
Genome-Wide Identification and Expression Pattern Analysis of the Late Embryogenesis Abundant (LEA) Family in Foxtail Millet (Setaria italica L.)

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Identification and Chromosomal Localization Analysis of SiLEA Gene Family Members in Foxtail Millet
2.2. Physicochemical Properties of SiLEA Gene Family Members in Foxtail Millet
2.3. Phylogenetic Analysis of Foxtail Millet SiLEA Gene Family Members
2.4. Conserved Domain and Gene Structure Analysis of Foxtail Millet SiLEA Genes
2.5. Analysis of SiLEA Gene Promoter Cis-Regulatory Elements in Foxtail Millet
2.6. Collinearity and Evolutionary Patterns of SiLEA Genes
2.7. Tissue-Specific Expression Analysis of Foxtail Millet SiLEA Gene Family Members
2.8. Expression Analysis of SiLEA Gene in Response to ABA in Different Tissues of Foxtail Millet by qRT-PCR
3. Results
3.1. Identification, Physicochemical Characterization, and Chromosomal Localization of SiLEA Gene Family Members in Foxtail Millet
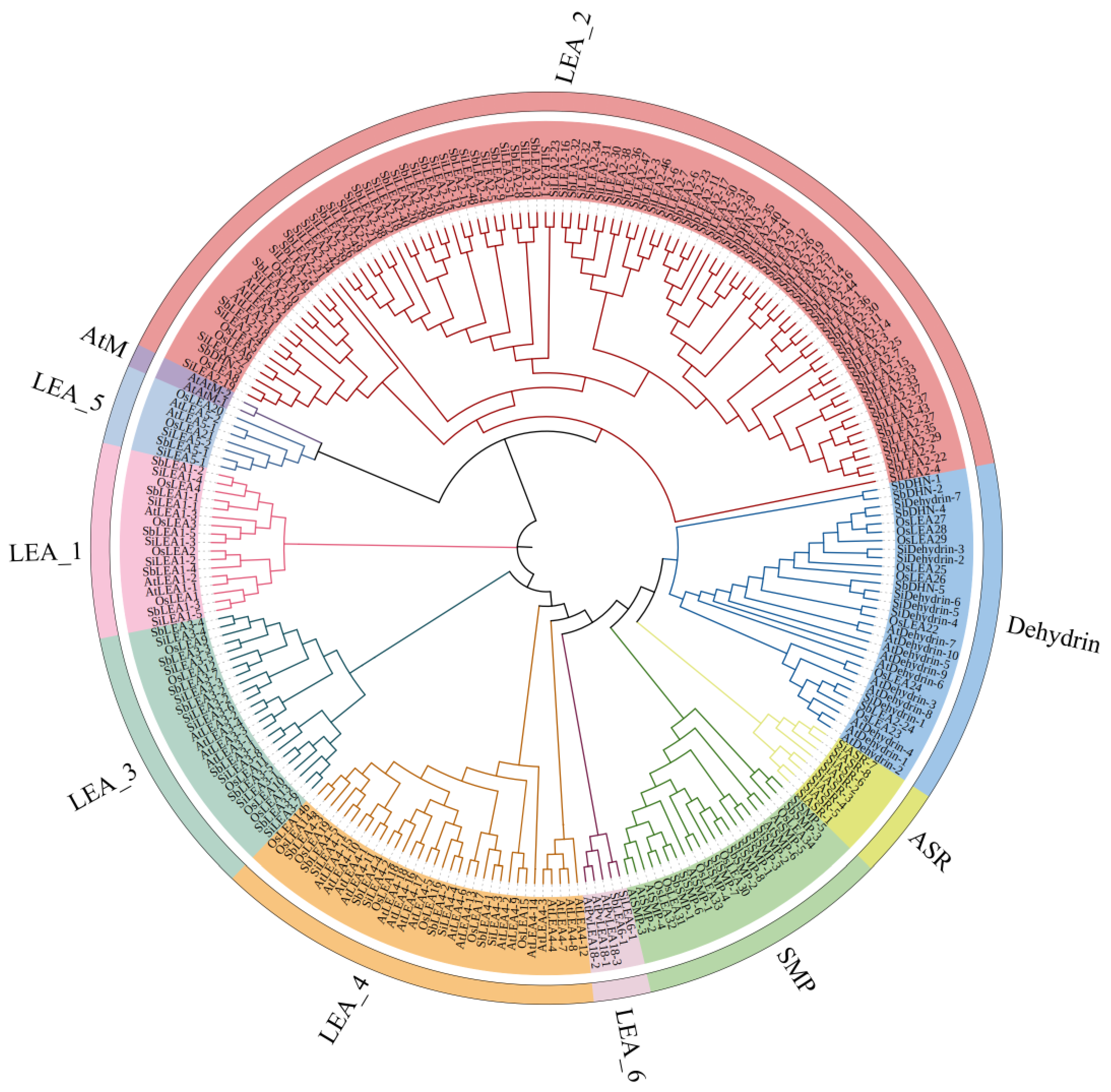
3.2. Phylogenetic Analysis of SiLEA Gene Family Members
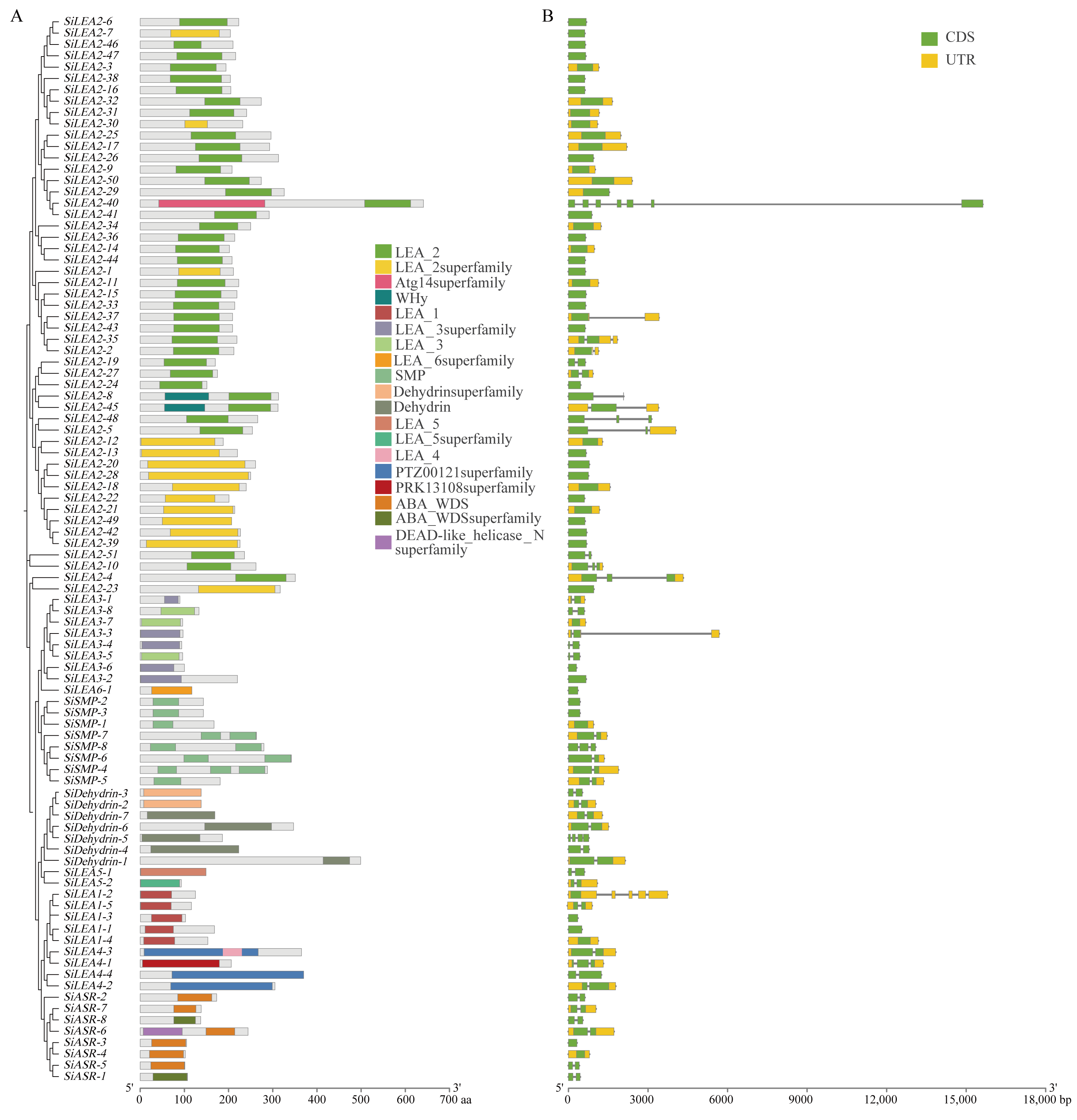
3.3. Conserved Domains and Gene Structures of SiLEA Genes in Foxtail Millet
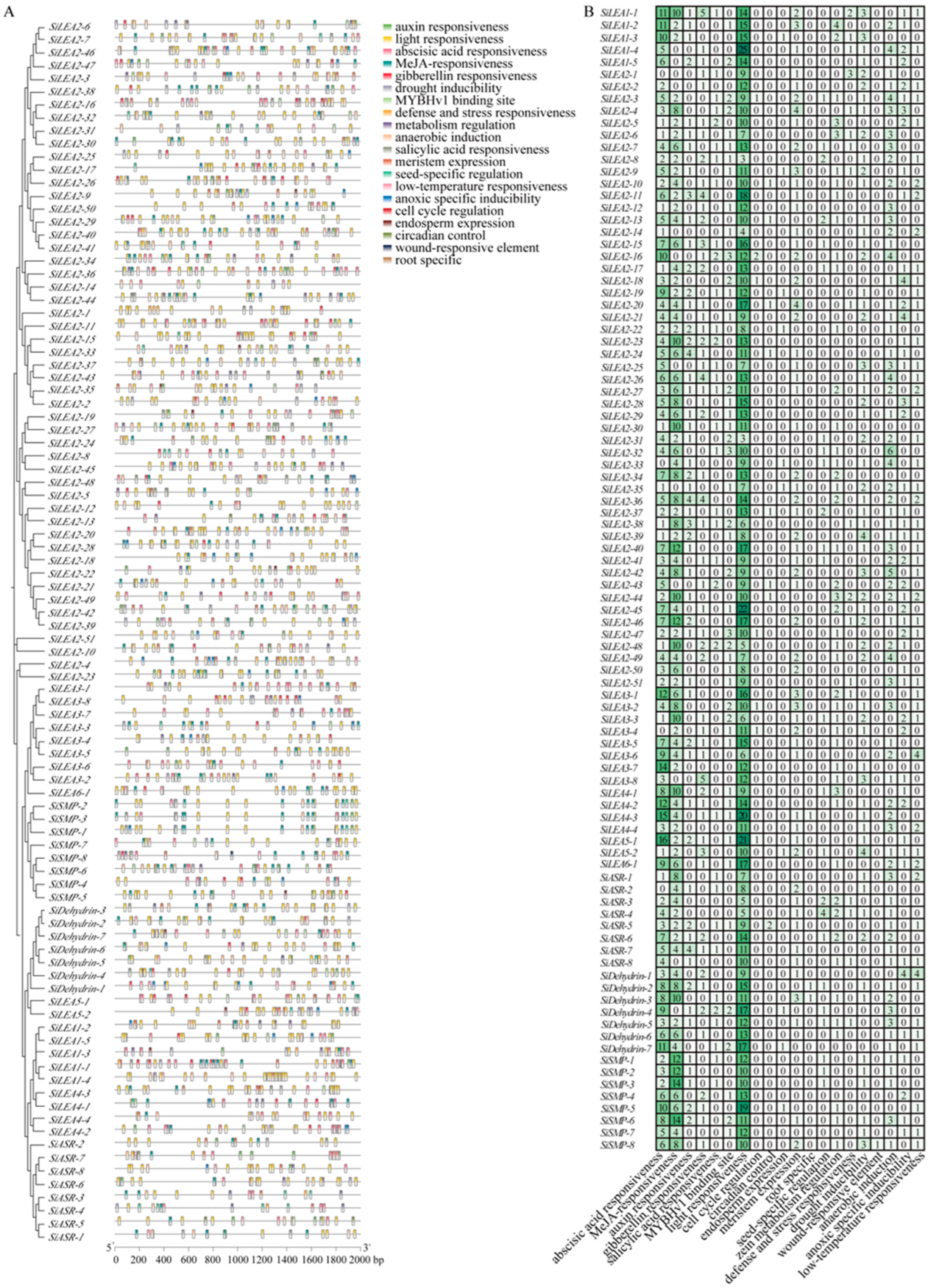
3.4. Promoter Cis-Acting Elements in SiLEA Genes
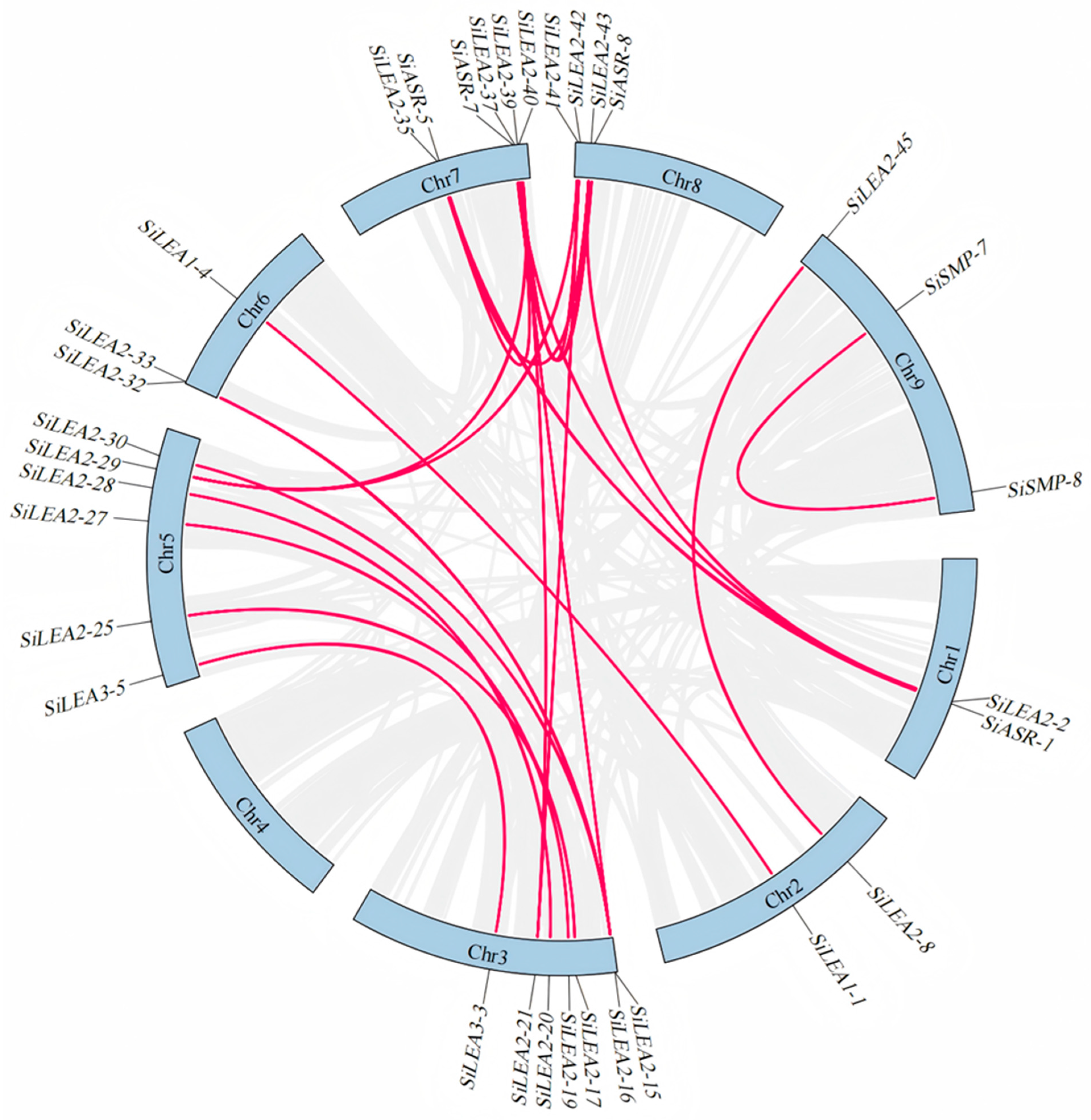
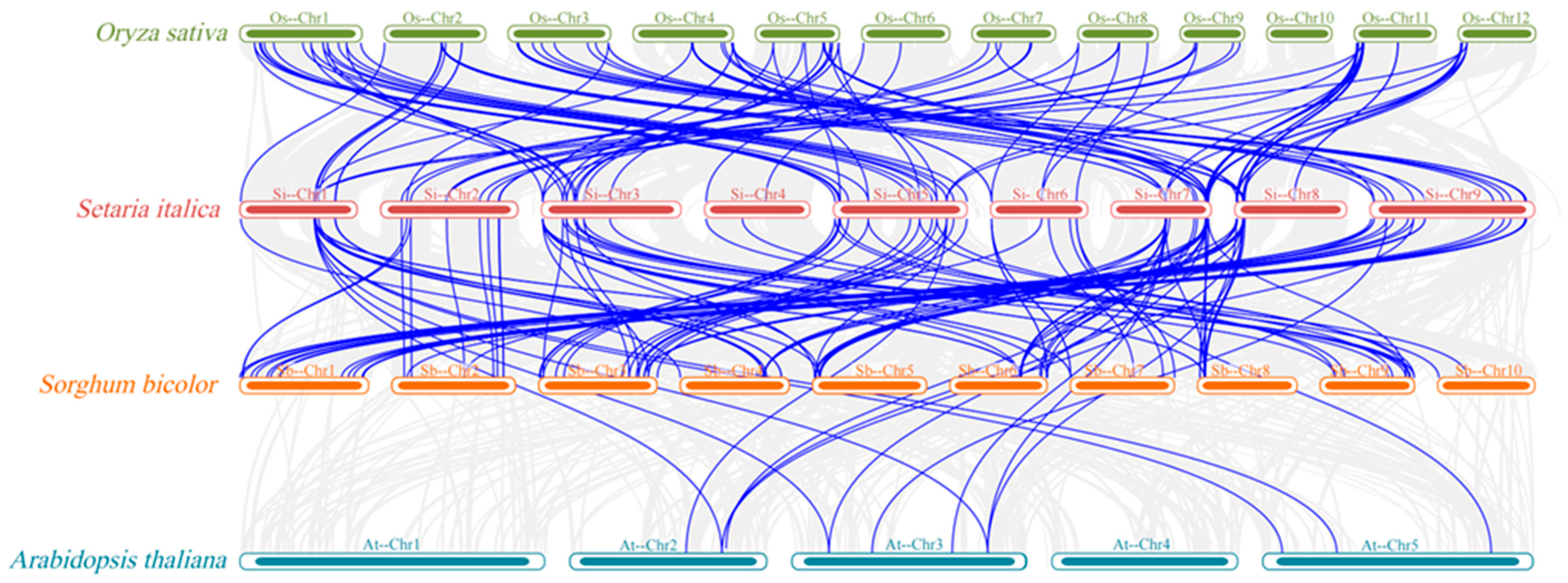
3.5. Synteny Analysis of the Foxtail Millet SiLEA Genes
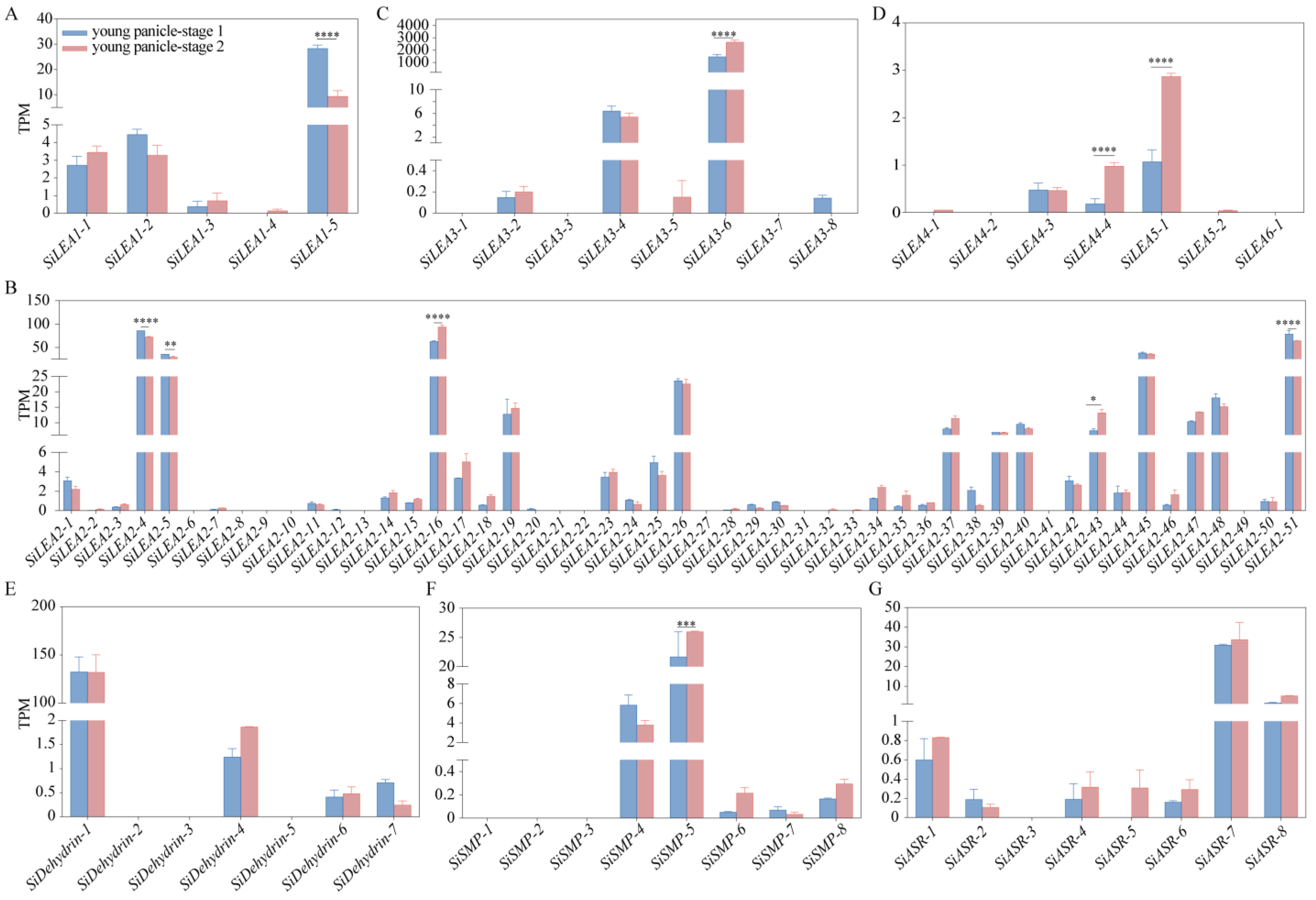
3.6. Tissue-Specific Expression Analysis of SiLEA Genes
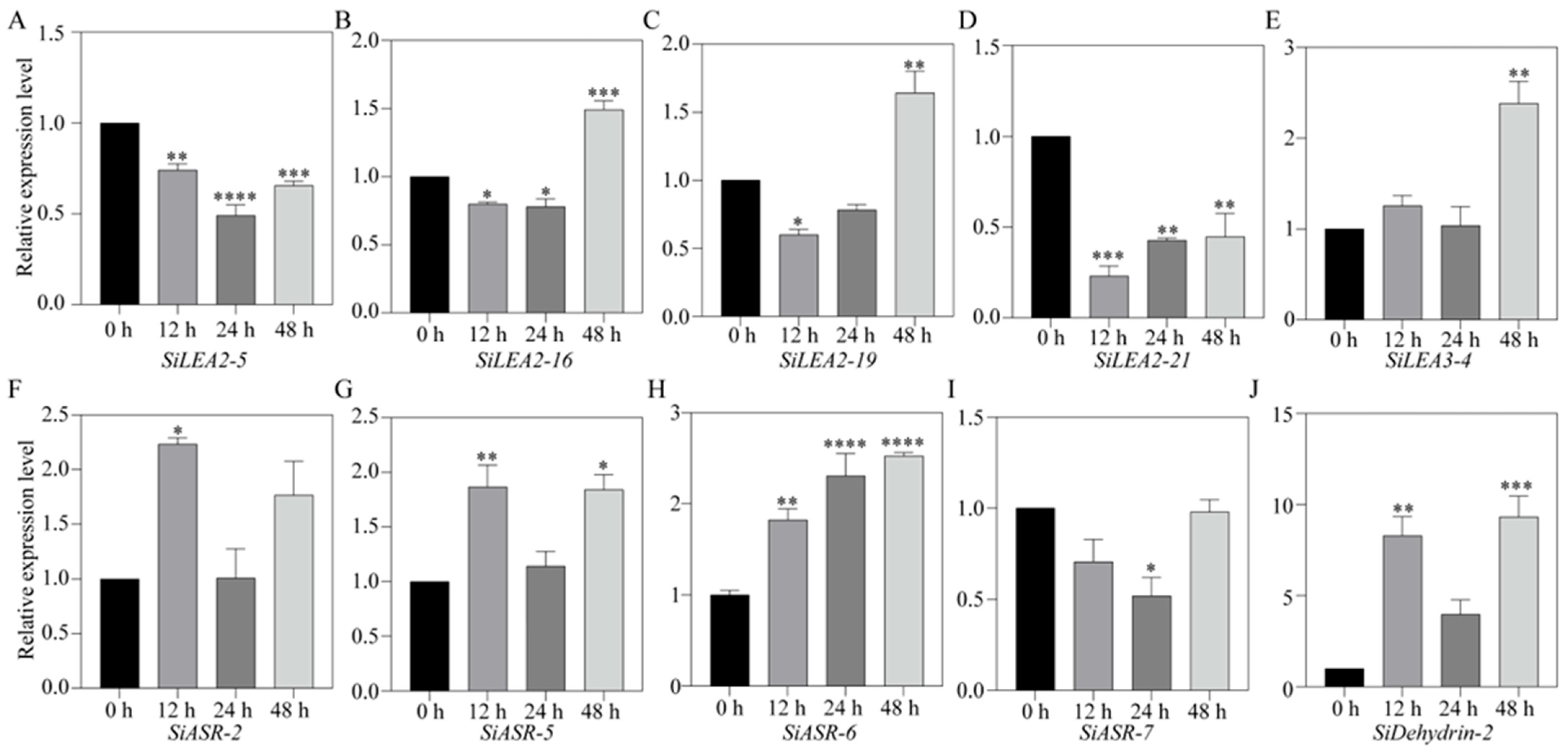
3.7. RT-qPCR Analysis of ABA-Responsive SiLEA Genes in Foxtail Millet
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shinozaki, K.; Yamaguchi-Shinozaki, K. Gene networks involved in drought stress response and tolerance. J. Exp. Bot. 2007, 58, 221–227. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, J.; Li, R.; Ge, Y.; Li, Y.; Li, R. Plants’ response to abiotic stress: Mechanisms and strategies. Int. J. Mol. Sci. 2023, 24, 10915. [Google Scholar] [CrossRef] [PubMed]
- Hundertmark, M.; Hincha, D.K. LEA (late embryogenesis abundant) proteins and their encoding genes in Arabidopsis thaliana. BMC Genom. 2008, 9, 118. [Google Scholar] [CrossRef] [PubMed]
- Bies-Etheve, N.; Gaubier-Comella, P.; Debures, A.; Lasserre, E.; Jobet, E.; Raynal, M.; Cooke, R.; Delseny, M. Inventory, evolution and expression profiling diversity of the LEA (late embryogenesis abundant) protein gene family in Arabidopsis thaliana. Plant Mol. Biol. 2008, 67, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Nagaraju, M.; Kumar, S.A.; Reddy, P.S.; Kumar, A.; Rao, D.M.; Kavi Kishor, P.B. Genome-scale identification, classification, and tissue specific expression analysis of late embryogenesis abundant (LEA) genes under abiotic stress conditions in Sorghum bicolor L. PLoS ONE 2019, 14, e0209980. [Google Scholar] [CrossRef]
- Wang, X.S.; Zhu, H.B.; Jin, G.L.; Liu, H.L.; Wu, W.R.; Zhu, J. Genome-scale identification and analysis of LEA genes in rice (Oryza sativa L.). Plant Sci. 2007, 172, 414–420. [Google Scholar] [CrossRef]
- Liu, D.; Sun, J.; Zhu, D.; Lyu, G.; Zhang, C.; Liu, J.; Wang, H.; Zhang, X.; Gao, D. Genome-wide identification and expression profiles of late embryogenesis-abundant (LEA) genes during grain maturation in wheat (Triticum aestivum L.). Genes 2019, 10, 696. [Google Scholar] [CrossRef]
- Koubaa, S.; Brini, F. Functional analysis of a wheat group 3 late embryogenesis abundant protein (TdLEA3) in Arabidopsis thaliana under abiotic and biotic stresses. Plant Physiol. Biochem. 2020, 156, 396–406. [Google Scholar] [CrossRef]
- Jin, F.; Yu, C.; Lai, L.; Liu, Z.; van der Spoel, D. Ligand clouds around protein clouds: A scenario of ligand binding with intrinsically disordered proteins. PLoS Comput. Biol. 2013, 9, e1003249. [Google Scholar] [CrossRef]
- Dure, L., III; Crouch, M.; Harada, J.; Ho, T.H.; Mundy, J.; Quatrano, R.; Thomas, T.; Sung, Z.R. Common amino acid sequence domains among the LEA proteins of higher plants. Plant Mol. Biol. 1989, 12, 475–486. [Google Scholar] [CrossRef]
- Bray, E.A. Molecular responses to water deficit. Plant Physiol. 1993, 103, 1035–1040. [Google Scholar] [CrossRef]
- Hunault, G.; Jaspard, E. LEAPdb: A database for the late embryogenesis abundant proteins. BMC Genom. 2010, 11, 221. [Google Scholar] [CrossRef]
- Battaglia, M.; Olvera-Carrillo, Y.; Garciarrubio, A.; Campos, F.; Covarrubias, A.A. The enigmatic LEA proteins and other hydrophilins. Plant Physiol. 2008, 148, 6–24. [Google Scholar] [CrossRef]
- Olvera-Carrillo, Y.; Campos, F.; Reyes, J.L.; Garciarrubio, A.; Covarrubias, A.A. Functional analysis of the group 4 late embryogenesis abundant proteins reveals their relevance in the adaptive response during water deficit in Arabidopsis. Plant Physiol. 2010, 154, 373–390. [Google Scholar] [CrossRef]
- Mowla, S.B.; Cuypers, A.; Driscoll, S.P.; Kiddle, G.; Thomson, J.; Foyer, C.H.; Theodoulou, F.L. Yeast complementation reveals a role for an Arabidopsis thaliana late embryogenesis abundant (LEA)-like protein in oxidative stress tolerance. Plant J. 2006, 48, 743–756. [Google Scholar] [CrossRef] [PubMed]
- Jia, F.; Qi, S.; Li, H.; Liu, P.; Li, P.; Wu, C.; Zheng, C.; Huang, J. Overexpression of Late Embryogenesis Abundant 14 enhances Arabidopsis salt stress tolerance. Biochem. Biophys. Res. Commun. 2014, 454, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.H.; Nguyen, C.T.T.; Jung, C.; Cheong, J.J. AtMYB44 suppresses transcription of the late embryogenesis abundant protein gene AtLEA4-5. Biochem. Biophys. Res. Commun. 2019, 511, 931–934. [Google Scholar] [CrossRef] [PubMed]
- Sivamani, E.; Bahieldin, A.; Wraith, J.M.; Al-Niemi, T.; Dyer, W.E.; Ho, T.D.; Qu, R. Improved biomass productivity and water use efficiency under water deficit conditions in transgenic wheat constitutively expressing the barley HVA1 gene. Plant Sci. 2000, 155, 1–9. [Google Scholar] [CrossRef]
- Xu, D.; Duan, X.; Wang, B.; Hong, B.; Ho, T.; Wu, R. Expression of a Late Embryogenesis Abundant Protein gene, HVA1, from barley confers tolerance to water deficit and salt stress in transgenic rice. Plant Physiol. 1996, 110, 249–257. [Google Scholar] [CrossRef]
- Yu, J.; Lai, Y.; Wu, X.; Wu, G.; Guo, C. Overexpression of OsEm1 encoding a group I LEA protein confers enhanced drought tolerance in rice. Biochem. Biophys. Res. Commun. 2016, 478, 703–709. [Google Scholar] [CrossRef]
- Huang, L.; Zhang, M.; Jia, J.; Zhao, X.; Huang, X.; Ji, E.; Ni, L.; Jiang, M. An atypical Late Embryogenesis Abundant Protein OsLEA5 plays a positive role in ABA-induced antioxidant defense in Oryza sativa L. Plant Cell Physiol. 2018, 59, 916–929. [Google Scholar] [CrossRef]
- Wang, G.; Su, H.; Abou-Elwafa, S.F.; Zhang, P.; Cao, L.; Fu, J.; Xie, X.; Ku, L.; Wen, P.; Wang, T.; et al. Functional analysis of a late embryogenesis abundant protein ZmNHL1 in maize under drought stress. J. Plant Physiol. 2023, 280, 153883. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Cao, L.; Ma, C.; Pang, Y.; Ye, F.; Lu, X. Molecular characterization, cloning, and expression analysis of maize ZmLEA34 gene in response to drought stress. J. Shandong Agric. Univ. (Nat. Sci. Ed.) 2022, 53, 665–672. [Google Scholar]
- Wang, M.; Li, P.; Li, C.; Pan, Y.; Jiang, X.; Zhu, D.; Zhao, Q.; Yu, J. SiLEA14, a novel atypical LEA protein, confers abiotic stress resistance in foxtail millet. BMC Plant Biol. 2014, 14, 290. [Google Scholar] [CrossRef]
- Tian, C.; Rehman, A.; Wang, X.; Wang, Z.; Li, H.; Ma, J.; Du, X.; Peng, Z.; He, S. Late embryogenesis abundant gene GhLEA-5 of semi-wild cotton positively regulates salinity tolerance in upland cotton. Gene 2025, 949, 149372. [Google Scholar] [CrossRef]
- Shiraku, M.L.; Magwanga, R.O.; Zhang, Y.; Hou, Y.; Kirungu, J.N.; Mehari, T.G.; Xu, Y.; Wang, Y.; Wang, K.; Cai, X.; et al. Late embryogenesis abundant gene LEA3 (Gh_A08G0694) enhances drought and salt stress tolerance in cotton. Int. J. Biol. Macromol. 2022, 207, 700–714. [Google Scholar] [CrossRef]
- Luo, D.; Wang, C.; Mubeen, S.; Rehman, M.; Cao, S.; Yue, J.; Pan, J.; Jin, G.; Li, R.; Chen, T.; et al. HcLEA113, a late embryogenesis abundant protein gene, positively regulates drought-stress responses in kenaf. Physiol. Plant 2024, 176, e14506. [Google Scholar] [CrossRef]
- Lim, C.W.; Lim, S.; Baek, W.; Lee, S.C. The pepper late embryogenesis abundant protein CaLEA1 acts in regulating abscisic acid signaling, drought and salt stress response. Physiol. Plant. 2015, 154, 526–542. [Google Scholar] [CrossRef]
- Lv, A.; Su, L.; Fan, N.; Wen, W.; Wang, Z.; Zhou, P.; An, Y. Chloroplast-targeted late embryogenesis abundant 1 increases alfalfa tolerance to drought and aluminum. Plant Physiol. 2023, 193, 2750–2767. [Google Scholar] [CrossRef]
- Guo, B.; Zhang, J.; Yang, C.; Dong, L.; Ye, H.; Valliyodan, B.; Nguyen, H.T.; Song, L. The Late Embryogenesis Abundant Proteins in soybean: Identification, expression analysis, and the roles of GmLEA4_19 in drought stress. Int. J. Mol. Sci. 2023, 24, 14834. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, S.; Xu, H.; Tian, H.; Zhang, M.; Zhu, S.; Wang, C.; Hou, J.; Chen, G.; Tang, X.; et al. Identification of the BcLEA gene family and functional analysis of the BcLEA73 gene in Wucai (Brassica campestris L.). Genes 2023, 14, 415. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lu, S.; Jiang, C.; Wang, Y.; Lv, B.; Shen, J.; Ming, F. RcLEA, a late embryogenesis abundant protein gene isolated from Rosa chinensis, confers tolerance to Escherichia coli and Arabidopsis thaliana and stabilizes enzyme activity under diverse stresses. Plant Mol. Biol. 2014, 85, 333–347. [Google Scholar] [CrossRef] [PubMed]
- Boucher, V.; Buitink, J.; Lin, X.; Boudet, J.; Hoekstra, F.A.; Hundertmark, M.; Renard, D.; Leprince, O. MtPM25 is an atypical hydrophobic late embryogenesis-abundant protein that dissociates cold and desiccation-aggregated proteins. Plant Cell Environ. 2010, 33, 418–430. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Hu, Y.; Dong, Y.; Zhang, L.; Wang, B. Abiotic stress-regulated LEA gene mediates the response to drought, salinity, and cold stress in Medicago sativa L. Plant Cell Physiol. 2025, 66, 781–796. [Google Scholar] [CrossRef]
- Wang, W.; Liu, Y.; Kang, Y.; Liu, W.; Li, S.; Wang, Z.; Xia, X.; Chen, X.; Qian, L.; Xiong, X.; et al. Genome-wide characterization of LEA gene family reveals a positive role of BnaA.LEA6.a in freezing tolerance in rapeseed (Brassica napus L.). BMC Plant Biol. 2024, 24, 433. [Google Scholar] [CrossRef]
- Zhou, C.; Niu, S.; El-Kassaby, Y.A.; Li, W. Genome-wide identification of late embryogenesis abundant protein family and their key regulatory network in Pinus tabuliformis cold acclimation. Tree Physiol. 2023, 43, 1964–1985. [Google Scholar] [CrossRef]
- Doust, A.N.; Kellogg, E.A.; Devos, K.M.; Bennetzen, J.L. Foxtail millet: A sequence-driven grass model system. Plant Physiol. 2009, 149, 137–141. [Google Scholar] [CrossRef]
- Yang, Z.; Zhang, H.; Li, X.; Shen, H.; Gao, J.; Hou, S.; Zhang, B.; Mayes, S.; Bennett, M.; Ma, J.; et al. A mini foxtail millet with an Arabidopsis-like life cycle as a C(4) model system. Nat. Plants 2020, 6, 1167–1178. [Google Scholar] [CrossRef]
- Lata, C.; Gupta, S.; Prasad, M. Foxtail millet: A model crop for genetic and genomic studies in bioenergy grasses. Crit. Rev. Biotechnol. 2013, 33, 328–343. [Google Scholar] [CrossRef]
- Li, X.; Hou, S.; Feng, M.; Xia, R.; Li, J.; Tang, S.; Han, Y.; Gao, J.; Wang, X. MDSi: Multi-omics Database for Setaria italica. BMC Plant Biol. 2023, 23, 223. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative Toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Liu, H.; Lyu, H.M.; Zhu, K.; Van de Peer, Y.; Max Cheng, Z.M. The emergence and evolution of intron-poor and intronless genes in intron-rich plant gene families. Plant J. 2021, 105, 1072–1082. [Google Scholar] [CrossRef]
- Hanin, M.; Brini, F.; Ebel, C.; Toda, Y.; Takeda, S.; Masmoudi, K. Plant dehydrins and stress tolerance: Versatile proteins for complex mechanisms. Plant Signal Behav. 2011, 6, 1503–1509. [Google Scholar] [CrossRef]
- Cakir, B.; Agasse, A.; Gaillard, C.; Saumonneau, A.; Delrot, S.; Atanassova, R. A grape ASR protein involved in sugar and abscisic acid signaling. Plant Cell 2003, 15, 2165–2180. [Google Scholar] [CrossRef]
- Wang, C.S.; Liau, Y.E.; Huang, J.C.; Wu, T.D.; Su, C.C.; Lin, C.H. Characterization of a desiccation-related protein in lily pollen during development and stress. Plant Cell Physiol. 1998, 39, 1307–1314. [Google Scholar] [CrossRef]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Genomic Location | Number of Amino Acids | Molecular Weight/KD | pI | GRAVY | Subcellular Localization |
|---|---|---|---|---|---|---|---|
| SiLEA1-1 | Si2g16530 | Chr2:23636372-23636878 (−) | 168 | 16.54 | 9.39 | −0.489 | cyto |
| SiLEA1-2 | Si4g01700 | Chr4:588119-591858 (+) | 125 | 13.49 | 9.84 | −0.889 | mito |
| SiLEA1-3 | Si4g12830 | Chr4:13541862-13542784 (−) | 102 | 11.17 | 8.73 | −1.012 | nucl |
| SiLEA1-4 | Si6g11990 | Chr6:18263649-18264769 (+) | 153 | 15.34 | 9.52 | −0.722 | nucl |
| SiLEA1-5 | Si7g22800 | Chr7:28695111-28695461 (+) | 116 | 11.98 | 9.33 | −0.928 | mito |
| SiLEA2-1 | Si1g01120 | Chr1:336572-337207 (+) | 211 | 21.66 | 10.67 | 0.08 | golg |
| SiLEA2-2 | Si1g19260 | Chr1:26685069-26686200 (−) | 212 | 22.88 | 9.23 | 0.212 | E.R. |
| SiLEA2-3 | Si1g27150 | Chr1:34072139-34073281 (+) | 194 | 21.25 | 9.81 | 0.128 | mito |
| SiLEA2-4 | Si1g27250 | Chr1:34138008-34142334 (−) | 351 | 37.84 | 10.19 | −0.15 | cyto |
| SiLEA2-5 | Si2g08600 | Chr2:7511879-7515924 (−) | 254 | 27.19 | 9.16 | 0.209 | chlo |
| SiLEA2-6 | Si2g10340 | Chr2:9405253-9405924 (−) | 223 | 23.90 | 8.79 | 0.253 | chlo |
| SiLEA2-7 | Si2g10360 | Chr2:9408452-9409066 (−) | 204 | 21.63 | 9.75 | 0.116 | cyto |
| SiLEA2-8 | Si2g11000 | Chr2:10739875-10741954 (−) | 313 | 34.89 | 4.84 | −0.39 | cyto |
| SiLEA2-9 | Si2g16390 | Chr2:23233404-23234406 (+) | 208 | 22.77 | 8.03 | 0.046 | chlo |
| SiLEA2-10 | Si2g22270 | Chr2:31975586-31976874 (+) | 262 | 28.69 | 10.12 | 0.093 | chlo |
| SiLEA2-11 | Si2g28910 | Chr2:38432920-38434042 (−) | 223 | 23.69 | 9.35 | 0.252 | E.R. |
| SiLEA2-12 | Si2g33820 | Chr2:42245927-42247204 (−) | 188 | 19.97 | 9.02 | 0.534 | extr |
| SiLEA2-13 | Si2g33830 | Chr2:42252870-42253532 (−) | 220 | 23.03 | 9.08 | 0.357 | plas |
| SiLEA2-14 | Si2g34050 | Chr2:42443262-42444234 (+) | 202 | 22.73 | 9.08 | −0.071 | chlo |
| SiLEA2-15 | Si3g01780 | Chr3:462543-463202 (+) | 219 | 24.23 | 8.14 | 0.144 | cyto |
| SiLEA2-16 | Si3g01790 | Chr3:466092-466709 (+) | 205 | 22.00 | 9.03 | 0.286 | chlo |
| SiLEA2-17 | Si3g11500 | Chr3:7657823-7660021 (−) | 293 | 30.05 | 10.07 | 0.084 | chlo |
| SiLEA2-18 | Si3g13070 | Chr3:8966951-8968514 (−) | 240 | 24.84 | 10.26 | 0.225 | cyto |
| SiLEA2-19 | Si3g13090 | Chr3:8974973-8975607 (+) | 170 | 18.77 | 5.56 | −0.155 | nucl |
| SiLEA2-20 | Si3g16980 | Chr3:12605559-12606344 (+) | 261 | 27.45 | 8.09 | 0.168 | cyto |
| SiLEA2-21 | Si3g19790 | Chr3:15198175-15199340 (−) | 214 | 22.66 | 10.33 | 0.43 | plas |
| SiLEA2-22 | Si3g28330 | Chr3:27459991-27460596 (−) | 201 | 20.54 | 9.41 | 0.514 | chlo |
| SiLEA2-23 | Si5g01580 | Chr5:514279-515232 (−) | 317 | 33.27 | 10.88 | −0.295 | chlo |
| SiLEA2-24 | Si5g02490 | Chr5:1883173-1883628 (+) | 151 | 16.30 | 6.11 | −0.028 | cyto |
| SiLEA2-25 | Si5g14240 | Chr5:12401284-12403251 (−) | 296 | 30.59 | 9.85 | 0.095 | chlo |
| SiLEA2-26 | Si5g21720 | Chr5:27447618-27448559 (−) | 313 | 32.30 | 11.03 | −0.07 | chlo |
| SiLEA2-27 | Si5g24710 | Chr5:30809909-30810829 (+) | 175 | 19.06 | 5.01 | −0.102 | cyto |
| SiLEA2-28 | Si5g31440 | Chr5:36949347-36950099 (+) | 250 | 26.85 | 5.30 | 0.006 | cyto |
| SiLEA2-29 | Si5g35890 | Chr5:40487066-40488603 (+) | 326 | 35.26 | 9.78 | −0.12 | chlo |
| SiLEA2-30 | Si5g39230 | Chr5:42968053-42969146 (−) | 232 | 25.36 | 9.57 | −0.14 | E.R. |
| SiLEA2-31 | Si5g39250 | Chr5:42971994-42973145 (−) | 241 | 26.70 | 9.49 | −0.26 | chlo |
| SiLEA2-32 | Si6g01320 | Chr6:331589-333238 (−) | 274 | 29.90 | 8.55 | 0.37 | plas |
| SiLEA2-33 | Si6g01330 | Chr6:334758-335402 (−) | 214 | 23.46 | 9.23 | 0.30 | cyto |
| SiLEA2-34 | Si6g19520 | Chr6:30784863-30786086 (−) | 250 | 28.05 | 10.31 | 0.09 | chlo |
| SiLEA2-35 | Si7g10930 | Chr7:19450879-19452721 (−) | 219 | 23.63 | 8.20 | 0.28 | extr |
| SiLEA2-36 | Si7g25560 | Chr7:30769688-30770332 (+) | 214 | 23.72 | 9.68 | −0.06 | chlo |
| SiLEA2-37 | Si7g30030 | Chr7:34042880-34046298 (+) | 209 | 22.95 | 8.28 | 0.137 | chlo |
| SiLEA2-38 | Si7g30060 | Chr7:34053641-34054255 (+) | 204 | 22.23 | 8.76 | 0.038 | chlo |
| SiLEA2-39 | Si7g30680 | Chr7:34528844-34529524 (−) | 226 | 24.16 | 9.43 | 0.123 | chlo |
| SiLEA2-40 | Si7g31120 | Chr7:34820864-34836496 (−) | 641 | 70.62 | 9.68 | −0.299 | E.R. |
| SiLEA2-41 | Si8g01520 | Chr8:523055-523933 (+) | 292 | 31.66 | 10.99 | −0.144 | cyto |
| SiLEA2-42 | Si8g02080 | Chr8:876708-877391 (+) | 227 | 24.16 | 9.43 | 0.176 | chlo |
| SiLEA2-43 | Si8g04550 | Chr8:2883748-2884377 (−) | 209 | 23.15 | 8.93 | 0.125 | chlo |
| SiLEA2-44 | Si9g01150 | Chr9:272503-273129 (−) | 208 | 22.08 | 9.12 | 0.084 | chlo |
| SiLEA2-45 | Si9g02550 | Chr9:1053385-1056778 (+) | 312 | 34.51 | 5.00 | −0.372 | cyto |
| SiLEA2-46 | Si9g02980 | Chr9:1300575-1301207 (−) | 210 | 22.62 | 8.00 | 0.249 | extr |
| SiLEA2-47 | Si9g02990 | Chr9:1322153-1322803 (−) | 216 | 23.28 | 9.10 | 0.133 | chlo |
| SiLEA2-48 | Si9g12880 | Chr9:8342829-8345960 (−) | 266 | 29.27 | 9.30 | −0.006 | cyto |
| SiLEA2-49 | Si9g18470 | Chr9:13483147-13483770 (−) | 207 | 22.10 | 8.97 | 0.263 | cyto |
| SiLEA2-50 | Si9g20420 | Chr9:15461936-15464333 (+) | 274 | 29.66 | 10.16 | −0.111 | E.R. |
| SiLEA2-51 | Si9g44540 | Chr9:50296499-50297353 (+) | 236 | 24.85 | 9.23 | 0.358 | chlo |
| SiLEA3-1 | Si1g20730 | Chr1:28206279-28206890 (+) | 90 | 9.41 | 9.99 | −0.186 | chlo |
| SiLEA3-2 | Si3g20020 | Chr3:15406559-15407221 (−) | 220 | 23.10 | 11.98 | −0.709 | nucl |
| SiLEA3-3 | Si3g26430 | Chr3:23674062-23679744 (+) | 97 | 10.39 | 5.45 | −0.218 | chlo |
| SiLEA3-4 | Si5g03150 | Chr5:2342818-2343221 (+) | 94 | 9.82 | 9.86 | −0.339 | chlo |
| SiLEA3-5 | Si5g03160 | Chr5:2347482-2347907 (+) | 96 | 10.25 | 10.08 | −0.165 | chlo |
| SiLEA3-6 | Si5g06480 | Chr5:5284677-5284979 (+) | 100 | 9.94 | 10.13 | 0 | chlo |
| SiLEA3-7 | Si6g17070 | Chr6:28370327-28370969 (+) | 96 | 10.28 | 9.18 | −0.309 | chlo |
| SiLEA3-8 | Si9g36850 | Chr9:44011390-44011988 (+) | 133 | 14.63 | 9.62 | −0.418 | chlo/mito |
| SiLEA4-1 | Si3g16110 | Chr3:11735612-11736926 (−) | 206 | 21.11 | 8.53 | −1.176 | nucl |
| SiLEA4-2 | Si7g24200 | Chr7:29717629-29719407 (+) | 305 | 32.20 | 9.02 | −0.89 | chlo |
| SiLEA4-3 | Si9g41270 | Chr9:47738247-47740023 (+) | 365 | 38.49 | 7.66 | −0.936 | mito |
| SiLEA4-4 | Si9g51210 | Chr9:55387138-55388377 (+) | 370 | 38.08 | 7.73 | −0.904 | extr |
| SiLEA5-1 | Si3g26920 | Chr3:24540051-24540652 (−) | 153 | 16.42 | 5.74 | −1.380 | nucl |
| SiLEA5-2 | Si3g26930 | Chr3:24544064-24545146 (−) | 93 | 9.80 | 5.48 | −1.280 | nucl |
| SiLEA6-1 | Si2g25470 | Chr2:35395735-35396088 (−) | 117 | 12.32 | 6.41 | −0.925 | nucl |
| SiDehydrin-1 | Si1g27340 | Chr1:34224000-34226135 (−) | 499 | 54.80 | 9.14 | −0.964 | chlo |
| SiDehydrin-2 | Si5g17250 | Chr5:16894582-16895605 (−) | 138 | 14.13 | 9.19 | −1.178 | nucl |
| SiDehydrin-3 | Si5g17260 | Chr5:16946844-16947365 (+) | 138 | 14.10 | 8.90 | −1.192 | nucl |
| SiDehydrin-4 | Si5g29160 | Chr5:35231709-35232485 (+) | 223 | 22.84 | 6.41 | −0.843 | nucl |
| SiDehydrin-5 | Si8g11920 | Chr8:21137495-21138260 (+) | 186 | 19.21 | 9.68 | −1.003 | chlo |
| SiDehydrin-6 | Si8g11930 | Chr8:21187756-21189264 (−) | 347 | 33.74 | 8.99 | −0.804 | nucl |
| SiDehydrin-7 | Si8g11970 | Chr8:21285230-21286503 (−) | 169 | 16.91 | 8.81 | −1.050 | nucl |
| SiSMP-1 | Si1g21980 | Chr1:29302808-29303751 (−) | 167 | 16.16 | 5.15 | −0.139 | cyto |
| SiSMP-2 | Si1g22010 | Chr1:29340219-29340650 (−) | 143 | 14.12 | 5.88 | −0.145 | cyto |
| SiSMP-3 | Si1g22030 | Chr1:29358375-29358806 (−) | 143 | 14.27 | 5.89 | −0.176 | cyto |
| SiSMP-4 | Si4g14860 | Chr4:21785714-21787597 (−) | 288 | 28.98 | 4.63 | −0.307 | cyto |
| SiSMP-5 | Si6g13010 | Chr6:20828940-20830271 (+) | 181 | 17.60 | 4.84 | −0.317 | cyto |
| SiSMP-6 | Si9g09570 | Chr9:5850754-5852099 (+) | 342 | 34.68 | 6.00 | −0.219 | chlo |
| SiSMP-7 | Si9g24110 | Chr9:19527196-19528646 (−) | 263 | 26.38 | 4.64 | −0.419 | nucl |
| SiSMP-8 | Si9g51720 | Chr9:55837685-55838706 (+) | 280 | 27.75 | 4.65 | −0.195 | cyto |
| SiASR-1 | Si1g19510 | Chr1:27087304-27087745 (−) | 107 | 11.82 | 6.76 | −1.19 | mito |
| SiASR-2 | Si5g45980 | Chr5:47661506-47662123 (+) | 173 | 19.38 | 6.26 | −1.277 | mito |
| SiASR-3 | Si5g45990 | Chr5:47665510-47665827 (−) | 105 | 11.71 | 9.73 | −1.147 | nucl |
| SiASR-4 | Si5g46000 | Chr5:47666703-47667492 (+) | 102 | 11.54 | 9.80 | −1.372 | nucl |
| SiASR-5 | Si7g11260 | Chr7:19766534-19766938 (−) | 101 | 11.48 | 6.81 | −1.366 | mito |
| SiASR-6 | Si7g11270 | Chr7:19769468-19771182 (−) | 244 | 26.00 | 5.03 | −1.636 | nucl |
| SiASR-7 | Si7g29600 | Chr7:33748100-33749135 (+) | 138 | 15.45 | 5.88 | −1.30 | mito |
| SiASR-8 | Si8g05130 | Chr8:3530617-3531156 (−) | 137 | 15.42 | 6.15 | −1.35 | mito |
| Homologous Gene | Non-Synonymous Substitution | Synonymous Substitution | Ka/Ks |
|---|---|---|---|
| SiLEA2-2-SiLEA2-35 | 0.25 | 0.57 | 0.44 |
| SiASR-1-SiASR-5 | 0.1 | 0.46 | 0.21 |
| SiLEA2-2-SiLEA2-37 | 0.4 | 0.34 | 1.18 |
| SiLEA2-2-SiLEA2-43 | 0.42 | 0.35 | 1.19 |
| SiLEA1-1-SiLEA1-4 | 0.38 | 0.68 | 0.56 |
| SiLEA2-8-SiLEA2-45 | 0.19 | NaN | NaN |
| SiLEA2-19-SiLEA2-27 | 0.24 | 0.42 | 0.56 |
| SiLEA2-20-SiLEA2-28 | 0.3 | 0.4 | 0.75 |
| SiLEA2-17-SiLEA2-25 | 0.28 | 0.41 | 0.68 |
| SiLEA3-3-SiLEA3-5 | 0.49 | 0.64 | 0.76 |
| SiLEA2-16-SiLEA2-30 | 0.88 | 0.65 | 1.34 |
| SiLEA2-16-SiLEA2-32 | 0.44 | 0.43 | 1.03 |
| SiLEA2-15-SiLEA2-37 | 0.45 | 0.44 | 1.02 |
| SiLEA2-15-SiLEA2-33 | 0.31 | 0.53 | 0.58 |
| SiLEA2-21-SiLEA2-39 | 0.55 | 0.6 | 0.92 |
| SiLEA2-21-SiLEA2-42 | 0.55 | 0.63 | 0.88 |
| SiLEA2-29-SiLEA2-40 | 0.61 | 0.62 | 0.98 |
| SiLEA2-29-SiLEA2-41 | 0.51 | 0.48 | 1.06 |
| SiLEA2-35-SiLEA2-37 | 0.39 | 0.49 | 0.79 |
| SiLEA2-35-SiLEA2-43 | 0.48 | 0.53 | 0.91 |
| SiLEA2-39-SiLEA2-42 | 0.02 | 0.08 | 0.24 |
| SiASR-7-SiASR-8 | 0.11 | 0.24 | 0.47 |
| SiLEA2-37-SiLEA2-43 | 0.18 | 0.43 | 0.42 |
| SiSMP-7-SiSMP-8 | 0.34 | 0.51 | 0.67 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, Y.; Zhao, Y.; Li, X.; Wang, R.; Chang, S.; Zhang, Y.; Ren, X.; Li, H. Genome-Wide Identification and Expression Pattern Analysis of the Late Embryogenesis Abundant (LEA) Family in Foxtail Millet (Setaria italica L.). Genes 2025, 16, 932. https://doi.org/10.3390/genes16080932
Qin Y, Zhao Y, Li X, Wang R, Chang S, Zhang Y, Ren X, Li H. Genome-Wide Identification and Expression Pattern Analysis of the Late Embryogenesis Abundant (LEA) Family in Foxtail Millet (Setaria italica L.). Genes. 2025; 16(8):932. https://doi.org/10.3390/genes16080932
Chicago/Turabian StyleQin, Yingying, Yiru Zhao, Xiaoyu Li, Ruifu Wang, Shuo Chang, Yu Zhang, Xuemei Ren, and Hongying Li. 2025. "Genome-Wide Identification and Expression Pattern Analysis of the Late Embryogenesis Abundant (LEA) Family in Foxtail Millet (Setaria italica L.)" Genes 16, no. 8: 932. https://doi.org/10.3390/genes16080932
APA StyleQin, Y., Zhao, Y., Li, X., Wang, R., Chang, S., Zhang, Y., Ren, X., & Li, H. (2025). Genome-Wide Identification and Expression Pattern Analysis of the Late Embryogenesis Abundant (LEA) Family in Foxtail Millet (Setaria italica L.). Genes, 16(8), 932. https://doi.org/10.3390/genes16080932