-QTOF-MS Phenolic Profiling and Activity of Cydonia oblonga Mill. Reveals a Promising Nutraceutical Potential. Foods 2021, 10, 1230. https://doi.org/10.3390/foods10061230
Zhang L, Rocchetti G, Zengin G, Ak G, Saber FR, Montesano D, Lucini L. The UHPLC-QTOF-MS Phenolic Profiling and Activity of Cydonia oblonga Mill. Reveals a Promising Nutraceutical Potential. Foods. 2021; 10(6):1230. https://doi.org/10.3390/foods10061230
Chicago/Turabian StyleZhang, Leilei, Gabriele Rocchetti, Gökhan Zengin, Gunes Ak, Fatema R. Saber, Domenico Montesano, and Luigi Lucini. 2021. "The UHPLC-QTOF-MS Phenolic Profiling and Activity of Cydonia oblonga Mill. Reveals a Promising Nutraceutical Potential" Foods 10, no. 6: 1230. https://doi.org/10.3390/foods10061230
APA StyleZhang, L., Rocchetti, G., Zengin, G., Ak, G., Saber, F. R., Montesano, D., & Lucini, L. (2021). The UHPLC-QTOF-MS Phenolic Profiling and Activity of Cydonia oblonga Mill. Reveals a Promising Nutraceutical Potential. Foods, 10(6), 1230. https://doi.org/10.3390/foods10061230