

Antifungal Activity of Thirty Essential Oils to Control Pathogenic Fungi of Postharvest Decay




Abstract
:1. Introduction
2. Results
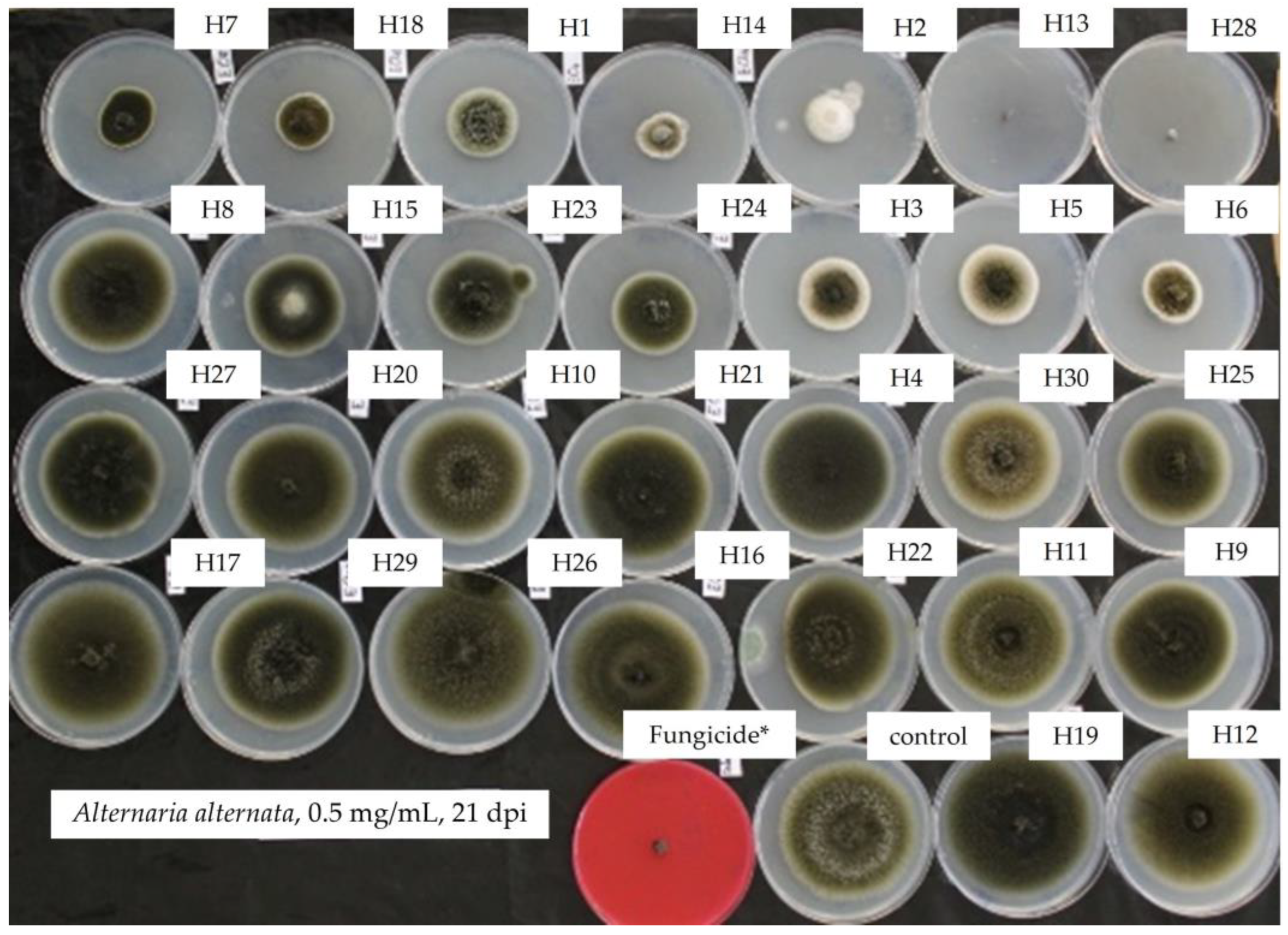
2.1. Response of Alternaria alternata towards 30 EOs
2.2. Response of Botrytis cinerea towards 30 EOs
2.3. Response of Penicillium italicum towards 30 EOs
2.4. Comparison between the Essential Oil Inhibitory Activity and between Fungi Tolerance
2.5. Minimum Inhibitory Concentration
3. Discussion
4. Materials and Methods
4.1. Origin of the Essential Oils
4.2. Fungal Species
4.3. In Vitro Antifungal Activities on Mycelial Growth
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO Fruit and Vegetables–Your Dietary Essentials. Available online: https://www.fao.org/documents/card/en/c/cb2395en (accessed on 16 September 2023).
- Moumni, M.; Romanazzi, G.; Najar, B.; Pistelli, L.; Ben Amara, H.; Mezrioui, K.; Karous, O.; Chaieb, I.; Allagui, M.B. Antifungal activity and chemical composition of seven essential oils to control the main seedborne fungi of cucurbits. Antibiotics 2021, 10, 104. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Khalek, H.H.; Hammad, A.A.; El-Kader, R.M.A.; Youssef, K.A.; Abdou, D.A. Combinational inhibitory action of essential oils and gamma irradiation for controlling Aspergillus flavus and Aspergillus parasiticus growth and their aflatoxins biosynthesis in vitro and in situ conditions. Food Sci. Technol. Int. 2022, 28, 703–715. [Google Scholar] [CrossRef]
- Zhou, X.; Zeng, M.; Huang, F.F.; Qin, G.; Song, Z.; Liu, F. The potential role of plant secondary metabolites on antifungal and immunomodulatory effect. Appl. Microbiol. Biotechnol. 2023, 107, 4471–4492. [Google Scholar] [CrossRef] [PubMed]
- Phuong, N.T.H.; Koga, A.; Nkede, F.N.; Tanaka, F.; Tanaka, F. Application of edible coatings composed of chitosan and tea seed oil for quality improvement of strawberries and visualization of internal structure changes using X-ray computed tomography. Prog. Org. Coat. 2023, 183, 107730. [Google Scholar] [CrossRef]
- Zhou, W.; Wang, Z.; Dong, L.; Wen, Q.; Huang, W.; Li, T.; YE, J.; Xu, L.A. Analysis on the character diversity of fruit and seed of Camellia chekiangoleosa. J. Nanjing Norm. Univ. (Nat. Sci. Ed.) 2021, 45, 51. [Google Scholar]
- Xiong, Y.; Li, S.; Warner, R.D.; Fang, Z. Effect of oregano essential oil and resveratrol nanoemulsion loaded pectin edible coating on the preservation of pork loin in modified atmosphere packaging. Food Control 2020, 114, 107226. [Google Scholar] [CrossRef]
- Basaglia, R.R.; Pizato, S.; Santiago, N.G.; de Almeida, M.M.M.; Pinedo, R.A.; Cortez-Vega, W.R. Effect of edible chitosan and cinnamon essential oil coatings on the shelf life of minimally processed pineapple (Smooth cayenne). Food Biosci. 2021, 41, 100966. [Google Scholar] [CrossRef]
- Wani, S.M.; Gull, A.; Ahad, T.; Malik, A.R.; Ganaie, T.A.; Masoodi, F.A.; Gani, A. Effect of gum Arabic, xanthan and carrageenan coatings containing antimicrobial agent on postharvest quality of strawberry: Assessing the physicochemical, enzyme activity and bioactive properties. Int. J. Biol. Macromol. 2021, 183, 2100–2108. [Google Scholar] [CrossRef]
- Lee, J.S.; Lee, E.S.; Han, J. Enhancement of the water-resistance properties of an edible film prepared from mung bean starch via the incorporation of sunflower seed oil. Sci. Rep. 2020, 10, 13622. [Google Scholar] [CrossRef]
- Das, S.; Vishakha, K.; Banerjee, S.; Mondal, S.; Ganguli, A. Sodium alginate-based edible coating containing nanoemulsion of Citrus sinensis essential oil eradicates planktonic and sessile cells of food-borne pathogens and increased quality attributes of tomatoes. Int. J. Biol. Macromol. 2020, 162, 1770–1779. [Google Scholar] [CrossRef]
- Hamedi, H.; Kargozari, M.; Shotorbani, P.M.; Mogadam, N.B.; Fahimdanesh, M. A novel bioactive edible coating based on sodium alginate and galbanum gum incorporated with essential oil of Ziziphora persica: The antioxidant and antimicrobial activity, and application in food model. Food Hydrocoll. 2017, 72, 35–46. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, C.; Liu, S.; Gao, J.; Cui, S.W.; Xia, W. Coating white shrimp (Litopenaeus vannamei) with edible fully deacetylated chitosan incorporated with clove essential oil and kojic acid improves preservation during cold storage. Int. J. Biol. Macromol. 2020, 162, 1276–1282. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, M.; Bhandari, B.; Bai, B. Nanoemulsion-based edible coatings loaded with fennel essential oil/cinnamaldehyde: Characterization, antimicrobial property and advantages in pork meat patties application. Food Control 2021, 127, 108151. [Google Scholar] [CrossRef]
- Sharma, A.; Gumber, K.; Gohain, A.; Bhatia, T.; Sohal, H.S.; Mutreja, V.; Bhardwaj, G. Chapter 3—Importance of essential oils and current trends in use of essential oils (aroma therapy, agrofood, and medicinal usage). In Essential Oils; Ahmad, G., Ansari, N.M.J., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 53–83. ISBN 9780323917407. [Google Scholar] [CrossRef]
- Khanam, M.; Dar, A.H.; Beg, F.; Khan, S.A.; Nayik, G.A.; Karabagias, I.K. Nutmeg essential oil. In Essential Oils; Academic Press: Cambridge, MA, USA, 2023; pp. 391–399. [Google Scholar]
- Oliveira Filho, J.G.D.; Duarte, L.G.; Silva, Y.B.; Milan, E.P.; Santos, H.V.; Moura, T.C.; Bandini, V.P.; Vitolano, L.E.S.; Nobre, J.J.; Moreira, C.T.; et al. Novel approach for improving papaya fruit storage with carnauba wax nanoemulsion in combination with Syzigium aromaticum and Mentha spicata essential oils. Coatings 2023, 13, 847. [Google Scholar] [CrossRef]
- Magri, A.; Curci, M.; Battaglia, V.; Fiorentino, A.; Petriccione, M. Essential oils in postharvest treatment against microbial spoilage of the rosaceae family fruits. AppliedChem 2023, 3, 196–216. [Google Scholar] [CrossRef]
- Rahman, M.M.; Wills, R.B.; Bowyer, M.C.; Golding, J.B.; Kirkman, T.; Pristijono, P. Potential control of postharvest fungal decay of citrus fruits by crude or photochemically changed essential oils—A review. Food Rev. Int. 2023, 1–18. [Google Scholar] [CrossRef]
- Fragkouli, R.; Antonopoulou, M.; Asimakis, E.; Spyrou, A.; Kosma, C.; Zotos, A.; Tsiamis, G.; Patakas, A.; Triantafyllidis, V. Mediterranean plants as potential source of biopesticides: An overview of current research and future trends. Metabolites 2023, 13, 967. [Google Scholar] [CrossRef]
- Javaid, A.; Ali, A.; Khan, I.H.; Ferdosi, M.F. Leaves of Chenopodium album as source of natural fungicides against Sclertium rolfsii. Arab. J. Chem. 2023, 16, 104677. [Google Scholar] [CrossRef]
- Assadpour, E.; Can Karaça, A.; Fasamanesh, M.; Mahdavi, S.A.; Shariat-Alavi, M.; Feng, J.; Kharazmi, M.S.; Rehman, A.; Jafari, S.M. Application of essential oils as natural biopesticides; recent advances. Crit. Rev. Food Sci. Nutr. 2023, 1–21. [Google Scholar] [CrossRef]
- Servili, A.; Feliziani, E.; Romanazzi, G. Exposure to volatiles of essential oils alone or under hypobaric treatment to control postharvest gray mold of table grapes. Postharvest Biol. Technol. 2017, 133, 36–40. [Google Scholar] [CrossRef]
- Mani-López, E.; Cortés-Zavaleta, O.; López-Malo, A. A review of the methods used to determine the target site or the mechanism of action of essential oils and their components against fungi. SN Appl. Sci. 2021, 3, 44. [Google Scholar] [CrossRef]
- Sukatta, U.; Haruthaithanasan, V.; Chantarapanont, W.; Dilokkunanant, U.; Suppakul, P. Antifungal activity of clove and cinnamon oil and their synergistic against postharvest decay fungi of grape in vitro. Kasetsart J. (Nat. Sci.) 2008, 42, 169–174. [Google Scholar]
- Parker, R.A.; Gabriel, K.T.; Graham, K.D.; Butts, B.K.; Cornelison, C.T. Antifungal activity of select essential oils against Candida auris and their interactions with antifungal drugs. Pathogens 2022, 11, 821. [Google Scholar] [CrossRef]
- Wang, S.Y.; Chen, P.F.; Chang, S.T. Antifungal activities of essential oils and their constituents from indigenous cinnamon (Cinnamomum osmophloeum) leaves against wood decay fungi. Bioresour. Technol. 2005, 96, 813–818. [Google Scholar] [CrossRef]
- Kishore, G.K.; Pande, S.; Harish, S. Evaluation of essential oils and their components for broad-spectrum antifungal activity and control of late leaf spot and crown rot diseases in peanut. Plant Dis. 2007, 91, 375–379. [Google Scholar] [CrossRef]
- Šernaitė, L.; Rasiukevičiūtė, N.; Valiuškaitė, A. The extracts of cinnamon and clove as potential biofungicides against strawberry grey mould. Plants 2020, 9, 613. [Google Scholar] [CrossRef] [PubMed]
- Shahina, Z.; Molaeitabari, A.; Sultana, T.; Dahms, T.E.S. Cinnamon leaf and clove essential oils are potent inhibitors of Candida albicans virulence traits. Microorganisms 2022, 10, 1989. [Google Scholar] [CrossRef]
- Al-Garadi, M.; Qaid, M.; Alqhtani, A.; Alhajj, M.; Al-abdullatif, A.; Al-Mufarrej, S. In vitro antimicrobial efficacy assessment of ethanolic and aqueous extracts of cinnamon (Cinnamomum verum) bark against selected microbes. Braz. J. Poult. Sci. 2023, 25, 1–15, eRBCA-2022-1682. [Google Scholar] [CrossRef]
- Singewar, K.; Fladung, M.; Robischon, M. Methyl salicylate as a signaling compound that contributes to forest ecosystem stability. Trees 2021, 35, 1755–1769. [Google Scholar] [CrossRef]
- Snoeren, T.A.L.; Mumm, R.; Poelman, E.H.; Yang, Y.; Pichersky, E.; Dicke, M. The herbivore-induced plant volatile methyl salicylate negatively affects attraction of the parasitoid Diadegma semiclausum. J. Chem. Ecol. 2010, 36, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Essien, E.E.; Newby, J.S.; Walker, T.M.; Setzer, W.N.; Ekundayo, O. Characterization and antimicrobial activity of volatile constituents from fresh fruits of Alchornea cordifolia and Canthium subcordatum. Medicines 2016, 3, 1. [Google Scholar] [CrossRef]
- Valverde, J.M.; Giménez, M.J.; Guillén, F.; Valero, D.; Martínez-Romero, D.; Serrano, M. Methyl salicylate treatments of sweet cherry trees increase antioxidant systems in fruit at harvest and during storage. Postharvest Biol. Technol. 2015, 109, 106–113. [Google Scholar] [CrossRef]
- Hao, Y.; Lin, Z.X.; Xiang, W.L.; Huang, M.; Tang, J.; Lu, Y.; Zhao, Q.H.; Zhang, Q.; Rao, Y.; Liu, L. Antifungal activity and mechanism of D-limonene against foodborne opportunistic pathogen Candida tropicalis. LWT-Food Sci. Technol. 2022, 159, 113144. [Google Scholar]
- Álvarez-García, S.; Moumni, M.; Romanazzi, G. Antifungal activity of volatile organic compounds from essential oils against the postharvest pathogens Botrytis cinerea, Monilinia fructicola, Monilinia fructigena, and Monilinia laxa. Front. Plant Sci. 2023, 14, 1274770. [Google Scholar] [CrossRef]
- Santoro, K.; Maghenzani, M.; Chiabrando, V.; Bosio, P.; Gullino, M.L.; Spadaro, D.; Giacalone, G. Thyme and savory essential oil vapor treatments control brown rot and improve the storage quality of peaches and nectarines, but could favor gray mold. Foods 2018, 7, 7. [Google Scholar] [CrossRef] [PubMed]
- Iacomi-Vasilescu, B.; Avenot, H.; Bataille-Simoneau, N.; Laurent, E.; Guénard, M.; Simoneau, P. In vitro fungicide sensitivity of Alternaria species pathogenic to crucifers and identification of Alternaria brassicicola field isolates highly resistant to both dicarboximides and phenylpyrroles. Crop Prot. 2004, 23, 481–488. [Google Scholar] [CrossRef]
- Avenot, H.F.; Michailides, T.J. Detection of isolates of Alternaria alternata with multiple-resistance to fludioxonil, cyprodinil, boscalid and pyraclostrobin in California pistachio orchards. Crop Prot. 2015, 78, 214–221. [Google Scholar] [CrossRef]
- Rosenzweig, N.; Hanson, L.E.; Mambetova, S.; Jiang, Q.W.; Guza, C.; Stewart, J.; Somohano, P. Fungicide sensitivity monitoring of Alternaria spp. causing leaf spot of sugarbeet (Beta vulgaris) in the Upper Great Lakes. Plant Dis. 2019, 103, 2263–2270. [Google Scholar] [CrossRef] [PubMed]
- Fagodiya, R.K.; Trivedi, A.; Fagodia, B.L. Impact of weather parameters on Alternaria leaf spot of soybean incited by Alternaria alternata. Sci. Rep. 2022, 12, 6131. [Google Scholar] [CrossRef] [PubMed]
- Oiki, S.; Yaguchi, T.; Urayama, S.I.; Hagiwara, D. Wide distribution of resistance to the fungicides fludioxonil and iprodione in Penicillium species. PLoS ONE 2022, 17, e0262521. [Google Scholar] [CrossRef]
- Moumni, M.; Allagui, M.B.; Mezrioui, K.; Ben Amara, H.; Romanazzi, G. Evaluation of seven essential oils as seed treatments against seedborne fungal pathogens of Cucurbita maxima. Molecules 2021, 26, 2354. [Google Scholar] [CrossRef] [PubMed]
- Bekker, T.F.; Kaiser, C.; vd Merwe, R.; Labuschagne, N. In-Vitro inhibition of mycelial growth of several phytopathogenic fungi by soluble potassium silicate. S. Afr. J. Plant Soil 2006, 23, 169–172. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Essential Oil | Inhibition of Mycelial Growth of A. alternata (%) at Different Essential Oil Concentrations (mg/mL) | ||||
|---|---|---|---|---|---|
| 0.05 | 0.1 | 0.2 | 0.3 | 0.4 | |
| Foeniculum vulgare (H2) | 3.8 ± 0.9 bc | 6.5 ± 1.5 bc | 9.4 ± 5.08 c | 17.6 ± 6.1 cd | 28.2 ± 8.8 b |
| Syzygium aromaticum (H13) | 18.7 ± 8.5 a | 17.8 ± 5.3 ab | 65.0 ± 2.8 a | 76.7 ± 4.4 b | 86.9 ± 6.7 a |
| Gautheria fragrantissima (H15) | 7.2 ± 5.2 ab | 2.9 ± 1.1 c | 27.5 ± 9.7 b | 26.4 ± 13.8 c | 28.4 ± 11.2 b |
| Cymbopogon nardus (H23) | −6 ± 0.3 c | −1.3 ± 3.7 c | 3.8 ± 1.48 d c | 5.8 ± 3.9 d | 21.4 ± 5.1 b |
| Pelargonium asperum (H24) | 1.3 ± 0.7 bc | 2.0 ± 1.7 c | 7.4 ± 2.7 c | 0.0 ± 0.6 d | 23.9 ± 21.4 b |
| Cinnamomum verrum (H28) | 10.3 ± 2.6 ab | 26.7 ± 13.5 a | 53.1 ± 7.4 a | 99.4 ± 0.4 a | 100.00 a |
| Essential Oil | Inhibition of Mycelial Growth of B. cinerea (%) at Increasing Essential Oil Concentrations (mg/mL) | ||||
|---|---|---|---|---|---|
| 0.05 | 0.1 | 0.2 | 0.3 | 0.4 | |
| Foeniculum vulgare (H2) | 0 ± 0 c | 31.9 ± 8.8 cd | 89.8 ± 0.7 ab | 84.4 ± 7.7 ab | 94.1 ± 1.5 ab |
| Syzygium aromaticum (H13) | 46.6 ± 7.7 a | 49.6 ± 1.3 bc | 63.1 ± 2.0 c | 73.5 ± 1.6 bc | 72.4 ± 0.5 c |
| Gautheria fragrantissima (H15) | 26.2 ± 24.9 abc | 58.1 ± 14.5 b | 88.7 ± 5.6 ab | 77.4 ± 2.7 bc | 90 ± 4.8 b |
| Cymbopogon nardus (H23) | 42.9 ± 4.2 ab | 85 ± 3.1 a | 89.8 ± 1.9 ab | 90.4 ± 0.9 ab | 99.1 ± 0.2 a |
| Pelargonium asperum (H24) | 12.3 ± 12.0 bc | 13.6 ± 5.4 d | 25.6 ± 6.3 d | 63.1 ± 14.5 c | 65.5 ± 3.6 c |
| Cinnamomum verrum (H28) | 12.3 ± 2.6 bc | 25.7 ± 0.7 d | 96.3 ± 2.1 a | 99.8 ± 0.1 a | 100 a |
| Essential Oil | Inhibition of Mycelial Growth of P. italicum (%) at Increasing Essential Oil Concentrations (mg/mL) | ||||
|---|---|---|---|---|---|
| 0.05 | 0.1 | 0.2 | 0.3 | 0.4 | |
| Syzygium aromaticum (H13) | 49.9 ± 15.5 a | 55.1 ± 14.0 a | 76.8 ± 3.3 a | 87.8 ± 2.3 a | 87.4 ± 3.3 b |
| Cinnamomum verrum (H28) | 14.0 ± 2.6 a | 10.4 ± 5.1 b | 30.3 ± 19.5 a | 76.1 ± 12.9 a | 100 a |
| Fungal Species | Code of EOs | Linear Regression | R² |
|---|---|---|---|
| Alternaria alternata | H2 | y = 0.0748x + 1.5905 | 0.96 |
| H13 | y = 0.1849x + 9.2307 | 0.88 | |
| H15 | y = 0.1199x − 3.0083 | 0.82 | |
| H23 | y = 0.0737x − 7.8006 | 0.85 | |
| H24 | y = 0.0607x − 3.8176 | 0.74 | |
| H28 | y = 0.2279x + 5.4806 | 0.90 | |
| Botrytis cinerea | H2 | y = 0.2143x + 9.6257 | 0.79 |
| H13 | y = 0.1268x + 27.743 | 0.72 | |
| H15 | y = 0.1751x + 22.99 | 0.76 | |
| H23 | y = 0.1585x + 37.839 | 0.62 | |
| H24 | y = 0.1133x + 66.675 | 0.27 | |
| H28 | y = 0.2816x + 12.828 | 0.77 | |
| Penicillium italicum | H13 | y = 0.1589x + 29.716 | 0.77 |
| H28 | y = 0.2258x − 2.5463 | 0.94 |
| Code | Plant Species | Vegetal Part of Oil Extraction | Major Chemical Components | Origin of Plant Product |
|---|---|---|---|---|
| H1 | Cymbopogon martinii | Aerial part of palmarosa | Geraniol, geranyl acetate | India |
| H2 | Foeniculum vulgare | Aerial part of sweet fennel | Trans-anethol, limonene | France |
| H3 | Laurus nobilis | Leaves of noble laurel | 1,8-cineole, terpenyl acetate | Balkans |
| H4 | Melaleuca alternifolia | Leaves of tea tree | Terpineol, alpha-terpinen | Australia |
| H5 | Salvia sclarea | flowering top of clary sage | Linalyl acetate, linalol | Bulgaria |
| H6 | Cananga odoranta | Flower of ylang ylang | Germacrene D, benzyle acetate and banzaoate, farnesene | Comores |
| H7 | Pogostemon cablin | Flowering top of patchouli | Patchoulol, alpha-patchoulene, alpha-bulnesene | Indonesia |
| H8 | Melaleuca quinquenervia | Leaves of niaouli | 1,8 cineole, α-pinene, viridiflorol | Madagascar |
| H9 | Boswellia carterii | Oleoresin | α-Pinene; β-Myrcene; β-Caryophyllene | Somalia |
| H10 | Lavandula angustifolia | Flowering top of fine lavender | Linalyle acetate, linalol, camphor | New Zealand |
| H11 | Citrus sinensis | Sweet orange zest | Limonene | Greece |
| H12 | Citrus paradisii | Grapefruit zest | Limonene | Argentina |
| H13 | Syzygium aromaticum | floral buds of cloves | Eugenol, eugenyl acetate | Madagascar |
| H14 | Mentha piperita | Aerial part of pepper mint | Menthol, menthone | France |
| H15 | Gautheria fragrantissima | Fragrant wintergreen leaves | Methyl aalicylate | Nepal |
| H16 | Artemisia dracunculus | Flowering top of tarragon | Estragole, chavicol | France |
| H17 | Citrus limon | Lemon zest | Limonene, limonum citrus | Italy |
| H18 | Daucus carota | Seeds of carrot | Carotol, alphapinene | France |
| H19 | Zingiber officinale | Rhizome ofginger | zingeberene, gingerol, bisabolene | China |
| H20 | Petroselinum crispum (Piper nigrum) | Fruit of black pepper | Caryophyllene, limonene | Madagascar |
| H21 | Citrus reticulata | Mandarin zest | Limonene, γ-Terpinene | Argentina |
| H22 | Citrus aurantium bergamia | Bergamot zest | Limonene, linalyl acetate, linalol | Italy |
| H23 | Cymbopogon nardus | Aerial part of Ceylon lemongrass | Citronellal, geraniol, citronellol, limonene, elemol | Ceylon |
| H24 | Pelargonium asperum | Geranium leaves | Citronellol, geraniol | Egypt |
| H25 | Juniperus communis | Branch of genevrier | α-Pinene, Sabinene | Bulgaria |
| H26 | Chamaemelum nobile | Flowers of chamomile | Isobutyl angelate, Isoamyl angelate, methylallyl angelate | Italy |
| H27 | Cedrus atlantica | Wood of Atlas cedar | Himachalene | Morocco |
| H28 | Cinnamomum verrum | Cinnamon bark | E-cinnamaldehyde, cinnamyl acetate | Madagascar |
| H29 | Cupressus sempervirens | Branch of cypress | Alpha pinene, Delta 3 carene | Morocco |
| H30 | Ocimum basilicum | Flowering top of basil | Estragol (Methylchavicol) | Italy |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Allagui, M.B.; Moumni, M.; Romanazzi, G. Antifungal Activity of Thirty Essential Oils to Control Pathogenic Fungi of Postharvest Decay. Antibiotics 2024, 13, 28. https://doi.org/10.3390/antibiotics13010028
Allagui MB, Moumni M, Romanazzi G. Antifungal Activity of Thirty Essential Oils to Control Pathogenic Fungi of Postharvest Decay. Antibiotics. 2024; 13(1):28. https://doi.org/10.3390/antibiotics13010028
Chicago/Turabian StyleAllagui, Mohamed Bechir, Marwa Moumni, and Gianfranco Romanazzi. 2024. "Antifungal Activity of Thirty Essential Oils to Control Pathogenic Fungi of Postharvest Decay" Antibiotics 13, no. 1: 28. https://doi.org/10.3390/antibiotics13010028
APA StyleAllagui, M. B., Moumni, M., & Romanazzi, G. (2024). Antifungal Activity of Thirty Essential Oils to Control Pathogenic Fungi of Postharvest Decay. Antibiotics, 13(1), 28. https://doi.org/10.3390/antibiotics13010028