
Reviewing the Benefits of Grazing/Browsing Semiarid Rangeland Feed Resources and the Transference of Bioactivity and Pro-Healthy Properties to Goat Milk and Cheese: Obesity, Insulin Resistance, Inflammation and Hepatic Steatosis Prevention



Abstract
:Simple Summary
Abstract
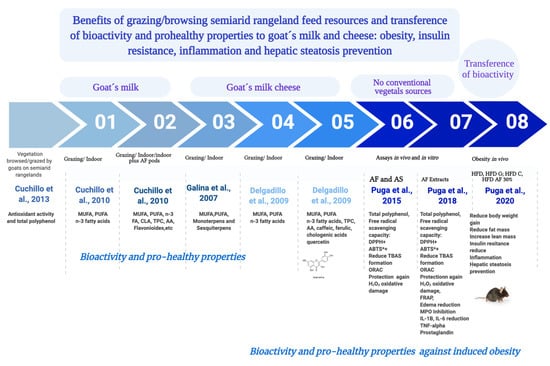
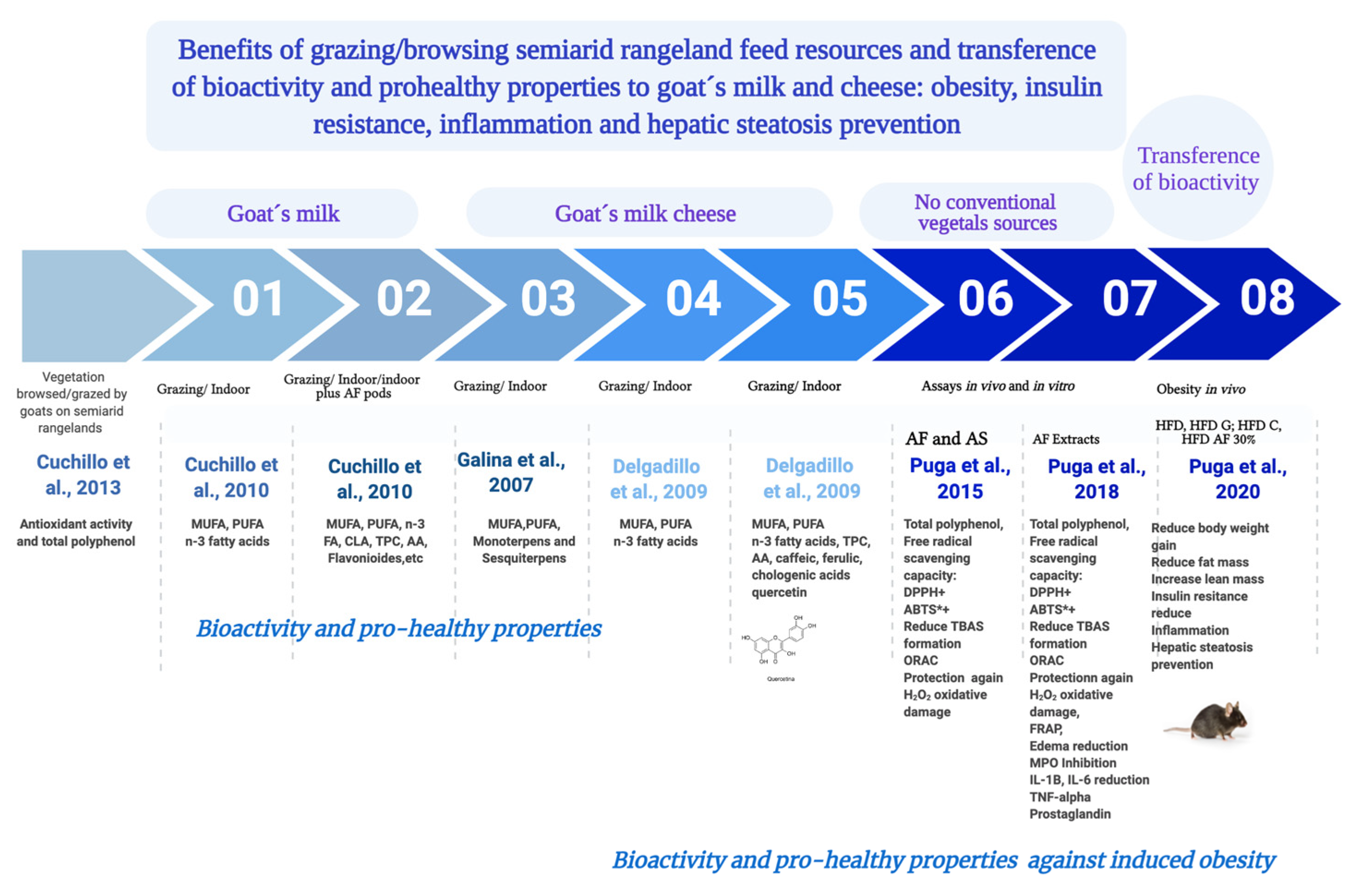
1. Introduction
2. Materials and Methods
2.1. Experiment 1
2.2. Experiment 2
2.3. Experiment 3
2.4. Experiment 4
2.5. Experiment 5
2.6. Experiment 6
2.7. Experiment 7
2.8. Experiment 8
2.9. Statistical Analyses
3. Results
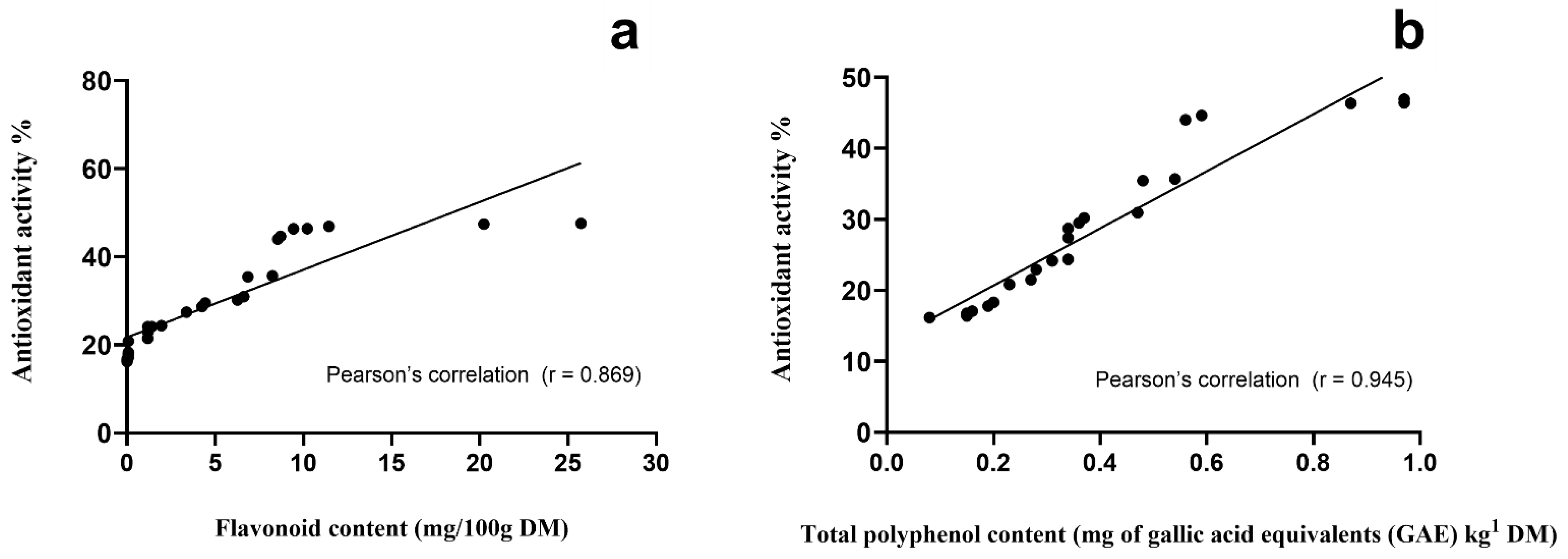
3.1. Antioxidant Activity and Total Polyphenol Content of Vegetation Species Browsed/Grazed by Goats on Semiarid Rangelands
3.2. Bioactive Compounds in Goas Milk and Cheese
3.2.1. Fatty Acids in Goat’s Milk and Cheese
3.2.2. Phenolic Content, Flavonoids, Terpenes, Hydroxycinnamic Acids, and Antioxidant Activity in Goat’s Milk and Cheese
3.3. In Vivo Prevention of Obesity, Hepatic Steatosis and Insulin Resistance
4. Discussion
4.1. Bioactive Compounds in Vegetation Species Browsed/Grazed by Goats on Semiarid Rangelands
4.2. Bioactive Compounds in Goat’s Milk and Cheese
4.3. Antioxidant and Anti-Inflammatory Activity of Acacia Pods
4.4. In Vivo Prevention of Obesity, Hepatic Steatosis and Insulin Resistance
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cabiddu, A.; Delgadillo-Puga, C.; Decandia, M.; Molle, G. Extensive ruminant production systems and milk quality with emphasis on unsaturated fatty acids, volatile compounds, antioxidant protection degree and phenol content. Animals 2019, 9, 771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrage, N.; Strodthoff, J.; Cuchillo, H.M.; Isselstein, J.; Kayser, M. Phytodiversity of temperate permanent grasslands: Ecosystem services for agriculture and livestock management for diversity conservation. Biodivers Conserv. 2011, 20, 3317–3339. [Google Scholar] [CrossRef] [Green Version]
- Claps, S.; Mecca, M.; Di Trana, A.; Sepe, L. Local small ruminant grazing in the monti foy area (Italy): The relationship between grassland biodiversity maintenance and added-value dairy products. Front. Vet. Sci. 2020, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Rudel, T.; Kwon, O.-J.; Paul, B.; Boval, M.; Rao, I.; Burbano, D.; McGroddy, M.; Lerner, A.; White, D.; Cuchillo, M.; et al. Do smallholder, mixed crop-livestock livelihoods encourage sustainable agricultural practices? A meta-analysis. Land 2016, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- van Vliet, S.; Provenza, F.D.; Kronberg, S.L. Health-promoting phytonutrients are higher in grass-fed meat and milk. Front. Sustain. Food Syst. 2021, 4, 1–20. [Google Scholar] [CrossRef]
- Galina, M.A.; Osnaya, F.; Cuchillo, H.M.; Haenlein, G.F.W. Cheese quality from milk of grazing or indoor fed Zebu cows and Alpine crossbred goats. Small Rumin. Res. 2007, 71, 264–272. [Google Scholar] [CrossRef]
- Cuchillo, H.M.; Puga, D.C.; Navarro, O.A.; Pérez-Gíl, R.F. Antioxidant activity, bioactive polyphenols in Mexican goats’ milk cheeses on summer grazing. J. Dairy Res. 2010, 77, 20–26. [Google Scholar] [CrossRef] [Green Version]
- Cuchillo, H.M.; Puga, D.C.; Wrage, N.; Pérez-Gíl, R.F. Feeding goats on scrubby Mexican rangeland and pasteurization: Influences on milk and artisan cheese quality. Trop. Anim. Health Prod. 2010, 42, 1127–1134. [Google Scholar] [CrossRef] [Green Version]
- Delgadillo, P.C.; Sánchez, M.B.; Nahed, T.J.; Cuchillo, H.M.; Díaz, M.M.; Solis, Z.R.; Reyes, H.A.; Castillo, D.R.M. Fatty acid content, health and risk indices, physicochemical composition, and somatic cell counts of milk from organic and conventional farming systems in tropical south-eastern Mexico. Trop. Anim. Health Prod. 2014, 46, 883–888. [Google Scholar] [CrossRef]
- Salami, S.A.; Luciano, G.; O’Grady, M.N.; Biondi, L.; Newbold, C.J.; Kerry, J.P.; Priolo, A. Sustainability of feeding plant by-products: A review of the implications for ruminant meat production. Anim. Feed Sci. Technol. 2019, 251, 37–55. [Google Scholar] [CrossRef]
- Ramírez-Bribiesca, J.E.; Hernández-Cruz, L.; Cruz-Monterrosa, R.G.; Guerrero-Legarreta, I.; Mota-Rojas, D.; Rayas-Amor, A.A.; Domínguez-Vara, I.A.; Avendaño-Reyes, L.; Corona-Gochi, L.; Cuchillo-Hilario, M. The effects of diets containing two corn stubble levels and three non-hydrogenated lipids sources on fattening performance, carcase, and meat quality of male hair-lambs. Ital. J. Anim. Sci. 2021, 20, 406–418. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Wu, J.H.Y. Flavonoids, dairy foods, and cardiovascular and metabolic health: A review of emerging biologic pathways. Circ. Res. 2018, 122, 369–384. [Google Scholar] [CrossRef]
- Delgadillo, P.C.; Galina, H.M.; Bonilla, C.A.; Cuchillo, H.M.; Montaño, B.S.; Castillo, D.R.; Villareal, E.; Pérez-Gil, R.F. Effect of feeding management on the nutritional composition of Mexican artisan soft cheese made with raw or pasteurized goats’ milk. Indian J. Anim. Sci. 2009, 79, 321–326. [Google Scholar]
- Delgadillo, P.C.; Cuchillo, H.M.; Pérez-Gil, R.F. Effect of feeding management and seasonal variation on fatty acid composition of Mexican soft raw goats’ milk cheese. Ital. J. Anim. Sci. 2009, 8, 402–404. [Google Scholar] [CrossRef] [Green Version]
- Delgadillo-Puga, C.; Cuchillo-Hilario, M.; León-Ortiz, L.; Ramírez-Rodríguez, A.; Cabiddu, A.; Navarro-Ocaña, A.; Morales-Romero, A.M.; Medina-Campos, O.N.; Pedraza-Chaverri, J. Goats’ feeding supplementation with Acacia farnesiana pods and their relationship with milk composition: Fatty acids, polyphenols, and antioxidant activity. Animals 2019, 9, 515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serra, V.; Salvatori, G.; Pastorelli, G. Dietary polyphenol supplementation in food producing animals: Effects on the quality of derived products. Animals 2021, 11, 401. [Google Scholar] [CrossRef]
- Innosa, D.; Ianni, A.; Faccia, M.; Martino, C.; Grotta, L.; Saletti, M.A.; Pomilio, F.; Martino, G. Physical, nutritional, and sensory properties of cheese obtained from goats fed a dietary supplementation with olive leaves. Animals 2020, 10, 2238. [Google Scholar] [CrossRef]
- Delgado-Pertíñez, M.; Martín-García, I.; Mena, Y.; Zarazaga, L.Á.; Guzmán, J.L. Supplementing the diet of dairy goats with dried orange pulp throughout lactation: Ii effect on milk fatty acids profile, phenolic compounds, fat-soluble vitamins and antioxidant capacity. Animals 2021, 11, 2421. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, D.; Sun, J.; Liu, X.; Jiang, L.; Guo, H.; Ren, F. Interaction of plant phenols with food macronutrients: Characterisation and nutritional-physiological consequences. Nutr. Res. Rev. 2014, 27, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Quideau, S.; Deffieux, D.; Douat-Casassus, C.; Pouységu, L. Plant polyphenols: Chemical properties, biological activities, and synthesis. Angew. Chem. Int. Ed. Engl. 2011, 50, 586–621. [Google Scholar] [CrossRef] [PubMed]
- Santurino, C.; Calvo, M.V.; Gómez-Candela, C.; Fontecha, J. Characterization of naturally goat cheese enriched in conjugated linoleic acid and omega-3 fatty acids for human clinical trial in overweight and obese subjects. PharmaNutrition 2017, 5, 8–17. [Google Scholar] [CrossRef]
- Fontecha, J.; Calvo, M.V.; Juarez, M.; Gil, A.; Martínez-Vizcaino, V. Milk and dairy product consumption and cardiovascular diseases: An overview of systematic reviews and meta-analyses. Adv. Nutr. 2019, 10, S164–S189. [Google Scholar] [CrossRef] [PubMed]
- Provenza, F.D.; Kronberg, S.L.; Gregorini, P. Is grassfed meat and dairy better for human and environmental health? Front. Nutr. 2019, 6. [Google Scholar] [CrossRef] [PubMed]
- Santurino, C.; López-Plaza, B.; Fontecha, J.; Calvo, M.V.; Bermejo, L.M.; Gómez-Andrés, D.; Gómez-Candela, C. Consumption of goat cheese naturally rich in omega-3 and conjugated linoleic acid improves the cardiovascular and inflammatory biomarkers of overweight and obese subjects: A randomized controlled trial. Nutrients 2020, 12, 1315. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.; Bronkema, S.M.; Yakah, W.; Rowntree, J.E.; Bitler, C.A.; Fenton, J.I. Seasonal differences exist in the polyunsaturated fatty acid, mineral and antioxidant content of U.S. grass-finished beef. PLoS ONE 2020, 15, e0229340. [Google Scholar] [CrossRef] [Green Version]
- Smith, J.; Sones, K.; Grace, D.; MacMillan, S.; Tarawali, S.; Herrero, M. Beyond milk, meat, and eggs: Role of livestock in food and nutrition security. Anim. Front. 2013, 3, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Röös, E.; Patel, M.; Spångberg, J.; Carlsson, G.; Rydhmer, L. Limiting livestock production to pasture and by-products in a search for sustainable diets. Food Policy 2016, 58, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Galina, M.A.; Puga, D.C.; Hernández, A.; Haenlein, G.F.W. Biodiverse and biosustainable production system with goats in Mexico: Importance of a forage bank. Small Rumin. Res. 1998, 27, 19–23. [Google Scholar] [CrossRef]
- Cuchillo, H.M.; Puga, D.C.; Wrage-Mönning, N.; Espinosa, M.J.G.; Montaño, B.S.; Navarro-Ocaña, A.; Ledesma, J.A.; Diaz, M.M.; Pérez-Gil, R.F. Chemical composition, antioxidant activity and bioactive compounds of vegetation species ingested by goats on semiarid rangelands. J. Anim. Feed Sci. 2013, 22, 106–115. [Google Scholar] [CrossRef]
- Puga, D.C.; Cuchillo, H.M.; Navarro, O.A.; Medina-Campos, O.N.; Nieto, C.A.; Lopez, T.Z.G.; Díaz, M.M.; Álvarez, I.M.A.; Cruz, Y.R.M. Phenolic compounds in organic and aqueous extracts from Acacia farnesiana pods analyzed by ULPS-ESI-Q-oa/TOF-MS. In vitro antioxidant activity and anti-inflammatory response in CD-1 mice. Molecules 2018, 23, 2386. [Google Scholar] [CrossRef] [Green Version]
- Delgadillo, P.C.; Cuchillo, H.M.; Espinosa, M.J.G.; Medina, C.O.; Molina, J.E.; Díaz, M.M.; Álvarez, I.M.A.; Ledesma, S.J.A.; Pedraza-Chaverri, J. Antioxidant activity and protection against oxidative-induced damage of Acacia shaffneri and Acacia farnesiana pods extracts: In vitro and in vivo assays. BMC Complement Altern. Med. 2015, 15, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Delgadillo-Puga, C.; Noriega, L.G.; Morales-Romero, A.M.; Nieto-Camacho, A.; Granados-Portillo, O.; Rodríguez-López, L.A.; Alemán, G.; Furuzawa-Carballeda, J.; Tovar, A.R.; Cisneros-Zevallos, L.; et al. Goat’s milk intake prevents obesity, hepatic steatosis and insulin resistance in mice fed a high-fat diet by reducing inflammatory markers and increasing energy expenditure and mitochondrial content in skeletal muscle. Int. J. Mol. Sci. 2020, 21, 5530. [Google Scholar] [CrossRef] [PubMed]
- Morales, A.R.; Galina, M.A.; Jimenez, S.; Haenlein, G.F.W. Improvement of biosustainability of a goat feeding system with key supplementation. Small Rumin. Res. 2000, 35, 97–105. [Google Scholar] [CrossRef]
- Nahed Toral, J.; López Tecpoyotl, Z.G.; Aguilar Jiménez, J.R.; Grande Cano, D.; Delgadillo Puga, C. Compliance of goat farming under extensive grazing with the organic standards and its contribution to sustainability in Puebla, Mexico. Sustainability 2021, 13, 6293. [Google Scholar] [CrossRef]
- Allen, V.G.; Batello, C.; Berretta, E.J.; Hodgson, J.; Kothmann, M.; Li, X.; McIvor, J.; Milne, J.; Morris, C.; Peeters, A.; et al. An international terminology for grazing lands and grazing animals. Grass Forage Sci. 2011, 66, 2–28. [Google Scholar] [CrossRef]
- Martirosyan, D.; Kanya, H.; Nadalet, C. Can functional foods reduce the risk of disease? Advancement of functional food definition and steps to create functional food products. Funct. Food Health Dis. 2021, 11, 213–221. [Google Scholar] [CrossRef]
- Sharma, O.P.; Bhat, T.K. DPPH antioxidant assay revisited. Food Chem. 2009, 113, 1202–1205. [Google Scholar] [CrossRef]
- Taga, M.; Miller, E.; Pratt, D. Chia seeds as a source of natural lipid antioxidants. J. Am. Oil Chem. Soc. 1984, 61, 928–931. [Google Scholar] [CrossRef]
- Folch, J.M.; Less, M.; Sloane-Stanley, G. A simple method of the isolation and purification of total lipids. J. Biol. Chem. 1957, 226, 497–504. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 23rd ed.; Association of Official Analytical Chemists: Washington, DC, USA, 2003. [Google Scholar]
- Chen, S.; Bobe, G.; Zimmerman, S.; Hammond, E.G.; Luhman, C.M.; Boylston, T.D.; Freeman, A.E.; Beitz, D.C. Physical and sensory properties of dairy products from cows with various milk fatty acid compositions. J. Agric. Food Chem. 2004, 52, 3422–3428. [Google Scholar] [CrossRef]
- Koren, E.; Kohen, R.; Ginsburg, I. Polyphenols enhance total oxidant-scavenging capacities of human blood by binding to red blood cells. Exp. Biol. Med. 2010, 235, 689–699. [Google Scholar] [CrossRef]
- Hatano, T.; Kagawa, H.; Yasuhara, T.; Okuda, T. Two new flavonoids and other constituents in liquorice root: Their relative astringency and radical scavenging effects. Chem. Pharmaceut. Bull. 1988, 36, 2090–2097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- von Gadow, A.; Joubert, E.; Hansmann, C.F. Comparison of the antioxidant activity of aspalathin with that of other plant phenols of rooibos tea (Aspalathus linearis), alpha-tocopherol, BHT, and BHA. J. Agric. Food Chem. 1997, 45, 632–638. [Google Scholar] [CrossRef]
- Tsuda, T.; Watanabe, M.; Ohshima, K.; Norinobu, S.; Choi, S.-W.; Kawakishi, S.; Osawa, T. Antioxidative Activity of the anthocyanin pigments cyanidin 3-O-.beta.-D-glucoside and cyanidin. J. Agric. Food Chem. 1994, 42, 2407–2410. [Google Scholar] [CrossRef]
- Hernández-Fonseca, K.; Cárdenas-Rodríguez, N.; Pedraza-Chaverri, J.; Massieu, L. Calcium-dependent production of reactive oxygen species is involved in neuronal damage induced during glycolysis inhibition in cultured hippocampal neurons. J. Neurosci. Res. 2008, 86, 1768–1780. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ou, B.; Hampsch-Woodill, M.; Flanagan, J.A.; Prior, R.L. High-throughput assay of oxygen radical absorbance capacity (ORAC) using a multichannel liquid handling system coupled with a microplate fluorescence reader in 96-well format. J. Agric. Food Chem. 2002, 50, 4437–4444. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Del-Ángel, M.; Nieto, A.; Ramírez-Apan, T.; Delgado, G. Anti-inflammatory effect of natural and semi-synthetic phthalides. Eur. J. Pharmacol. 2015, 752, 40–48. [Google Scholar] [CrossRef]
- Delgadillo, P.C.; Castillo, D.R.M.; Cuchillo, H.M.; Díaz, M.M.; Pérez-Gíl, R.F.; Montaño, B.S. Radical scavenging activity and health and risk fatty acid indices of soft goats’ milk cheeses. Arch. Latinoam. Prod. Anim. 2015, 23, 21–26. [Google Scholar]
- SAS. Statistical Analysis System. User’s Guide, 1st ed.; SAS. Institute Inc.: Cary, NC, USA, 2003; p. 120. [Google Scholar]
- IBM Corp. Released 2010. IBM SPSS Statistics for Windows, Version 19.0; IBM Corp: Armonk, NY, USA, 2010. [Google Scholar]
- Cuchillo, H.M.; Wrage-Mönnig, N.; Isselstein, J. Behavioral patterns of (co-)grazing cattle and sheep on swards differing in plant diversity. Appl. Anim. Behav. Sci. 2017, 191, 17–23. [Google Scholar] [CrossRef]
- Cuchillo, H.M.; Wrage-Mönnig, N.; Isselstein, J. Forage selectivity by cattle and sheep co-grazing swards differing in plant species diversity. Grass Forage Sci. 2018, 73, 320–329. [Google Scholar] [CrossRef]
- Juárez-Silva, M.E.; Cuchillo-Hilario, M.; Villarreal-Delgado, E. Dietary supplementation of inulin or flavomycin and type of cut of meat: Effects on fatty acid profile, sensorial characteristics and consumer preference of rabbits meat. Rev. Mex Cienc. Pecu. 2019, 10, 552–570. [Google Scholar] [CrossRef]
- Simopoulos, A. An increase in the omega-6/omega-3 fatty acid ratio increases the risk for obesity. Nutrients 2016, 8, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Intorre, F.; Foddai, M.S.; Azzini, E.; Martin, B.; Montel, M.-C.; Catasta, G.; Toti, E.; Finotti, E.; Palomba, L.; Venneria, E.; et al. Differential effect of cheese fatty acid composition on blood lipid profile and redox status in normolipidemic volunteers: A pilot study. Int. J. Food Nutr. Sci. 2011, 62, 660–669. [Google Scholar] [CrossRef]
- Jakobsen, M.U.; Trolle, E.; Outzen, M.; Mejborn, H.; Grønberg, M.G.; Lyndgaard, C.B.; Stockmarr, A.; Venø, S.K.; Bysted, A. Intake of dairy products and associations with major atherosclerotic cardiovascular diseases: A systematic review and meta-analysis of cohort studies. Sci. Rep. 2021, 11, 1303. [Google Scholar] [CrossRef]
- Palacios-González, B.; Vargas-Castillo, A.; Velázquez-Villegas, L.A.; Vasquez-Reyes, S.; López, P.; Noriega, L.G.; Aleman, G.; Tovar-Palacio, C.; Torre-Villalvazo, I.; Yang, L.J.; et al. Genistein increases the thermogenic program of subcutaneous WAT and increases energy expenditure in mice. J. Nutr. Biochem. 2019, 68, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Avior, Y.; Bomze, D.; Ramon, O.; Nahmias, Y. Flavonoids as dietary regulators of nuclear receptor activity. Food Funct. 2013, 4, 831–844. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
| Specie | Portion | Antioxidant Activity (AA, %) | Total Polyphenol Content (TPC) (mg of Gallic Acid Equivalents/kg DM) |
|---|---|---|---|
| Aristida adscensionis | Complete | 24.34 g | 160 l,m |
| Acacia schaffneri | Complete 1 | 47.38 a | 2730 b |
| Bouteloua curtipendula | Complete | 35.42 d | 314 g,h,i |
| Bouteloua repens | Complete | 30.92 b,c | 364 g |
| Chloris virgata | Complete | 27.39 i | 474 f |
| Jatropha dioica | Complete | 24.13 g | 189l k |
| Leptochloa dubia | Complete | 21.48 h | 146 l,m,n |
| Mimosa biuncifera | Complete | 28.70 e,f | 341 g,h |
| Rhynchelytrum roseum | Complete | 44.63 b,c | 23 i,j,k |
| Urochloa fasciculata | Complete | 30.19 e | 966 c |
| Mean value | 31.46 A * | 592 A * | |
| Acacia farnesiana | Fruits | 47.59 a | 38170 a |
| Opuntia amyctaea | Fruits | 20.83 h | 587 e |
| Opuntia hytiacantha | Fruits | 18.30 i | 343 g,h |
| Prosopis laevigata | Fruits | 22.92 g,h | 314 g,h |
| Mean value | 27.41 B * | 9854 B ** | |
| Celtis pallida | Leaves | 29.49 e,f | 280 h,i,j |
| Prosopis laevigata | Leaves | 35.68 d | 968 c |
| Verbesina serrata | Leaves | 17.04 i | 272 i,j |
| Mean value | 27.40 B * | 507 C ** | |
| Celtis pallida | Stems | 44.02 c | 370 g |
| Verbesina serrata | Stems | 17.78 i | 874 d |
| Zalazania augusta | Stems | 46.41 a,b | 480 f |
| Mean value | 36.07 A * | 575 D * | |
| Opuntia affasiacantha | Cladodes | 24.13 g | 202 k |
| Opuntia hytiacantha | Cladodes | 46.92 a | 77 n |
| Opuntia robusta | Cladodes | 16.14 i | 537 e,f |
| Opuntia streptacantha | Cladodes | 16.39 i | 147 l,m,n |
| Opuntia tomentosa | Cladodes | 16.77 i | 335 g,h,i |
| Mean value | 24.07 B,C ** | 259 E * |
| Animal Food Delivery | Milk | Cheese | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Experiment 1 | Experiment 2 | Experiment 3 | Experiment 4 | Experiment 5 | |||||||
| Geographical coordinates | 20°35′ Latitude North and 100°18′ Longitude West | 20°31′ Latitude North and 100°48′ Longitude West | 20°35′ Latitude North and 100°18′ Longitude West | ||||||||
| Dominant vegetation | Shrubby rangeland | Shrubby rangeland | Shrubby rangeland | Shrubby rangeland | Shrubby rangeland | ||||||
| Altitude a.s.l. (m) | 1950 | 1767 | 1950 | 1950 | 1950 | ||||||
| Treatments | Grazing | Indoor | Grazing | Indoor diet plus pods of A. farnesiana | Indoor | Grazing | Indoor | Grazing | Indoor | Grazing | Indoor |
| Number of animals | 10 | 10 | 10 | 10 | 10 | 60 | 60 | 40 | 40 | 10 | 10 |
| Goat breed | French Alpine | French Alpine | French Alpine | French Alpine | French Alpine | Crossbred French Alpine with Saanen and Toggenburg | Crossbred French Alpine with Saanen and Toggenburg | French Alpine | French Alpine | French Alpine | French Alpine |
| Analysis | |||||||||||
| Fatty acids | Gas chromatography | Gas chromatography | Gas chromatography | Gas chromatography | Gas chromatography | ||||||
| Monoterpenes | Mass spectrometry | ||||||||||
| Sesquiterpenes | Mass spectrometry | ||||||||||
| Total polyphenols | Folin–Ciocalteu colorimetric method | Folin–Ciocalteu colorimetric method | |||||||||
| Antioxidant activity | DPPH• scavenging activity | DPPH• scavenging activity | |||||||||
| Other bioactive compounds | High-performance liquid chromatography (HPLC) | High-performance liquid chromatography (HPLC) | |||||||||
| Compounds | % of total fatty acids | ||||||||||
| PUFA | 4.73 | 3.44 | 4.05 | 4.74 | 5.64 | 6.07 | 5.24 | 3.90 | 5.05 | 4.80 | 5.42 |
| MUFA | 25.21 | 19.85 | 26.22 | 25.85 | 31.62 | 23.49 | 23.83 | 22.03 | 22.08 | 25.29 | 23.88 |
| SFA | 64.35 | 70.29 | 69.73 | 69.42 | 62.74 | 68.88 | 69.38 | 57.30 | 62.21 | 66.29 | 65.62 |
| n-3 | 0.94 | 0.72 | 0.87 | 0.67 | 0.96 | 1.27 | 1.21 | 1.03 | 1.0 | 0.34 | 0.33 |
| n-3:n-6 ratio | 0.35 | 0.35 | 0.30 | 0.21 | 0.17 | 0.60 | 0.69 | 0.36 | 0.26 | 0.36 | 0.29 |
| CLA (conjugate linoleic acid) | - | - | 0.29 | 0.23 | 0.20 | - | - | - | - | - | - |
| Monoterpenes (ng/kg of cheese) | - | - | - | - | - | 460 | 221 | ||||
| Sesquiterpenes (ng/kg of cheese) | - | - | - | - | - | 850 | 415 | - | - | - | - |
| Total polyphenol content (mg of gallic acid equivalents/L of milk or kg of cheese) | - | - | 231.6 | 159.4 | 305.5 | - | - | - | - | 300 | 60 |
| Antioxidant activity (%) | - | - | 42.1 | 30.8 | 27.7 | - | - | - | - | 24.1 | 15.2 |
| Other bioactive compounds | - | - | mg/L | mg/kg | |||||||
| Chlorogenic acid | - | - | 17.64 | 12.04 | - | - | - | - | 119 | 53 | |
| Ferulic acid | - | - | 6.23 | 0.0 | 3.19 | - | - | - | - | 0.0 | 165 |
| Gallic acid | - | - | 1.43 | 2.94 | - | - | - | - | nd | nd | |
| Catechin | - | - | 5.27 | 1.61 | - | - | - | - | 0.23 | 0.16 | |
| Quercetin | - | - | nd | nd | - | - | - | - | 4.2 | nd | |
| Caffeic acid | - | - | nd | nd | - | - | - | - | 16 | nd | |
| Experiment 6 | Experiment 7 | |||||||
|---|---|---|---|---|---|---|---|---|
| In Vitro | ||||||||
| Variables | AS | AF | Acacia farnesiana Pods Extracts | |||||
| CE | HE | KE | ME | MEAE | AE | |||
| Total polyphenol (TP) mg of equivalent of gallic acid/g of extract | 213 a | 76 b | TPC, mg of equivalent of gallic acid/100 g pods, dry matter | |||||
| 506 c | 620 a | 594 b | 378 d | 399 d | 565 b | |||
| Free radical scavenging capacity % (DPPH+) | 95 a | 95 a | 90 a | 80 b | 79 b | 82 b | 80 b | 80 b |
| Free radical scavenging capacity % (ABTS•+) | 10 a | 10 a | - | - | - | - | - | - |
| Inhibition of TBARS formation, % | 66 a | 66 a | IC50 on TBARS ug/mL | |||||
| 9 b | 12 a | 4.0 c | 4.5 c | 4.5 c | 17 a | |||
| Protection against H2O2 oxidative-induced damage †; (%) | ++ | +++ | 37 a | 25 b | 36 a | 18 c | 23 b | 18 c |
| Oxygen radical absorbance capacity, (ORAC) § | - | - | 150 c | 100 d | 450 b | 450 b | 500 a | 50 e |
| Ferric-reducing antioxidant power (FRAP) ¥ | - | - | 1.5 a | 1.4 a | 1.5 a | 2.0 a | 1.7 a | 1.4 a |
| In Vivo | ||||||||
| Free radical scavenging capacity, % (DPPH+) | 29 | 30 | - | - | - | - | - | - |
| Oxygen radical absorbance capacity (ORAC)≠ | 2207 | 1561 | - | - | - | - | - | - |
| Edema (mg) | - | - | 12 a | 13 a | 13 a | 11 a | 14 a | 14 a |
| MPO inhibition % | - | - | 34.9 b | 28.5 b | 65.9 a | 31.0 b | 73.3 a | 37.1 b |
| Interleukins IL-1β (pg/biopsy) Š | - | - | 52 b | 60 a | 55 b | 52 b | 50 b | 52 b |
| Interleukins IL-6 (pg/biopsy) Š | - | - | 800 b | 1200 a | 780 b | 1100 a | 1200 a | 1100 a |
| TNF-α Š | - | - | 15 b | 12 b | 17 a | 13 b | 22 a | 11 b |
| Ear thickness (µm) | - | - | 400 b | 420 b | 580 a | 580 a | 300 c | 400 b |
| Prostaglandin (µg/mL) | - | - | 1200 b | 900 d | 3000 c | 16,000 a | 11,000 b | 12,000 b |
| Experiment 8 | |||||
|---|---|---|---|---|---|
| Variables | Control | HFD | CD | G | CD + AF |
| Energy intake and body changes | |||||
| Final body weight (g) | 30 b | 42 a | 29.2 b | 34.5 b | 33.8 b |
| Body weight gain (g) | 9.5 c | 21. 6 a | 9.6 c | 16.0 a,b | 11.8 b,c |
| Food intake (g/day) | 3.2 a,b | 3 d | 3.7 c | 4.4 a | 3.9 b |
| Energy intake (kcal/day) | 16.1 c | 16 c | 18.5 b | 21.7 a | 18.9 b |
| Mice body composition Fat mass (%) | 20 b | 35 a | 10.5 c | 19.5 b,c | 19.1 b |
| Mice body composition Lean mass (%) | 75 b | 62 c | 86.0 a | 78.5 a,b | 77.7 d |
| Serum and insulin resistance related-parameters | |||||
| Serum glucose (mg/dL) | 200 b | 310 a | 228 b | 263 a,b | 242 a,b |
| Serum insulin (ng/mL)+ | 1 a.b | 2.8 a | 0.63 b | 0.65 b | 0.70 b |
| ipGTT AUC | 25,000 b | 45,000 a | 19,584 b | 28,415 b | 22,689 b |
| ipITT AUC | 13,240 a,b | 20,000 a | 14,599 a,b | 13,240 a,b | 1741 b |
| Pancreatic islets size (µm2) | 9800b | 20,000 a | 8508 b | 10,110 a,b | 6683 b |
| Energy expenditure and thermogenic changes | |||||
| Average VO2 (oxygen consumption; mL/kg/h) feeding | 5700 b | 5450 c | 6035.1 a | 6059 a | 5989 a |
| Lipid content BODIPY staining in skeletal muscle (relative units) | 50 b | 150 a | 38.5 b | 24.33 b | 16.57 b |
| Mitochondrial activity SDH staining in skeletal muscle (relative units) | 1900 b | 900 c | 2237 a,b | 2612 a | 3075.0 a |
| UCP-1/GAPDH immunoblotting BAT (relative units) | 1 c | 1.5 b | 1.5 b | 2.0 a | 2.0 a |
| Hepatic energy improvement and anti-inflammatory prevention | |||||
| Lipid content ORO staining in hepatic tissue (relative units) | 105.8 b | 300 a | 105 b | 93.48 b | 104.4 b |
| p-AMPK/AMPK ratio in hepatic tissue (relative units) by immunoblotting | 1 c | 1.0 c | 1.41 b,c | 1.82 b | 2.59 a |
| p-JNK/JNK ration in hepatic tissue (relative units) by immunoblotting | 1 a | 3.00 a | 0.85 a | 0.37 b | 0.56 a |
| (EPA+DHA)/AA ratio in liver (mg/g of liver) | 0.2 c | 0.2 c | 1.39 b,c | 2.07 a,b | 0.82 b,c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgadillo-Puga, C.; Cuchillo-Hilario, M. Reviewing the Benefits of Grazing/Browsing Semiarid Rangeland Feed Resources and the Transference of Bioactivity and Pro-Healthy Properties to Goat Milk and Cheese: Obesity, Insulin Resistance, Inflammation and Hepatic Steatosis Prevention. Animals 2021, 11, 2942. https://doi.org/10.3390/ani11102942
Delgadillo-Puga C, Cuchillo-Hilario M. Reviewing the Benefits of Grazing/Browsing Semiarid Rangeland Feed Resources and the Transference of Bioactivity and Pro-Healthy Properties to Goat Milk and Cheese: Obesity, Insulin Resistance, Inflammation and Hepatic Steatosis Prevention. Animals. 2021; 11(10):2942. https://doi.org/10.3390/ani11102942
Chicago/Turabian StyleDelgadillo-Puga, Claudia, and Mario Cuchillo-Hilario. 2021. "Reviewing the Benefits of Grazing/Browsing Semiarid Rangeland Feed Resources and the Transference of Bioactivity and Pro-Healthy Properties to Goat Milk and Cheese: Obesity, Insulin Resistance, Inflammation and Hepatic Steatosis Prevention" Animals 11, no. 10: 2942. https://doi.org/10.3390/ani11102942
APA StyleDelgadillo-Puga, C., & Cuchillo-Hilario, M. (2021). Reviewing the Benefits of Grazing/Browsing Semiarid Rangeland Feed Resources and the Transference of Bioactivity and Pro-Healthy Properties to Goat Milk and Cheese: Obesity, Insulin Resistance, Inflammation and Hepatic Steatosis Prevention. Animals, 11(10), 2942. https://doi.org/10.3390/ani11102942