Impact of Fermentable Protein, by Feeding High Protein Diets, on Microbial Composition, Microbial Catabolic Activity, Gut Health and beyond in Pigs

 , ,
, ,
Abstract
1. Introduction
2. Changes in Microbial Composition
3. Impact on Microbial Catabolic Activity
3.1. Ammonia
3.2. SCFA/BCFA
3.3. Biogenic Amines
3.4. Indolic and Phenolic Compounds
3.5. Other Metabolites
3.6. Overall Impact on Microbial Catabolic Activity
4. Impact on the Gut and Host Health
4.1. Intestinal Morphology
4.2. Intestinal Barrier Function
4.3. Diarrhoea Incidence and Growth Performance
5. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Davila, A.M.; Blachier, F.; Gotteland, M.; Andriamihaja, M.; Benetti, P.H.; Sanz, Y.; Tome, D. Intestinal luminal nitrogen metabolism: Role of the gut microbiota and consequences for the host. Pharmacol. Res. 2013, 68, 95–107. [Google Scholar] [CrossRef]
- Smith, E.A.; Macfarlane, G.T. Enumeration of amino acid fermenting bacteria in the human large intestine: Effects of pH and starch on peptide metabolism and dissimilation of amino acids. FEMS Microbiol. Ecol. 1998, 25, 355–368. [Google Scholar] [CrossRef]
- Oliphant, K.; Allen-Vercoe, E. Macronutrient metabolism by the human gut microbiome: Major fermentation by-products and their impact on host health. Microbiome 2019, 7, 91. [Google Scholar] [CrossRef]
- Gilbert, M.S.; Ijssennagger, N.; Kies, A.K.; van Mil, S.W.C. Protein fermentation in the gut; implications for intestinal dysfunction in humans, pigs, and poultry. Am. J. Physiol. Gastrointest. Liver Physiol. 2018, 315, G159–G170. [Google Scholar] [CrossRef]
- Williams, B.A.; Verstegen, M.W.; Tamminga, S. Fermentation in the large intestine of single-stomached animals and its relationship to animal health. Nutr. Res. Rev. 2001, 14, 207–228. [Google Scholar] [CrossRef]
- Nyachoti, C.; Lange, C.D.; McBride, B.; Schulze, H. Significance of endogenous gut nitrogen losses in the nutrition of growing pigs: A review. Can. J. Anim. Sci. 1997, 77, 149–163. [Google Scholar]
- Stein, H.H.; Seve, B.; Fuller, M.F.; Moughan, P.J.; de Lange, C.F. Invited review: Amino acid bioavailability and digestibility in pig feed ingredients: Terminology and application. J. Anim. Sci. 2007, 85, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.M.; Kim, J.C.; Hansen, C.F.; Mullan, B.P.; Hampson, D.J.; Pluske, J.R. Feeding a diet with a decreased protein content reduces both nitrogen content in the gastrointestinal tract and post-weaning diarrhoea, but does not affect apparent nitrogen digestibility in weaner pigs challenged with an enterotoxigenic strain of Escherichia coli. Anim. Feed Sci. Technol. 2010, 160, 148–159. [Google Scholar] [CrossRef]
- Yang, Y.; Kim, Y.; Lohakare, J.; Yun, J.; Lee, J.; Kwon, M.; Park, J.; Choi, J.; Chae, B. Comparative efficacy of different soy protein sources on growth performance, nutrient digestibility and intestinal morphology in weaned pigs. Asian Australas. J. Anim. Sci. 2007, 20, 775–783. [Google Scholar]
- Jha, R.; Berrocoso, J.F.D. Dietary fiber and protein fermentation in the intestine of swine and their interactive effects on gut health and on the environment: A review. Anim. Feed Sci. Technol. 2016, 212, 18–26. [Google Scholar] [CrossRef]
- Pieper, R.; Kroger, S.; Richter, J.F.; Wang, J.; Martin, L.; Bindelle, J.; Htoo, J.K.; von Smolinski, D.; Vahjen, W.; Zentek, J.; et al. Fermentable fiber ameliorates fermentable protein-induced changes in microbial ecology, but not the mucosal response, in the colon of piglets. J. Nutr. 2012, 142, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Wellock, I.J.; Fortomaris, P.D.; Houdijk, J.G.M.; Kyriazakis, I. The effect of dietary protein supply on the performance and risk of post-weaning enteric disorders in newly weaned pigs. J. Nutr. 2006, 82, 327–335. [Google Scholar] [CrossRef]
- Cummings, J.H.; Antoine, J.M.; Azpiroz, F.; Bourdet-Sicard, R.; Brandtzaeg, P.; Calder, P.C.; Gibson, G.R.; Guarner, F.; Isolauri, E.; Pannemans, D.; et al. PASSCLAIM--gut health and immunity. Eur. J. Nutr. 2004, 43 (Suppl. 2), II118–II173. [Google Scholar] [CrossRef]
- Flint, H.J.; Scott, K.P.; Louis, P.; Duncan, S.H. The role of the gut microbiota in nutrition and health. Nat. Rev. Gastro. Hepat. 2012, 9, 577–589. [Google Scholar] [CrossRef]
- Slifierz, M.J.; Friendship, R.M.; Weese, J.S. Longitudinal study of the early-life fecal and nasal microbiotas of the domestic pig. BMC. Microbiol. 2015, 15, 184. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.B.; Borewicz, K.; White, B.A.; Singer, R.S.; Sreevatsan, S.; Tu, Z.J.; Isaacson, R.E. Longitudinal investigation of the age-related bacterial diversity in the feces of commercial pigs. Vet. Microbiol. 2011, 153, 124–133. [Google Scholar] [CrossRef]
- Rist, V.T.; Weiss, E.; Eklund, M.; Mosenthin, R. Impact of dietary protein on microbiota composition and activity in the gastrointestinal tract of piglets in relation to gut health: A review. Animal 2013, 7, 1067–1078. [Google Scholar] [CrossRef]
- Kiros, T.G.; Pinloche, E.; D’Inca, R.; Auclair, E.; Van Kessel, A. Model development: Establishing pigs with homogenous microbial profile in the hind gut. Can. J. Anim. Sci. 2018, 98, 498–507. [Google Scholar] [CrossRef]
- Bikker, P.; Dirkzwager, A.; Fledderus, J.; Trevisi, P.; le Huerou-Luron, I.; Lalles, J.P.; Awati, A. The effect of dietary protein and fermentable carbohydrates levels on growth performance and intestinal characteristics in newly weaned piglets. J. Anim. Sci. 2006, 84, 3337–3345. [Google Scholar] [CrossRef]
- Htoo, J.K.; Araiza, B.A.; Sauer, W.C.; Rademacher, M.; Zhang, Y.; Cervantes, M.; Zijlstra, R.T. Effect of dietary protein content on ileal amino acid digestibility, growth performance, and formation of microbial metabolites in ileal and cecal digesta of early-weaned pigs. J. Anim. Sci. 2007, 85, 3303–3312. [Google Scholar] [CrossRef]
- Diether, N.E.; Willing, B.P. Microbial Fermentation of Dietary Protein: An Important Factor in Diet(-)Microbe(-)Host Interaction. Microorganisms 2019, 7, 19. [Google Scholar] [CrossRef]
- Wellock, I.J.; Fortomaris, P.D.; Houdijk, J.G.; Kyriazakis, I. Effects of dietary protein supply, weaning age and experimental enterotoxigenic Escherichia coli infection on newly weaned pigs: Health. Animal 2008, 2, 834–842. [Google Scholar] [CrossRef]
- Lynch, B.; Callan, J.J.; O’Doherty, J.V. The interaction between dietary crude protein and fermentable carbohydrate source on piglet post weaning performance, diet digestibility and selected faecal microbial populations and volatile fatty acid concentration. Livest. Sci. 2009, 124, 93–100. [Google Scholar] [CrossRef]
- Lynch, M.B.; Sweeney, T.; Callan, B.F.J.J.; O’Doherty, J.V. The effect of high and low dietary crude protein and inulin supplementation on nutrient digestibility, nitrogen excretion, intestinal microflora and manure ammonia emissions from finisher pigs. Animal 2007, 1, 1112–1121. [Google Scholar] [CrossRef] [PubMed]
- Leser, T.D.; Amenuvor, J.Z.; Jensen, T.K.; Lindecrona, R.H.; Boye, M.; Møller, K. Culture-Independent Analysis of Gut Bacteria: The Pig Gastrointestinal Tract Microbiota Revisited. Appl. Environ. Microbiol. 2002, 68, 673–690. [Google Scholar] [CrossRef]
- Claesson, M.J.; Wang, Q.; O’Sullivan, O.; Greene-Diniz, R.; Cole, J.R.; Ross, R.P.; O’Toole, P.W. Comparison of two next-generation sequencing technologies for resolving highly complex microbiota composition using tandem variable 16S rRNA gene regions. Nucleic Acids Res. 2010, 38, e200. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.S.; Spakowicz, D.J.; Hong, B.-Y.; Petersen, L.M.; Demkowicz, P.; Chen, L.; Leopold, S.R.; Hanson, B.M.; Agresta, H.O.; Gerstein, M.; et al. Evaluation of 16S rRNA gene sequencing for species and strain-level microbiome analysis. Nat. Commun. 2019, 10, 5029. [Google Scholar] [CrossRef]
- Vandeputte, D.; Kathagen, G.; D’hoe, K.; Vieira-Silva, S.; Valles-Colomer, M.; Sabino, J.; Wang, J.; Tito, R.Y.; De Commer, L.; Darzi, Y.; et al. Quantitative microbiome profiling links gut community variation to microbial load. Nature 2017, 551, 507–511. [Google Scholar] [CrossRef]
- Hillman, E.T.; Lu, H.; Yao, T.; Nakatsu, C.H. Microbial Ecology along the Gastrointestinal Tract. Microbes Environ. 2017, 32, 300–313. [Google Scholar] [CrossRef] [PubMed]
- Fan, P.; Liu, P.; Song, P.; Chen, X.; Ma, X. Moderate dietary protein restriction alters the composition of gut microbiota and improves ileal barrier function in adult pig model. Sci. Rep. 2017, 7, 43412. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Song, P.; Fan, P.; He, T.; Jacobs, D.; Levesque, C.L.; Johnston, L.J.; Ji, L.; Ma, N.; Chen, Y.; et al. Moderate Dietary Protein Restriction Optimized Gut Microbiota and Mucosal Barrier in Growing Pig Model. Front. Cell. Infect. Microbiol. 2018, 8, 246. [Google Scholar] [CrossRef]
- Opapeju, F.O.; Krause, D.O.; Payne, R.L.; Rademacher, M.; Nyachoti, C.M. Effect of dietary protein level on growth performance, indicators of enteric health, and gastrointestinal microbial ecology of weaned pigs induced with postweaning colibacillosis. J. Anim. Sci. 2009, 87, 2635–2643. [Google Scholar] [CrossRef]
- Luo, Z.; Li, C.; Cheng, Y.; Hang, S.; Zhu, W. Effects of low dietary protein on the metabolites and microbial communities in the caecal digesta of piglets. Arch. Anim. Nutr. 2015, 69, 212–226. [Google Scholar] [CrossRef]
- Peng, Y.; Yu, K.; Mu, C.; Hang, S.; Che, L.; Zhu, W. Progressive response of large intestinal bacterial community and fermentation to the stepwise decrease of dietary crude protein level in growing pigs. Appl. Microbiol. Biotechnol. 2017, 101, 5415–5426. [Google Scholar] [CrossRef]
- Kriss, M.; Hazleton, K.Z.; Nusbacher, N.M.; Martin, C.G.; Lozupone, C.A. Low diversity gut microbiota dysbiosis: Drivers, functional implications and recovery. Curr. Opin. Microbiol. 2018, 44, 34–40. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef]
- Mikkelsen, L.L.; Naughton, P.J.; Hedemann, M.S.; Jensen, B.B. Effects of Physical Properties of Feed on Microbial Ecology and Survival of Salmonella enterica Serovar Typhimurium in the Pig Gastrointestinal Tract. Appl. Environ. Microbiol. 2004, 70, 3485–3492. [Google Scholar] [CrossRef]
- Nyachoti, C.M.; Omogbenigun, F.O.; Rademacher, M.; Blank, G. Performance responses and indicators of gastrointestinal health in early-weaned pigs fed low-protein amino acid-supplemented diets. J. Anim. Sci. 2006, 84, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Smith, E.A.; Macfarlane, G.T. Dissimilatory Amino Acid Metabolism in Human Colonic Bacteria. Anaerobe 1997, 3, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Fang, L.; Sun, Y.; Su, Y.; Zhu, W. Effects of the dietary protein level on the microbial composition and metabolomic profile in the hindgut of the pig. Anaerobe 2016, 38, 61–69. [Google Scholar] [CrossRef]
- Zhang, C.; Yu, M.; Yang, Y.; Mu, C.; Su, Y.; Zhu, W. Effect of early antibiotic administration on cecal bacterial communities and their metabolic profiles in pigs fed diets with different protein levels. Anaerobe 2016, 42, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Zhang, C.; Yang, Y.; Mu, C.; Su, Y.; Yu, K.; Zhu, W. Long-term effects of early antibiotic intervention on blood parameters, apparent nutrient digestibility, and fecal microbial fermentation profile in pigs with different dietary protein levels. J. Anim. Sci. Biotechnol. 2017, 8, 60. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Van Kruiningen, H.J.; West, A.B.; Cartun, R.W.; Cortot, A.; Colombel, J.-F. Immunocytochemical evidence of Listeria, Escherichia coil, and Streptococcus antigens in Crohn’s disease. Gastroenterology 1995, 108, 1396–1404. [Google Scholar] [CrossRef]
- Gresse, R.; Chaucheyras-Durand, F.; Fleury, M.A.; Van de Wiele, T.; Forano, E.; Blanquet-Diot, S. Gut Microbiota Dysbiosis in Postweaning Piglets: Understanding the Keys to Health. Trends Microbiol. 2017, 25, 851–873. [Google Scholar] [CrossRef]
- Pluske, J.; Turpin, D.; Kim, J.C. Gastrointestinal tract (gut) health in the young pig. Anim. Nutr. 2018, 4. [Google Scholar] [CrossRef]
- Yu, D.; Zhu, W.; Hang, S. Effects of Long-Term Dietary Protein Restriction on Intestinal Morphology, Digestive Enzymes, Gut Hormones, and Colonic Microbiota in Pigs. Animals 2019, 9, 180. [Google Scholar] [CrossRef]
- Opapeju, F.O.; Rademacher, M.; Blank, G.; Nyachoti, C.M. Effect of low-protein amino acid-supplemented diets on the growth performance, gut morphology, organ weights and digesta characteristics of weaned pigs. Animal 2008, 2, 1457–1464. [Google Scholar] [CrossRef]
- Yu, D.; Zhu, W.; Hang, S. Effects of low-protein diet on the intestinal morphology, digestive enzyme activity, blood urea nitrogen, and gut microbiota and metabolites in weaned pigs. Arch. Anim. Nutr. 2019, 73, 287–305. [Google Scholar] [CrossRef]
- Wellock, I.J.; Fortomaris, P.D.; Houdijk, J.G.; Kyriazakis, I. Effects of dietary protein supply, weaning age and experimental enterotoxigenic Escherichia coli infection on newly weaned pigs: Performance. Animal 2008, 2, 825–833. [Google Scholar] [CrossRef]
- Fan, P.; Li, L.; Rezaei, A.; Eslamfam, S.; Che, D.; Ma, X. Metabolites of Dietary Protein and Peptides by Intestinal Microbes and their Impacts on Gut. Curr. Protein Pept. Sci. 2015, 16, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Nagase, M.; Matsuo, T. Interactions between amino-acid-degrading bacteria and methanogenic bacteria in anaerobic digestion. Biotechnol. Bioeng. 1982, 24, 2227–2239. [Google Scholar] [CrossRef]
- Portune, K.J.; Beaumont, M.; Davila, A.-M.; Tomé, D.; Blachier, F.; Sanz, Y. Gut microbiota role in dietary protein metabolism and health-related outcomes: The two sides of the coin. Trends Food Sci. Technol. 2016, 57, 213–232. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Barrett, A.J.; Thomas, P.D.; Huang, X.; Bateman, A.; Finn, R.D. The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database. Nucleic Acids Res. 2018, 46, D624–D632. [Google Scholar] [CrossRef]
- Booijink, C.C.G.M.; Boekhorst, J.; Zoetendal, E.G.; Smidt, H.; Kleerebezem, M.; de Vos, W.M. Metatranscriptome Analysis of the Human Fecal Microbiota Reveals Subject-Specific Expression Profiles, with Genes Encoding Proteins Involved in Carbohydrate Metabolism Being Dominantly Expressed. Appl. Environ. Microbiol. 2010, 76, 5533. [Google Scholar] [CrossRef] [PubMed]
- Leimena, M.M.; Ramiro-Garcia, J.; Davids, M.; van den Bogert, B.; Smidt, H.; Smid, E.J.; Boekhorst, J.; Zoetendal, E.G.; Schaap, P.J.; Kleerebezem, M. A comprehensive metatranscriptome analysis pipeline and its validation using human small intestine microbiota datasets. BMC Genom. 2013, 14, 530. [Google Scholar] [CrossRef]
- Ugarte, A.; Vicedomini, R.; Bernardes, J.; Carbone, A. A multi-source domain annotation pipeline for quantitative metagenomic and metatranscriptomic functional profiling. Microbiome 2018, 6, 149. [Google Scholar] [CrossRef]
- Shakya, M.; Lo, C.-C.; Chain, P.S.G. Advances and Challenges in Metatranscriptomic Analysis. Front. Genet. 2019, 10, 904. [Google Scholar] [CrossRef]
- Yao, C.K.; Muir, J.G.; Gibson, P.R. Review article: Insights into colonic protein fermentation, its modulation and potential health implications. Aliment. Pharmacol. Ther. 2016, 43, 181–196. [Google Scholar] [CrossRef]
- Macfarlane, G.T.; Cummings, J.H. The colonic flora, fermentation and large bowel digestive function. In The Large Intestine: Physiology, Pathophysiology and Disease; Philips, S.F., Pemberton, J.H., Shorter, R.G., Eds.; Raven Press: New York, NY, USA, 1991; pp. 51–92. [Google Scholar]
- Smith, E.A.; Macfarlane, G.T. Enumeration of human colonic bacteria producing phenolic and indolic compounds: Effects of pH, carbohydrate availability and retention time on dissimilatory aromatic amino acid metabolism. J. Appl. Microbiol. 1996, 81, 288–302. [Google Scholar] [CrossRef]
- Smith, E.A.; Macfarlane, G.T. Studies on Amine Production in the Human Colon: Enumeration of Amine forming Bacteria and Physiological Effects of Carbohydrate and pH. Anaerobe 1996, 2, 285–297. [Google Scholar] [CrossRef]
- Libao-Mercado, A.J.O.; Zhu, C.L.; Cant, J.P.; Lapierre, H.l.n.; Thibault, J.-N.l.; Sève, B.; Fuller, M.F.; de Lange, C.F.M. Dietary and Endogenous Amino Acids Are the Main Contributors to Microbial Protein in the Upper Gut of Normally Nourished Pigs. J. Nutr. 2009, 139, 1088–1094. [Google Scholar] [CrossRef]
- Pieper, R.; Boudry, C.; Bindelle, J.; Vahjen, W.; Zentek, J. Interaction between dietary protein content and the source of carbohydrates along the gastrointestinal tract of weaned piglets. Arch. Anim. Nutr. 2014, 68, 263–280. [Google Scholar] [CrossRef]
- Mouillé, B.; Delpal, S.; Mayeur, C.; Blachier, F. Inhibition of human colon carcinoma cell growth by ammonia: A non-cytotoxic process associated with polyamine synthesis reduction. Biochim. Biophys. Acta Gen. Subj. 2003, 1624, 88–97. [Google Scholar] [CrossRef]
- Pieper, R.; Villodre Tudela, C.; Taciak, M.; Bindelle, J.; Perez, J.F.; Zentek, J. Health relevance of intestinal protein fermentation in young pigs. Anim. Health Res. Rev. 2016, 17, 137–147. [Google Scholar] [CrossRef]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef]
- Aguirre, M.; Eck, A.; Koenen, M.E.; Savelkoul, P.H.; Budding, A.E.; Venema, K. Diet drives quick changes in the metabolic activity and composition of human gut microbiota in a validated in vitro gut model. Res. Microbiol. 2016, 167, 114–125. [Google Scholar] [CrossRef]
- Villodre Tudela, C.; Boudry, C.; Stumpff, F.; Aschenbach, J.R.; Vahjen, W.; Zentek, J.; Pieper, R. Down-regulation of monocarboxylate transporter 1 (MCT1) gene expression in the colon of piglets is linked to bacterial protein fermentation and pro-inflammatory cytokine-mediated signalling. Br. J. Nutr. 2015, 113, 610–617. [Google Scholar] [CrossRef]
- Heo, J.M.; Kim, J.C.; Hansen, C.F.; Mullan, B.P.; Hampson, D.J.; Pluske, J.R. Feeding a diet with decreased protein content reduces indices of protein fermentation and the incidence of postweaning diarrhea in weaned pigs challenged with an enterotoxigenic strain of Escherichia coli. J. Anim. Sci. 2009, 87, 2833–2843. [Google Scholar] [CrossRef]
- Hughes, R.; Magee, E.A.; Bingham, S. Protein degradation in the large intestine: Relevance to colorectal cancer. Curr. Issues Intestig. Microbiol. 2000, 1, 51–58. [Google Scholar]
- Ahrens, R.; Gäbel, G.; Garz, B.; Aschenbach, J.R. Histamine-induced chloride secretion is mediated via H2-receptors in the pig proximal colon. Inflamm. Res. 2003, 52, 79–85. [Google Scholar] [CrossRef]
- Kroger, S.; Pieper, R.; Schwelberger, H.G.; Wang, J.; Villodre Tudela, C.; Aschenbach, J.R.; Van Kessel, A.G.; Zentek, J. Diets high in heat-treated soybean meal reduce the histamine-induced epithelial response in the colon of weaned piglets and increase epithelial catabolism of histamine. PLoS ONE 2013, 8, e80612. [Google Scholar] [CrossRef]
- Andriamihaja, M.; Lan, A.; Beaumont, M.; Audebert, M.; Wong, X.; Yamada, K.; Yin, Y.; Tomé, D.; Carrasco-Pozo, C.; Gotteland, M.; et al. The deleterious metabolic and genotoxic effects of the bacterial metabolite p-cresol on colonic epithelial cells. Free Radic. Biol. Med. 2015, 85, 219–227. [Google Scholar] [CrossRef]
- Bansal, T.; Alaniz, R.C.; Wood, T.K.; Jayaraman, A. The bacterial signal indole increases epithelial-cell tight-junction resistance and attenuates indicators of inflammation. Proc. Natl. Acad. Sci. USA 2010, 107, 228–233. [Google Scholar] [CrossRef] [PubMed]
- Richter, J.F.; Pieper, R.; Zakrzewski, S.S.; Gunzel, D.; Schulzke, J.D.; Van Kessel, A.G. Diets high in fermentable protein and fibre alter tight junction protein composition with minor effects on barrier function in piglet colon. Br. J. Nutr. 2014, 111, 1040–1049. [Google Scholar] [CrossRef]
- Linden, D.R. Hydrogen Sulfide Signaling in the Gastrointestinal Tract. Antioxid. Redox Signal. 2014, 20, 818–830. [Google Scholar] [CrossRef] [PubMed]
- Cone, J.W.; Jongbloed, A.W.; Gelder, A.H.V.; Lange, L.d. Estimation of protein fermentation in the large intestine of pigs using a gas production technique. Anim. Feed Sci. Technol. 2005, 123–124, 463–472. [Google Scholar] [CrossRef]
- Roediger, W.E. Utilization of nutrients by isolated epithelial cells of the rat colon. Gastroenterology 1982, 83, 424–429. [Google Scholar] [CrossRef]
- Scheppach, W. Effects of short chain fatty acids on gut morphology and function. Gut 1994, 35, S35–S38. [Google Scholar] [CrossRef]
- Macfarlane, G.T.; Steed, H.; Macfarlane, S. Bacterial metabolism and health-related effects of galacto-oligosaccharides and other prebiotics. J. Appl. Microbiol. 2008, 104, 305–344. [Google Scholar] [CrossRef] [PubMed]
- Hermes, R.G.; Molist, F.; Ywazaki, M.; Nofrarias, M.; Gomez de Segura, A.; Gasa, J.; Perez, J.F. Effect of dietary level of protein and fiber on the productive performance and health status of piglets. J. Anim. Sci. 2009, 87, 3569–3577. [Google Scholar] [CrossRef]
- Gilbert, M.S.; van der Hee, B.; Gerrits, W.J.J. The role of protein fermentation metabolites in post-weaning diarrhoea in piglets. In Energy and Protein Metabolism and Nutrition; Wageningen Academic Publishers: Wageningen, The Netherlands, 2019; Volume 138, pp. 361–362. [Google Scholar]
- Kaiko, G.E.; Ryu, S.H.; Koues, O.I.; Collins, P.L.; Solnica-Krezel, L.; Pearce, E.J.; Pearce, E.L.; Oltz, E.M.; Stappenbeck, T.S. The Colonic Crypt Protects Stem Cells from Microbiota-Derived Metabolites. Cell 2016, 165, 1708–1720. [Google Scholar] [CrossRef]
- Peng, X.; Hu, L.; Liu, Y.; Yan, C.; Fang, Z.F.; Lin, Y.; Xu, S.Y.; Li, J.; Wu, C.M.; Chen, D.W.; et al. Effects of low-protein diets supplemented with indispensable amino acids on growth performance, intestinal morphology and immunological parameters in 13 to 35 kg pigs. Animal 2016, 10, 1812–1820. [Google Scholar] [CrossRef]
- Vente-Spreeuwenberg, M.A.M.; Verdonk, J.M.A.J.; Bakker, G.C.M.; Beynen, A.C.; Verstegen, M.W.A. Effect of dietary protein source on feed intake and small intestinal morphology in newly weaned piglets. Livest. Prod. Sci. 2004, 86, 169–177. [Google Scholar] [CrossRef]
- Chelakkot, C.; Ghim, J.; Ryu, S.H. Mechanisms regulating intestinal barrier integrity and its pathological implications. Exp. Mol. Med. 2018, 50, 103. [Google Scholar] [CrossRef]
- Boudry, G.; Jamin, A.; Chatelais, L.; Gras-Le Guen, C.; Michel, C.; Le Huërou-Luron, I. Dietary Protein Excess during Neonatal Life Alters Colonic Microbiota and Mucosal Response to Inflammatory Mediators Later in Life in Female Pigs. J. Nutr. 2013, 143, 1225–1232. [Google Scholar] [CrossRef]
- Heo, J.M.; Kim, J.C.; Yoo, J.; Pluske, J.R. A between-experiment analysis of relationships linking dietary protein intake and post-weaning diarrhea in weanling pigs under conditions of experimental infection with an enterotoxigenic strain of Escherichia coli. Anim. Sci. J. 2015, 86, 286–293. [Google Scholar] [CrossRef]
- Roediger, W.E.W.; Moore, J.; Babidge, W. Colonic Sulfide in Pathogenesis and Treatment of Ulcerative Colitis. Dig. Dis. Sci. 1997, 42, 1571–1579. [Google Scholar] [CrossRef]
- Yen, J.T.; Kerr, B.J.; Easter, R.A.; Parkhurst, A.M. Difference in rates of net portal absorption between crystalline and protein-bound lysine and threonine in growing pigs fed once daily. J. Anim. Sci. 2004, 82, 1079–1090. [Google Scholar] [CrossRef]
- van den Borne, J.J.G.C.; Schrama, J.W.; Heetkamp, M.J.W.; Verstegen, M.W.A.; Gerrits, W.J.J. Synchronising the availability of amino acids and glucose increases protein retention in pigs. Animal 2007, 1, 666–674. [Google Scholar] [CrossRef]
- Yue, L.Y.; Qiao, S.Y. Effects of low-protein diets supplemented with crystalline amino acids on performance and intestinal development in piglets over the first 2 weeks after weaning. Livest. Sci. 2008, 115, 144–152. [Google Scholar] [CrossRef]
- Bindelle, J.; Buldgen, A.; Delacollette, M.; Wavreille, J.; Agneessens, R.; Destain, J.P.; Leterme, P. Influence of source and concentrations of dietary fiber on in vivo nitrogen excretion pathways in pigs as reflected by in vitro fermentation and nitrogen incorporation by fecal bacteria123. J. Anim. Sci. 2009, 87, 583–593. [Google Scholar] [CrossRef] [PubMed]
- Jérôme, B.; Pascal, L.; Buldgen, A. Nutritional and environmental consequences of dietary fibre in pig nutrition: A review. Biotechnol. Agron. Soc. Environ. 2008, 12, 69–80. [Google Scholar]


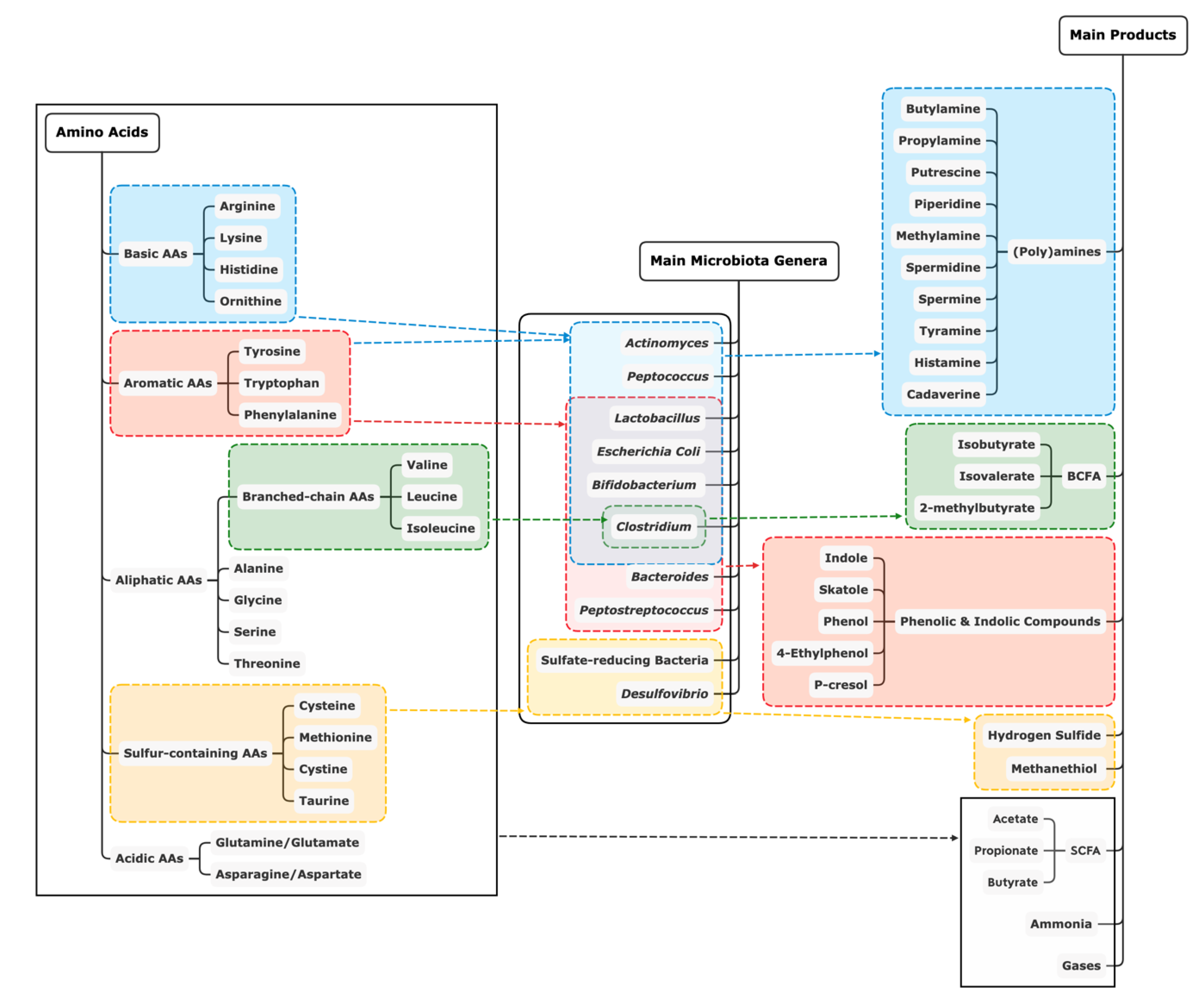{kind=link}
| Age (d) | Crude Protein Level (%) | Period (d) | Method | Microbiota Responses | Source | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Richness | Diversity | Phylum | Class | Order | Family | Genus | Species | |||||
| 17 | 19 vs. 21 | 21 | Culture-Based | Caecum = | [47] | |||||||
| 17 | 18 vs. 23 | 14 | Culture-based; TRFLP | - | Ileum, colon and rectum =: Total Coliforms, lactic acid producing bacteria | [32] a | ||||||
| Colon ↑ 7d post challenge | Colon = | = | ↓: Clostridiales ↑: Unclassified Clostridiales | ↓: Lachnospiraceae | ↓: Roseburia | - | ||||||
| 18 | 17 vs. 19 vs. 21 vs. 23 | 21 | Culture-Based | - | - | Ileum =: Aerobic spore formers, Anaerobic spore formers, Enterobacteriaceae, Enterococci, E. coli, Total Coliforms | [38] | |||||
| 24 | 16 vs. 20 | 21 | Culture-based | - | - | Faeces =: Enterobacteria spp., Lactobacilli spp. | [23] | |||||
| 25 | 15 vs. 20 | 21-23 | Real-time PCR | - | - | Proximal colon =: Total bacteria | - | - | =: Enterobacteria | =: Lactobacilli, Bacteroides | =: Cl. coccoides ↑: Cl. leptum | [11] |
| 26 | 15 vs. 22 | 28 | Culture-based | - | - | Ileum and colon =: E. coli, Total Coliforms, Lactobacilli | [19] | |||||
| 28 | 14 vs. 17 vs. 20 | 45 | 16S rRNA, V3-V4 regions | Jejunum and colon = | = | - | - | - | = | - | [48] | |
| 28 | 13 vs. 18 vs. 23 | 14 | Culture based | - | - | Ileum = Proximal colon ↑: Coliforms ↓: lactobacillus to Coliforms ratios Faeces ↑: Coliforms | [49] | |||||
| 28/40 | 13 vs. 23 | 14 | Culture based | - | - | Colon ↑: Lactobacilli =: Coliform, Lactobacillus to Coliforms ratio Faeces numerical ↑enterotoxigenic E. coli | [49] | |||||
| ~35 | 14 vs. 20 | 45 | 16S rDNA, V6–V8 regions; DGGE | - | Caecum ↑ (d25) | ↑: Firmicutes (d25) =: total bacteria, Bacteroidetes | - | - | - | ↑: Clostridium cluster IV (d25) =: Clostridium cluster XIVa (d10, 45) | [33] | |
| 35 | 10 vs. 13 vs. 16 | 112 | 16S rRNA, V3–V4 regions | Colon = | - | - | - | - | ↑: Streptococcus, Lactobacillus, Turicibacter ↓: Prevotella, Lachnospira Ruminococcus, Dorea, Candidatus, Unclassified Clostridiales, Uncultured Peptococcaceae | - | [46] b | |
| 42 | 14 vs. 18 | 143 | DGGE; Real-time PCR; 16S rRNA, V6–V8 regions | Caecum = Faeces | Caecum =: total bacteria, Firmicutes ↓: Bacteroidetes Faeces =: total bacteria, Firmicutes, Bacteroidetes | - | - | - | - | Caecum ↑: E. coli (d77) ↓: Clostridium cluster IV, Clostridium cluster XIVa Faeces ↑: E. coli (d77) =: Clostridium cluster IV, Clostridium cluster XIVa | [41,42] b | |
| 45 | 14 vs. 15 vs. 17 vs. 20 | 28 | Real-time PCR | - | Ileum, caecum = Colon: ↑ ↓= | Ileum, caecum, colon =: total bacteria, Firmicutes, Bacteroidetes | - | - | - | Caecum ↑: Bifidobacterium (15, 20%) Colon ↑: Bifidobacterium Ileum, caecum, colon =: Lactobacillus | Colon↓: E. coli (15%) Ileum, caecum, colon =: Clostridium cluster IV, Clostridium cluster XIVa | [34] |
| 70 | 13 vs. 16 | 100 | 16S rRNA, V1-V3 regions | Caecum, colon = | = | - | - | Caecum ↓: unclassified Peptostreptococcaceae, uncultured Lachnospiraceae, and uncultured Erysipelotrichaceae Colon ↓: Unclassified Clostridiaceae, and Erysipelotrichaceae incertae sedis | Caecum ↑: Lactobacillus ↓: Prevotella, Coprococcus Colon ↑: Streptococcus ↓: Sarcina, Peptostreptococcaceae incertae sedis, Mogibacterium, Subdoligranulum, Coprococcus | - | [40] b | |
| ~80 | 12 vs. 15 vs. 18 | 30 | 16S rRNA, V3–V4 regions | Ileum, colon = | Ileum ↑↓: Firmicutes ↓↑: Proteobacteria Colon ↓↑: Bacteroides ↑↓: Firmicutes, Spirochaetae ↑: Verrucomicrobia | - | - | Ileum ↓↑: Streptococcaceae, Enterobacteriaceae, Leuconostocaceae ↑↓: Lactobacillaceae, Closridiaceae_1, Micrococcaceae Colon = ↓: Lachnospiraceae, ↑↓: Veillonellaceae ↑: Ruminococcaceae | Ileum ↓↑: Streptococcus, Escherichia-Shigella, Weissella ↑↓: Lactobacillus, Clostridium_sensu_stricto_1 Colon ↓↑: Ruminococcaceae_UCG- 005, Norank_f _Bacteroidales_S24-7_group ↑↓: Streptococcus ↑: Prevotellaceae_NK3B31_group | - | [31] | |
| Finishing | 10 vs. 13 vs. 16 | 50 | 16S RNA, V3–V4 regions | Ileum, Colon = | Ileum =↑: Firmicutes =↓: Proteobacteria =: Actinobacteria Colon=: Firmicutes, Bacteroidetes, Spirochaetae | - | - | Ileum =↑: Clostridiaceae_1 =↓: Enterobacteria Colon ↓: Clostridiaceae_1, Erysipelotrichaceae =↑: Rikenellaceae ↑↓: Peptostreptococcaceae =: Spirochaetaceae | Ileum = ↑: Clostridium_sensu_stricto_1 ↓: Escherichia-Shigella Colon =↓: Clostridium_sensu_stricto_1, Turicibacter | - | [30] | |
| Finishing | 15 vs. 20 | ~30 | Culture-based | - | - | - | - | - | - | - | Caecum = Colon ↓: Bifidobacteria spp. =: Lactobacilli spp., Enterobacteria spp. | [24] |
| Age (d) | Duration (d) | Crude Protein Level (%) | Segment | Change in Digesta Concentrations | Source | ||||
|---|---|---|---|---|---|---|---|---|---|
| pH | Short-Chain Fatty Acids | Branched-Chain Fatty Acids | Biogenic Amines/Indolic and Phenolic Compounds | Ammonia | |||||
| 17 | 14 | 18 vs. 23 | Colon | - | = | = | - | ↑ | [32] a |
| 17 | 21 | 19 vs. 21 | Caecum | - | = | - | - | ↑ | [47] |
| 18 | 21 | 17 vs. 19 vs. 21 vs. 23 | Duodenum | = | ↑: propionic, valeric | = | - | ↑ | [38] |
| Jejunum | = | = | = | - | ↑ | ||||
| Ileum | ↑ | ↑: acetic, propionic, valeric | ↑: isobutyric, isovaleric | - | ↑ | ||||
| 21 | 28 | 18 vs. 26 | Faeces | - | ↑ | = (%) | - | ↑ | [69] a |
| 21 | 21 | 15 vs. 19 | Colon | - | ↑ | = | - | - | [81] |
| 24 | 21 | 16 vs. 20 | Faeces | = | ↑: total SCFA, butyric acid (%) | = | - | - | [23] |
| 25 | 21-23 | 15 vs. 20 | Colon | - | ↑ | ↑ | ↑: putrescine, histamine, spermidine | ↑ | [11] |
| 25 | 21 | 18 vs. 26 | Stomach | = | = | - | ↓: cadaverine, phenol | ↑ | [63] |
| Ileum | = | =: total SCFA ↑: acetate (%), butyrate (%), propionate (%) | - | ↑: total amines, putrescine, ↓: phenol | ↑ | ||||
| Caecum | = | ↑: total SCFA, acetate, butyrate, propionate (%), butyrate (%) | = | ↑: histamine, 4-ethylphenol | ↑ | ||||
| Proximal colon | = | = | = | ↑: total amines | ↑ | ||||
| Distal colon | = | =: total SCFA ↑: butyrate ↓: acetate (%) | = | ↑: cadaverine, phenol, p-cresol, skatole | = | ||||
| 25 | 22 | 18 vs. 26 | Proximal colon | - | ↑: total SCFA, butyrate | ↑ | ↑: putrescine, histamine, and spermidine | ↑ | [68] |
| 26 | 28 | 15 vs. 22 | Ileum | - | = | = | - | ↑ | [19] |
| Colon | = | = | - | = | |||||
| 28 | 14 | 20 vs. 24 | Caecum | = | ↑: acetic acid | ↑: isobutyric acid, isovaleric acid | ↑: putrescine | ↑ | [20] |
| 28 | 45 | 14 vs. 17 vs. 20 | Jejunum | - | = | = | = ↑: tyramine | = | [48] |
| Colon | - | = | = | = | = ↑ | ||||
| 33 | 14 | 20 vs. 24 | Ileum | = | = | = | = | = | [20] |
| ~35 | 10 | 14 vs. 20 | Caecum | - | = | = | = | = | [33] |
| 25 | ↑: acetate =: total SCFA | ↑ | ↑: cadaverine | ↑ | |||||
| 45 | ↑: acetate, total SCFA | ↑ | ↑: cadaverine | ↑ | |||||
| 35 | 112 | 10 vs. 13 | Colon | - | = | ↑: isobutyrate, isovalerate | = | = | [46] c |
| 13 vs. 16 | - | ↑: total SCFA, acetate | = | ↑: tryptamine, putrescine, cadaverine | ↑ | ||||
| 42 | 78 | 10 vs. 14 | Caecum | - | ↓: acetate, propionate | ↑: isobutyrate | ↑: tyramine, cadaverine, phenol and indole | ↑ | [41] b,c |
| 35 | Faeces | - | =: total SCFA, acetate, propionate, butyrate, valerate | = | ↑: total amines, methylamine, tryptamine, cadaverine, tyramine, skatole, p-cresol, indole =: putrescine, spermidine, spermine, phenol | ↑ | [42] b | ||
| 143 | ↑: total SCFA, acetate, propionate =: butyrate, valerate | ↑: isobutyrate, isovalerate | ↑: total amines, putrescine, spermidine, methylamine, tryptamine, cadaverine, tyramine, skatole, phenol ↓: p-cresol =: spermine, indole | ||||||
| 45 | 28 | 14 vs. 15 vs. 17 vs. 20 | Ileum | - | ↓: total SCFA, butyrate ↑=: Valerate | ↓: isobutyrate ↑: isovalerate = BCFA | ↑: total amines, cadaverine, putrescine | ↑ | [34] |
| Caecum | - | ↑: total SCFA, butyrate | ↓: isovalerate, BCFA | ↑: histamine, spermidine | ↑ | ||||
| Colon | - | ↑: total SCFA, acetate, valerate | = | ↑: total amines, tryptamine, putrescine, spermidine | ↑ | ||||
| 70 | 100 | 15/13 vs. 18/16 | Caecum | - | =: total SCFA, acetate, propionate, butyrate, valerate | ↑: isobutyrate, isovalerate; BCFA/SCFA | - | - | [40] c |
| Colon | - | = | = | - | - | ||||
| ~80 | 30 | 12 vs. 15 vs. 18 | Ileum | - | = | = | ↑: putrescine, histamine, spermidine | - | [31] |
| colon | - | ↑: acetic acid | = | ↑: cadaverine, spermidine | - | ||||
| Finishing | 50 | 10 vs. 13 vs. 16 | Ileum | - | ↑: acetic acid ↓: valeric acid | = | ↑: methylamine, cadaverine, putrescine, histamine, spermidine | - | [30] |
| Colon | ↑: acetic acid, propionic acid, butyric acid | ↑: isobutyric acid, isovaleric acid | ↑: methylamine, putrescine, histamine, spermidine | - | |||||
| Finishing | ~30 | 15 vs. 20 | Caecum | = | = | = (%) | - | - | [24] |
| Colon | = | =: total SCFA ↑: butyric acid (%) | = (%) | - | - | ||||
| Faeces | ↑ | - | - | - | ↑ | ||||
| Age (d) | Duration (d) | Crude Protein Level (%) | Growth Performance | Faecal Fluidity | Organ Weight | Intestinal Responses | Source | ||
|---|---|---|---|---|---|---|---|---|---|
| Morphology | Integrity | Immunity | |||||||
| 17 | 14 | 18 vs. 23 | ↑: ADG ↓: FCR (Before) | ↑ (After) | - | Ileum ↑: CD (before); ↓: VH (after), VCR | - | - | [32] a |
| 17 | 21 | 19 vs. 21 | ↑: ADG ↓: FCR | ↑ | - | Duodenum, ileum ↑: CD | - | - | [47] |
| 18 | 21 | 17 vs. 19 vs. 21 vs. 23 | ↑: ADG, ADFI, BW ↓: FCR | = | =: spleen ↑: ST, liver | Duodenum = Jejunum ↑: VH, CD ↓: VCR | - | - | [38] |
| 18 | 14 | 17 vs. 19 vs. 21 vs. 22 | =: ADFI ↑: ADG ↓: FCR | ↑ | - | Duodenum, jejunum ↑: VH Ileum = | - | - | [92] |
| 25 | 21/23 | 15 vs. 20 | = | - | - | - | Proximal colon ↑: PCNA, | Proximal colon ↑: IL1β, IL10, TGFβ, MUC1, MUC2 and MUC20 | [11] |
| 26 | 28 | 15 vs. 22 | = | - | - | = VH, CD | =: density of gut wall muscularis, serosa, mucosa | - | [19] |
| 29 | 14 | 13 vs. 18 vs. 23 | ↑: ADG, =: ADFI ↓: FCR | ↑ | ↑: ST, SI | - | - | - | [12] |
| 28/40 | 14 | 13 vs. 23 | ↓: ADFI, FCR | ↑ | - | = VH, CD | - | - | [49] a |
| 31 | 45 | 14 vs. 17 vs. 20 | ↑: BW, ADG, ADFI ↓: FCR | - | - | Duodenum↑: VH, CD, VCR Jejunum↑: VH, VCR Ileum = | - | - | [48] |
| 35 | 112 | 10 vs. 13 vs. 16 | ↑: BW, ADG, ADFI, FCR | - | - | Duodenum↑: VH, CD; =: VCR Jejunum↑: VCR Ileum = | - | - | [46] b |
| 35 | 21 | 16 vs. 20 | = | = | ↑: ST, LT (%BW) =: liver (%BW) | - | Proximal colon ↑: goblet cells | Proximal colon ↓: intraepithelial lymphocytes | [81] |
| 45 | 28 | 14 vs. 15 vs. 17 vs. 20 | =: ADFI ↑: BW, ADG ↓: FCR | - | =: heart, spleen, kidney ↑: liver, pancreas | Duodenum ↑: VH, CD Jejunum, ileum ↑: CD; ↓: VCR | - | Ileum ↑ mRNA: TLR-4, MyD88, NF-kB; ↓: TOLLIP Plasma ↑: CD3+T cells, IgG =: CD3+CD4+T cells, CD3+CD8+T cells | [84] |
| ~80 | 30 | 12 vs. 15 vs. 18 | - | - | - | Ileum ↑: VH (18%), CD (15%) colon ↑: CD (15, 18%) | Serum ↓ =: LPS Ileum ↑: claudin-3 (18%), claudin-7 (15, 18%)Colon ↑: occludin, ZO-3, claudin-1, claudin-7 | - | [31] |
| Finishing | 50 | 10 vs. 13 vs. 16 | ↑: BW | - | - | Ileum ↑: VH ↓: CD | Ileum ↑: claudin-1, occludin (13%) Colon =: tight junction proteins | - | [30] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Wielen, N.v.d.; Hee, B.v.d.; Wang, J.; Hendriks, W.; Gilbert, M. Impact of Fermentable Protein, by Feeding High Protein Diets, on Microbial Composition, Microbial Catabolic Activity, Gut Health and beyond in Pigs. Microorganisms 2020, 8, 1735. https://doi.org/10.3390/microorganisms8111735
Zhang H, Wielen Nvd, Hee Bvd, Wang J, Hendriks W, Gilbert M. Impact of Fermentable Protein, by Feeding High Protein Diets, on Microbial Composition, Microbial Catabolic Activity, Gut Health and beyond in Pigs. Microorganisms. 2020; 8(11):1735. https://doi.org/10.3390/microorganisms8111735
Chicago/Turabian StyleZhang, Hanlu, Nikkie van der Wielen, Bart van der Hee, Junjun Wang, Wouter Hendriks, and Myrthe Gilbert. 2020. "Impact of Fermentable Protein, by Feeding High Protein Diets, on Microbial Composition, Microbial Catabolic Activity, Gut Health and beyond in Pigs" Microorganisms 8, no. 11: 1735. https://doi.org/10.3390/microorganisms8111735
APA StyleZhang, H., Wielen, N. v. d., Hee, B. v. d., Wang, J., Hendriks, W., & Gilbert, M. (2020). Impact of Fermentable Protein, by Feeding High Protein Diets, on Microbial Composition, Microbial Catabolic Activity, Gut Health and beyond in Pigs. Microorganisms, 8(11), 1735. https://doi.org/10.3390/microorganisms8111735