
Metabolomics Analysis Reveals Molecular Signatures of Metabolic Complexity in Children with Hypercholesterolemia

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Serum Cholesterol and Lipid Profile Measurement
2.3. H–NMR Spectroscopy
2.4. NMR Data Processing and Analysis
2.5. Statistical Analysis
3. Results
3.1. Population Characteristics
3.2. Identification of Blood Metabolites for the Different Cholesterol Statuses
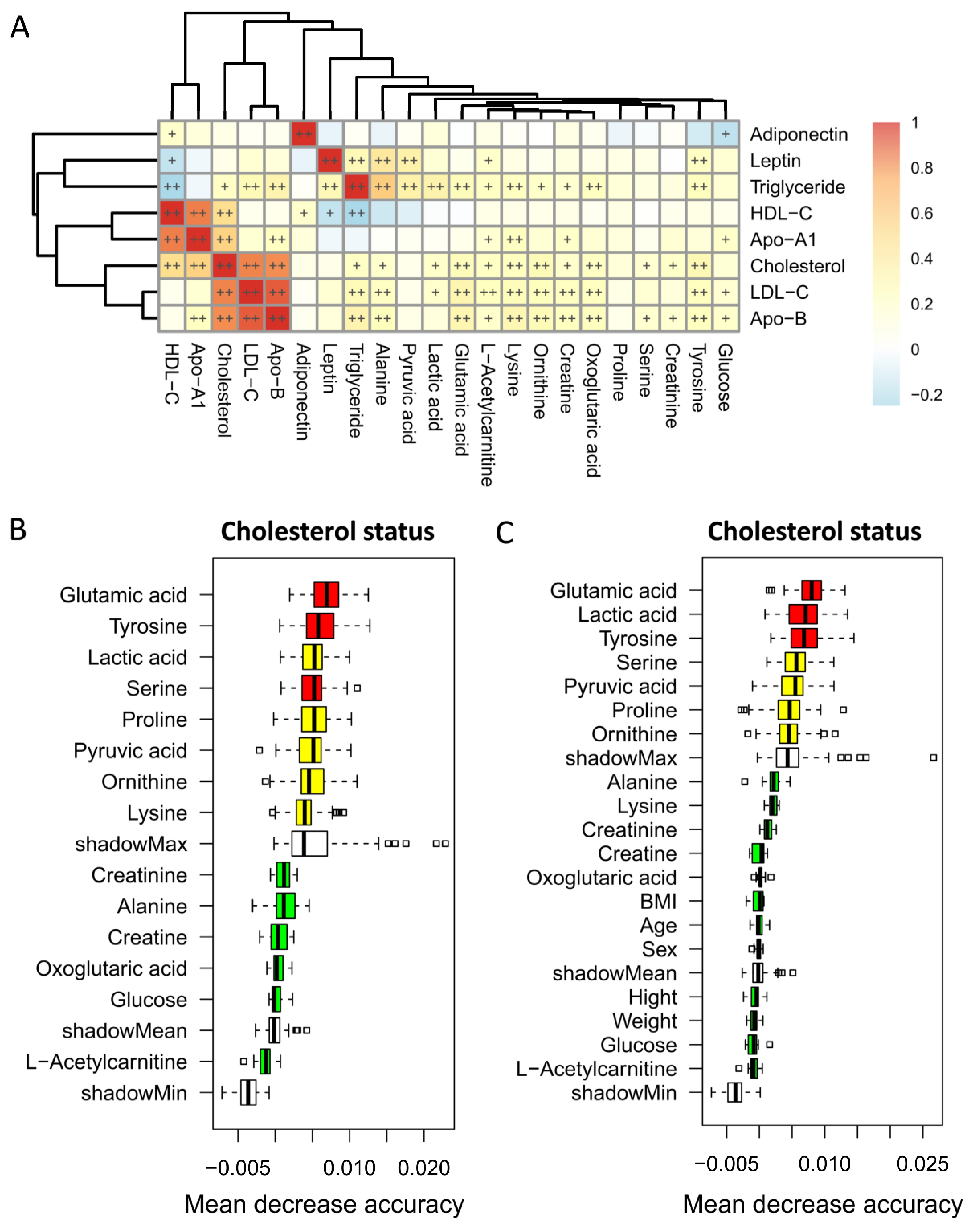
3.3. Correlation between Blood Metabolites and Lipid Profiles
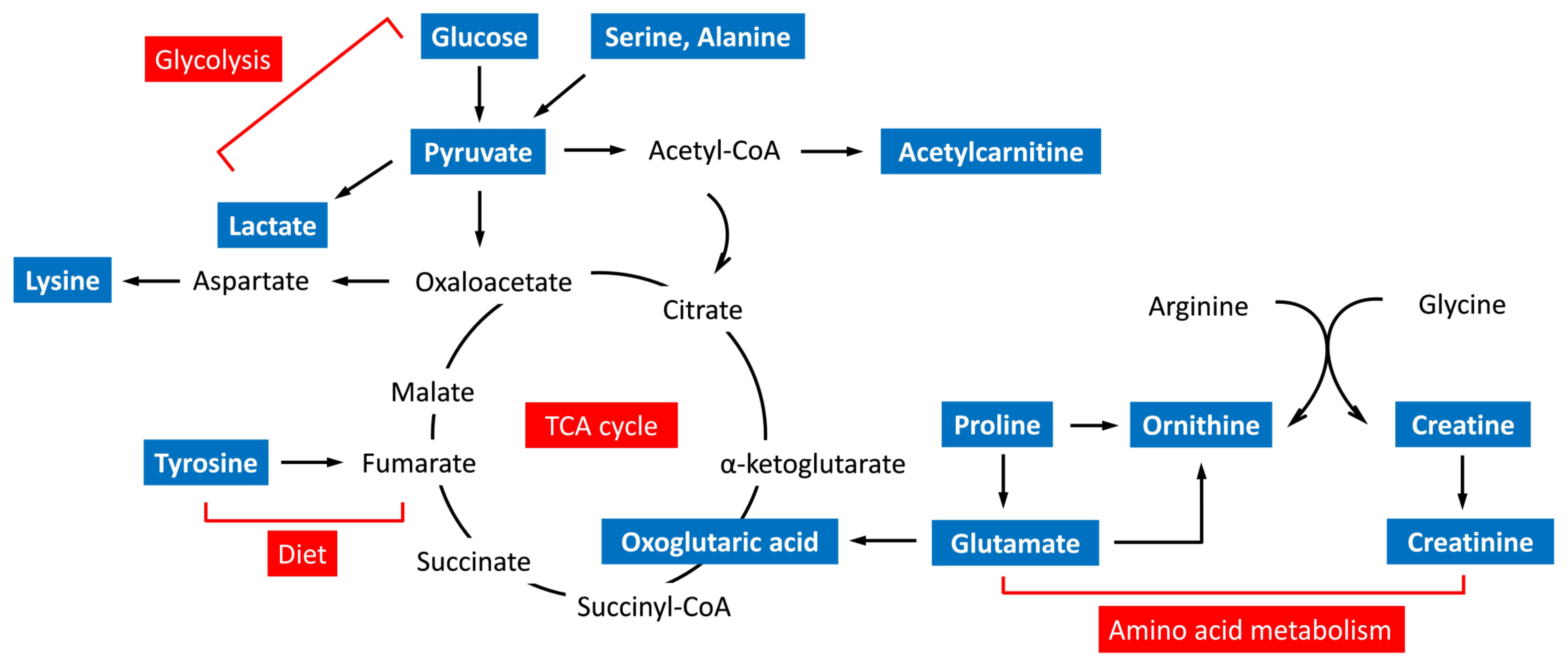
3.4. Metabolic Pathway and Function Analysis
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McGill, H.C., Jr.; McMahan, C.A.; Herderick, E.E.; Malcom, G.T.; Tracy, R.E.; Strong, J.P. Origin of atherosclerosis in childhood and adolescence. Am. J. Clin. Nutr. 2000, 72, 1307s–1315s. [Google Scholar] [CrossRef] [PubMed]
- Daniels, S.R.; Greer, F.R. Lipid screening and cardiovascular health in childhood. Pediatrics 2008, 122, 198–208. [Google Scholar] [CrossRef] [PubMed]
- Lozano, P.; Henrikson, N.B.; Morrison, C.C.; Dunn, J.; Nguyen, M.; Blasi, P.R.; Whitlock, E.P. Lipid Screening in Childhood and Adolescence for Detection of Multifactorial Dyslipidemia: Evidence Report and Systematic Review for the US Preventive Services Task Force. JAMA 2016, 316, 634–644. [Google Scholar] [CrossRef] [PubMed]
- Jacobs, D.R.; Woo, J.G.; Sinaiko, A.R.; Daniels, S.R.; Ikonen, J.; Juonala, M.; Kartiosuo, N.; Lehtimäki, T.; Magnussen, C.G.; Viikari, J.S.A.; et al. Childhood Cardiovascular Risk Factors and Adult Cardiovascular Events. N. Engl. J. Med. 2022, 386, 1877–1888. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Chang, P.Y.; Zhang, Y.; Kizer, J.R.; Best, L.G.; Howard, B.V. Triglyceride and HDL-C Dyslipidemia and Risks of Coro nary Heart Disease and Ischemic Stroke by Glycemic Dysregulation Status: The Strong Heart Study. Diabetes Care 2017, 40, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Behbodikhah, J.; Ahmed, S.; Elyasi, A.; Kasselman, L.J.; De Leon, J.; Glass, A.D.; Reiss, A.B. Apolipoprotein B and Cardiovascular Disease: Biomarker and Potential Therapeutic Target. Metabolites 2017, 11, 690. [Google Scholar] [CrossRef]
- de Ferranti, S.D.; Rodday, A.M.; Mendelson, M.M.; Wong, J.B.; Leslie, L.K.; Sheldrick, R.C. Prevalence of Familial Hypercholesterolemia in the 1999 to 2012 United States National Health and Nutrition Examination Surveys (NHANES). Circulation 2016, 133, 1067–1072. [Google Scholar] [CrossRef]
- Christensen, J.J.; Ulven, S.M.; Retterstøl, K.; Narverud, I.; Bogsrud, M.P.; Henriksen, T.; Bollerslev, J.; Halvorsen, B.; Aukrust, P.; Holven, K.B. Comprehensive lipid and metabolite profiling of children with and without familial hypercholesterolemia: A cross-sectional study. Atherosclerosis 2017, 266, 48–57. [Google Scholar] [CrossRef]
- Soliman, G.A. Dietary Cholesterol and the Lack of Evidence in Cardiovascular Disease. Nutrients 2018, 10, 780. [Google Scholar] [CrossRef]
- Collado, A.; Domingo, E.; Piqueras, L.; Sanz, M.J. Primary hypercholesterolemia and development of cardiovascular disorders: Cellular and molecular mechanisms involved in low-grade systemic inflammation and endothelial dysfunction. Int. J. Biochem. Cell Biol. 2021, 139, 106066. [Google Scholar] [CrossRef]
- Yu, L.; Zhu, Q.; Li, Y.; Song, P.; Zhang, J. Dietary Branched-Chain Amino Acids (BCAAs) and Risk of Dyslipidemia in a Chinese Population. Nutrients 2022, 14, 1824. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Zhang, A.H.; Miao, J.H.; Sun, H.; Han, Y.; Yan, G.L.; Wu, F.F.; Wang, X.J. Metabolomics biotechnology, applications, and future trends: A systematic review. RSC Adv. 2019, 9, 37245–37257. [Google Scholar] [CrossRef] [PubMed]
- Medina, S.; Dominguez-Perles, R.; Gil, J.I.; Ferreres, F.; Gil-Izquierdo, A. Metabolomics and the diagnosis of human diseases a guide to the markers and pathophysiological pathways affected. Curr. Med. Chem. 2014, 21, 823–848. [Google Scholar] [CrossRef] [PubMed]
- Grundy, S.M.; Stone, N.J.; Bailey, A.L.; Beam, C.; Birtcher, K.K.; Blumenthal, R.S.; Braun, L.T.; Ferranti, S.d.; Faiella-Tommasino, J.; Forman, D.E.; et al. 2018 AHA/ACC/AACVPR/AAPA/ABC/ACPM/ADA/AGS/APhA/ASPC/NLA/PCNA Guideline on the Management of Blood Cholesterol. J. Am. Coll. Cardiol. 2019, 73, e285–e350. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.-Y.; Lin, G.; Cheng, M.-L.; Chiang, M.-H.; Tsai, M.-H.; Lai, S.-H.; Wong, K.-S.; Hsieh, S.-Y. Metabolomic Profiling of Infectious Parapneumonic Effusions Reveals Biomarkers for Guiding Management of Children with Streptococcus pneumoniae Pneumonia. Sci. Rep. 2016, 6, 24930. [Google Scholar] [CrossRef] [PubMed]
- Jacob, D.; Deborde, C.; Lefebvre, M.; Maucourt, M.; Moing, A. NMRProcFlow: A graphical and interactive tool dedicated to 1D spectra processing for NMR-based metabolomics. Metabolomics 2017, 13, 36. [Google Scholar] [CrossRef]
- de Ferranti, S.D.; Steinberger, J.; Ameduri, R.; Baker, A.; Gooding, H.; Kelly, A.S.; Mietus-Snyder, M.; Mitsnefes, M.M.; Peterson, A.L.; St-Pierre, J.; et al. Cardiovascular Risk Reduction in High-Risk Pediatric Patients: A Scientific Statement From the American Heart Association. Circulation 2019, 139, e603–e634. [Google Scholar] [CrossRef]
- Schienkiewitz, A.; Truthmann, J.; Ernert, A.; Wiegand, S.; Schwab, K.O.; Scheidt-Nave, C. Age, Maturation and Serum Lipid Parameters: Findings from The German Health Survey for Children and Adolescents. BMC Public Health 2019, 19, 1627. [Google Scholar] [CrossRef]
- Kuo, P.; Syu, J.-T.; Tzou, I.L.; Chen, P.-Y.; Su, H.-Y.; Chu, N.-F. Prevalence and trend of dyslipidaemia from 1996 to 2006 among normal and overweight adolescents in Taiwan. BMJ Open 2014, 4, e003800. [Google Scholar] [CrossRef]
- Hickman, T.B.; Briefel, R.R.; Carroll, M.D.; Rifkind, B.M.; Cleeman, J.I.; Maurer, K.R.; Johnson, C.L. Distributions and trends of serum lipid levels among United States children and adolescents ages 4-19 years: Data from the Third National Health and Nutrition Examination Survey. Prev Med. 1998, 27, 879–890. [Google Scholar] [CrossRef]
- Labarthe, D.R.; Dai, S.; Day, R.S.; Fulton, J.E.; Grunbaum, J.A.; Shah, S.M.; Wen, E. Project HeartBeat! Concept, development, and design. Am. J. Prev. Med. 2009, 37, S9–S16. [Google Scholar] [CrossRef] [PubMed]
- Kreisberg, R.A.; Kasim, S. Cholesterol metabolism and aging. Am. J. Med. 1987, 82, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Robinson, G.A.; Peng, J.; Peckham, H.; Radziszewska, A.; Butler, G.; Pineda-Torra, I.; Jury, E.C.; Ciurtin, C. Sex hormones drive changes in lipoprotein metabolism. iScience 2021, 24, 103257. [Google Scholar] [CrossRef]
- Xiang, A.S.; Kingwell, B.A. Rethinking good cholesterol: A clinicians’ guide to understanding HDL. Lancet Diabetes Endocrinol. 2019, 7, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-F.; Zhang, Y.-Z. Targeted profiling of amino acid metabolome in serum by a liquid chromatography-mass spectrometry method: Application to identify potential markers for diet-induced hyperlipidemia. Anal. Methods 2020, 12, 2355–2362. [Google Scholar] [CrossRef]
- Xu, D.; Wang, S.; Feng, M.; Shete, V.; Chu, Y.; Kamil, A.; Yang, C.; Liu, H.; Xia, H.; Wang, X.; et al. Serum Metabolomics Reveals Underlying Mechanisms of Cholesterol-Lowering Effects of Oat Consumption: A Randomized Controlled Trial in a Mildly Hypercholesterolemic Population. Mol. Nutr.Food Res. 2021, 65, e2001059. [Google Scholar] [CrossRef]
- Maltais-Payette, I.; Boulet, M.-M.; Prehn, C.; Adamski, J.; Tchernof, A. Circulating glutamate concentration as a biomarker of visceral obesity and associated metabolic alterations. Nutr. Metab. 2018, 15, 78. [Google Scholar] [CrossRef]
- Boulet, M.M.; Chevrier, G.; Grenier-Larouche, T.; Pelletier, M.; Nadeau, M.; Scarpa, J.; Prehn, C.; Marette, A.; Adamski, J.; Tchernof, A. Alterations of plasma metabolite profiles related to adipose tissue distribution and cardiometabolic risk. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E736–E746. [Google Scholar] [CrossRef]
- Yamakado, M.; Tanaka, T.; Nagao, K.; Ishizaka, Y.; Mitushima, T.; Tani, M.; Toda, A.; Toda, E.; Okada, M.; Miyano, H.; et al. Plasma amino acid profile is associated with visceral fat accumulation in obese Japanese subjects. Clin. Obes. 2012, 2, 29–40. [Google Scholar] [CrossRef]
- Liu, L.; Feng, J.; Zhang, G.; Yuan, X.; Li, F.; Yang, T.; Hao, S.; Huang, D.; Hsue, C.; Lou, Q. Visceral adipose tissue is more strongly associated with insulin resistance than subcutaneous adipose tissue in Chinese subjects with pre-diabetes. Curr. Med. Res. Opin. 2018, 34, 123–129. [Google Scholar] [CrossRef]
- Hellmuth, C.; Kirchberg, F.F.; Lass, N.; Harder, U.; Peissner, W.; Koletzko, B.; Reinehr, T. Tyrosine Is Associated with Insulin Resistance in Longitudinal Metabolomic Profiling of Obese Children. J. Diabetes Res. 2016, 2016, 2108909. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, S.; Miyazaki, H.; Oda, H.; Aoyama, Y.; Yoshida, A. Effects of excess dietary tyrosine on cholesterol, bile acid metabolism and mixed-function oxidase system in rats. J. Nutr. 1990, 120, 1134–1139. [Google Scholar] [CrossRef]
- Jacques, H.; Deshaies, Y.; Savoie, L. Relationship between dietary tyrosine and plasma cholesterol in the rat. Can. J. Physiol. Pharmacol. 1988, 66, 1023–1027. [Google Scholar] [CrossRef] [PubMed]
- Chris, D.W.M.B.; Adams, N. Impacts of dietary tyrosine on serum cholesterol portions in rodents. Adv. Res. J. Biochem. 2019, 4, 179–185. [Google Scholar]
- Akram, M. Citric Acid Cycle and Role of its Intermediates in Metabolism. Cell Biochem. Biophys. 2013, 68, 475–478. [Google Scholar] [CrossRef] [PubMed]
- Bays, H.E.; Toth, P.P.; Kris-Etherton, P.M.; Abate, N.; Aronne, L.J.; Brown, W.V.; Gonzalez-Campoy, J.M.; Jones, S.R.; Kumar, R.; La Forge, R.; et al. Obesity, adiposity, and dyslipidemia: A consensus statement from the National Lipid Association. J. Clin. Lipidol. 2013, 7, 304–383. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Characteristics | Acceptable (n = 42) | Borderline (n = 52) | High (n = 31) | p-Values |
|---|---|---|---|---|
| Age (yr) | 7.12 ± 0.25 | 7.20 ± 0.31 | 7.12 ± 0.06 | 0.683 |
| Sex, male | 27 (64.3%) | 31 (60.8%) | 11 (35.5%) | 0.031 |
| Height (m) | 1.23 ± 0.06 | 1.24 ± 0.05 | 1.23 ± 0.06 | 0.829 |
| Weight (kg) | 24.92 ± 6.77 | 25.08 ± 5.31 | 23.96 ± 5.63 | 0.574 |
| BMI (kg/m2) | 16.33 ± 3.52 | 16.29 ± 2.50 | 15.82 ± 2.84 | 0.406 |
| Obesity | 5 (12%) | 7 (13%) | 6 (19%) | 0.648 |
| Lipid profile | ||||
| Cholesterol | 151.64 ± 16.92 | 185.25 ± 8.48 | 223.72 ± 21.45 | <0.001 |
| TG | 47.55 ± 22.68 | 57.60 ± 36.90 | 69.28 ± 34.86 | 0.010 |
| LDL | 82.92 ± 14.46 | 103.9 ± 14.92 | 130.28 ± 12.48 | <0.001 |
| HDL | 52.33 ± 9.10 | 60.99 ±12.74 | 68.89 ± 13.22 | <0.001 |
| Apo-A1 | 134.64 ± 18.05 | 148.85 ± 18.42 | 165.48 ± 20.95 | <0.001 |
| Apo-B | 56.95 ± 9.92 | 70.87 ±9.47 | 87.17 ± 11.02 | <0.001 |
| Adiponectin | 21.62 + 12.02 | 20.53 ± 9.17 | 23.51 ± 10.30 | 0.275 |
| Leptin | 5.07 ± 7.03 | 6.25 ± 8.58 | 5.53 ± 5.93 | 0.571 |
| High vs. Acceptable | Borderline vs. Acceptable | High vs. Borderline | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Metabolites | Chemical Shift, ppm (Multiplicity) | VIP Score * | Fold Change † | p ‡ | VIP Score | Fold Change | p | VIP Score | Fold Change | p |
| Tyrosine | 6.860−6.917(dt) | 2.26 | 1.16 | 0.000 | 1.62 | 1.14 | 0.004 | 0.85 | 1.02 | 0.455 |
| Glutamic acid | 2.094−2.130(m) | 1.47 | 1.08 | 0.001 | 1.37 | 1.10 | 0.002 | 0.70 | 0.98 | 0.774 |
| Ornithine | 3.041−3.075(t) | 1.65 | 1.10 | 0.004 | 1.73 | 1.13 | 0.001 | 0.75 | 0.98 | 0.646 |
| Lysine | 3.010−3.030(t) | 1.30 | 1.07 | 0.007 | 1.33 | 1.09 | 0.003 | 0.66 | 0.98 | 0.683 |
| Alanine | 1.455−1.490(q) | 1.69 | 1.14 | 0.013 | 1.86 | 1.18 | 0.002 | 0.64 | 0.96 | 0.612 |
| Creatinine | 3.036−3.041(s) | 1.16 | 1.06 | 0.013 | 1.26 | 1.08 | 0.004 | 0.60 | 0.98 | 0.571 |
| Lactic acid | 1.304−1.340(t) | 1.79 | 1.14 | 0.020 | 0.99 | 1.12 | 0.168 | 1.10 | 1.02 | 0.434 |
| Oxoglutaric acid | 2.980−3.010(dt) | 1.10 | 1.06 | 0.024 | 1.24 | 1.08 | 0.005 | 0.76 | 0.98 | 0.568 |
| L-Acetylcarnitine | 3.182−3.187(d) | 1.22 | 1.08 | 0.029 | 0.87 | 1.06 | 0.079 | 0.99 | 1.01 | 0.648 |
| Creatine | 3.917−3.926(s) | 1.18 | 1.08 | 0.044 | 1.31 | 1.11 | 0.013 | 0.68 | 0.97 | 0.661 |
| Serine | 3.937−3.957(q) | 0.85 | 1.05 | 0.085 | 1.36 | 1.11 | 0.005 | 1.13 | 0.94 | 0.219 |
| Glucose | 4.622−4.656(d) | 0.85 | 1.05 | 0.099 | 1.04 | 1.07 | 0.026 | 0.85 | 0.98 | 0.563 |
| Pyruvic acid | 2.358−2.372(s) | 0.71 | 1.06 | 0.301 | 1.69 | 1.17 | 0.005 | 1.48 | 0.91 | 0.134 |
| Proline | 3.309−3.339(m) | 0.60 | 1.04 | 0.359 | 1.89 | 1.18 | 0.001 | 2.00 | 0.88 | 0.031 |
| Pathway Names | Total | Hits | Matched Metabolites | RawP | FDR | Function |
|---|---|---|---|---|---|---|
| Arginine and proline metabolism | 38 | 5 | Creatine, Proline, Pyruvate Glutamic acid, Ornithine | 1.700 × 10−5 | 0.001 | Amino acid metabolism |
| Alanine, aspartate, and glutamate metabolism | 28 | 4 | Alanine, Glutamic acid, Pyruvate, Oxoglutaric acid | 1.016 × 10−4 | 0.003 | Amino acid metabolism |
| Arginine biosynthesis | 14 | 3 | Glutamic acid, Ornithine, Oxoglutaric acid | 2.507 × 10−4 | 0.005 | Amino acid metabolism |
| D-Glutamine and D-glutamate metabolism | 6 | 2 | Glutamic acid, Oxoglutaric acid | 1.282 × 10−3 | 0.021 | Amino acid metabolism |
| Glycolysis/gluconeogenesis | 26 | 3 | Pyruvate, Lactic acid, Glucose | 1.669 × 10−3 | 0.023 | Carbohydrate metabolism |
| Butanoate metabolism | 15 | 2 | Glutamic acid, Oxoglutaric acid | 8.538 × 10−3 | 0.102 | Carbohydrate metabolism |
| Citrate cycle (TCA cycle) | 20 | 2 | Oxoglutaric acid, Pyruvate | 1.502 × 10−2 | 0.157 | Carbohydrate metabolism |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, P.-S.; Su, K.-W.; Yeh, K.-W.; Huang, J.-L.; Lo, F.-S.; Chiu, C.-Y. Metabolomics Analysis Reveals Molecular Signatures of Metabolic Complexity in Children with Hypercholesterolemia. Nutrients 2023, 15, 1726. https://doi.org/10.3390/nu15071726
Gu P-S, Su K-W, Yeh K-W, Huang J-L, Lo F-S, Chiu C-Y. Metabolomics Analysis Reveals Molecular Signatures of Metabolic Complexity in Children with Hypercholesterolemia. Nutrients. 2023; 15(7):1726. https://doi.org/10.3390/nu15071726
Chicago/Turabian StyleGu, Pei-Shin, Kuan-Wen Su, Kuo-Wei Yeh, Jing-Long Huang, Fu-Sung Lo, and Chih-Yung Chiu. 2023. "Metabolomics Analysis Reveals Molecular Signatures of Metabolic Complexity in Children with Hypercholesterolemia" Nutrients 15, no. 7: 1726. https://doi.org/10.3390/nu15071726
APA StyleGu, P.-S., Su, K.-W., Yeh, K.-W., Huang, J.-L., Lo, F.-S., & Chiu, C.-Y. (2023). Metabolomics Analysis Reveals Molecular Signatures of Metabolic Complexity in Children with Hypercholesterolemia. Nutrients, 15(7), 1726. https://doi.org/10.3390/nu15071726