
Effects of Lactation Lactoferrin Deficiency on Intestinal Microbiota in Different Mice Models


Abstract
1. Introduction
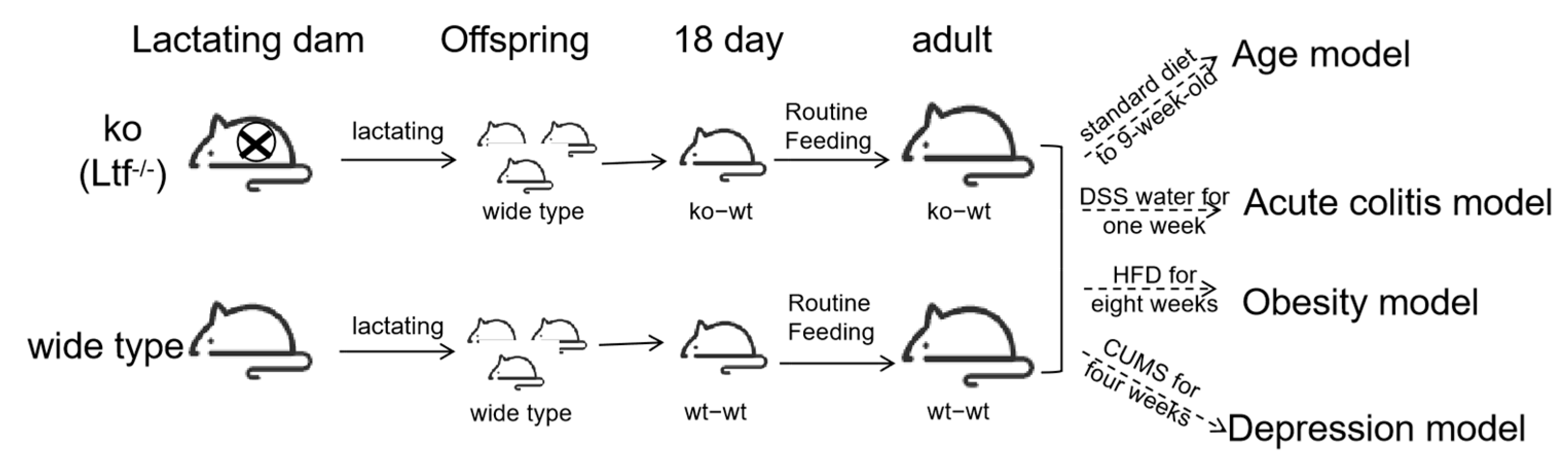
2. Method
2.1. Animal
2.1.1. Age Model
2.1.2. High-Fat Diet Model
2.1.3. DSS-Induced Colitis
2.1.4. CUMS-Induced Depression-like Behavior
2.2. Microbiome Composition Assessment via 16S rRNA Amplicon Sequencing
2.3. Statistical Analysis
3. Results
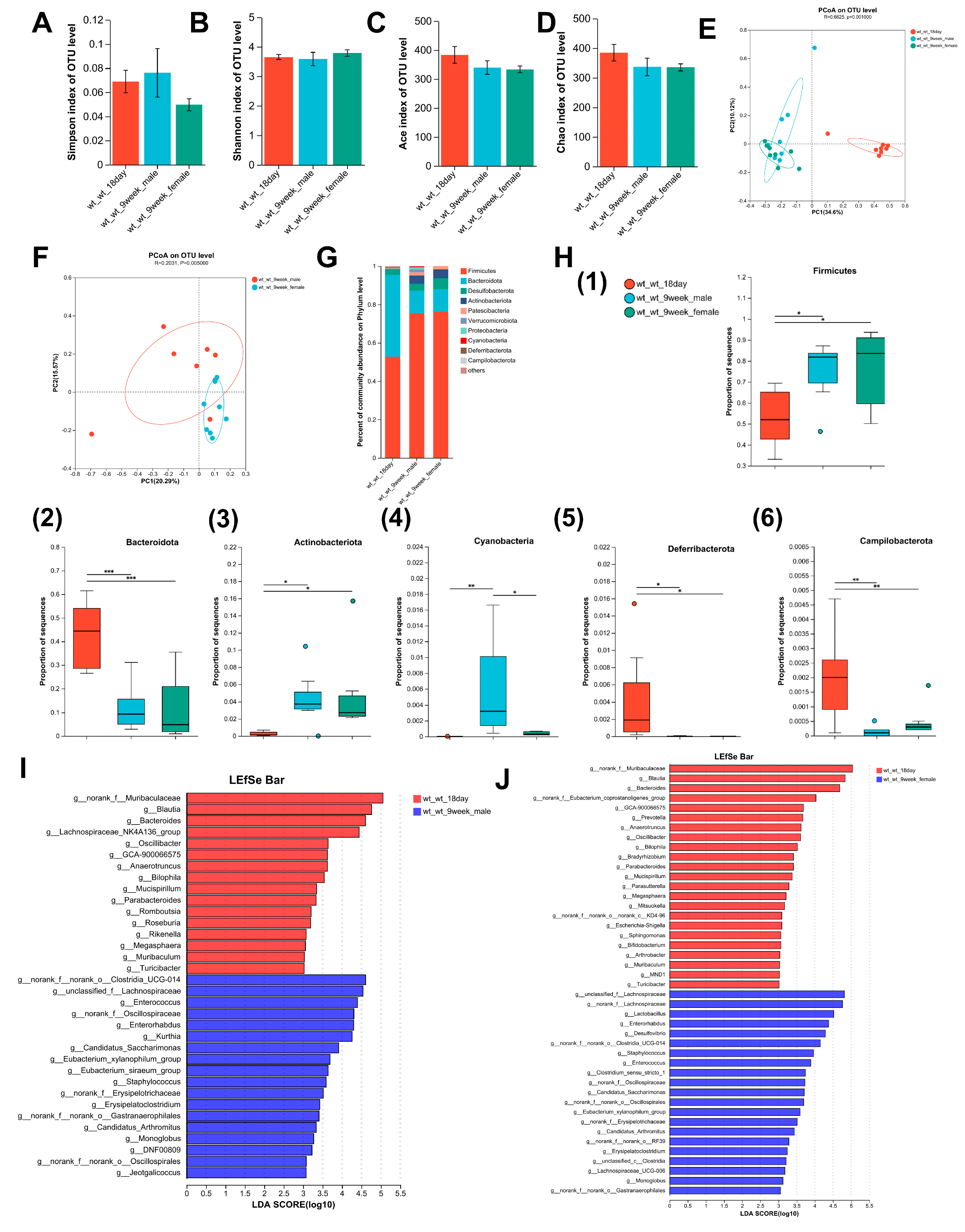
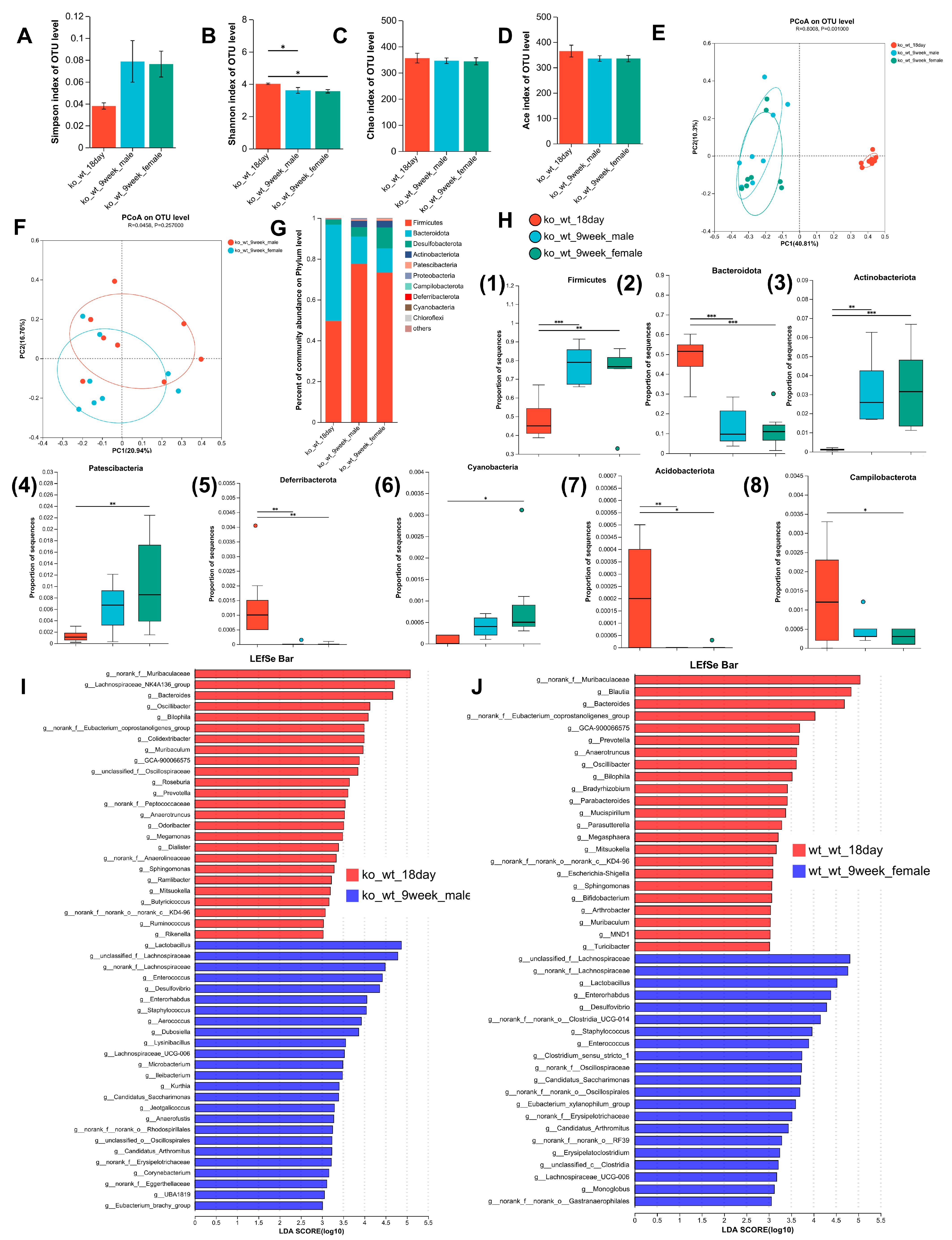
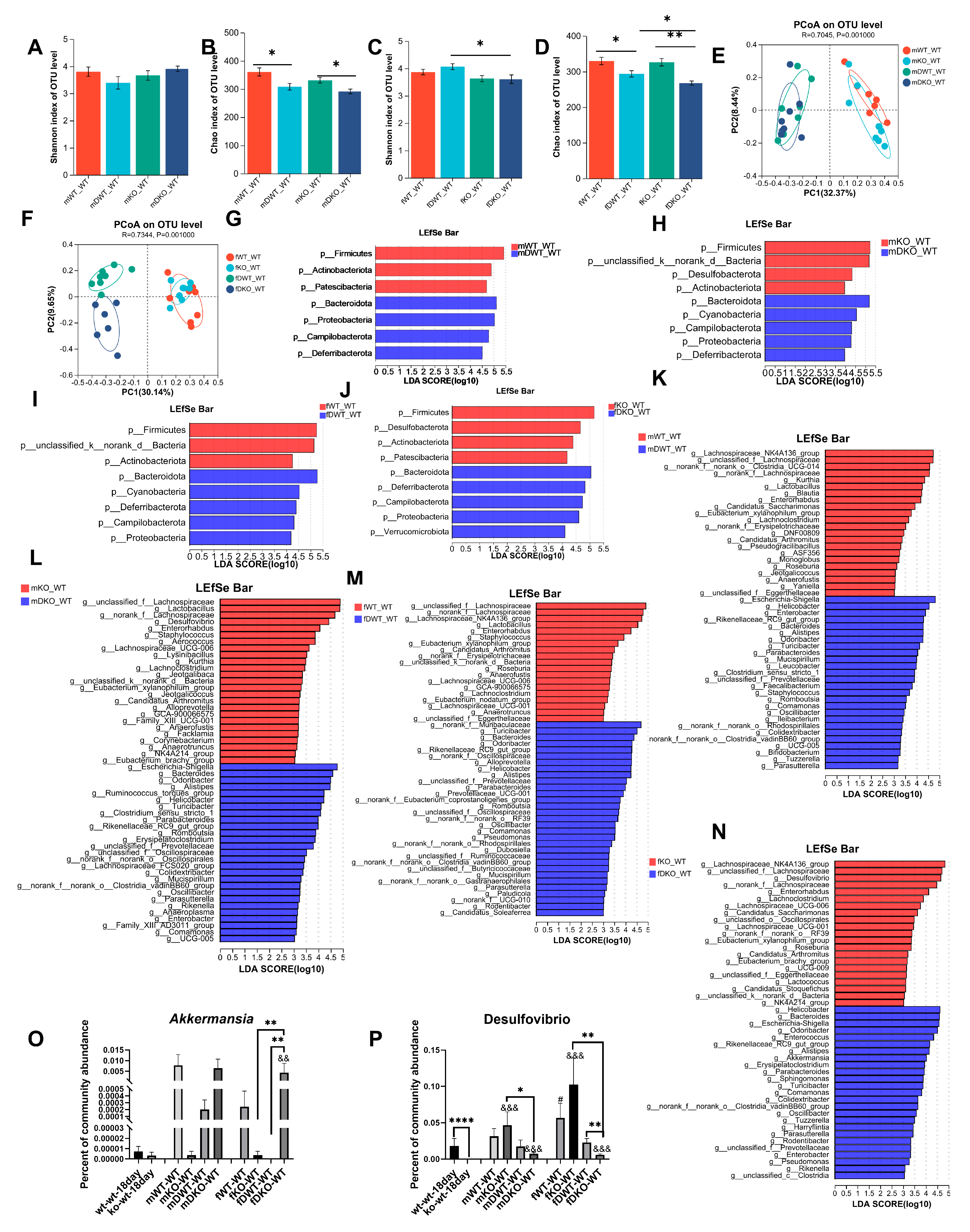
3.1. Alterations in the Composition of the Intestinal Microbiome with Age in Mice
3.2. Alterations in the Composition of the Intestinal Microbiome with Age in Lactation LF Feeding-Deficient Mice
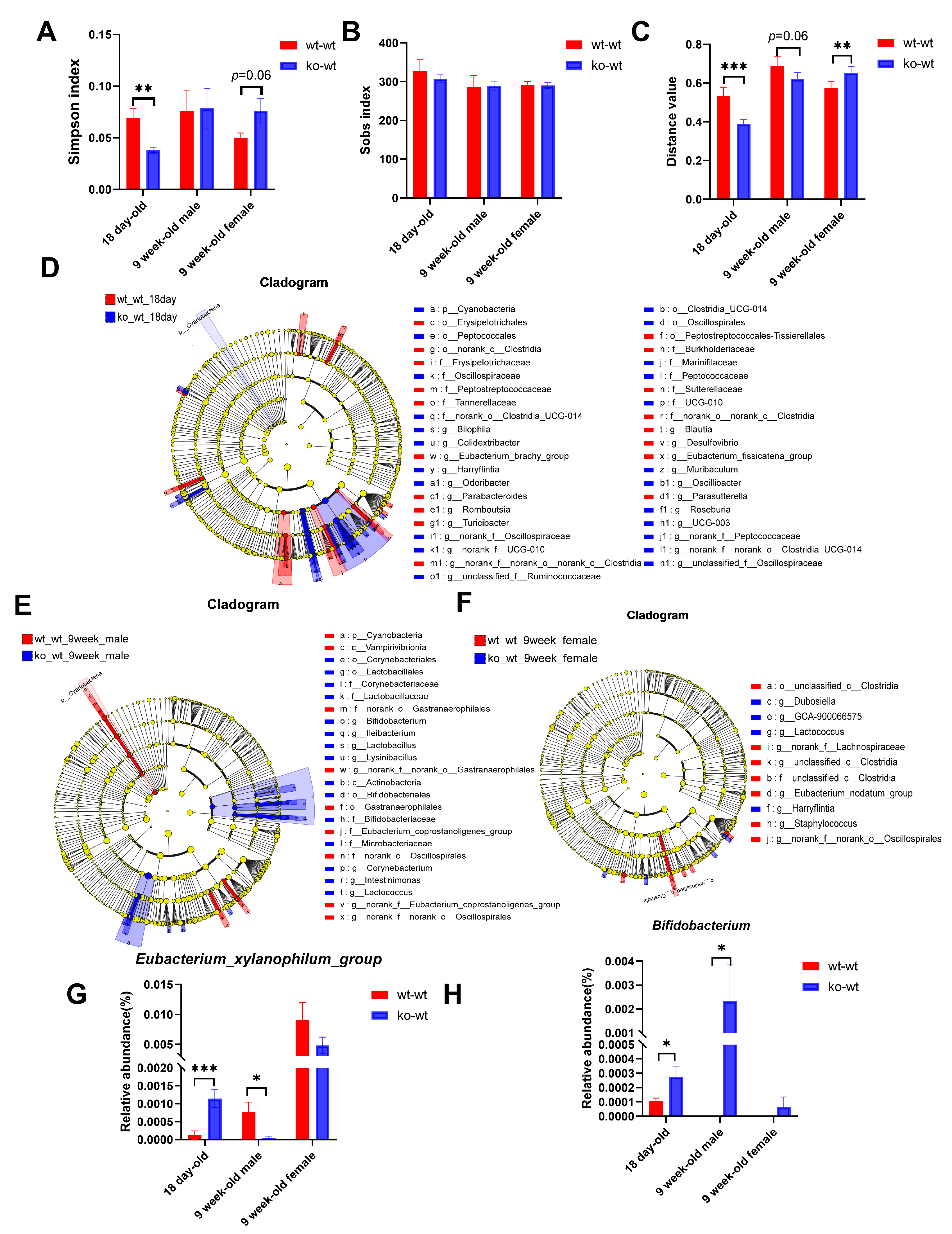
3.3. Effects of LF Deficiency During Lactation on Intestinal Microbial Composition in 18-Day-Old and Adult Mice
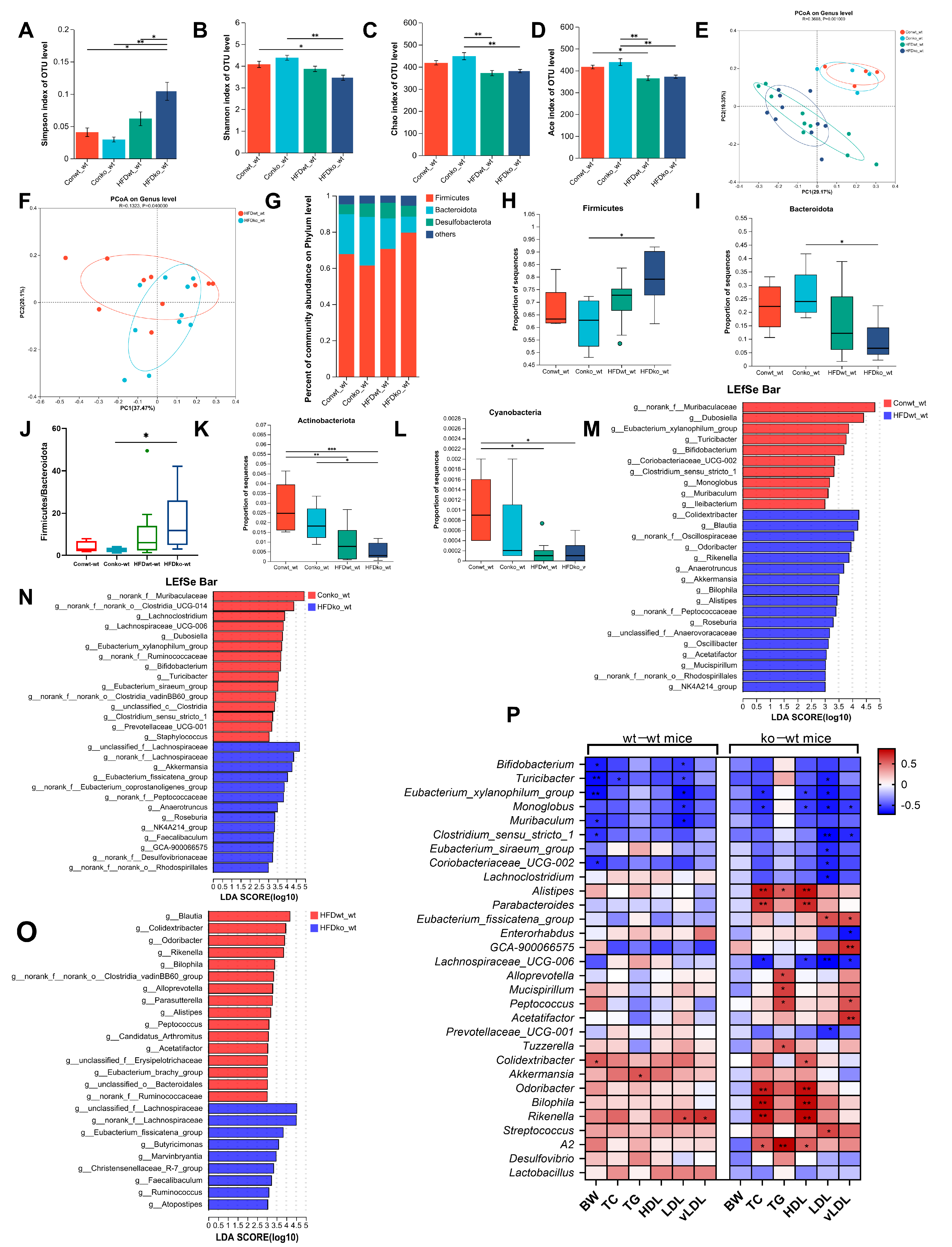
3.4. Effects of LF Deficiency During Lactation on Intestinal Microbial Composition in Adult Obese Mice Induced by HFD
3.5. Alterations in the Gut Microbiota of Mice with Acute Colitis Induced by DSS
3.6. Effects of LF Deficiency During Lactation on Intestinal Microbial Composition in CUMS-Induced Depression-like Behavior Mice
3.7. The Connections Between Bacteria That Are Enriched in Various Models
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| IBD | inflammatory bowel disease |
| LF | lactoferrin |
| HFD | high-fat diet |
| DSS | dextran sodium sulfate |
| KO | knockout |
| WT | wild-type |
| OTU | operational taxonomic units |
| PCoA | principal coordinate analysis plots |
| TC | total cholesterol |
| HDL | high-density lipoprotein |
| LDL | low-density lipoprotein |
| CUMS | chronic unpredictable mild stress |
| CRC | colorectal cancer |
| SCFA | short-chain fatty acid |
References
- Goodrich, J.K.; Davenport, E.R.; Clark, A.G.; Ley, R.E. The Relationship Between the Human Genome and Microbiome Comes into View. Annu. Rev. Genet. 2017, 51, 413–433. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, L.; Chen, H.; Zhang, S.; Zhuang, J.; Li, Q.; Feng, Z. Intestinal Microbiota in Early Life and Its Implications on Childhood Health. Genom. Proteom. Bioinform. 2019, 17, 13–25. [Google Scholar] [CrossRef]
- Simpson, C.A.; Diaz-Arteche, C.; Eliby, D.; Schwartz, O.S.; Simmons, J.G.; Cowan, C.S.M. The gut microbiota in anxiety and depression—A systematic review. Clin. Psychol. Rev. 2021, 83, 101943. [Google Scholar] [CrossRef]
- Lee, J.; Cevallos, S.A.; Byndloss, M.X.; Tiffany, C.R.; Olsan, E.E.; Butler, B.P.; Young, B.M.; Rogers, A.W.L.; Nguyen, H.; Kim, K.; et al. High-Fat Diet and Antibiotics Cooperatively Impair Mitochondrial Bioenergetics to Trigger Dysbiosis that Exacerbates Pre-inflammatory Bowel Disease. Cell Host Microbe 2020, 28, 273–284. [Google Scholar] [CrossRef]
- Tsai, S.F.; Hsu, P.L.; Chen, Y.W.; Hossain, M.S.; Chen, P.C.; Tzeng, S.F.; Chen, P.S.; Kuo, Y.M. High-fat diet induces depression-like phenotype via astrocyte-mediated hyperactivation of ventral hippocampal glutamatergic afferents to the nucleus accumbens. Mol. Psychiatry 2022, 27, 4372–4384. [Google Scholar] [CrossRef] [PubMed]
- Bisgaard, T.H.; Allin, K.H.; Keefer, L.; Ananthakrishnan, A.N.; Jess, T. Depression and anxiety in inflammatory bowel disease: Epidemiology, mechanisms and treatment. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 717–726. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Xu, J.; Li, J.; Deng, M.; Shen, Z.; Nie, K.; Luo, W.; Zhang, C.; Ma, K.; Chen, X.; et al. Bacteroides vulgatus alleviates dextran sodium sulfate-induced colitis and depression-like behaviour by facilitating gut-brain axis balance. Front. Microbiol. 2023, 14, 1287271. [Google Scholar] [CrossRef]
- Wang, W.; An, Q.; Huang, K.; Dai, Y.; Meng, Q.; Zhang, Y. Unlocking the power of Lactoferrin: Exploring its role in early life and its preventive potential for adult chronic diseases. Food Res. Int. 2024, 182, 114143. [Google Scholar] [CrossRef]
- Wang, W.; Cheng, Z.; Wang, X.; An, Q.; Huang, K.; Dai, Y.; Meng, Q.; Zhang, Y. Lactoferrin deficiency during lactation increases the risk of depressive-like behavior in adult mice. BMC Biol. 2023, 21, 242. [Google Scholar] [CrossRef]
- Hill, J.H.; Round, J.L. SnapShot: Microbiota effects on host physiology. Cell 2021, 184, 2796. [Google Scholar] [CrossRef]
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Cresci, G.A.; Bawden, E. Gut Microbiome: What we do and don’t know. Nutr. Clin. Pract. 2015, 30, 734–746. [Google Scholar] [CrossRef]
- Wu, C.S.; Muthyala, S.; Klemashevich, C.; Ufondu, A.U.; Menon, R.; Chen, Z.; Devaraj, S.; Jayaraman, A.; Sun, Y. Age-dependent remodeling of gut microbiome and host serum metabolome in mice. Aging 2021, 13, 6330–6345. [Google Scholar] [CrossRef] [PubMed]
- Rautava, S.; Luoto, R.; Salminen, S.; Isolauri, E. Microbial contact during pregnancy, intestinal colonization and human disease. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 565–576. [Google Scholar] [CrossRef]
- Yin, J.; Li, Y.; Han, H.; Chen, S.; Gao, J.; Liu, G.; Wu, X.; Deng, J.; Yu, Q.; Huang, X.; et al. Melatonin reprogramming of gut microbiota improves lipid dysmetabolism in high-fat diet-fed mice. J. Pineal Res. 2018, 65, e12524. [Google Scholar] [CrossRef]
- de Wit, N.; Derrien, M.; Bosch-Vermeulen, H.; Oosterink, E.; Keshtkar, S.; Duval, C.; de Vogel-van, D.B.J.; Kleerebezem, M.; Müller, M.; van der Meer, R. Saturated fat stimulates obesity and hepatic steatosis and affects gut microbiota composition by an enhanced overflow of dietary fat to the distal intestine. Am. J. Physiol. Gastroint. Liver Physiol. 2012, 303, G589–G599. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Kong, Q.; Li, X.; Zhao, J.; Zhang, H.; Chen, W.; Wang, G. A High-Fat Diet Increases Gut Microbiota Biodiversity and Energy Expenditure Due to Nutrient Difference. Nutrients 2020, 12, 3197. [Google Scholar] [CrossRef]
- Xu, H.M.; Huang, H.L.; Liu, Y.D.; Zhu, J.Q.; Zhou, Y.L.; Chen, H.T.; Xu, J.; Zhao, H.L.; Guo, X.; Shi, W.; et al. Selection strategy of dextran sulfate sodium-induced acute or chronic colitis mouse models based on gut microbial profile. BMC Microbiol. 2021, 21, 279. [Google Scholar] [CrossRef]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef]
- Mariat, D.; Firmesse, O.; Levenez, F.; Guimaraes, V.; Sokol, H.; Dore, J.; Corthier, G.; Furet, J.P. The Firmicutes/Bacteroidetes ratio of the human microbiota changes with age. BMC Microbiol. 2009, 9, 123. [Google Scholar] [CrossRef]
- Tseng, C.; Wu, C. The gut microbiome in obesity. J. Formos. Med. Assoc. 2019, 118, S3–S9. [Google Scholar] [CrossRef] [PubMed]
- Yee, S.; Choi, H.; Seon, J.; Ban, Y.; Kim, M.; Seo, J.; Seo, J.H.; Kim, S.; Moon, S.H.; Yun, C.; et al. Axl alleviates DSS-induced colitis by preventing dysbiosis of gut microbiota. Sci. Rep. 2023, 13, 5371. [Google Scholar] [CrossRef]
- Wu, F.; Lei, H.; Chen, G.; Chen, C.; Song, Y.; Cao, Z.; Zhang, C.; Zhang, C.; Zhou, J.; Lu, Y.; et al. In Vitro and In Vivo Studies Reveal that Hesperetin-7-O-glucoside, a Naturally Occurring Monoglucoside, Exhibits Strong Antiinflammatory Capacity. J. Agric. Food Chem. 2021, 69, 12753–12762. [Google Scholar] [CrossRef]
- Liu, X.; Mao, B.; Gu, J.; Wu, J.; Cui, S.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Blautia-a new functional genus with potential probiotic properties? Gut Microbes 2021, 13, 1875796. [Google Scholar] [CrossRef] [PubMed]
- Xue, M.; Teng, X.; Liang, H.; Zhao, J.; Jiang, Y.; Qiu, X.; Zhang, Z.; Pei, Z.; Zhang, N.; Qin, Y. Neuroprotective effect of fucoidan by regulating gut-microbiota-brain axis in alcohol withdrawal mice. J. Funct. Foods 2021, 86, 104726. [Google Scholar] [CrossRef]
- Parker, B.J.; Wearsch, P.A.; Veloo, A.C.M.; Rodriguez-Palacios, A. The Genus Alistipes: Gut Bacteria with Emerging Implications to Inflammation, Cancer, and Mental Health. Front. Immunol. 2020, 11, 906. [Google Scholar] [CrossRef]
- Yu, M.; Yue, J.; Hui, N.; Zhi, Y.; Hayat, K.; Yang, X.; Zhang, D.; Chu, S.; Zhou, P. Anti-Hyperlipidemia and Gut Microbiota Community Regulation Effects of Selenium-Rich Cordyceps militaris Polysaccharides on the High-Fat Diet-Fed Mice Model. Foods 2021, 10, 2252. [Google Scholar] [CrossRef]
- Sheng, K.; Zhang, G.; Sun, M.; He, S.; Kong, X.; Wang, J.; Zhu, F.; Zha, X.; Wang, Y. Grape seed proanthocyanidin extract ameliorates dextran sulfate sodium-induced colitis through intestinal barrier improvement, oxidative stress reduction, and inflammatory cytokines and gut microbiota modulation. Food Funct. 2020, 11, 7817–7829. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Zhai, Z.; Wang, X.; Ding, Y.; He, L.; Li, L.; Mo, Q.; Mu, C.; Xie, R.; Liu, T.; et al. Targeting Desulfovibrio vulgaris flagellin-induced NAIP/NLRC4 inflammasome activation in macrophages attenuates ulcerative colitis. J. Adv. Res. 2023, 52, 219–232. [Google Scholar] [CrossRef]
- Van Hecke, T.; De Vrieze, J.; Boon, N.; De Vos, W.H.; Vossen, E.; De Smet, S. Combined Consumption of Beef-Based Cooked Mince and Sucrose Stimulates Oxidative Stress, Cardiac Hypertrophy, and Colonic Outgrowth of Desulfovibrionaceae in Rats. Mol. Nutr. Food Res. 2019, 63, e1800962. [Google Scholar] [CrossRef]
- Shao, X.; Liu, L.; Zhou, Y.; Zhong, K.; Gu, J.; Hu, T.; Yao, Y.; Zhou, C.; Chen, W. High-fat diet promotes colitis-associated tumorigenesis by altering gut microbial butyrate metabolism. Int. J. Biol. Sci. 2023, 19, 5004–5019. [Google Scholar] [CrossRef]
- Kong, C.; Gao, R.; Yan, X.; Huang, L.; Qin, H. Probiotics improve gut microbiota dysbiosis in obese mice fed a high-fat or high-sucrose diet. Nutrition 2019, 60, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhang, S.; Zhong, R.; Wan, F.; Chen, L.; Liu, L.; Yi, B.; Zhang, H. Olive Fruit Extracts Supplement Improve Antioxidant Capacity via Altering Colonic Microbiota Composition in Mice. Front. Nutr. 2021, 8, 645099. [Google Scholar] [CrossRef]
- Wu, T.; Zhang, Y.; Li, W.; Zhao, Y.; Long, H.; Muhindo, E.M.; Liu, R.; Sui, W.; Li, Q.; Zhang, M. Lactobacillus rhamnosus LRa05 Ameliorate Hyperglycemia through a Regulating Glucagon-Mediated Signaling Pathway and Gut Microbiota in Type 2 Diabetic Mice. J. Agric. Food Chem. 2021, 69, 8797–8806. [Google Scholar] [CrossRef] [PubMed]
- Dang, Y.; Ma, C.; Chen, K.; Chen, Y.; Jiang, M.; Hu, K.; Li, L.; Zeng, Z.; Zhang, H. The Effects of a High-Fat Diet on Inflammatory Bowel Disease. Biomolecules 2023, 13, 905. [Google Scholar] [CrossRef] [PubMed]
- Yamane, T.; Handa, S.; Imai, M.; Harada, N.; Sakamoto, T.; Ishida, T.; Nakagaki, T.; Nakano, Y. Exopolysaccharides from a Scandinavian fermented milk viili increase butyric acid and Muribaculum members in the mouse gut. Food Chem. Mol. Sci. 2021, 3, 100042. [Google Scholar] [CrossRef]
- Yan, S.; Yang, B.; Zhao, J.; Zhao, J.; Stanton, C.; Ross, R.P.; Zhang, H.; Chen, W. A ropy exopolysaccharide producing strain Bifidobacterium longum subsp. longum YS108R alleviates DSS-induced colitis by maintenance of the mucosal barrier and gut microbiota modulation. Food Funct. 2019, 10, 1595–1608. [Google Scholar] [CrossRef]
- Zhao, Q.; Hou, D.; Fu, Y.; Xue, Y.; Guan, X.; Shen, Q. Adzuki Bean Alleviates Obesity and Insulin Resistance Induced by a High-Fat Diet and Modulates Gut Microbiota in Mice. Nutrients 2021, 13, 3240. [Google Scholar] [CrossRef]
- Sun, Q.; Ji, Y.; Wang, Z.; She, X.; He, Y.; Ai, Q.; Li, L. Sodium Butyrate Alleviates Intestinal Inflammation in Mice with Necrotizing Enterocolitis. Mediat. Inflamm. 2021, 2021, 6259381. [Google Scholar] [CrossRef]
- Pang, Y.; Zheng, Y.; Yang, N.; Zan, M.; Zhang, L.; Ding, W. Potential novel biomarkers in small intestine for obesity/obesity resistance revealed by multi-omics analysis. Lipids Health Dis. 2022, 21, 98. [Google Scholar] [CrossRef]
- Henning, S.M.; Yang, J.; Hsu, M.; Lee, R.P.; Grojean, E.M.; Ly, A.; Tseng, C.H.; Heber, D.; Li, Z. Decaffeinated green and black tea polyphenols decrease weight gain and alter microbiome populations and function in diet-induced obese mice. Eur. J. Nutr. 2018, 57, 2759–2769. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.B.; Gonzalez, E.L.; Choy, K.; Faull, K.F.; Jewell, T.; Arellano, A.; Liang, J.; Yu, K.B.; Paramo, J.; Hsiao, E.Y. Gut microbiota Turicibacter strains differentially modify bile acids and host lipids. Nat. Commun. 2023, 14, 3669. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Zhuang, L.; Gao, S.; Ma, H.; Cheng, J.; Liu, J.; Liu, D.; Fu, S.; Hu, G. Orally Administered Ginkgolide C Attenuates DSS-Induced Colitis by Maintaining Gut Barrier Integrity, Inhibiting Inflammatory Responses, and Regulating Intestinal Flora. J. Agric. Food Chem. 2022, 70, 14718–14731. [Google Scholar] [CrossRef] [PubMed]
- Moorthy, M.; Sundralingam, U.; Palanisamy, U.D. Polyphenols as Prebiotics in the Management of High-Fat Diet-Induced Obesity: A Systematic Review of Animal Studies. Foods 2021, 10, 299. [Google Scholar] [CrossRef]
- Huang, Z.R.; Chen, M.; Guo, W.L.; Li, T.T.; Liu, B.; Bai, W.D.; Ai, L.Z.; Rao, P.F.; Ni, L.; Lv, X.C. Monascus purpureus-fermented common buckwheat protects against dyslipidemia and non-alcoholic fatty liver disease through the regulation of liver metabolome and intestinal microbiome. Food Res. Int. 2020, 136, 109511. [Google Scholar] [CrossRef]
- Kaur, N.; Chen, C.; Luther, J.; Kao, J.Y. Intestinal dysbiosis in inflammatory bowel disease. Gut Microbes 2011, 2, 211–216. [Google Scholar] [CrossRef]
- Wang, W.; An, Q.; Zou, Y.; Dai, Y.; Meng, Q.; Zhang, Y. Lactoferrin alleviates the adverse effects of early-life inflammation on depression in adults by regulating the activation of microglia. Mol. Med. 2025, 31, 50. [Google Scholar] [CrossRef]
- Grodner, B.; Shi, H.; Farchione, O.; Vill, A.C.; Ntekas, I.; Diebold, P.J.; Wu, D.T.; Chen, C.Y.; Kim, D.M.; Zipfel, W.R.; et al. Spatial mapping of mobile genetic elements and their bacterial hosts in complex microbiomes. Nat. Microbiol. 2024, 9, 2262–2277. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | HFD | DSS | ||
|---|---|---|---|---|
| wt-wt | ko-wt | wt-wt | ko-wt | |
| Alistipes | ↑ | - | ↑ ↑ | ↑ ↑ |
| Colidextribacter | ↑ | - | ↑ - | ↑ ↑ |
| Mucispirillum | ↑ | - | ↑ ↑ | ↑ ↑ |
| Odoribacter | ↑ | - | ↑ ↑ | ↑ ↑ |
| Oscillibacter | ↑ | - | ↑ ↑ | ↑ ↑ |
| Monoglobus | ↓ | - | ↓ - | - - |
| Muribaculum | ↓ | - | ↓ - | ↓ ↓ |
| Eubacterium_xylanophilum_group | ↓ | ↓ | ↓ ↓ | ↓ ↓ |
| Lachnoclostridium | - | ↓ | ↓ ↓ | ↓ ↓ |
| Lachnospiraceae_UCG-06 | - | ↓ | - ↓ | ↓ - |
| Staphylococcus | - | ↓ | ↑ ↓ | ↓ - |
| Clostridium_sensu_stricto_1 | ↓ | ↓ | ↑ - | ↑ - |
| Turicibacter | ↓ | ↓ | ↑ ↑ | ↑ ↑ |
| NK4A214_group | ↑ | ↑ | ↓ - | ↓ ↓ |
| Roseburia | ↑ | ↑ | ↓ ↓ | ↓ ↓ |
| GCA-900066575 | - | ↑ | - ↓ | ↓ - |
| Anaerotruncus | ↑ | ↑ | - ↓ | ↓ ↓ |
| Genus | Age | DSS | ||
|---|---|---|---|---|
| wt-wt | ko-wt | wt-wt | ko-wt | |
| Candidatus_Arthromitus | ↑↑ | ↑↑ | ↓↓ | ↓↓ |
| Candidatus_Saccharimonas | ↑↑ | ↑↑ | ↓- | -↓ |
| Enterorhabdus | ↑↑ | ↑↑ | ↓↓ | ↓↓ |
| Eubacterium_xylanophilum_group | ↑↑ | -- | ↓↓ | ↓↓ |
| Jeotgalicoccus | ↑- | ↑- | ↓↓ | ↓- |
| Kurthia | ↑- | ↑- | ↓- | ↓- |
| Monoglobus | ↑↑ | -- | ↓- | -- |
| Bacteroides | ↓↓ | ↓↓ | ↑- | ↑↑ |
| Mucispirillum | ↓↓ | -- | ↑↑ | ↑↑ |
| Oscillibacter | ↓↓ | ↓↓ | ↑↑ | ↑↑ |
| Parabacteroides | ↓↓ | -- | ↑↑ | ↑↑ |
| Romboutsia | ↓- | -- | ↑↑ | ↑- |
| Turicibacter | ↓↓ | -- | ↑↑ | ↑↑ |
| Aerococcus | -- | ↑- | ↓- | ↓- |
| Desulfovibrio | -↑ | ↑↑ | -- | ↓↓ |
| Eubacterium_brachy_group | -- | ↑↑ | ↓↓ | ↓↓ |
| Corynebacterium | -- | ↑- | -- | ↓- |
| Lachnospiraceae_UCG-06 | -↑ | ↑- | -↓ | ↓- |
| Lactobacillus | -↑ | ↑↑ | ↓↓ | ↓- |
| Lysinibacillus | -- | ↑- | -- | ↓- |
| Microbacterium | -- | ↑- | -- | ↓- |
| Staphylococcus | ↑↑ | ↑- | ↑↓ | ↓- |
| Colidextribacter | -- | ↓↓ | ↑- | ↑↑ |
| Odoribacter | -- | ↓↓ | ↑↑ | ↑↑ |
| Rikenella | ↓- | ↓- | -- | ↑↑ |
| Lachnoclostridium | -- | -↑ | ↓↓ | ↓↓ |
| Genus | Age | HFD | ||
|---|---|---|---|---|
| wt-wt | ko-wt | wt-wt | ko-wt | |
| Eubacterium_xylanophilum_group | ↑ ↑ | - - | ↓ | ↓ |
| Monoglobus | ↑ ↑ | - - | ↓ | - |
| Anaerotruncus | ↓ ↓ | ↓ ↓ | ↑ | ↑ |
| Bilophila | ↓ ↓ | ↓ ↓ | ↑ | - |
| Blautia | ↓ ↓ | - - | ↑ | - |
| Mucispirillum | ↓ ↓ | - - | ↑ | - |
| Oscillibacter | ↓ ↓ | ↓ ↓ | ↑ | - |
| Rikenella | ↓ - | ↓ - | ↑ | - |
| Roseburia | ↓ - | ↓ - | ↑ | ↑ |
| Muribaculum | ↓ ↓ | ↓ ↓ | ↓ | - |
| Turicibacter | ↓ ↓ | - - | ↓ | ↓ |
| Dubosiella | - - | ↑ ↑ | ↓ | ↓ |
| Lachnospiraceae_UCG-06 | - ↑ | ↑ - | - | ↓ |
| Staphylococcus | ↑ ↑ | ↑ - | - | ↓ |
| GCA-900066575 | ↓ ↓ | ↓ ↓ | - | ↑ |
| Age | HFD | DSS | ||||
|---|---|---|---|---|---|---|
| wt-wt | ko-wt | wt-wt | ko-wt | wt-wt | ko-wt | |
| Firmicutes | ↑ | ↑ | - | ↑ | ↓ | ↓ |
| Bacteroidota | ↓ | ↓ | - | ↓ | ↑ | ↑ |
| Actinobacteriota | ↑ | ↑ | ↓ | ↓ | ↓ | ↓ |
| Cyanobacteria | ↑ | ↑ | ↓ | - | ↑ | ↑ |
| Deferribacterota | ↓ | ↓ | ↑ | - | ↑ | ↑ |
| Campilobacterota | ↓ | ↓ | ↑ | - | ↑ | ↑ |
| Patescibacteria | ↑ | ↑ | - | - | - | ↓ |
| Acidobacteriota | ↓ | ↓ | - | - | - | - |
| Desulfobacterota | - | - | - | - | - | ↓ |
| Proteobacteria | - | - | - | - | ↑ | ↑ |
| Verrucomicrobiota | - | - | ↑ | ↑ | - | ↑ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; An, Q.; Zou, Y.; Meng, Q.; Zhang, Y. Effects of Lactation Lactoferrin Deficiency on Intestinal Microbiota in Different Mice Models. Nutrients 2025, 17, 2248. https://doi.org/10.3390/nu17132248
Wang W, An Q, Zou Y, Meng Q, Zhang Y. Effects of Lactation Lactoferrin Deficiency on Intestinal Microbiota in Different Mice Models. Nutrients. 2025; 17(13):2248. https://doi.org/10.3390/nu17132248
Chicago/Turabian StyleWang, Wenli, Qin An, Yunxia Zou, Qingyong Meng, and Yali Zhang. 2025. "Effects of Lactation Lactoferrin Deficiency on Intestinal Microbiota in Different Mice Models" Nutrients 17, no. 13: 2248. https://doi.org/10.3390/nu17132248
APA StyleWang, W., An, Q., Zou, Y., Meng, Q., & Zhang, Y. (2025). Effects of Lactation Lactoferrin Deficiency on Intestinal Microbiota in Different Mice Models. Nutrients, 17(13), 2248. https://doi.org/10.3390/nu17132248