Selection and Micropropagation of an Elite Melatonin Rich Tulsi (Ocimum sanctum L.) Germplasm Line

 , ,
, ,  and
and
Abstract
1. Introduction
2. Materials and Methods
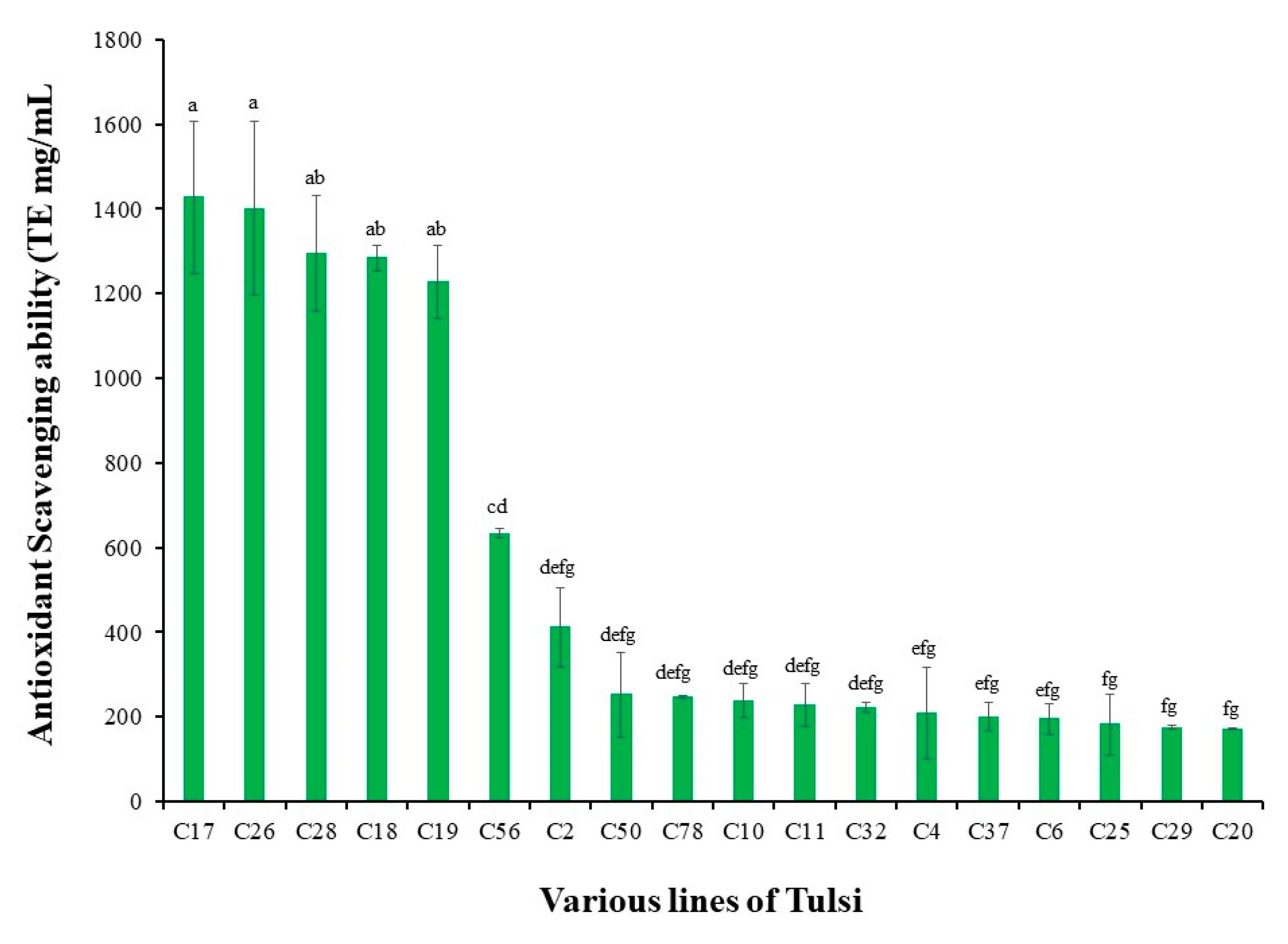
2.1. Plant Materials and Germplasm Line Development
2.2. Determination of Antioxidant Potential Using DPPH Bioassay
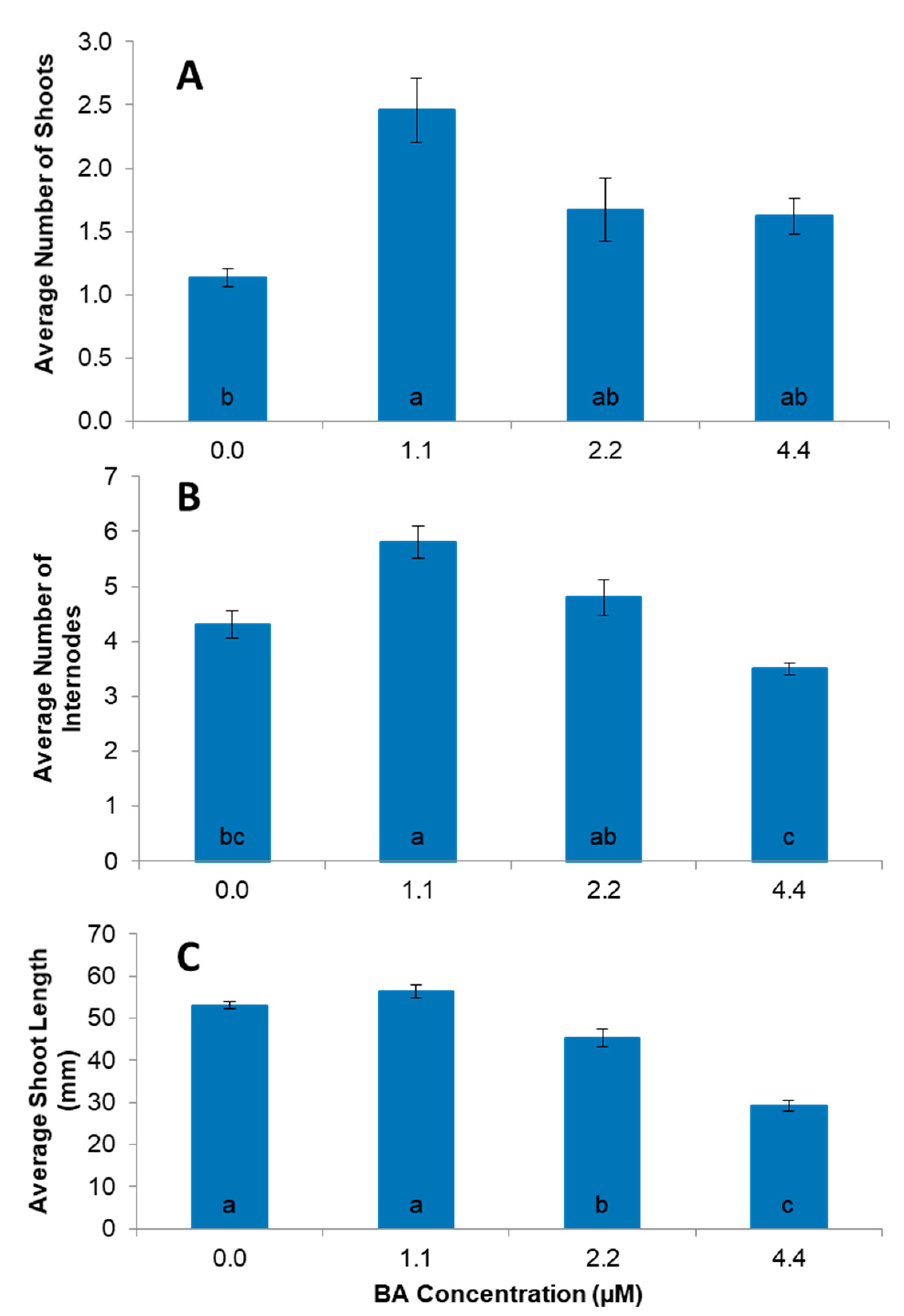
2.3. Culture Establishment and Propagation
2.4. Shoot Multiplication
2.5. Rooting and Acclimatization in the Greenhouse
2.6. Folin-Ciocalteu Phenolic Assay
2.7. Detection and Quantification of Neurotransmitters
2.8. Statistical Analysis
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bano, N.; Ahmed, A.; Tanveer, M.; Khan, G.; Ansari, M. Pharmacological evaluation of Ocimum sanctum. J. Bioequivalence Bioavailab. 2017, 9, 387–392. [Google Scholar] [CrossRef]
- Dafni, A.; Petanidou, T.; Vallianatou, I.; Kozhuharova, E.; Blanche, C.; Pacini, E.; Peyman, M.; Stevanoic, Z.D.; Franchi, G.G.; Benitez, G. Myrtle, basil, rosemary, and three-lobed sage as ritual plants in the monotheistic religions: An Historical-ethnobotanical comparison. Econ. Bot. 2020, 74, 330–355. [Google Scholar] [CrossRef]
- Ved, D.K.; Sureshchandra, S.T.; Barve, V.; Srinivas, V.; Sangeetha, S.; Ravikumar, K.; Kartikeyan, R.; Kulkarni, V.; Kumar, A.S.; Venugopal, S.N.; et al. FRLHT’s ENVIS Centre on Medicinal Plants, Bengaluru. Available online: Envis.frlht.org/frlhtenvis.nic.in (accessed on 8 November 2020).
- Makri, O.; Kintzios, S. Ocimum sp. (Basil): Botany, cultivation, pharmaceutical properties, and biotechnology. J. Herbs Spices Med. Plants 2008, 13, 123–150. [Google Scholar] [CrossRef]
- Duke, J.A. The Garden Pharmacy: Basil as the Holy Hindu Highness. Altern. Complement. Ther. 2008, 14, 5–8. [Google Scholar] [CrossRef]
- Zahran, E.M.; Abdelmohsen, U.R.; Khalil, H.E.; Desoukey, S.Y.; Fouad, M.A.; Kamel, M.S. Diversity, phytochemical and medicinal potential of the genus Ocimum L. (Lamiaceae). Phytochem Rev. 2020, 19, 907–953. [Google Scholar] [CrossRef]
- Engels, G.; Brinckmann, J. Holy Basil. In HerbalGram; American Botanical Council: Austin, TX, USA, 2013; pp. 1–6. [Google Scholar]
- Singh, D.; Chaudhuri, P.K. A review on phytochemical and pharmacological properties of Holy basil (Ocimum sanctum L.). Ind Crop. Prod. 2018, 118, 367–382. [Google Scholar] [CrossRef]
- Maharjan, S. Ocimum sanctum (Linn.); The queen of herbs. Eur. J. Biomed. Pharm. Sci. 2019, 6, 106–109. [Google Scholar]
- Pandey, V.K.; Upadhyay, S.N.; Mishra, P.K. Light-induced synthesis of silver nanoparticles using Ocimum tenuiflorum extract: Characterisation and application. J. Chem Res. 2020, 174751982093651. [Google Scholar] [CrossRef]
- Cohen, M.M. Tulsi—Ocimum sanctum: A herb for all reasons. J. Ayurveda Integr. Med. 2014, 5, 251–259. [Google Scholar] [CrossRef]
- Pathania, M.; Bhardwaj, P.; Pathania, N.; Rathaur, V.; Amisha. A review on exploring evidence-based approach to harnessing the immune system in times of corona virus pandemic: Best of modern and traditional Indian system of medicine. J. Fam Med. Prim. Care 2020, 9, 3826. [Google Scholar] [CrossRef]
- Singh, N.K.; Sehgal, C.B. Micropropagation of ‘Holy Basil’ (Ocimum sanctum Linn.) from young inflorescences of mature plants. Plant Growth Regul. 1999, 29, 161–166. [Google Scholar] [CrossRef]
- Shilpa, K.; Selvakkumar, C.; Senthil, A.K.; Lakshmi, B.S. In vitro root culture of Ocimum sanctum L. and evaluation of its free radical scavenging activity. Plant Cell Tissue Organ Cult. 2010, 101, 105–109. [Google Scholar] [CrossRef]
- Banu, L.; Bari, M. Protocol establishment for multiplication and regeneration of Ocimum sanctum Linn. an important medicinal plant with high religious value in Bangladesh. J. Plant Sci. 2007, 2, 530–537. [Google Scholar] [CrossRef][Green Version]
- Lim, Z.; Ling, A.; Hussein, S. Callus Induction of Ocimum sanctum and estimation of its total flavonoids Content. Asian J. Agric. Sci. 2009, 1, 55–61. [Google Scholar]
- Mishra, T. Protocol establishment for multiplication and regeneration of ‘Holy Basil’ (Ocimum sanctum Linn). An important medicinal plant with high religious value in India. J. Med. Plants Stud. 2015, 3, 16–19. [Google Scholar]
- Mandal, J.; Pattnaik, S.; Chand, P.K. Alginate encapsulation of axillary buds of Ocimum americanum L. (hoary basil), O. basilicum L. (sweet basil), O. gratissimum L. (shrubby basil), and O. sanctum. L. (sacred basil). Vitr. Cell Dev. Biol. Plant 2000, 36, 287–292. [Google Scholar] [CrossRef]
- Tyub, S.; Kamili, A. Enhanced axillary shoot proliferation in Ocimum sanctum Linn. via shoot tip culture using various concentrations of BAP. J. Res. Dev. 2008, 8, 80–85. [Google Scholar]
- Gogoi, K.; Kumara, S. Callus-Mediated plantlet regeneration of Ocimum tenuiflorum L. using axillary buds as explants. Plant Sci. 2011, v.2, 1–5. [Google Scholar]
- Girija, S.; Kavitha, S.; Deepavathi, S. Direct multiple shoot regeneration from shoot tip and nodal explants of Ocimum sanctum L. (Tulsi): A medicinal herb. Plant Cell Biotechnol. 2006, 7, 23–28. [Google Scholar]
- Aggarwal, D.; Neeti, N.; Reddy, M.S.; Kumar, A. Shoot organogenesis and assessment of clonal fidelity of regenerated plants of Ocimum tenuiflorum L.: Queen of Herbs. Vegetos 2020, 33, 420–429. [Google Scholar] [CrossRef]
- Bhuvaneshwari, K.; Gokulanathan, A.; Jayanthi, M.; Govindasamy, V.; Milella, L.; Lee, S.; Yang, D.C.; Girija, S. Can Ocimum basilicum L. and Ocimum tenuiflorum L. in vitro culture be a potential source of secondary metabolites? Food Chem. 2016, 194, 55–60. [Google Scholar] [CrossRef]
- Rebollo-Hernanz, M.; Aguilera, Y.; Herrera, T.; Cayuelas, L.T.; Dueñas, M.; Rodríguez-Rodríguez, P.; Ramiro-Cortijo, D.; Arribas, S.M.; Martín-Cabrejas, M.A. Bioavailability of melatonin from lentil sprouts and its role in the plasmatic antioxidant status in rats. Foods 2020, 9, 330. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.-X.; Reiter, R.J.; Manchester, L.C.; Yan, M.; El-Sawi, M.; Sainz, R.M.; Mayo, J.C.; Kohen, R.; Allegra, M.; Hardeland, R. Chemical and physical properties and potential mechanisms: Melatonin as a broad spectrum antioxidant and free radical scavenger. Curr. Top. Med. Chem. 2002, 2, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Naranjo, M.I.; Moya, M.L.; Cantos-Villar, E.; Garcia-Parrilla, M.C. Comparative evaluation of the antioxidant activity of melatonin and related indoles. J. Food Compos. Anal. 2012, 28, 16–22. [Google Scholar] [CrossRef]
- Bajwa, V.S.; Shukla, M.R.; Sherif, S.M.; Murch, S.J.; Saxena, P.K. Identification and characterization of serotonin as an anti-browning compound of apple and pear. Postharvest Biol. Technol. 2015, 110, 183–189. [Google Scholar] [CrossRef]
- Erland, L.A.E.; Murch, S.J.; Reiter, R.J.; Saxena, P.K. A new balancing act: The many roles of melatonin and serotonin in plant growth and development. Plant Signal. Behav. 2015, 10, e1096469-15. [Google Scholar] [CrossRef]
- Erland, L.A.E.; Saxena, P.K. Melatonin in morphogenesis. Vitr. Cell Dev. Biol. Plant 2018, 54, 3–24. [Google Scholar] [CrossRef]
- Erland, L.A.E.; Turi, C.E.; Saxena, P.K. Serotonin: An ancient molecule and an important regulator of plant processes. Biotechnol. Adv. 2016, 8, 1347–1361. [Google Scholar] [CrossRef]
- Basheer, M.; Rai, S.; Hsu, T.-C. Melatonin vs. phytomelatonin: Therapeutic uses with special reference to polycystic ovarian syndrome (PCOS). Cogent. Biol. 2016, 2, 1136257. [Google Scholar] [CrossRef]
- Lanfumey, L.; Mongeau, R.; Hamon, M. Biological rhythms and melatonin in mood disorders and their treatments. Pharm. Ther. 2013, 138, 176–184. [Google Scholar] [CrossRef]
- Reiter, R.J.; Abreu-Gonzalez, P.; Marik, P.E.; Dominguez-Rodriguez, A. Therapeutic algorithm for use of melatonin in patients with COVID-19. Front. Med. 2020, 7, 226. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.X.; Hardeland, R. Potential utility of melatonin in deadly infectious diseases related to the overreaction of innate immune response and destructive inflammation: Focus on COVID-19. Melatonin Res. 2020, 3, 120–143. [Google Scholar] [CrossRef]
- Anderson, G.; Reiter, R.J. Melatonin: Roles in influenza, Covid-19, and other viral infections. Rev. Med. Virol. 2020, 30, e2109. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Llamas, F.; Hernández-Ruiz, J.; Cuesta, A.; Zamora, S.; Arnao, M.B. Development of a phytomelatonin-rich extract from cultured plants with excellent biochemical and functional properties as an alternative to synthetic melatonin. Antioxidants 2020, 9, 158. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.; Hernández-Ruiz, J. The potential of phytomelatonin as a nutraceutical. Molecules 2018, 23, 238. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Fukumoto, L.R.; Mazza, G. Assessing antioxidant and prooxidant activities of phenolic compounds. J. Agric. Food Chem. 2000, 48, 3597–3604. [Google Scholar] [CrossRef]
- Herald, T.J.; Gadgil, P.; Tilley, M. High-throughput micro plate assays for screening flavonoid content and DPPH-scavenging activity in sorghum bran and flour. J. Sci. Food Agric. 2012, 92, 2326–2331. [Google Scholar] [CrossRef]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. Methods Enzym. 1999, 299, 152–178. [Google Scholar] [CrossRef]
- Saremba, B.M.; Tymm, F.J.M.; Baethke, K.; Rheault, M.R.; Sherif, S.M.; Saxena, P.K.; Murch, S.J. Plant signals during beetle (Scolytus multistriatus) feeding in American elm Ulmus americana Planch). Plant Signal. Behav. 2017, 12, e1296997. [Google Scholar] [CrossRef]
- Beilby, M.J.; Turi, C.E.; Baker, T.C.; Tymm, F.J.M.; Murch, S.J. Circadian changes in endogenous concentrations of indole-3-acetic acid, melatonin, serotonin, abscisic acid and jasmonic acid in Characeae (Chara australis Brown). Plant Signal. Behav. 2015, 10, e1082697. [Google Scholar] [CrossRef] [PubMed]
- Ulewicz-Magulska, B.; Wesolowski, M. Total phenolic contents and antioxidant potential of herbs used for medical and culinary purposes. Plant Food Hum. Nutr. 2019, 74, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Khan, M.M.; Raza, S.S.; Javed, H.; Ashafaq, M.; Islam, F.; Safhi, M.M.; Islam, F. Ocimum sanctum attenuates oxidative damage and neurological deficits following focal cerebral ischemia/reperfusion injury in rats. Neurol. Sci. 2012, 33, 1239–1247. [Google Scholar] [CrossRef] [PubMed]
- Zobayed, S.M.A.; Saxena, P.K. In vitro-grown roots: A superior explant for prolific shoot regeneration of St. John’s wort (Hypericum perforatum L. cv ‘New Stem’) in a temporary immersion bioreactor. Plant Sci. 2003, 165, 463–470. [Google Scholar] [CrossRef]
- Liu, C.Z.; Murch, S.J.; El-Demerdash, M.; Saxena, P.K. Artemisia judaica L.: Micropropagation and antioxidant activity. J. Biotechnol. 2004, 110, 63–71. [Google Scholar] [CrossRef]
- Murch, S.J.; Liu, C.; Romero, R.M.; Saxena, P.K. In vitro culture and temporary immersion bioreactor production of Crescentia cujete. Plant Cell Tissue Org. Cult. 2004, 78, 63–68. [Google Scholar] [CrossRef]
- Murch, S.J.; Saxena, P.K. St. John’s wort (Hypericum perforatum L.): Challenges and strategies for production of chemically-consistent plants. Can. J. Plant Sci. 2006, 86, 765–771. [Google Scholar] [CrossRef]
- Pattnaik, S.; Chand, P.K. In vitro propagation of the medicinal herbs Ocimum americanum L. syn. O. canum Sims. (hoary basil) and Ocimum sanctum L. (holy basil). Plant Cell Rep. 1996, 15, 846–850. [Google Scholar] [CrossRef]
- Cole, I.B.; Saxena, P.K.; Murch, S.J. Medicinal biotechnology in the genus scutellaria. Vitr. Cell Dev. Biol. Plant 2007, 43, 318–327. [Google Scholar] [CrossRef]
- Jamal, M.A.H.M.; Sharif, I.H.; Shakil, M.M.; Rahman, A.N.M.R.-B.; Banu, N.A.; Islam, M.R.; Nazmuzzaman, M. In vitro regeneration of a common medicinal plant, Ocimum sanctum L. for mass propagation. Afr. J. Biotechnol. 2016, 15, 1269–1275. [Google Scholar] [CrossRef]
- Sahoo, Y.; Pattnaik, S.K.; Chand, P.K. In vitro clonal propagation of an aromatic medicinal herb Ocimum basilicum L. (sweet basil) by axillary shoot proliferation. Vitr. Cell Dev. Biol. Plant 1997, 33, 293–296. [Google Scholar] [CrossRef]
- Mao, A.A.; Wetten, A.; Fay, M.; Caligari, P.D.S. In vitro propagation of Clerodendrum colebrookianum Walp., a potential natural anti-hypertension medicinal plant. Plant Cell Rep. 1995, 14, 493–496. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Tiwari, K.N. In vitro plant regeneration from decapitated embryonic axes of Clitoria ternatea L.—An important medicinal plant. Ind. Crop. Prod. 2012, 35, 224–229. [Google Scholar] [CrossRef]
- Gaspar, T.; Kevers, C.; Penel, C.; Greppin, H.; Reid, D.M.; Thorpe, T.A. Plant hormones and plant growth regulators in plant tissue culture. Vitr. Cell Dev. Biol. Plant 1996, 32, 272–289. [Google Scholar] [CrossRef]
- Shukla, M.R.; Jones, A.M.P.; Sullivan, J.A.; Liu, C.; Gosling, S.; Saxena, P.K. In vitro conservation of American elm (Ulmus americana): Potential role of auxin metabolism in sustained plant proliferation. Can. J. For. Res. 2012, 42, 686–697. [Google Scholar] [CrossRef]
- Thomas, T.D. The role of activated charcoal in plant tissue culture. Biotechnol. Adv. 2008, 26, 618–631. [Google Scholar] [CrossRef]
- Pan, M.J.; Staden, J. van. The use of charcoal in in vitro culture–A review. Plant Growth Regul. 1998, 26, 155–163. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin: A new plant hormone and/or a plant master regulator? Trend Plant Sci. 2019, 24, 38–48. [Google Scholar] [CrossRef]
- Reiter, R.; Tan, D.-X.; Zhou, Z.; Cruz, M.; Fuentes-Broto, L.; Galano, A. Phytomelatonin: Assisting plants to survive and thrive. Molecules 2015, 20, 7396–7437. [Google Scholar] [CrossRef]
- Erland, L.A.E.; Shukla, M.R.; Singh, A.S.; Murch, S.J.; Saxena, P.K. Melatonin and serotonin: Mediators in the symphony of plant morphogenesis. J. Pineal Res. 2018, 64, e12452. [Google Scholar] [CrossRef]
- Bahcesular, B.; Yildirim, E.D.; Karaçocuk, M.; Kulak, M.; Karaman, S. Seed priming with melatonin effects on growth, essential oil compounds and antioxidant activity of basil (Ocimum basilicum L.) under salinity stress. Ind. Crop. Prod. 2020, 146, 112165. [Google Scholar] [CrossRef]
- Nazir, M.; Ullah, M.A.; Mumtaz, S.; Siddiquah, A.; Shah, M.; Drouet, S.; Hano, C.; Abbasi, B.H. Interactive effect of melatonin and UV-C on phenylpropanoid metabolite production and antioxidant potential in callus cultures of purple basil (Ocimum basilicum L. var purpurascens). Molecules 2020, 25, 1072. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; Saxena, P.K. A melatonin-rich germplasm line of St John’s wort (Hypericum perforatum L.). J. Pineal Res. 2006, 41, 284–287. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Transition | Cone Voltage | Collision Voltage |
|---|---|---|---|
| Serotonin | 177 > 160 | 45 | 10 |
| 177 > 115 | 27 | ||
| Melatonin | 233 > 159 | 30 | 23 |
| 233 > 174 | 15 |
| Source | Melatonin (ng/g) | Serotonin (ng/g) |
|---|---|---|
| Wildtype | 42.8 ± 4.95 b | 1684.7 ± 82.04 a |
| Vrinda | 66.4 ± 16.5 a | 1305.1 ± 139.43 b |
| Source/Plant Parts | Leaf | Root | ||
|---|---|---|---|---|
| MEL (ng/g) | SER (ng/g) | MEL (ng/g) | SER (ng/g) | |
| Field | 327.17 a | 685.71 a | 365.05 a | 605.26 a |
| Greenhouse | 341.83 a | 497.95 a | 378.64 a | 501.79 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shukla, M.R.; Kibler, A.; Turi, C.E.; Erland, L.A.E.; Sullivan, J.A.; Murch, S.J.; Saxena, P.K. Selection and Micropropagation of an Elite Melatonin Rich Tulsi (Ocimum sanctum L.) Germplasm Line. Agronomy 2021, 11, 207. https://doi.org/10.3390/agronomy11020207
Shukla MR, Kibler A, Turi CE, Erland LAE, Sullivan JA, Murch SJ, Saxena PK. Selection and Micropropagation of an Elite Melatonin Rich Tulsi (Ocimum sanctum L.) Germplasm Line. Agronomy. 2021; 11(2):207. https://doi.org/10.3390/agronomy11020207
Chicago/Turabian StyleShukla, Mukund R., Annaliese Kibler, Christina E. Turi, Lauren A. E. Erland, J. Alan Sullivan, Susan J. Murch, and Praveen K. Saxena. 2021. "Selection and Micropropagation of an Elite Melatonin Rich Tulsi (Ocimum sanctum L.) Germplasm Line" Agronomy 11, no. 2: 207. https://doi.org/10.3390/agronomy11020207
APA StyleShukla, M. R., Kibler, A., Turi, C. E., Erland, L. A. E., Sullivan, J. A., Murch, S. J., & Saxena, P. K. (2021). Selection and Micropropagation of an Elite Melatonin Rich Tulsi (Ocimum sanctum L.) Germplasm Line. Agronomy, 11(2), 207. https://doi.org/10.3390/agronomy11020207