
Germination Requirement and Suitable Storage Method of Hydrocharis dubia Seeds

 ,
,  ,
,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
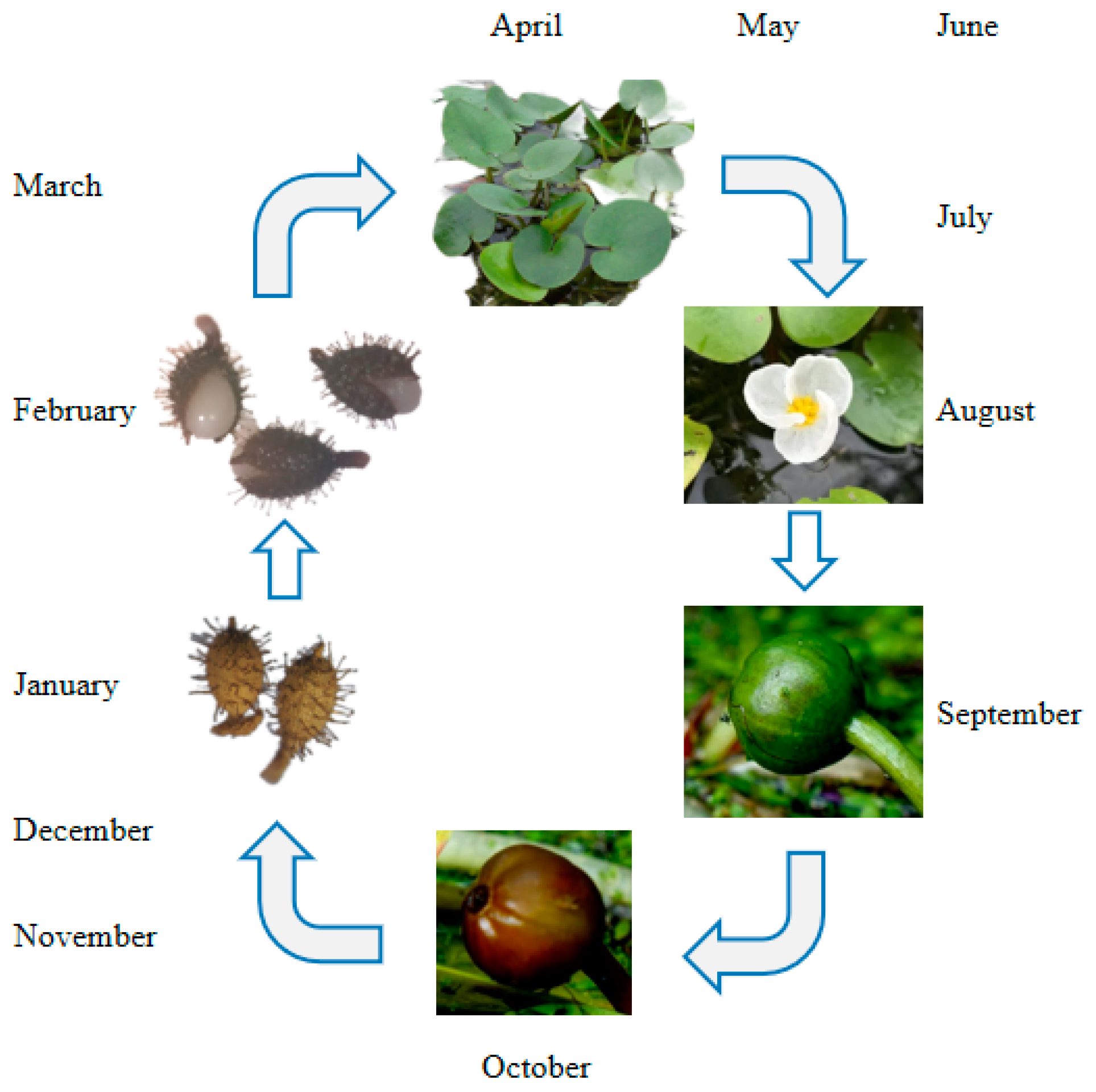
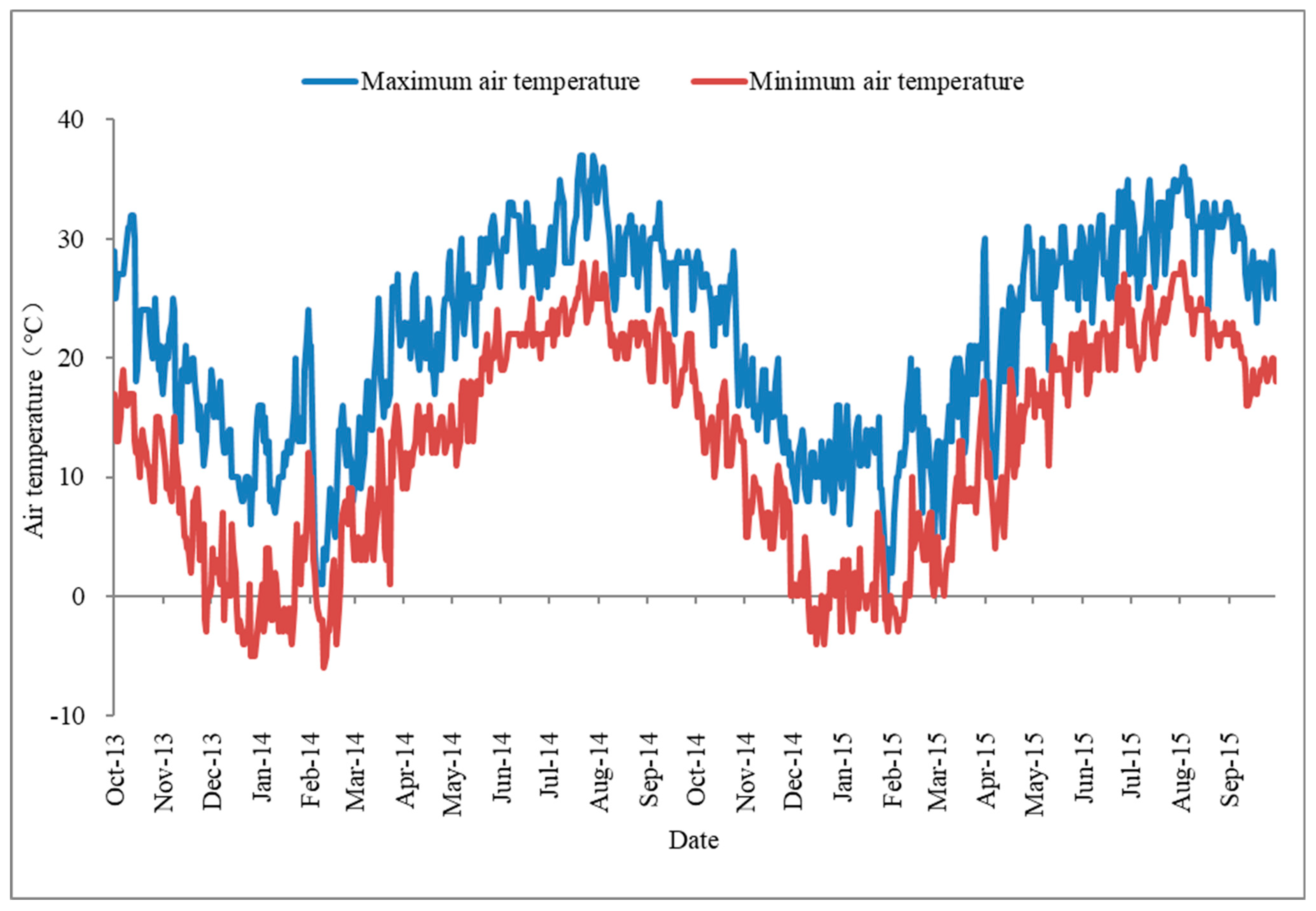
2.2. Seed Collection
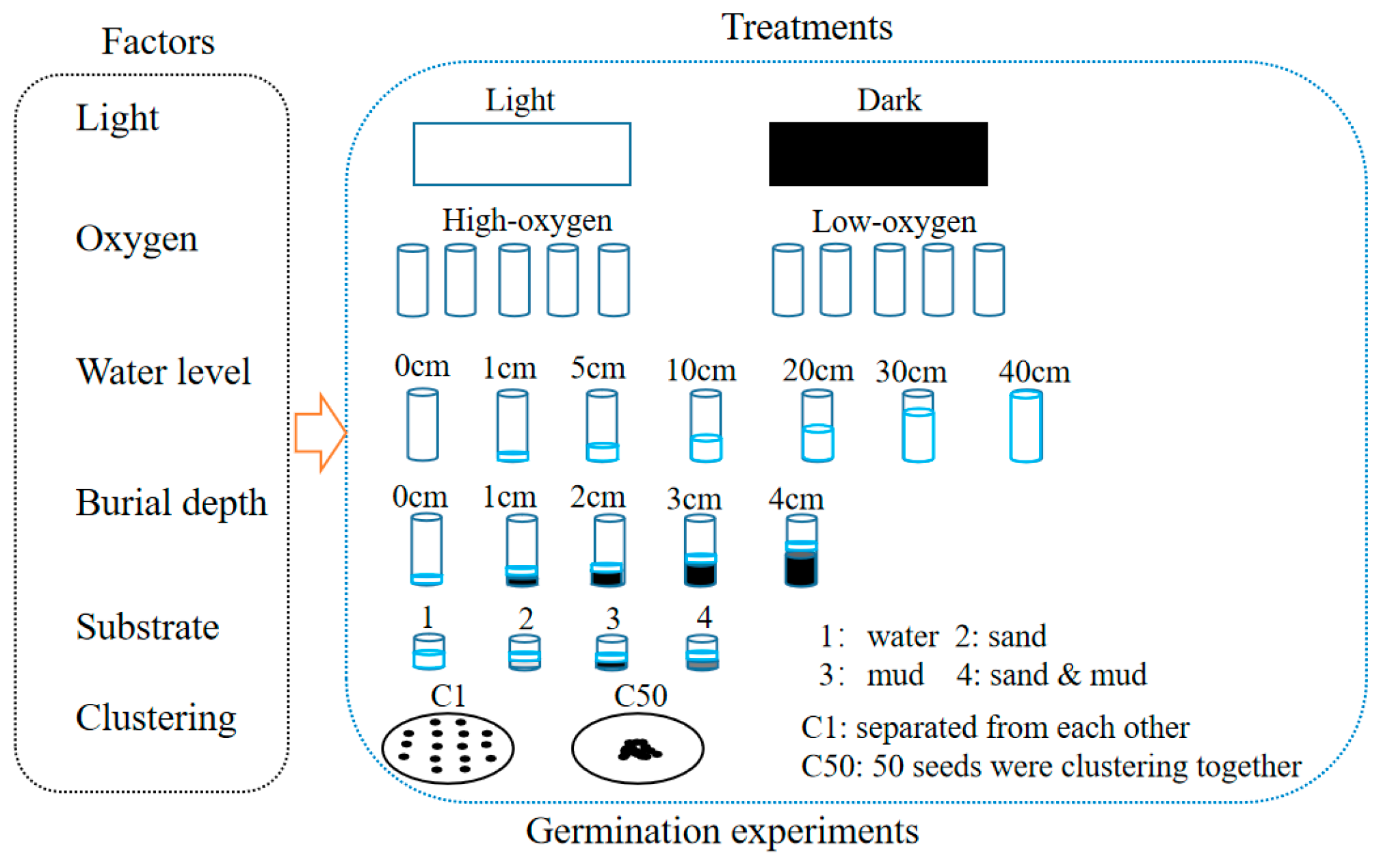
2.3. Germination Experiments
2.3.1. Effect of Light Availability on Germination
2.3.2. Effect of Oxygen Availability on Germination
2.3.3. Effect of Water Level on Germination
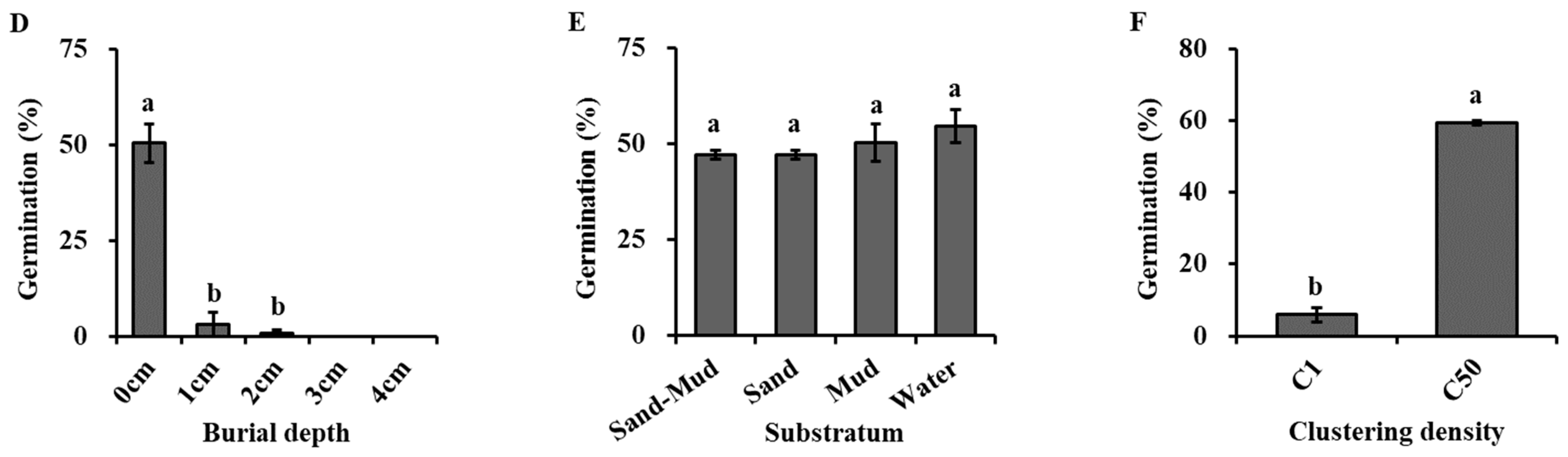
2.3.4. Effect of Burial Depth on Germination
2.3.5. Effect of Substratum on Germination
2.3.6. Effect of Clustering Density on Germination
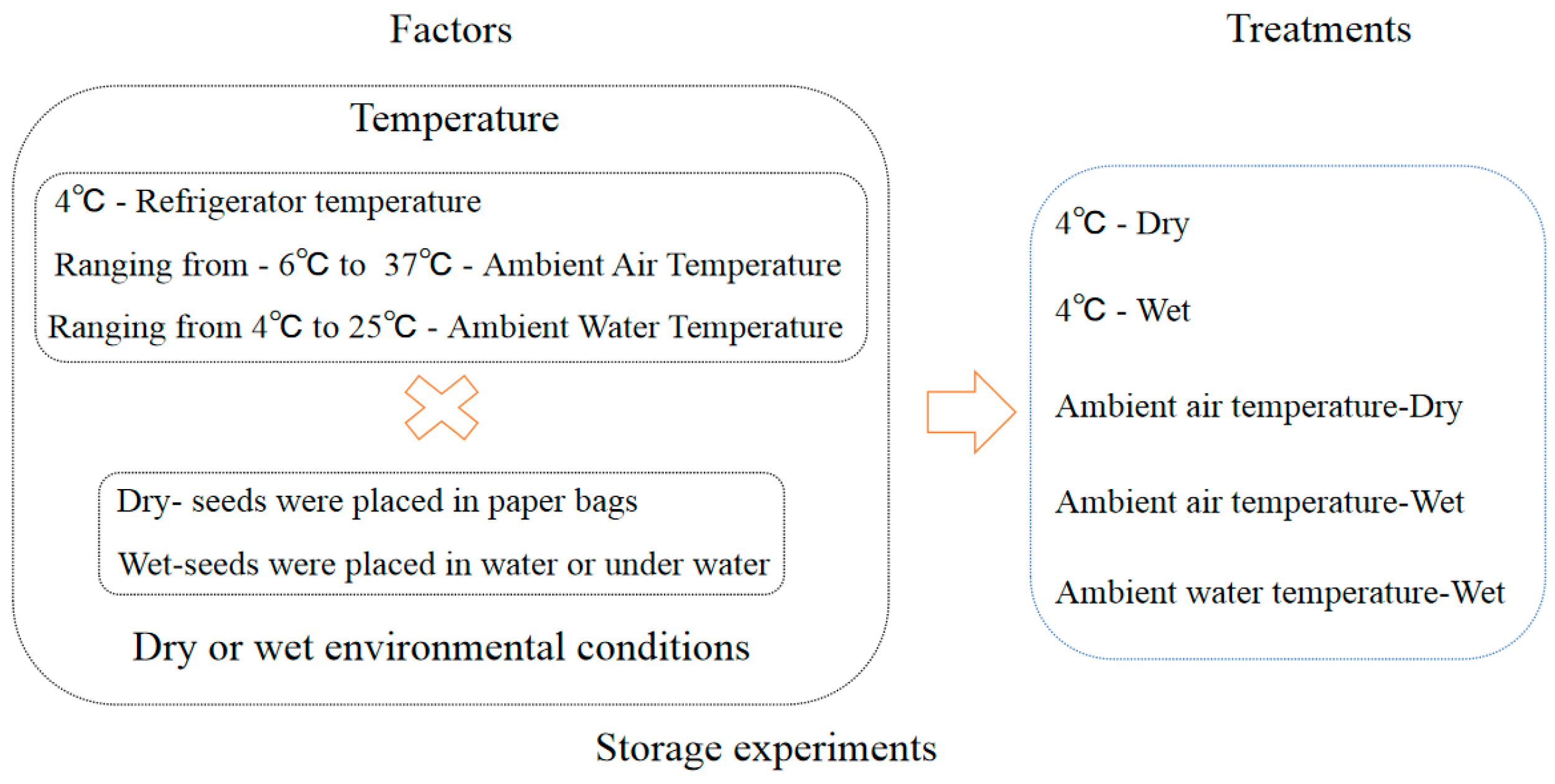
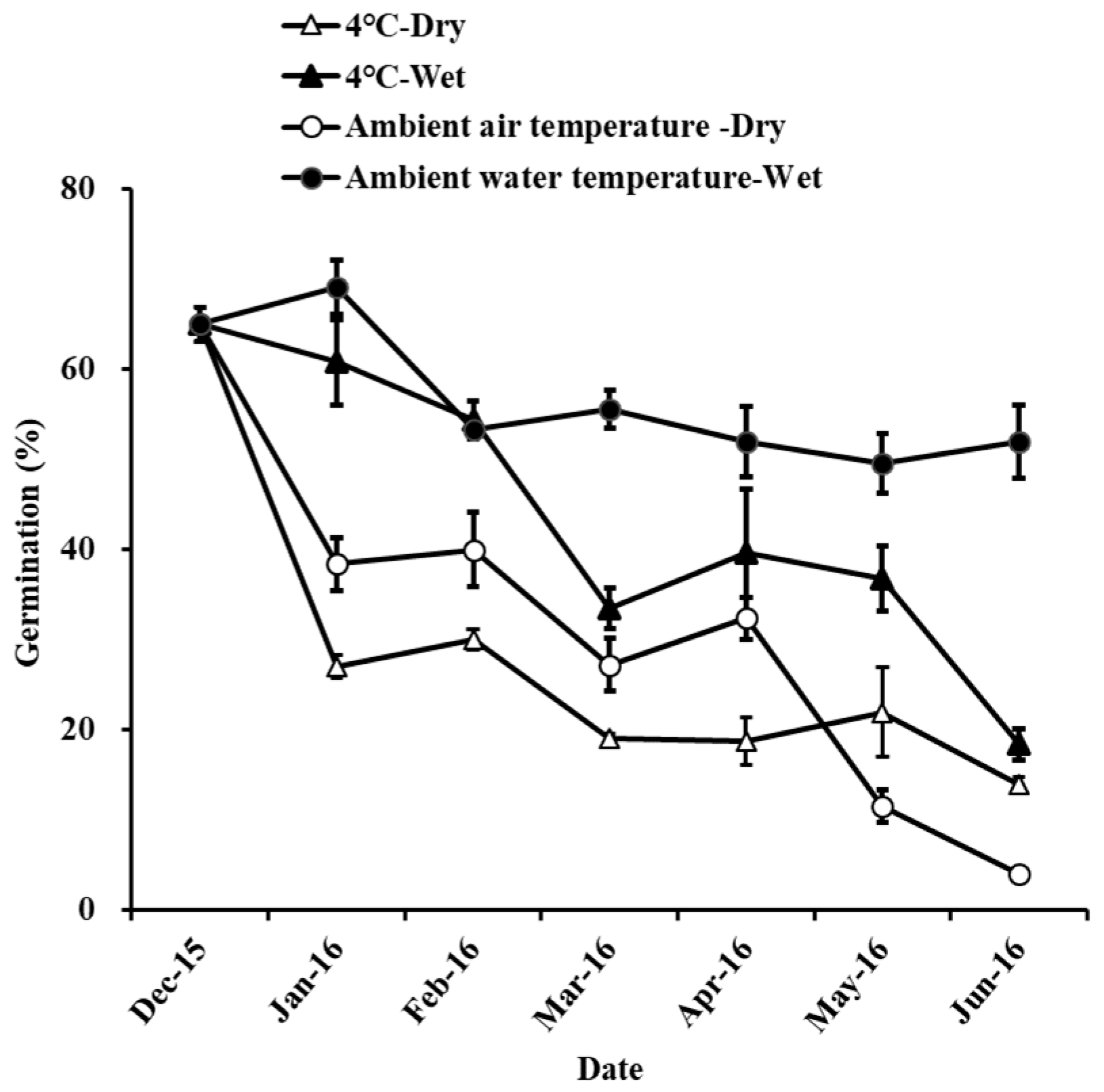
2.4. Storage Experiments
3. Data Analysis
4. Results
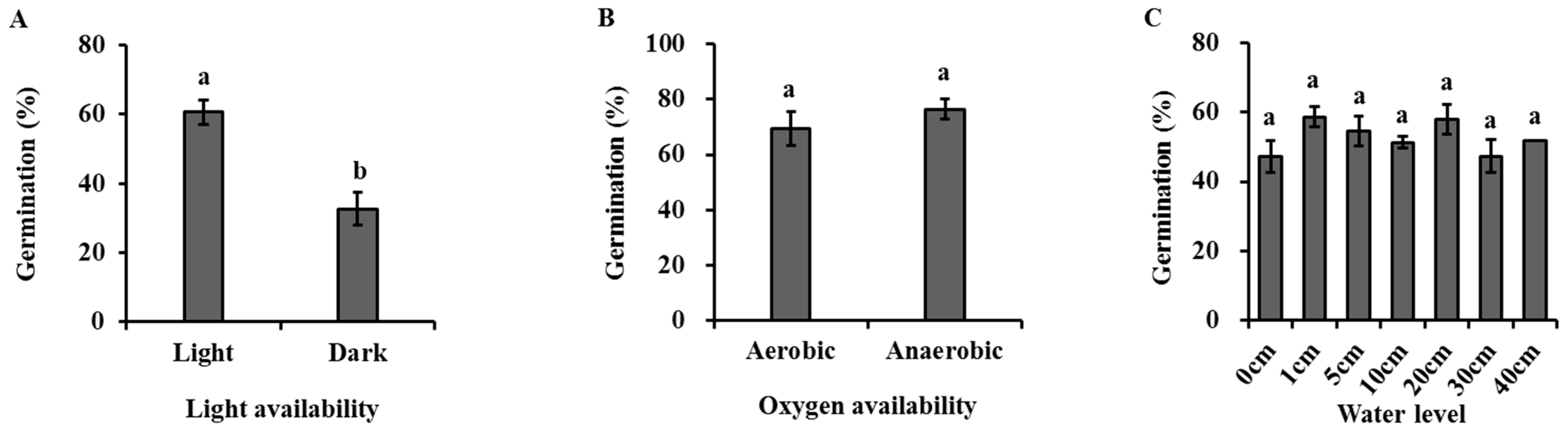
4.1. Germination Requirement in Seeds of H. dubia
4.2. Suitable Storage Method in Seeds of H. dubia
5. Discussion
6. Conclusions and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A


References
- Troia, A. Macrophytes in Inland Waters: From Knowledge to Management. Plants 2023, 12, 582. [Google Scholar] [CrossRef] [PubMed]
- Chao, C.X.; Wang, L.G.; Li, Y.; Yan, Z.W.; Liu, H.M.; Yu, D.; Liu, C.H. Response of sediment and water microbial communities to submerged vegetations restoration in a shallow eutrophic lake. Sci. Total Environ. 2021, 801, 149701. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.T.; Wang, G.D.; Zhao, M.L.; Wang, M.; Xue, Z.S.; Liu, B.; Jiang, M. Seed limitation and saline-alkaline stress restrict wetland restoration potential in the Songnen Plain, northeastern China. Ecol. Indic. 2021, 129, 107998. [Google Scholar] [CrossRef]
- Zhao, D.J.; Sun, M.Q.; Xue, Y.W.; Yang, Q.M.; Liu, B.H.; Jia, B.Q.; Song, C.H.; Zhang, S.H.; Zhang, Z.Q. Spatial variations of plant species diversity in urban soil seed banks in Beijing, China: Implications for plant regeneration and succession. Urban For. Urban Green. 2023, 86, 128012. [Google Scholar] [CrossRef]
- Li, Z.Q.; Lu, W.; Yang, L.; Kong, X.H.; Deng, X.W. Seed weight and germination behavior of the submerged plant Potamogeton pectinatus in the arid zone of northwest China. Ecol. Evol. 2015, 5, 1504–1512. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.T.; Zhang, R.F.; Liu, Y.; Yin, L.Y.; Wang, C.Y.; Li, W. The effect of storage condition on seed germination of six Hydrocharitaceae and Potamogetonaceae species. Aquat. Bot. 2017, 143, 49–53. [Google Scholar] [CrossRef]
- Rosbakh, S.; Poschlod, P. Broad Seed Germination Niche as an Adaptation to Heterogeneous Aquatic Habit-a Case Study of Four Potamogeton species. Inland Water Biol. 2019, 12, 68–73. [Google Scholar] [CrossRef]
- Temmink, R.J.M.; Dorenbosch, M.; Lamers, L.P.M.; Smolders, A.J.P.; Rip, W.; Lengkeek, W.; Didderen, K.; Fivash, G.S.; Bouma, T.J.; van der Heide, T. Growth forms and life-history strategies predict the occurrence of aquatic macrophytes in relation to environmental factors in a shallow peat lake complex. Hydrobiologia 2021, 848, 3987–3999. [Google Scholar] [CrossRef]
- Rohal, C.B.; Adams, C.R.; Martin, C.W.; Tevlin, S.; Reynolds, L.K. Intraspecific variation in Potamogeton illinoensis life history and seed germination has implications for restoration in eutrophic lakes. Hydrobiologia 2022, 849, 3481–3500. [Google Scholar] [CrossRef]
- Fraser, L.H.; Mulac, K.; Moore, F.B.G. Germination of 14 freshwater wetland plants as affected by oxygen and light. Aquat. Bot. 2014, 114, 29–34. [Google Scholar] [CrossRef]
- Vázquez-Benavides, J.; Moreno-Casasola, P.; López-Rosas, H. Effect of the grass Leersia hexandra on the dispersal, seed germination, and establishment of Pachira aquatica seedlings. Freshwater Biol. 2020, 65, 1702–1717. [Google Scholar] [CrossRef]
- Waite, B.; Statton, J.; Kendrick, G.A. Temperature Stratification and Monochromatic Light Break Dormancy and Facilitate On-Demand In Situ Germination in the Seagrass Halophila ovalis, with Seed Viability Determined by a Novel X-ray Analysis. Estuaries Coasts 2021, 44, 412–421. [Google Scholar] [CrossRef]
- Zhao, F.B.; Fang, X.; Zhao, Z.Y.; Chai, X.L. Effects of Water Level Fluctuations on the Growth Characteristics and Community Succession of Submerged Macrophytes: A Case Study of Yilong Lake, China. Water 2021, 13, 2900. [Google Scholar] [CrossRef]
- Ke, X.S.; Li, W. Germination requirement of Vallisneria natans seeds: Implications for restoration in Chinese lakes. Hydrobiologia 2006, 559, 357–362. [Google Scholar] [CrossRef]
- Yin, L.Y.; Zhang, R.F.; Xie, Z.M.; Wang, C.Y.; Li, W. The effect of temperature, substrate, light, oxygen availability and burial depth on Ottelia alismoides seed germination. Aquat. Bot. 2013, 111, 50–53. [Google Scholar] [CrossRef]
- Dunne, M.A.; Newman, R.M. Effect of light on macrophyte sprouting and assessment of viable seedbank to predict community composition. J. Aquat. Plant Manage. 2019, 57, 90–98. [Google Scholar]
- Zhang, X.K.; Liu, X.Q.; Wang, H.Z. Developing water level regulation strategies for macrophytes restoration of a large river-disconnected lake, China. Ecol. Eng. 2014, 68, 25–31. [Google Scholar] [CrossRef]
- Murray, B.R. Density-dependent germination and the role of seed leachate. Aust. J. Ecol. 1998, 23, 411–418. [Google Scholar] [CrossRef]
- Yin, L.Y.; Wang, C.Y.; Chen, Y.Y.; Yu, C.; Cheng, Y.; Li, W. Cold stratification, light and high seed density enhance the germination of Ottelia alismoides. Aquat. Bot. 2009, 90, 85–88. [Google Scholar] [CrossRef]
- Imanishi, A.; Imanishi, J. Seed dormancy and germination traits of an endangered aquatic plant species, Euryale ferox Salisb. (Nymphaeaceae). Aquat. Bot. 2014, 119, 80–83. [Google Scholar] [CrossRef]
- Wagner, E.J.; Oplinger, R.W. Effect of overwinter hydration, seed storage time, temperature, photoperiod, water depth, and scarification on seed germination of some Schoenoplectus, Polygonum, Eleocharis and Alisma species. Aquat. Bot. 2017, 136, 164–174. [Google Scholar] [CrossRef]
- Moore, K.A.; Orth, R.J.; Nowak, J.F. Environmental-regulation of seed-germination in Zostera-marina L (eelgrass) in chesapeake bay-effects of light, oxygen and sediment burial. Aquat. Bot. 1993, 45, 79–91. [Google Scholar] [CrossRef]
- Ruiz-Merino, M.; Campos-Cuéllar, R.; Germán-Gómez, A.; Aponte, H. Characteristics, natural history, and applications of Hydrocharis laevigata: A review. Caldasia 2022, 44, 432–441. [Google Scholar] [CrossRef]
- Cutter, E.G.; Hung, C.Y. Symmetric and asymmetric mitosis and cytokinesis in root tip of Hydrocharis-morsus-ranae L. J. Cell Sci. 1972, 11, 723. [Google Scholar] [CrossRef] [PubMed]
- Cook, C.D.K.; Luond, R. A revision of the genus Hydrocharis (Hydrocharitaceae). Aquat. Bot. 1982, 14, 177–204. [Google Scholar] [CrossRef]
- Efremov, A.N.; Sviridenko, B.F.; Li, Z.Z.; Mesterhazy, A.; Tchatchouange, E.N.; Toma, C.; Grishina, V.S.; Murashko, Y.A. Genus Hydrocharis (Hydrocharitaceae): Ecology of Communities and Their Tolerance to Abiotic Factors. Inland Water Biol. 2023, 16, 428–438. [Google Scholar] [CrossRef]
- Efremov, A.N.; Grishina, V.S.; Toma, C.; Mesterházy, A.; Tchatchouang, E.N. Comparative Morphology of the Genus Hydrocharis L. (Hydrocharitaceae). Inland Water Biol. 2021, 14, 638–651. [Google Scholar] [CrossRef]
- Catling, P.M.; Mitrow, G.; Haber, E.; Posluszny, U.; Charlton, W.A. The biology of Canadian weeds: Hydrocharis morsus-ranae L. Can. J. Plant Sci. 2003, 83, 1001–1016. [Google Scholar] [CrossRef]
- Scribailo, R.W.; Posluszny, U. The reproductive-biology of Hydrocharis-morsus-ranae. Can. J. Bot. 1984, 62, 2779–2787. [Google Scholar] [CrossRef]
- Scribailo, R.W.; Carey, K.; Posluszny, U. Isozyme variation and the reproductive-biology of Hydrocharis-morsus-ranae L (Hydrocharitaceae). Bot. J. Linn. Soc. 1984, 89, 305–312. [Google Scholar] [CrossRef]
- Garcia-Murillo, P. Hydrocharis laevigata in Europe. Plants 2023, 12, 701. [Google Scholar] [CrossRef] [PubMed]
- Xue, T.T.; Sun, Q.Y.; Tu, J.H.; Liu, J.; Cui, M.X. Germination and secondary dormancy of Hydrocharis dubia (blume) seed. Bangladesh J. Bot. 2023, 52, 429–435. [Google Scholar] [CrossRef]
- Ru, J.; Liu, M.; Cheng, X.Y.; Wang, C. The morphological study of the fruit, seed and seedling of Hydrocharis dubia (Hydrocharitaceae). Pak. J. Bot. 2015, 47, 1467–1472. [Google Scholar]
- Zhao, S.T.; Huang, W.M.; Jiang, H.S.; Sun, J.Y.; Yin, L.Y.; Li, W. Hydrocharis dubia seeds maintain high seed vigor in ambient wet storage condition through scavenging hydrogen peroxide by antioxidant systems. Aquat. Bot. 2017, 143, 18–24. [Google Scholar] [CrossRef]
- Koutsovoulou, K.; Daws, M.I.; Thanos, C.A. Campanulaceae: A family with small seeds that require light for germination. Ann. Bot. 2014, 113, 135–143. [Google Scholar] [CrossRef] [PubMed]
- Magnee, K.J.H.; Scholten, O.E.; Kodde, J.; Postma, J.; Gort, G.; van Bueren, E.T.L.; Groot, S.P.C. Higher seed maturity levels, darker pericarp, and smaller seed size relate to improved damping-off tolerance in spinach. Sci. Hortic. 2023, 321, 112219. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Kamiya, Y. Gibberellins and light-stimulated seed germination. J. Plant Growth Regul. 2001, 20, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Teubner, K.; Grossschartner, M.; Teubner, I.E. Response of Zooplankton to Restoration and Climate Warming in Alte Donau. In Successful Restoration and Sustainable Management-an Ecosystem Case Study of a Shallow Urban Lake; Springer: Berlin/Heidelberg, Germany, 2018; Volume 1010, pp. 163–212. [Google Scholar]
- Rosbakh, S.; Phartyal, S.S.; Poschlod, P. Seed germination traits shape community assembly along a hydroperiod gradient. Ann. Bot. 2020, 125, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Brenchley, J.L.; Probert, R.J. Seed germination responses to some environmental factors in the seagrass Zostera capricorni from eastern Australia. Aquat. Bot. 1998, 62, 177–188. [Google Scholar] [CrossRef]
- Rocha, M.D.D.; Neto, I.E.L. Internal phosphorus loading and its driving factors in the dry period of Brazilian semiarid reservoirs. J. Environ. Manag. 2022, 312, 114983. [Google Scholar] [CrossRef]
- Wu, J.; Cheng, S.P.; Liang, W.; He, F.; Wu, Z.B. Effects of sediment anoxia and light on turion germination and early growth of Potamogeton crispus. Hydrobiologia 2009, 628, 111–119. [Google Scholar] [CrossRef]
- Kato, R.; Kadono, Y. Seed germination traits of Trapella sinensis (Trapellaceae), an endangered aquatic plant in Japan: Conservation implications. Aquat. Bot. 2011, 95, 258–261. [Google Scholar] [CrossRef]
- Li, W. Environmental opportunities and constraints in the reproduction and dispersal of aquatic plants. Aquat. Bot. 2014, 118, 62–70. [Google Scholar] [CrossRef]
- Sridharan, S.; Saha, M.; Singh, L. Evidence of Soil Microplastics Inhibiting the Germination of Commercial Coriander Seeds Under Field Conditions. Water Air Soil Pollut. 2023, 234, 675. [Google Scholar] [CrossRef]
- Xiao, C.; Wang, X.F.; Xia, J.; Li, G.H. The effect of temperature, water level and burial depth on seed germination of Myriophyllum spicatum and Potamogeton malaianus. Aquat. Bot. 2010, 92, 28–32. [Google Scholar] [CrossRef]
- Boedeltje, G.; ter Heerdt, G.N.J.; Bakker, J.P. Applying the seedling-emergence method under waterlogged conditions to detect the seed bank of aquatic plants in submerged sediments. Aquat. Bot. 2002, 72, 121–128. [Google Scholar] [CrossRef]
- Pulido, C.; Sebesta, N.; Richards, J.H. Effects of salinity on sawgrass (Cladium jamaicense Crantz) seed germination. Aquat. Bot. 2020, 166, 103277. [Google Scholar] [CrossRef]
- Brock, T.C.M.; Mielo, H.; Oostermeijer, G. On the life-cycle and germination of hottonia-palustris l in a wetland forest. Aquat. Bot. 1989, 35, 153–166. [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M.; Chester, E.W. Seed germination ecology of the aquatic winter annual Hottonia inflata. Aquat. Bot. 1996, 54, 51–57. [Google Scholar] [CrossRef]
- Hartleb, C.F.; Madsen, J.D.; Boylen, C.W. Environmental-factors affecting seed-germination in Myriophyllum-spicatum L. Aquat. Bot. 1993, 45, 15–25. [Google Scholar] [CrossRef]
- Dugdale, T.M.; De Winton, M.D.; Clayton, J.S. Burial limits to the emergence of aquatic plant propagules. N. Z. J. Mar. Freshw. Res. 2001, 35, 147–153. [Google Scholar] [CrossRef][Green Version]
- Mai, K.; Williams, R.A. Response of Oak and Maple Seed Germination and Seedling Growth to Di ff erent Manganese Fertilizers in a Cultured Substratum. Forests 2019, 10, 547. [Google Scholar] [CrossRef]
- Li, J.L.; Yin, L.Y.; Chen, Y.; Gu, S.; Li, W. Preliminary Study on Clustering Effect on Seed Germination for Several Aquatic Plants. J. Wuhan Bot. Res. 2010, 28, 330–335. (In Chinese) [Google Scholar] [CrossRef]
- Baskin, C.C.; Baskin, J.M.; Chester, D.W.; Smith, M. Ethylene as a possible cue for seed germination of Schoenoplectus hallii (Cyperaceae), a rare summer annual of occasionally flooded sites. Am. J. Bot. 2003, 90, 620–627. [Google Scholar] [CrossRef] [PubMed]
- Azarkovich, M.I. Dehydrins in Orthodox and Recalcitrant Seeds. Russ. J. Plant Physiol. 2020, 67, 221–230. [Google Scholar] [CrossRef]
- Xia, K.; Zhu, Z.Q. Characterization of physiological traits during development of the recalcitrant seeds of Quercus serrata. Plant Biol. 2021, 23, 1000–1005. [Google Scholar] [CrossRef]
- Kauth, P.J.; Biber, P.D. Testa imposed dormancy in Vallisneria americana seeds from the Mississippi Gulf Coast. J. Torrey Bot. Soc. 2014, 141, 80–90. [Google Scholar] [CrossRef]
- Roberts, E.H. Predicting the storage life of seeds. Seed Sci. Technol. 1973, 143, 499–514. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Temperature | Relative Humidity | Storage Media | Seed Status | Storage Place | Storage Time |
|---|---|---|---|---|---|---|
| 4 °C-Dry | 4 °C | 55% | paper bag | dry | refrigerator | 2 years |
| 4 °C-Wet | 4 °C | 100% | water | wet | refrigerator | 2 years |
| AA-Dry | ambient air temperature a | 75% | paper bag | dry | laboratory room | 2 years |
| AA-Wet | ambient air temperature | 100% | water | wet | laboratory room | 2 years |
| AW-Wet | ambient water temperature b | 100% | wet sand | wet | pond | 2 years |
| Source | t-Value a or F-Value b | p |
|---|---|---|
| Light availability | 4.696 ** | 0.009 |
| Oxygen availability | −0.979 | 0.366 |
| Water level | 1.601 | 0.219 |
| Burial depth | 69.102 ** | <0.001 |
| Substratum | 0.984 | 0.428 |
| Clustering density | −25.298 ** | <0.001 |
| Storage Conditions | Germination (%) | Moisture Content (% FW) | Respiration Rate (μmol O2 h−1 mg−1 FW) |
|---|---|---|---|
| 4 °C-Dry | 64.0 ± 1.7 c | 13.80 ± 0.62 b | 13.55 ± 1.36 ab |
| 4 °C-Wet | 85.2 ± 5.1 b | 35.44 ± 0.50 a | 11.09 ± 0.51 ab |
| AA-Dry | 0 ± 0 d | 11.51 ± 0.57 b | 7.36 ± 2.25 b |
| AA-Wet | - | - | - |
| AW-Wet | 92.8 ± 4.3 a | 32.49 ± 1.13 a | 15.25 ± 2.10 a |
| Parameters | F-Value a | p |
|---|---|---|
| Germination | 263.499 ** | <0.001 |
| Moisture content | 201.162 ** | <0.001 |
| Respiration rate | 11.85 ** | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, S.; Jiang, H.; Liu, Y.; Xian, L.; Fu, W.; Yuan, S.; Yin, L.; Li, W. Germination Requirement and Suitable Storage Method of Hydrocharis dubia Seeds. Biology 2024, 13, 246. https://doi.org/10.3390/biology13040246
Zhao S, Jiang H, Liu Y, Xian L, Fu W, Yuan S, Yin L, Li W. Germination Requirement and Suitable Storage Method of Hydrocharis dubia Seeds. Biology. 2024; 13(4):246. https://doi.org/10.3390/biology13040246
Chicago/Turabian StyleZhao, Suting, Hongsheng Jiang, Yang Liu, Ling Xian, Wenlong Fu, Saibo Yuan, Liyan Yin, and Wei Li. 2024. "Germination Requirement and Suitable Storage Method of Hydrocharis dubia Seeds" Biology 13, no. 4: 246. https://doi.org/10.3390/biology13040246
APA StyleZhao, S., Jiang, H., Liu, Y., Xian, L., Fu, W., Yuan, S., Yin, L., & Li, W. (2024). Germination Requirement and Suitable Storage Method of Hydrocharis dubia Seeds. Biology, 13(4), 246. https://doi.org/10.3390/biology13040246