Abstract
In wheat, anthocyanin pigments can be accumulated in pericarp tissues (under control of the Pp genes) resulting in purple-colored grain. In the current study, a strategy, based on the use of molecular and morphological markers, was applied to create purple-grained bread wheat cultivars adapted to the West Siberian region. The breeding scheme started from crossing of recipients (elite cultivars and lines) with donor lines carrying dominant alleles of the complementary genes Pp3 and Pp-D1. The F2 hybrids passed three-step marker-assisted selection, and those having dominant Pp-D1Pp-D1Pp3Pp3 genotypes were backcrossed with the recurrent parents. The desired BC1F2-3 progenies were selected using morphological marker, while BC1F3 also passed through field evaluation. At this stage, 120 lines were selected and planted in individual 1 m2 “breeding nursery (BN) plots” for assessment of heading dates, duration of vegetation period, resistance to powdery mildew, stem and leaf rusts, protein and gluten content, as well as productivity. After these investigations, a total of 17 promising anthocyanin-rich purple-grained lines characterized by multiple resistance and having best yield/quality characteristics were finally candidates for selection of commercial cultivars adapted to the West Siberian climate and suitable for functional food production.
1. Introduction
In the late 1980s, a new concept in nutritional science, known as functional foods, emerged in Japan and spread over the world. The main idea was improving health benefits of foods to prevent a wide range of human diseases [1]. Research programs were initiated to determine nutritional value of food ingredients. Furthermore, breeding programs were developed to improve crop cultivars for producing functional foods. Special attention was paid to wheat as a main staple food in many countries. One promising direction for improving wheat is the use of breeding blue- and purple-grained cultivars with an increased content of anthocyanins, which are biologically active substances known for both plant protection against severe environments and positive effect on human health [2,3,4]. They possess antioxidant, anti-inflammatory, hypoglycemic, antimutagenic, and neuroprotective activities; they can prevent cardiovascular disease, metabolic syndrome, different types of cancer, as well as they are beneficial for eye vision [5,6]. For example, in vitro studies have demonstrated that wheat or rye bread fortified with anthocyanin-rich berries caused a delay in the glycemic response after eating and significantly improved the glycemic profile of bread, which could help to prevent type 2 diabetes [7]. A study of the effects of anthocyanin-rich wheat grain diet on working memory in mouse models of Alzheimer’s and Parkinson’s diseases showed that cereal diets, independent of pigmentation, prevented the development of disturbances in working memory, which, in the case of Parkinson’s, was even improved by a diet with anthocyanin’s [8]. The anthocyanin-rich cereal diet demonstrated its effectiveness for preventing the development of tumors and metastasis in a mouse model of Lewis lung carcinoma [9]. After consumption for eight weeks, of whole purple wheat bars by obese adults with chronic inflammation, reduced concentrations of plasma marker IL-6 and fasting glucose were observed [10].
In wheats, the purple color of grains caused by anthocyanins accumulating in pericarp tissues was discovered, for the first time, in tetraploid wheat species T. aethiopicum Jakubz. from Abyssinia (now Ethiopia), in the 1870s [11]. T. aethiopicum originated from the Ethiopian emmer wheat, and then distributed throughout Ethiopia, Eritrea, Yemen, where, for a long time, it was considered to be an endemic species [12]. Currently, purple-grained tetraploid wheat is still cultivated in mountain regions of Ethiopia, where it demonstrates earlier maturity; shorter height; and higher fertility, tillering capacity, and harvest index than the white- or red-grained wheats. Purple-grained wheat is used for preparation of Arekie, a local distilled spirituous liquor consumed as a warming drink in the cool and wet highlands and as a traditional medicine against stomach disorders [13]. According to Belay et al. [13], some other tetraploid wheats (T. polonicum, T. carthlicum, and T. durum) which are found in Ethiopia may possess the purple pericarp color.
With the idea of using purple grain color to clearly mark feed-type common wheat from bread-making types, the first purple-grained hexaploid wheat was obtained by crossing the tetraploid wheat E0450 that originated from Abyssinia and the hexaploid wheat cultivar Arawa [14]. In 1977, winter wheat Charcoal was released in United States [15]. In 1986, the purple wheat lines from New Zealand were used to develop wheat cultivars for animal feed and for bioethanol production in Western Canada [16]. First reports on breeding purple-grained cultivars for human nutrition refer to cv Konini that was released for wholemeal bread-making, in 1981, in New Zealand [17]. Nowadays beneficial commercial cultivars of wheat with increased anthocyanin content have been released in Canada, China, Japan, and several European countries. The Austrian company Backaldrin is one of the main world leaders in the production of ingredients for the bakery and confectionery industry, has registered new commercial products from purple wheat under the brand “PurPur” (The Kornspitz Company, Asten, Austria) [18]. Whole flour, bran, and flakes from Canadian-bred purple wheat have been produced under the registered trademark AnthoGrainTM (InfraReady Products Ltd., Saskatoon, SK, Canada) [19]. Purple wheat grains have been evaluated as a novel brewing material [20].
The high anthocyanin content in purple grains enriches traditional bakery products with new nutritional properties [21]. Although some degradation of anthocyanins takes place during processing [22], they have been demonstrated in end-use bakery products, i.e., wholegrain bread [23], biscuits [24,25], pasta [26], pancakes, porridge, crackers, and bars [27]. The consumption of assimilable anthocyanins, up to 1.03 mg or 0.83 mg per 100 g, has been reported for wholegrain bread or biscuits baked from purple wheat grain, respectively [23,25]. No deterioration of bread-making quality by anthocyanins was detected [23,28,29]. Biscuits prepared from whole-wheat purple grains had higher antioxidant capacity, and lower oxidative degradation of lipids than common non-pigmented biscuits [24]. Ficco et al. [26] described that the quality of whole-grained pasta from the purple hard wheat T. durum was not significantly different from commercial durum. Furthermore, the presence of anthocyanins in grain increased the shelf life of the bakery products and their resistance to the development of fungi in provocative conditions [23].
The anthocyanins biosynthesis in wheat has been well studied and fully reviewed in [30]. The two complementary genes, Pp3 and Pp-1, mapped to chromosomes 2A and homoeologous group of chromosomes 7, respectively, are responsible for activation of the anthocyanin biosynthesis pathway in wheat pericarp. Molecular functions of the genes have been recently revealed. The TaMyc1 gene, encoding for regulatory factor with bHLH domain, was suggested as a gene candidate for Pp3 [31]. The functional difference of dominant and recessive alleles of this gene is due to the copy number variation of tandem repeat sequence in its promoter [32]. The complementary gene Pp-1, required for activation of anthocyanin biosynthesis having pleiotropic effect on pigmentation of other parts of wheat plant, encodes the regulatory factor from the R2R3-MYB family [33,34].
Using molecular-genetic mechanisms underlying the phenotype of the purple-grained wheat, we developed a strategy for accelerated breeding of anthocyanin-rich bread wheat cultivars. Here, we describe the strategy and demonstrate its successful application for development of purple-grained breeding lines of spring bread wheat, adapted to conditions of the West Siberian region, resistant to a wide range of unfavorable biotic stresses and abiotic environmental factors, and finally, suitable for modern responsible farming.
2. Materials and Methods
2.1. Plant Material and Breeding Strategy
To get anthocyanin-rich cultivars adapted to the West Siberian region we used marker-assisted transfer of dominant alleles of the regulatory genes Pp3 and Pp-D1 from donor lines to elite cultivars or breeding lines having potential for production in this region. MAS was applied twice for selection of genotypes Pp-D1Pp-D1Pp3Pp3 as follows: (1) in F2 (by using molecular and morphological marker) for immediate crossing with a recipient line and (2) in BC1F2/F3 (by morphological markers only) (Figure 1).
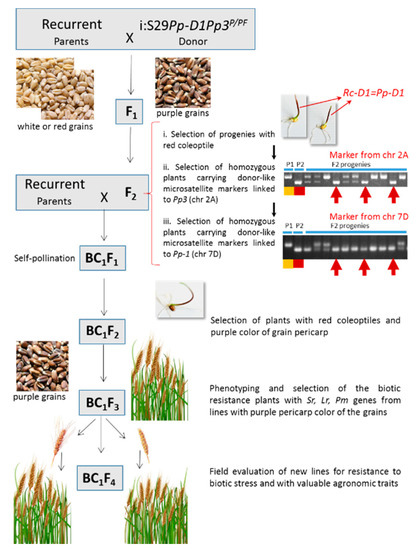
Figure 1.
Breeding strategy for development of anthocyanin-rich cultivars of bread wheat (Triticum aestivum L.) based on the elite spring wheat cultivars and promising breeding lines. The near-isogenic line i:S29PpD1Pp3PF with cv Purple Feed (PF) introgressions in chromosomes 2A and 7D, carrying, respectively, the Pp3 and Pp-D1 genes, responsible for purple pigmentation of grain was used as a donor. The three-step selection of the homozygous F2 plants for the target genes based on the coleoptile pigmentation assessment (i), donor-like homozygous SSR-loci linked to the Pp3 (ii), and Pp-D1 (iii) genes was carried out. The selected plants underwent backcrossing with the recurrent parents. The BC1F2 generation with dark-red coleoptiles were self-pollinated to get BC1F3 seeds that were collected separately from individual plants and the resulted families were evaluated in the field conditions. The individual plant from the families without segregation of the pericarp color with resistance to mildew and rusts diseases were selected, and then their grain quality and agronomic traits were characterized in BC1F4.
A description of the parental recipient materials is given in Table 1. Near-isogenic lines, i:S29Pp-1DPp3PF (abbreviation—i:S29PF) and i:S29Pp-1DPp3P (i:S29P), inherited dark-red coleoptile color and purple pericarp color from their donor spring breeding lines, Purple Feed (k-49426, Canada) and Purple (k-46990, Australia), respectively, and were used as donors of the anthocyanin grain pigmentation. The lines were both created in the genetics background of red-grained spring bread wheat cultivar Saratovskaya 29 (S29) having uncolored pericarp. Derivatives carrying both complementary Pp-D1 and Pp3 were created by [35]; the introgressions of purple-grained donors’ chromatin were mapped to the short arm of chromosome 7D and the long arm of chromosome 2A, respectively (Figure A1) [36].

Table 1.
The recurrent parental cultivars and lines of Triticum aestivum L.
2.2. Molecular Markers
Gatersleben wheat microsatellite (GWM) markers [39,40] flanking target genes Pp3 (on chromosome 2A) and Pp-D1 (on chromosome 7D) (Figure A1 and Table 2), shown previously to be efficient in MAS-based development of wheat lines with desired Pp alleles [41], were used for screenings. DNA was extracted from young leaves of the parental and individual progeny plants, according to a procedure described by Plaschke et al. [42]. The conditions of PCR are described in Röder et al. [39]. Amplicons were separated through either 2% or 5% ACTGene agarose gels (ACTGene, Inc., Piscataway, NJ, USA).
Table 2.
The GWM markers polymorphic among donor lines i:S29Pp-D1Pp3PF, i:S29Pp-D1Pp3P, and recipient cultivars used for marker-assisted breeding.
2.3. Morphological Markers
Before planting F2 and BC1F2 plants, the coleoptile color was assessed by sowing seeds in Petri dishes and growing them for one week on filter paper at 20 °C under a 12 h photoperiod in a “Rubarth Apparate” growth chamber (RUMED GmbH, Laatzen, Germany). Dark-red coleoptile color was used as a morphological marker for Pp-D1 (according to Gordeeva et al., 2015 [41]). To select homozygous families in BC1F2/F3, purple pericarp color was used as the morphological marker.
2.4. Sowing, Phenotyping, and Grain Quality Analysis
A total of 5 vegetation periods, over 17 months, passed from initial crosses till growing BC1F2 plants. The periods were the following: (1) October 2016–January 2017 and (2) February 2017–April 2017 in the Greenhouse Core Facilities of the ICG SB RAS; (3) May 2017–August 2017 in the field experimental plot at the Selection and Genetics Complex in ICG SB RAS (55°02′ N, 82°56′ E), Novosibirsk, Russia; (4) October 2017–January 2018; and (5) February 2018–April 2018 again in the Greenhouse Core Facilities of the ICG SB RAS.
Phenotyping of BC1F3 material and further selection were carried out from May 2018 in the breeding nursery (BN) and the experimental fields at the Omsk State Agrarian University (55°02′ N, 73°32′ E), Omsk, Russia. The lines with purple-colored grains were phenotyped in 1 m2 plots of the BN. Sowing, in the middle of May, was carried out with a SSFK-7 seeder with a sowing rate of 500 seeds per 1 m2. The predecessor was fallow. The cultivars Pamyati Azieva, Duet, and Element 22, as local checks, were sown every 20 numbers. The harvest was carried out in late August. Resistance to powdery mildew (Pm) caused by Erysiphe (syn., Blumeria) graminis DC. f. sp. tritici Marchal, leaf (syn., brown) rust caused by Puccinia triticina Eriks f. sp. Tritici, and stem rust caused by Puccinia graminis Pers. f. sp. tritici Erikss.et Henn. was assessed in the experimental field at the Omsk State Agrarian University (Figure A2).
Reactions to Pm were scored based on the modified version of Saari and Prescott for foliar diseases [43], where 1–10% = resistance (R), 11–25% = moderately resistance (MR), 26–40% = moderately susceptible (MS), 41–65% = susceptible (S), and 66% and above = highly susceptible (HS). Reactions to stem and leaf rusts were evaluated at least 4–5 times during the growing season, according to the scales suggested by Peterson [44], where 0—immunity, uredo pustules are not formed; R (resistance—high resistance), 1 score, the degree of damage is 5–10%; MR (moderately resistance—medium resistance), 2 score, the degree of damage is 10–25%; M (heterogeneous type), pustules of various sizes, surrounded by chlorotic and necrotic spots or without them; MS (moderately susceptible—average susceptibility), 3 score, the degree of damage is 40–50%; S (susceptible-susceptibility), 4 score, the degree of damage is more than 60%.
Grain yield was recorded by weight method at 14% moisture content. Protein and gluten content were detected using IR-Fourier spectrometer “InfroLum FT-10M” (“Lumex”, St. Petersburg).
3. Results
3.1. Marker-Assisted Selection
The crosses and number of plants obtained and assessed during MAS breeding (according to the scheme described in the Section 2 and illustrated in Figure 1) are given in Table 3.
Table 3.
The crosses and number of plants obtained and assessed during MAS breeding.
Homozygous plants having the donor-like microsatellite alleles linked to the Pp3 gene (as an example, microsatellite genotyping results are given at Figure A3) were selected and further genotyped by the markers linked to the Pp-D1 gene. Finally, the homozygous plants having Pp-D1Pp-D1Pp3Pp3 genotype were selected and backcrossed with the recurrent parental cultivars (Table 3).
A low number of F2 seeds was obtained in crosses of cv Aina with both donors and Tobol’skaya with i:S29PF. Among their offspring, a large percentage of sterile bush-shaped plants was observed. This can be related to hybrid dysgenesis in these crosses.
The BC1F1 hybrids were planted and self-pollinated to obtain BC1F2 grains. Around 120 seeds of each combination were germinated and assessed for coleoptile color. Eighty seedlings with the dark-red coleoptile color were taken for each combination and further planted to obtained BC1F3 seeds. The BC1F3 seeds were collected separately from individual plants. The only seeds with purple color from individual plants were planted in particular 1 m2 “BN plots”, in the experimental field at the Omsk State Agrarian University. The individual BC1F3 plants that were resistant to powdery mildew and rust fungi diseases from each plot, which did not have segregation by purple grain color trait, were collected to get BC1F4. In total, 120 BC1F4 lines were selected for field testing.
3.2. Field Screening and Characterization of the Lines
The BC1F4 lines were planted in individual 1 m2 “BN plots” in the experimental field at the Omsk State Agrarian University, in May, and harvested in August 2019, and were assessed in respect to the heading dates, duration of vegetation period, resistance to powdery mildew, stem and leaf rusts, protein and gluten content, and productivity (Table A1). The data on the best lines evaluated in the 1 m2 “BN plots” are presented in Table 4.
Table 4.
Characterization of the best BC1F4 lines in the “breeding nursery (BN) plots” (1 m2), in the experimental field at the Omsk State Agrarian University, 2019.
All lines were divided, according to the maturity dates, into early with the length of the growing season from 72 to 76 days, intermediate with the length of the growing season from 78 to 79 days, and late maturing with a growing season from 84 to 86 days.
In the group of early lines, two genotypes, 275 and 281, developed in the background of BW49880 out yielded check cv Pamyati Azieva with yields of 440 and 417 g/m2, respectively.
In the intermediate group, three lines, i.e., 297, 298, and 301, in the background of cv Aina stood out with yields of 522, 461, and 417 g/m2, respectively.
In the late group, twelve lines 239, 240, 249, 250, 251, 253, 254, 255, 260, 264, 266, and 267 in the background of cv Element 22 surpassed the check cv Element 22 in yield, which varied from 522 to 687 g/m2.
The lines were different in grain protein content, ranging from 12.22% for the line 307 (background of cv Tobol’skaya), having low productivity 318 g/m2 (less than all checks), to 19.18% for the line 264 (background of cv Element 22) characterized by higher productivity 546 g/m2 than its recurrent parent cv Element 22 (Table A1). The highest productivity was shown by line 272 (background of STRU 0732) with a yield of 852 g/m2 and 16.73% protein content. Unfortunately, this line was highly susceptible to stem rust infection.
The biotic resistance of bread wheat plants promotes photosynthesis and grain filling, which significantly improves grain quality and yield. For all the lines, the degree of resistance to powdery mildew varied among the genotypes. Most of the lines showed susceptibility to stem rust and resistance to leaf rust. Only nine lines were moderately resistant to stem rust as follows: late maturing lines 238, 239, 249, 255, 260, and 266 (background of cv Element 22), and medium maturing lines 275, 279, and 281 (background of BW49880). The lines derived from STRU 0732 showed the worst results and were discarded. The lines derived from cv Tobol’skaya were resistant to leaf rust, but susceptible to stem rust. Tobol’skaya is a medium late high-yielding cultivar of the steppe ecotype, characterized by high drought tolerance, good grain quality, and resistance to head smut. The lines derived from it, however, did not demonstrate high productivity and protein content. The best results for disease resistance and yield were demonstrated by lines derived from BW49880 (275, 281), Element 22 (239, 240, 249, 250, 251, 253, 254, 255, 260, 264, 266, and 267), and Aina (297, 298, and 301). These lines were selected for further advanced trials and potential release as the new commercial cultivars in Western Siberia.
4. Discussion
Breeding programs aimed at obtaining anthocyanin-rich wheat cultivars are under way in many countries. A major goal in breeding of purple wheat is to develop high-yielding cultivars adapted to regions of their cultivation, disease resistant, and having good baking and milling quality [45]. This time-consuming process took fourteen years to breed the first commercial purple hard bread wheat cv CDC Primepurple at the University of Saskatchewan (USask) in Canada by Dr. Pierre Hucl. [19]. In the current work, the accelerated breeding strategy based on the achievements by studying molecular-genetics mechanisms for regulation of anthocyanin biosynthesis in wheat grain [30] was proposed and successfully applied to breed promising anthocyanin, rich lines of spring bread wheat adapted to growing conditions of the West Siberian region. The region is characterized by extreme continental climate with frequent droughts (two to three years out of five years), often early summer droughts, severe cold and long winters (150–170 days), relatively short warm summers, low precipitation, sharp temperature fluctuations throughout the year, and even during the day [46]. The climatic conditions of Western Siberia are generally suitable for growing spring-type wheats. Despite the low agro-climatic potential, an abundance of sunlight and heat during the growing season makes the region applicable for production of high-quality wheat grain [47]. However, due to climate warming and, as a consequence, the emergence of aggressive races of fungal diseases, there are significant fluctuations in grain yield from year to year. Cultivars resistant to stem rust occupy about 10–15% of all wheat-cultivated areas in the region [48]. For effective genetic protection of wheat plants, the diversity of cultivated cultivars, in terms of resistance types and genes, plays an important role. The growing of resistant cultivars is the most effective and environmentally friendly approach, as this method eliminates the need to use fungicides and reduces the cost of production. Thus, the main directions of wheat breeding programs in this zone is the development of early maturing, highly productive, resistant to biotic and abiotic stresses spring wheat cultivars. As the purple-grained wheat isogenic donor lines i:S29P and i:S29PF having good levels of grain protein were susceptible to disease, and therefore had low yields [29], the breeding strategy was aimed at developing cultivars not only accumulating anthocyanins in grain but also resistant to fungal diseases and adaptive for growing conditions for this region.
Application of the molecular markers substantially facilitates the selection of the plants with the required genotype and finally speeds up the breeding process [49]. It is especially efficient in case of the grain pericarp color, which is inherited with maternal tissue and has a complex control by two complementary dominant genes, since only 6.25% of the F2 progeny can be used for backcrossing. The breeding strategy for development of anthocyanin-rich cultivars was based on the transfer of the regulatory genes Pp3 and Pp-D1 controlling purple color of wheat grain and mapped on the chromosomes 2A and 7D, respectively, into background of elite cultivars and promising breeding lines adapted for growing conditions of Western Siberia. The critical point of the strategy was the selection of the homozygous plants for the Pp genes in the F2 generation after crossing the donors with the recurrent cultivars/lines. However, before this critical step, it was possible to reduce the cost for molecular screening by selection of progeny with at least one dominant Pp-D1 allele based on a morphological marker, i.e., dark-red coleoptile color due to pleotropic effect of this R2R3-Myb encoding gene on the short arm of chromosome 7D. The use of this morphological marker reduced the progeny size to be analyzed with microsatellite markers by 25%. The remaining F2 progeny was reduced by 75% based on analysis of microsatellites flanking the Pp3 gene. Finally, one-third of the plants selected with microsatellites flanking the Pp-D1 gene were suitable for backcrossing. These were Pp-D1Pp-D1Pp3Pp3 genotypes. Today instead of microsatellites flanking the Pp3 gene, one intragenic marker can be potentially used, since Jiang et al. [32] described promoter region polymorphism between Pp3 and pp3 alleles representing, respectively, expressed and silent TaMyc1 (syn. of Pp3) gene, known earlier from Shoeva et al. [31].
The resulting BC1F3-4 lines were characterized by different intensities of the anthocyanin pigmentation of grains that depended on the genotype. The lightest grain color was in progeny derived from STRU0732, the darkest in BW49880-derived lines. The grain color of the latter line was even deeper than the donor line (Figure A4). The difference can be explained by genes modifying the color. A probable explanation for BW49880-derived lines can also be an increased content of iron and zinc in the grain of this line affecting the formation of stable chelate bonds with anthocyanins [50,51].
We used a breeding scheme without backcross beyond BC1. This scheme which produced progeny with three quarters of the genetic material from recipient has been demonstrated earlier to be efficient to select proper recipient-like lines by consequent field screenings [52]. Especially, since Saratovskaya 29 background in the donor lines is a source of good quality and productivity. Therefore, the main purpose of the backcrossing was to overcome its sensitivity to diseases. The development of the rust resistant cultivars is the focus of the breeding program in Western Siberia and worldwide. The late maturing lines created in the background of cv Element 22 showed the high level of resistance to leaf and stem rusts, inherited from the cv Element 22 which is one of the few cultivars with effective group resistance against all local pathogen populations of stem and leaf rusts [48,53]. The cultivar has wheat-rye translocation 1BL.1RS with the Sr31 gene, co-segregated with Lr26, and Yr9. At present, African stem rust strain Ug99, originated in Uganda, overcame resistance underlined by the Sr31 gene [54]. In Element 22, this gene work is complimented by the Sr35 gene transferred from diploid wheat T. monococcum [55]. Together they provide a high level of resistance to Ug99. Many of the purple-grained lines developed using Element 22 genetic background had higher productivity and grain protein content than its check cv Element 22.
The intermediate maturing purple-grained lines derived from Aina showed resistance to leaf rust, which was connected to the Lr24 gene derived from Thinopyrum ponticum located on chromosome 3DL [56], which always presents with the Sr24 gene with the low level of effectiveness to stem rust. Recently, a new pathotype of Ug99 with virulence to Sr24 has been identified in Kenya [57]. However, the genes Lr24 and Sr24 retain their effectiveness in most regions of Russia and Kazakhstan [48,58]. The cultivar is highly drought resistant, able to withstand a prolonged absence of precipitations and high temperatures and is resistant to lodging. Solid stem of cv Aina protects against harmful insects. Therefore, the best purple-grained lines derived from Aina were also chosen for future tests.
Early maturing lines developed using BW49880 as recipient were resistant to local populations of leaf and stem rusts under condition of Western Siberia. The resistance could be associated with the slow rusting Sr2 gene complex, Sr2+Yr30+Lr27+Pbc (pseudo-black chaff), located on chromosome 3BS [59,60]. The Sr2-complex originating from the cv Hope could be identified by the pseudo-black chaff or brown necrosis phenotype on the glumes and below the nodes [61], that were observed on the glumes of the purple-grained lines developed in the background of BW49880 during their ripening (Figure A5). This complex reduced epidemic development and provided durable and adult plant resistance to a broad spectrum of rusts including Ug99 in the CIMMYT-derived spring wheat germplasm [62]. The lines developed in the genetic background of BW49880 also demonstrated good productivity and high protein content.
The lines derived from cv Tobol’skaya were resistant to leaf rust, but susceptible to stem rust; cv Tobol’skaya is a medium late high-yielding cultivar of the steppe ecotype, characterized by high drought resistance, good grain quality, and resistance to head smut. The lines did not show good results in productivity and protein content as compared with the parental cultivar and were rejected from the further testing.
Thus, the purple-grained lines in the background of cvs Aina, Element 22, and the line BW49880 were selected for further evaluation and potential release as new spring wheat commercial cultivars for Western Siberia.
5. Conclusions
New high-yielding and resistant to fungal disease wheat cultivars with high anthocyanin content in grain is a resource for high added-value products such as healthy functional and dietary food. We demonstrated the efficiency of the breeding strategy which lasting only three years from the first cross between purple-grained donor and regional elite cultivar as recipient until the BC1F4 lines ready for the state cultivar competitive testing. The anthocyanin-rich lines are characterized by increased productivity combined with resistance to the local populations of leaf and stem rust pathogens, which make them environmentally friendly and suitable for modern responsible farming.
Author Contributions
Crossing and evaluating with morphological markers, E.G. and T.K.; molecular marker screening, E.G.; field screenings and analysis of grain quality, V.S.; manuscript draft preparation, E.G. and V.S.; participation data interpretation and revising the manuscript critically, O.S.; coordinating the study, contributing to its conception and design, and revising the manuscript critically, A.M. and E.K. All authors have read and agreed to the published version of the manuscript.
Funding
Development of the strategy for breeding of anthocyanin-rich cultivars and MAS of the hybrids was supported by the Russian Foundation for Basic Research (RFBR), grant no. 19-016-00140. The phenotyping of the lines and grain quality assessment carried out in the experimental field and laboratory at the Omsk State Agrarian University, were supported by the Russian Science Foundation (RSF), grant no. 16-16-10005.
Acknowledgments
We thank Galina Generalova and Nina Kovaleva for technical assistance. Growing of wheat plants in ICG Greenhouse Core Facility was supported by ICG project No. 0324-2019-0039-C-01.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Appendix A

Figure A1.
Schematic representation of introgressions from cvs Purple Feed and Purple into the wheat chromosomes 2A and 7D carrying the regulatory Pp genes determining purple grain color which were used as donor lines for breeding anthocyanin-rich cultivars according to Tereschenko et al. (2012).


Figure A2.
The traits of the fungal pathogens diseases affected wheat. Powdery mildew caused by Erysiphe (Blumeria) graminis (left picture in the top panel); leaf rust caused by Puccinia triticina (right picture in the top panel); and stem rust caused by Puccinia graminis (in the bottom row).


Figure A3.
Electrophoregram of PCR products obtained using primers for the microsatellite locus Xgwm0312 (chromosome 2A) on DNA of the isogenic donor line i:S29Pp-D1Pp3PF (P1, lane 1), cv Aina DNA (P2, lane 2), and corresponding F2 heterozygous / homozygous progenies.

Table A1.
The results of the assessment of the purple-grained wheat lines (BC1F4) from “BN plots” in the experimental field at the Omsk State Agrarian University, 2019.

Figure A4.
Grains of wheat cultivars/lines without anthocyanins (in the top row) and that of their purple-grained derivatives (in the bottom row). (1) ‘Saratovskaya 29′ and i:S29PF; (2) ‘Element 22′ and its BC1F4 line; (3) BW49880 and its BC1F4 line.


Figure A5.
Brown necrosis or pseudo-black chaff on the glumes of wheat spikes of the purple-grained lines on BW49880 genetic background, which markers of the Sr2-complex.

References
- Weststrate, J.A.; van Poppe, G.; Verschuren, P.M. Functional foods, trends and future. Br. J. Nutr. 2002, 88, S233–S235. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Aal, E.S.M.; Young, J.C.; Rabalski, I. Anthocyanin composition in black, blue, pink, purple, and red cereal grains. J. Agric. Food Chem. 2006, 54, 4696–4704. [Google Scholar] [CrossRef] [PubMed]
- Landi, M.; Tattini, M.; Gould, K.S. Multiple functional roles of anthocyanins in plant-environment interactions. Environ. Exp. Bot. 2015, 119, 4–17. [Google Scholar] [CrossRef]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef]
- Li, D.; Wang, P.; Luo, Y.; Zhao, M.; Chen, F. Health benefits of anthocyanins and molecular mechanisms: Update from recent decade. Crit. Rev. Food Sci. Nutr. 2017, 57, 1729–1741. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, J.; Liu, F.; Tong, L.; Chen, Z.; Chen, J.; Huang, C. Neuroprotective effects of anthocyanins and its major component cyanidin-3-O-glucoside (C3G) in the central nervous system: An outlined review. Eur. J. Pharmacol. 2019, 858, 172500. [Google Scholar] [CrossRef]
- Törrönen, R.; Kolehmainen, M.; Sarkkinen, E.; Poutanen, K.; Mykkänen, H.; Niskanen, L. Berries reduce postprandial insulin responses to wheat and rye breads in healthy women. J. Nutr. 2013, 143, 430–436. [Google Scholar] [CrossRef]
- Tikhonova, M.A.; Amstislavskaya, T.G.; Akopyan, A.A.; Ovsyukova, M.V.; Tenditnik, M.V.; Khlestkina, E.K. Effects of diets rich in plant polyphenols in mouse models of neurodegenerative disorders. In Proceedings of the 12th International Multiconference: Bioinformatics of Genome Regulation and Structure/Systems Biology (BGRS/SB-2020), Novosibirsk, Russia, 6–10 July 2020; p. 292. [Google Scholar] [CrossRef]
- Tenditnik, M.V.; Tikhonova, M.A.; Litvinova, E.A.; Popova, N.A.; Amstislavskaya, T.G.; Khlestkina, E.K. Effects of anthocyanin-rich grain diet on growth and metastasis of Lewis lung carcinoma in mice. In Proceedings of the 12th International Multiconference: Bioinformatics of Genome Regulation and Structure/Systems Biology (BGRS/SB-2020), Novosibirsk, Russia, 6–10 July 2020; p. 359. [Google Scholar] [CrossRef]
- Gamel, T.H.; Abdel-Aal, E.S.M.; Tucker, A.J.; Pare, S.M.; Faughnan, K.; O’Brien, C.D.; Wright, A.J. Consumption of whole purple and regular wheat modestly improves metabolic markers in adults with elevated high-sensitivity C-reactive protein: A randomised, single-blind parallel-arm study. Br. J. Nutr. 2020, 1–11. [Google Scholar] [CrossRef]
- Zeven, A.C. Wheats with purple and blue grains: A review. Euphytica 1991, 56, 243–258. [Google Scholar] [CrossRef]
- Badaeva, E.D.; Shishkina, A.A.; Goncharov, N.P.; Zuev, E.V.; Lysenko, N.S.; Mitrofanova, O.P.; Dragovich, A.Y.; Kudriavtsev, A.M. Evolution of Triticum aethiopicum Jakubz. from the position of chromosome analysis. Russ. J. Genet. 2018, 54, 629–642. [Google Scholar] [CrossRef]
- Belay, G.; Tesemma, T.; Bechere, E.; Mitiku, D. Natural and human selection for purple-grain tetraploid wheats in the Ethiopian highlands. Gen. Res. Crop Evol. 1995, 42, 387–391. [Google Scholar] [CrossRef]
- Copp, L.G.L. Purple grain in hexaploid wheat. Wheat Inf. Serv. 1965, 20, 18. [Google Scholar]
- Jensen, N.F. Registration of Charcoal wheat germplasm. Crop Sci. 1977, 17, 983. [Google Scholar] [CrossRef]
- Morin, C. Crop Development Breeds Patience for USask Wheat Researcher. 2019. Available online: https://agbio.usask.ca/news/2019/12/news-crop-development-breeds-patience-for-usask-wheat-researcher.php (accessed on 20 August 2020).
- Griffin, W.B. Outcrossing in New Zealand wheats measured by occurrence of purple grain. New Zealand J. Agric. Res. 1987, 30, 287–290. [Google Scholar] [CrossRef]
- Syed Jaafar, S.N.; Baron, J.; Siebenhandl-Ehn, S.; Rosenau, T.; Böhmdorfer, S.; Grausgruber, H. Increased anthocyanin content in purple pericarp× blue aleurone wheat crosses. Plant Breed. 2013, 132, 546–552. [Google Scholar] [CrossRef]
- King, C. New Possibilities with Purple Wheat. 5 May 2017. Available online: https://www.topcropmanager.com/new-possibilities-with-purple-wheat-20050/ (accessed on 20 September 2020).
- Li, W.; Pickard, M.D.; Beta, T. Evaluation of antioxidant activity and electronic taste and aroma properties of antho-beers from purple wheat grain. J. Agric. Food Chem. 2007, 55, 8958–8966. [Google Scholar] [CrossRef]
- Corrêa, R.C.G.; Garcia, J.A.A.; Correa, V.G.; Vieira, T.F.; Bracht, A.; Peralta, R.M. Pigments and vitamins from plants as functional ingredients: Current trends and perspectives. Adv. Food Nutr. Res. 2019, 90, 259–303. [Google Scholar] [CrossRef]
- Bartl, P.; Albreht, A.; Skrt, M.; Tremlová, B.; Ošťádalová, M.; Šmejkal, K.; Ulrih, N.P. Anthocyanins in purple and blue wheat grains and in resulting bread: Quantity, composition, and thermal stability. Int. J. Food Sci. Nutr. 2015, 66, 514–519. [Google Scholar] [CrossRef]
- Khlestkina, E.K.; Usenko, N.I.; Gordeeva, E.I.; Stabrovskaya, O.I.; Sharfunova, I.B.; Otmakhova, Y.S. Evaluation of wheat products with high flavonoid content: Justification of importance of marker-assisted development and production of flavonoid-rich wheat cultivars. Vavilov. J. Genet. Breed. 2017, 21, 545–553. [Google Scholar] [CrossRef]
- Pasqualone, A.; Bianco, A.M.; Paradiso, V.M.; Summo, C.; Gabarcorta, G.; Caponio, F.; Blanco, A. Production and characterization of functional biscuits obtained from purple wheat. Food Chem. 2015, 180, 64–70. [Google Scholar] [CrossRef]
- Usenko, N.I.; Khlestkina, E.K.; Asavasanti, S.; Gordeeva, E.I.; Yudina, R.S.; Otmakhova, Y.S. Possibilities of enriching food products with anthocyanins by using new forms of cereals. Foods Raw Mater. 2018, 6, 128–135. [Google Scholar] [CrossRef]
- Ficco, D.B.M.; De Simone, V.; De Leonardis, A.M.; Giovanniello, V.; Del Nobile, M.A.; Padalino, L.; De Vita, P. Use of purple durum wheat to produce naturally functional fresh and dry pasta. Food Chem. 2016, 100, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Gamel, T.H.; Wright, A.J.; Pickard, M.; Abdel-Aal, E.S.M. Characterization of anthocyanin-containing purple wheat prototype products as functional foods with potential health benefits. Cereal Chem. 2020, 97, 34–38. [Google Scholar] [CrossRef]
- Burešová, I.; Trojan, V.; Helis, M. Characteristics of flour and dough from purple and blue wheat grain. Potravin. Slovak J. Food Sci. 2019, 13, 163–166. [Google Scholar] [CrossRef]
- Morgounov, A.; Karaduman, Y.; Akin, B.; Aydogan, S.; Baenziger, P.S.; Bhatta, M.; Guzman, C. Yield and quality in purple-grained wheat isogenic lines. Agronomy 2020, 10, 86. [Google Scholar] [CrossRef]
- Khlestkina, E.K.; Shoeva, O.Y.; Gordeeva, E.I. Flavonoid biosynthesis genes in wheat. Russ. J. Genet. Appl. Res. 2015, 5, 268–278. [Google Scholar] [CrossRef]
- Shoeva, O.; Gordeeva, E.; Khlestkina, E. The regulation of anthocyanin synthesis in the wheat pericarp. Molecules 2014, 19, 20266–20279. [Google Scholar] [CrossRef]
- Jiang, W.; Liu, T.; Nan, W.; Jeewani, D.C.; Niu, Y.; Li, C.; Wang, Y.; Shi, X.; Wang, C.; Wang, J.; et al. Two transcription factors TaPpm1 and TaPpb1 co-regulate anthocyanin biosynthesis in purple pericarps of wheat. J. Exp. Botany 2018, 69, 2555–2567. [Google Scholar] [CrossRef]
- Himi, E.; Taketa, S. Isolation of candidate genes for the barley Ant1 and wheat Rc genes controlling anthocyanin pigmentation in different vegetative tissues. Mol. Genet. Genom. 2015, 290, 1287–1298. [Google Scholar] [CrossRef]
- Gordeeva, E.I.; Glagoleva, A.Y.; Kukoeva, T.V.; Khlestkina, E.K.; Shoeva, O.Y. Purple-grained barley (Hordeum vulgare L.): Marker-assisted development of NILs for investigating peculiarities of the anthocyanin biosynthesis regulatory network. BMC Plant. Biol. 2019, 19, 49–57. [Google Scholar] [CrossRef]
- Arbuzova, V.S.; Maystrenko, O.I.; Popova, O.M. Development of near-isogenic lines of the common wheat cultivar ‘Saratovskaya 29’. Cereal Res. Commun. 1998, 26, 39–46. [Google Scholar] [CrossRef]
- Tereshchenko, O.; Gordeeva, E.; Arbuzova, V.; Börner, A.; Khlestkina, E. The D genome carries a gene determining purple grain colour in wheat. Cereal Res. Com. 2012, 40, 334–341. [Google Scholar] [CrossRef]
- Chatrath, R.; Gupta, V.; Parkash, O.; Singh, G.P. Evaluation of biofortified spring wheat genotypes for yield and micronutrients. J. Appl. Nat. Sci. 2018, 10, 210–215. [Google Scholar] [CrossRef]
- Ghimire, S.; Thapa, D.B.; Paudel, A.; Adhikari, N.R. Variability study of biofortified bread wheat genotypes for grain Zinc and Iron concentration, yield and yield associated traits at Khumaltar, Lalitpur, Nepal. Int. J. Appl. Sci. Biotechnol. 2019, 7, 184–194. [Google Scholar] [CrossRef][Green Version]
- Röder, M.S.; Korzun, V.; Wendehake, K.; Plaschke, J.; Tixier, M.H.; Leroy, P.; Ganal, M.W. A microsatellite map of wheat. Genetics 1998, 149, 2007–2023. [Google Scholar]
- Ganal, M.W.; Röder, M.S. Microsatellite and SNP markers in wheat breeding. In Genomics-Assisted Crop Improvement; Springer: Dordrecht, Germany, 2007; pp. 1–24. [Google Scholar]
- Gordeeva, E.I.; Shoeva, O.Y.; Khlestkina, E.K. Marker-assisted development of bread wheat near-isogenic lines carrying various combinations of purple pericarp (Pp) alleles. Euphytica 2015, 203, 469–476. [Google Scholar] [CrossRef]
- Plaschke, J.; Ganal, M.W.; Röder, M.S. Detection of genetic diversity in closely related bread wheat using microsatellite markers. Theor. Appl. Genet. 1995, 91, 1001–1007. [Google Scholar] [CrossRef]
- Koyshybaev, M.; Shamanin, V.P.; Morgunov, A.I. Screening of Wheat for Resistance to Major Diseases; FAO-SEK: Ankara, Turkey, 2014. (In Russian) [Google Scholar]
- Peterson, R.F.; Campbell, A.B.; Hannah, A.E. A diagrammatic scale for estimating rust intensity on leaves and stems of cereals. Can. J. Res. 1948, 26, 496–500. [Google Scholar] [CrossRef]
- Martinek, P.; Jirsa, O.; Vaculova, K.; Chrpova, J.; Watanabe, N.; Buresova, V.; Kopecky, D.; Stiasna, K.; Vyhnanek, T.; Trojan, V. Use of wheat gene resources with different colour in breeding. Proc. Tag. Pflanz. 2014, 64, 1–4. [Google Scholar]
- Zykin, V.A.; Shamanin, V.P.; Belan, I.A. Ecology of Wheat: Monograph; Publishing house Omsk state Agrarian University: Omsk, Russia, 2000; p. 124. (In Russian) [Google Scholar]
- Likhenko, I.E.; Sovetov, V.V.; Anosov, S.I.; Likhenko, N.N. Formation of grain yield of Siberian cultivars of spring soft wheat under conditions of continental climate of Western Siberia. Res. Tech. Adv. Agribus. Sector 2014, 1, 27–30. (In Russian) [Google Scholar]
- Shamanin, V.P.; Pototskaya, I.V.; Shepelev, S.S.; Pozherukova, V.E.; Salina, Е.А.; Skolotneva, Е.S.; Morgounov, A.I. Stem rust in Western Siberia–race composition and effective resistance genes. Vavilov. J. Genet. Breed. 2020, 24, 131–138. [Google Scholar] [CrossRef]
- Khlestkina, E.K. Molecular markers in genetic studies and breeding. Russ. J. Genet. Appl. Res. 2014, 4, 236–244. [Google Scholar] [CrossRef]
- Luna-Vital, D.; Cortez, R.; Ongkowijoyo, P.; de Mejia, E.G. Protection of color and chemical degradation of anthocyanin from purple corn (Zea mays L.) by zinc ions and alginate through chemical interaction in a beverage model. Food Res. Int. 2018, 105, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Zhu, X.; Li, Y.; Wang, C. Analysis of the ph-dependent fe (iii) ion chelating activity of anthocyanin extracted from black soybean [Glycine max (L.) Merr.] coats. J. Agric. Food Chem. 2018, 66, 1131–1139. [Google Scholar] [CrossRef] [PubMed]
- Collard, B.C.Y.; Jahufer, M.Z.Z.; Brouwer, J.B.; Pang, E.C.K. An introduction to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: The basic concepts. Euphytica 2005, 142, 169–196. [Google Scholar] [CrossRef]
- Shamanin, V.; Salina, E.; Wanyera, R.; Zelenskiy, Y.; Olivera, P.; Morgounov, A. Genetic diversity of spring wheat from Kazakhstan and Russia for resistance to stem rust Ug99. Euphytica 2016, 212, 287–296. [Google Scholar] [CrossRef]
- Singh, R.P.; Hodson, D.P.; Huerta-Espino, J.; Jin, Y.; Njau, P.; Wanyera, R.; Ward, R.W. Will stem rust destroy the world’s wheat crop? Adv. Agron. 2008, 98, 271–309. [Google Scholar]
- Babiker, E.; Ibrahim, A.M.; Yen, Y.; Stein, J. Identification of a microsatellite marker associated with stem rust resistance gene ‘sr35’ in wheat. Aust. J. Crop. Sci. 2009, 3, 195–200. [Google Scholar]
- Gupta, S.K.; Charpe, A.; Koul, S.; Haque, Q.M.R.; Prabhu, K.V. Development and validation of SCAR markers co-segregating with an Agropyron elongatum derived leaf rust resistance gene Lr24 in wheat. Euphytica 2006, 150, 233–240. [Google Scholar] [CrossRef]
- Jin, Y.; Szabo, L.J.; Pretorius, Z.A.; Singh, R.P.; Ward, R.; Fetch, T. Detection of virulence to resistance gene Sr24 within race TTKS of Puccinia graminis f. sp tritici. Plant Dis. 2008, 92, 923–926. [Google Scholar] [CrossRef]
- Gultyaeva, E.I.; Shaydayuk, E.L.; Shamanin, V.P.; Akhmetova, A.K.; Tyunin, V.A.; Shreider, E.R.; Morgunov, A.I. Genetic structure of Russian and Kazakhstani leaf rust causative agent Puccinia triticina Erikss. populations as assessed by virulence profiles and SSR markers. Agric. Biol. 2018, 53, 85–95. [Google Scholar] [CrossRef]
- Spielmeyer, W.; Sharp, P.J.; Lagudah, E.S. Identification and validation of markers linked to broad-spectrum stem rust resistance gene Sr2 in wheat (Triticum aestivum L.). Crop. Sci. 2003, 43, 333–336. [Google Scholar]
- Aktar-Uz-Zaman, M.; Tuhina-Khatun, M.; Hanafi, M.M.; Sahebi, M. Genetic analysis of rust resistance genes in global wheat cultivars: An overview. Biotechnol. Biotechnol. Equip. 2017, 31, 431–445. [Google Scholar] [CrossRef]
- Huerta-Espino, J.; Singh, R.; Crespo-Herrera, L.A.; Villaseñor-Mir, H.E.; Rodriguez-Garcia, M.F.; Dreisigacker, S.; Lagudah, E. Adult plant slow rusting genes confer high levels of resistance to rusts in bread wheat cultivars from Mexico. Front. Plant Sci. 2020, 11, 824. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Huerta-Espino, J.; Bhavani, S.; Herrera-Foessel, S.A.; Singh, D.; Singh, P.K.; Crossa, J. Race non-specific resistance to rust diseases in CIMMYT spring wheats. Euphytica 2011, 179, 175–186. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).