
Isocaloric Diets with Different Protein-Carbohydrate Ratios: The Effect on Sleep, Melatonin Secretion and Subsequent Nutritional Response in Healthy Young Men

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
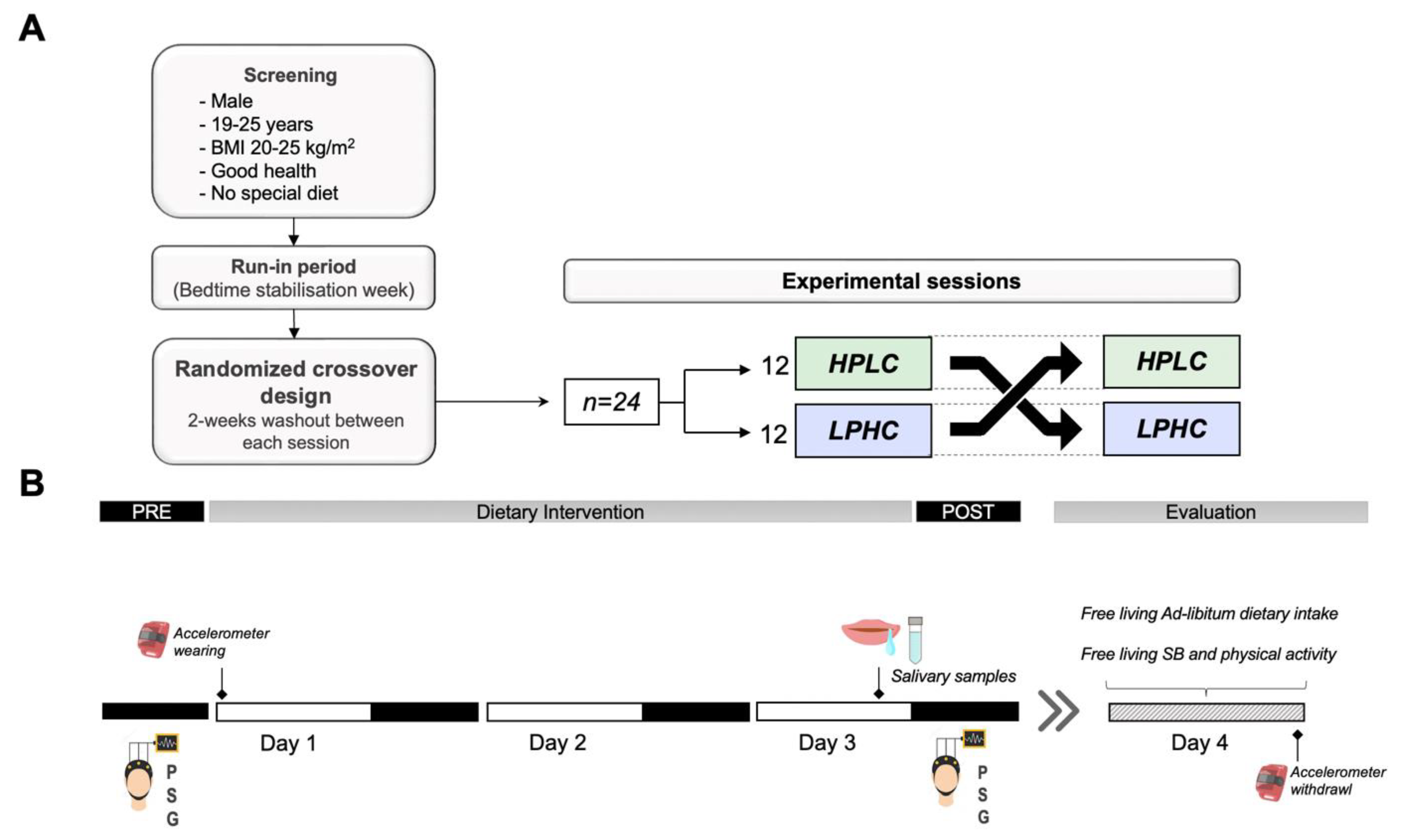
2.1. Participants
2.2. Study Procedure
2.3. Nutritional Intervention
2.4. Measurements
2.4.1. Anthropometric Characteristics and Body Composition
2.4.2. Physical Activity and Sedentary Time
2.4.3. Melatonin Profile/Phase Angles
2.4.4. Sleep
2.4.5. Ad-Libitum Dietary Intake
2.5. Statistical Analysis
3. Results
3.1. Participants
3.2. Melatonin Secretion
3.3. Sleep
3.4. Day 4 Free Living Ad-Libitum Dietary Intake
3.5. Day 4 Free Living Time Spent on Sedentary Behaviors and Physical Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Huang, W.; Ramsey, K.M.; Marcheva, B.; Bass, J. Circadian Rhythms, Sleep, and Metabolism. J. Clin. Investig. 2011, 121, 2133–2141. [Google Scholar] [CrossRef] [PubMed]
- St-Onge, M.-P. The Role of Sleep Duration in the Regulation of Energy Balance: Effects on Energy Intakes and Expenditure. J. Clin. Sleep Med. 2013, 9, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Morselli, L.; Leproult, R.; Balbo, M.; Spiegel, K. Role of Sleep Duration in the Regulation of Glucose Metabolism and Appetite. Best Pract. Res. Clin. Endocrinol. Metab. 2010, 24, 687–702. [Google Scholar] [CrossRef] [PubMed]
- St-Onge, M.-P.; Mikic, A.; Pietrolungo, C.E. Effects of Diet on Sleep Quality. Adv. Nutr. 2016, 7, 938–949. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.; Gonzalez, K.; Lee-Ang, L.; Young, M.C.; Tamez, M.; Mattei, J. Diet and Sleep Physiology: Public Health and Clinical Implications. Front. Neurol. 2017, 8, 393. [Google Scholar] [CrossRef]
- Zuraikat, F.M.; Wood, R.A.; Barragán, R.; St-Onge, M.-P. Sleep and Diet: Mounting Evidence of a Cyclical Relationship. Annu. Rev. Nutr. 2021, 41, 309–332. [Google Scholar] [CrossRef]
- Khan, M.K.; Faught, E.L.; Chu, Y.L.; Ekwaru, J.P.; Storey, K.E.; Veugelers, P.J. Is It Nutrients, Food Items, Diet Quality or Eating Behaviours That Are Responsible for the Association of Children’s Diet with Sleep? J. Sleep Res. 2017, 26, 468–476. [Google Scholar] [CrossRef]
- Wilson, K.; St-Onge, M.-P.; Tasali, E. Diet Composition and Objectively Assessed Sleep Quality: A Narrative Review. J. Acad. Nutr. Diet. 2022, 122, 1182–1195. [Google Scholar] [CrossRef]
- Du, C.; Almotawa, J.; Feldpausch, C.E.; Folk, S.Y.L.; Parag, H.; Tucker, R.M. Effects of Macronutrient Intake on Sleep Duration and Quality: A Systematic Review. Nutr. Diet. 2022, 79, 59–75. [Google Scholar] [CrossRef]
- Tanaka, E.; Yatsuya, H.; Uemura, M.; Murata, C.; Otsuka, R.; Toyoshima, H.; Tamakoshi, K.; Sasaki, S.; Kawaguchi, L.; Aoyama, A. Associations of Protein, Fat, and Carbohydrate Intakes with Insomnia Symptoms among Middle-Aged Japanese Workers. J. Epidemiol. 2013, 23, 132–138. [Google Scholar] [CrossRef]
- Spaeth, A.M.; Dinges, D.F.; Goel, N. Objective Measurements of Energy Balance Are Associated with Sleep Architecture in Healthy Adults. Sleep 2017, 40, zsw018. [Google Scholar] [CrossRef] [PubMed]
- Lindseth, G.; Lindseth, P.; Thompson, M. Nutritional Effects on Sleep. West. J. Nurs. Res. 2013, 35, 497–513. [Google Scholar] [CrossRef]
- Lindseth, G.; Murray, A. Dietary Macronutrients and Sleep. West J. Nurs. Res. 2016, 38, 938–958. [Google Scholar] [CrossRef]
- Zhou, J.; Kim, J.E.; Armstrong, C.L.; Chen, N.; Campbell, W.W. Higher-Protein Diets Improve Indexes of Sleep in Energy-Restricted Overweight and Obese Adults: Results from 2 Randomized Controlled Trials. Am. J. Clin. Nutr. 2016, 103, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Tan, X.; Alén, M.; Cheng, S.M.; Mikkola, T.M.; Tenhunen, J.; Lyytikäinen, A.; Wiklund, P.; Cong, F.; Saarinen, A.; Tarkka, I.; et al. Associations of Disordered Sleep with Body Fat Distribution, Physical Activity and Diet among Overweight Middle-Aged Men. J. Sleep Res. 2015, 24, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Afaghi, A.; O’Connor, H.; Chow, C.M. Acute Effects of the Very Low Carbohydrate Diet on Sleep Indices. Nutr. Neurosci. 2008, 11, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Phillips, F.; Crisp, A.H.; McGuinness, B.; Kalucy, E.C.; Chen, C.N.; Koval, J.; Kalucy, R.S.; Lacey, J.H. Isocaloric Diet Changes and Electroencephalographic Sleep. Lancet 1975, 306, 723–725. [Google Scholar] [CrossRef]
- Kwan, R.M.; Thomas, S.; Mir, M.A. Effects of a Low Carbohydrate Isoenergetic Diet on Sleep Behavior and Pulmonary Functions in Healthy Female Adult Humans. J. Nutr. 1986, 116, 2393–2402. [Google Scholar] [CrossRef]
- Nehme, P.; Marqueze, E.C.; Ulhôa, M.; Moulatlet, E.; Codarin, M.A.; Moreno, C.R. Effects of a Carbohydrate-Enriched Night Meal on Sleepiness and Sleep Duration in Night Workers: A Double-Blind Intervention. Chronobiol. Int. 2014, 31, 453–460. [Google Scholar] [CrossRef]
- Saidi, O.; Rochette, E.; Doré, É.; Maso, F.; Raoux, J.; Andrieux, F.; Fantini, M.L.; Merlin, E.; Pereira, B.; Walrand, S. Randomized Double-Blind Controlled Trial on the Effect of Proteins with Different Tryptophan/Large Neutral Amino Acid Ratios on Sleep in Adolescents: The PROTMORPHEUS Study. Nutrients 2020, 12, 1885. [Google Scholar] [CrossRef]
- Boelsma, E.; Brink, E.J.; Stafleu, A.; Hendriks, H.F.J. Measures of Postprandial Wellness after Single Intake of Two Protein–Carbohydrate Meals. Appetite 2010, 54, 456–464. [Google Scholar] [CrossRef] [PubMed]
- Netzer, N.C.; Stoohs, R.A.; Netzer, C.M.; Clark, K.; Strohl, K.P. Using the Berlin Questionnaire to Identify Patients at Risk for the Sleep Apnea Syndrome. Ann. Intern. Med. 1999, 131, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Horne, J.A.; Östberg, O. A Self-Assessment Questionnaire to Determine Morningness-Eveningness in Human Circadian Rhythms. Int. J. Chronobiol. 1976, 4, 97–110. [Google Scholar] [PubMed]
- Buysse, D.J.; Reynolds, C.F.; Monk, T.H.; Berman, S.R.; Kupfer, D.J. The Pittsburgh Sleep Quality Index: A New Instrument for Psychiatric Practice and Research. Psychiatry Res. 1989, 28, 193–213. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.H.; Macfarlane, D.J.; Lam, T.; Stewart, S.M. Validity of the International Physical Activity Questionnaire Short Form (IPAQ-SF): A Systematic Review. Int. J. Behav. Nutr. Phys. Act. 2011, 8, 115. [Google Scholar] [CrossRef]
- Freedson, P.S.; Melanson, E.; Sirard, J. Calibration of the Computer Science and Applications, Inc. Accelerometer. Med. Sci. Sports Exerc. 1998, 30, 777–781. [Google Scholar] [CrossRef]
- Pullman, R.E.; Roepke, S.E.; Duffy, J.F. Laboratory Validation of an In-Home Method for Assessing Circadian Phase Using Dim Light Melatonin Onset (DLMO). Sleep Med. 2012, 13, 703–706. [Google Scholar] [CrossRef]
- Popovic, D.; Khoo, M.; Westbrook, P. Automatic Scoring of Sleep Stages and Cortical Arousals Using Two Electrodes on the Forehead: Validation in Healthy Adults. J. Sleep Res. 2014, 23, 211–221. [Google Scholar] [CrossRef]
- Levendowski, D.J.; Popovic, D.; Berka, C.; Westbrook, P.R. Retrospective Cross-Validation of Automated Sleep Staging Using Electroocular Recording in Patients with and without Sleep Disordered Breathing. Int. Arch. Med. 2012, 5, 21. [Google Scholar] [CrossRef]
- Westbrook, P.R.; Levendowski, D.J.; Zavora, T.; Davis, G.; Popovic, D.; Berka, C.; Mitrovic, M.; Veljkovic, B. System for the Assessment of Sleep Quality in Adults and Children. U.S. Patent 8,355,769, 15 January 2014. [Google Scholar]
- Berry, R.B.; Brooks, R.; Gamaldo, C.; Harding, S.M.; Lloyd, R.M.; Quan, S.F.; Troester, M.T.; Vaughn, B.V. AASM Scoring Manual Updates for 2017 (Version 2.4); American Academy of Sleep Medicine: Darien, IL, USA, 2017; ISBN 15509389. [Google Scholar]
- Monti, J.M. Serotonin Control of Sleep-Wake Behavior. Sleep Med. Rev. 2011, 15, 269–281. [Google Scholar] [CrossRef]
- Davidson, J.; Sjoerdsma, A.; Loomis, L.N.; Udenfriend, S. Studies with the Serotonin Precursor, 5-Hydroxytryptophan, in Experimental Animals and Man. J. Clin. Investig. 1957, 36, 1594–1599. [Google Scholar] [CrossRef] [PubMed]
- McMenamy, R.H.; Oncley, J.L. The Specific Binding of L-Tryptophan to Serum Albumin. J. Biol. Chem. 1958, 1436–1447. [Google Scholar] [CrossRef]
- Pardridge, W.M. Blood-Brain Barrier Carrier-Mediated Transport and Brain Metabolism of Amino Acids. Neurochem. Res. 1998, 23, 635–644. [Google Scholar] [CrossRef]
- Halson, S.L. Sleep in Elite Athletes and Nutritional Interventions to Enhance Sleep. Sports Med. 2014, 44, 13–23. [Google Scholar] [CrossRef]
- Fukagawa, N.K.; Minaker, K.L.; Rowe, J.W.; Young, V.R. Plasma Tryptophan and Total Neutral Amino Acid Levels in Men: Influence of Hyperinsulinemia and Age. Metabolism 1987, 36, 683–686. [Google Scholar] [CrossRef]
- Voderholzer, U.; Hornyak, M.; Thiel, B.; Huwig-Poppe, C.; Kiemen, A.; König, A.; Backhaus, J.; Riemann, D.; Berger, M.; Hohagen, F. Impact of Experimentally Induced Serotonin Deficiency by Tryptophan Depletion on Sleep EEC in Healthy Subjects. Neuropsychopharmacology 1998, 18, 112–124. [Google Scholar] [CrossRef]
- Bhatti, T.; Gillin, J.C.; Seifritz, E.; Moore, P.; Clark, C.; Golshan, S.; Stahl, S.; Rapaport, M.; Kelsoe, J. Effects of a Tryptophan-Free Amino Acid Drink Challenge on Normal Human Sleep Electroencephalogram and Mood. Biol. Psychiatry 1998, 43, 52–59. [Google Scholar] [CrossRef]
- Arnulf, I.; Quintin, P.; Alvarez, J.-C.; Vigil, L.; Touitou, Y.; Lèbre, A.-S.; Bellenger, A.; Varoquaux, O.; Derenne, J.-P.; Allilaire, J.-F. Mid-Morning Tryptophan Depletion Delays REM Sleep Onset in Healthy Subjects. Neuropsychopharmacology 2002, 27, 843–851. [Google Scholar] [CrossRef] [PubMed]
- Bell, C.J.; Hood, S.D.; Nutt, D.J. Acute Tryptophan Depletion. Part II: Clinical Effects and Implications. Aust. N. Z. J. Psychiatry 2005, 39, 565–574. [Google Scholar] [CrossRef]
- Wurtman, R.J.; Wurtman, J.J.; Regan, M.M.; McDermott, J.M.; Tsay, R.H.; Breu, J.J. Effects of Normal Meals Rich in Carbohydrates or Proteins on Plasma Tryptophan and Tyrosine Ratios. Am. J. Clin. Nutr. 2003, 77, 128–132. [Google Scholar] [CrossRef]
- Lacey, J.H.; Stanley, P.; Hartmann, M.; Koval, J.; Crisp, A.H. The Immediate Effect of Intravenous Specific Nutrients on EEG Sleep. Electroencephalogr. Clin. Neurophysiol. 1978, 44, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Karklin, A.; Driver, H.S.; Buffenstein, R. Restricted Energy Intake Affects Nocturnal Body Temperature and Sleep Patterns. Am. J. Clin. Nutr. 1994, 59, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T. Roles of Orexin/Hypocretin in Regulation of Sleep/Wakefulness and Energy Homeostasis. Sleep Med. Rev. 2005, 9, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Burdakov, D.; González, J.A. Physiological Functions of Glucose-inhibited Neurones. Acta Physiol. 2009, 195, 71–78. [Google Scholar] [CrossRef]
- Saito, Y.C.; Tsujino, N.; Abe, M.; Yamazaki, M.; Sakimura, K.; Sakurai, T. Serotonergic Input to Orexin Neurons Plays a Role in Maintaining Wakefulness and REM Sleep Architecture. Front. Neurosci. 2018, 12, 892. [Google Scholar] [CrossRef]
- Chieffi, S.; Carotenuto, M.; Monda, V.; Valenzano, A.; Villano, I.; Precenzano, F.; Tafuri, D.; Salerno, M.; Filippi, N.; Nuccio, F. Orexin System: The Key for a Healthy Life. Front. Physiol. 2017, 8, 357. [Google Scholar] [CrossRef]
- Latifi, B.; Adamantidis, A.; Bassetti, C.; Schmidt, M.H. Sleep-Wake Cycling and Energy Conservation: Role of Hypocretin and the Lateral Hypothalamus in Dynamic State-Dependent Resource Optimization. Front. Neurol. 2018, 9, 790. [Google Scholar] [CrossRef]
- Aston-Jones, G.; Smith, R.J.; Moorman, D.E.; Richardson, K.A. Role of Lateral Hypothalamic Orexin Neurons in Reward Processing and Addiction. Neuropharmacology 2009, 56, 112–121. [Google Scholar] [CrossRef]
- Choi, D.L.; Davis, J.F.; Fitzgerald, M.E.; Benoit, S.C. The Role of Orexin-A in Food Motivation, Reward-Based Feeding Behavior and Food-Induced Neuronal Activation in Rats. Neuroscience 2010, 167, 11–20. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Mean (SD) | 95% CI | |
|---|---|---|
| Age (year) | 22.80 (1.90) | (22–23) |
| Height (cm) | 181 (7.36) | (178–184) |
| Weight (kg) | 73.30 (7.30) | (70.21–76.38) |
| BMI (kg·m−2) | 22.30 (1.54) | (21.68–22.98) |
| FM (%) | 11.70 (3.64) | (10.21–13.28) |
| FFM (kg) | 64.50 (5.17) | (62.34–66.71) |
| Circadian typology (MEQscore) | 49.10 (10.30) | (44.79–53.46) |
| Sleep quality (PSQIscore) | 4.79 (1.89) | (3.99–5.59) |
| Sleepiness (ESSscore) | 5.96 (3.08) | (4.65–7.26) |
| Sleep stabilization week sleep onset (time) | 22:39 (00:13) | (1353–1364) |
| Sleep stabilization week total time in bed (h:min) | 8:04 (00:15) | (476–490) |
| RMR (kcal·day−1) | 1813 (135) | (1756–1870) |
| HPLC (n = 24) | LPHC (n = 24) | HPLC vs. LPHC | ||||||
|---|---|---|---|---|---|---|---|---|
| PRE Mean (SD) | POST Mean (SD) | p-Value (ES) of Differences | PRE Mean (SD) | POST Mean (SD) | p-Value (ES) of Differences | Delta HPLC-LPHC (SD) | p-Value (ES) of Differences | |
| TST (min) | 423 (24.00) | 419 (25.30) | 0.126 (0.11) | 426 (24.60) | 426 (24.40) | 0.987 (0.02) | −4.50 (14.84) | 0.290 (0.17) |
| REM (min) | 108 (21.60) | 102 (27.50) | 0.119 (0.10) | 109 (22.10) | 112 (17.00) | 0.145 (0.10) | −9.10 (23.82) | 0.039 (0.32) |
| REM (%) | 25.60 (4.94) | 24.50 (6.44) | 0.381 (0.08) | 25.70 (5.43) | 26.40 (4.16) | 0.186 (0.10) | −3.00 (7.77) | 0.053 (0.37) |
| N1 (min) | 30.3 (5.33) | 29.90 (10.20) | 0.617 (0.11) | 29.90 (5.05) | 22.30 (5.32) | <0.001 (0.59) | 7.30 (13.34) | 0.023 (0.41) |
| N1 (%) | 7.17 (1.31) | 7.16 (2.45) | 0.689 (0.08) | 7.05 (1.28) | 5.25 (1.24) | <0.001 (0.59) | 1.80 (3.10) | 0.013 (0.42) |
| N2 (min) | 197 (21.10) | 200 (26.20) | 0.617 (0.04) | 198 (23.20) | 197 (31.20) | 0.668 (0.06) | 4.40 (31.92) | 0.345 (0.09) |
| N2 (%) | 46.70 (6.07) | 47.90 (6.52) | 0.265 (0.11) | 46.70 (5.85) | 46.40 (7.43) | 0.565 (0.06) | 1.50 (8.21) | 0.259 (0.17) |
| N3 (min) | 88 (37.80) | 86.40 (45.60) | 0.700 (0.12) | 88.70 (40.40) | 94.20 (40.10) | 0.359 (0.09) | −7.10 (55.33) | 0.317 (0.16) |
| N3 (%) | 20.60 (8.27) | 20.40 (10.20) | 0.684 (0.12) | 20.60 (8.88) | 21.90 (9.03) | 0.304 (0.12) | −1.50 (12.60) | 0.391 (0.16) |
| SE (%) | 88.70 (3.58) | 87.80 (3.47) | 0.057 (0.13) | 89 (3.23) | 89.50 (3.37) | 0.188 (0.09) | −1.20 (2.46) | 0.038 (0.32) |
| SOL (min) | 15.50 (8.57) | 15.20 (6.45) | 0.667 (0.001) | 15.80 (7.78) | 11.40 (4.49) | <0.001 (0.36) | 4.30 (6.10) | 0.004 (0.31) |
| WASO (min) | 38.50 (13.80) | 42.40 (12.40) | 0.055 (0.21) | 36.40 (12.00) | 38.50 (13.80) | 0.128 (0.07) | 1.70 (10.87) | 0.361 (0.11) |
| HPLC (n = 24) | LPHC (n = 24) | HPLC vs. LPHC | ||
|---|---|---|---|---|
| Mean (SD) | Mean (SD) | Delta HPLC-LPHC (SD) | p-Value (ES) of Differences | |
| Total EI (kcal) | 2878.15 (218.65) | 2814.38 (185.48) | 63.77 (104.87) | 0.027 (0.16) |
| Total CHO (g) | 337.43 (22.10) | 328.50 (18.01) | 8.93 (14.15) | 0.007 (0.19) |
| Total Fat (g) | 114.25 (10.34) | 112.17 (14.11) | 2.08 (8.63) | 0.290 (0.09) |
| Total Protein (g) | 125.05 (28.51) | 122.71 (29.30) | 2.34 (16.01) | 0.875 (0.06) |
| Breakfast EI (kcal) | 606.29 (97.15) | 617.13 (121.36) | −10.83 (75.39) | 0.681 (0.03) |
| Breakfast CHO (g) | 86.30 (14.12) | 79.19 (15.39) | 7.12 (15.73) | 0.023 (0.27) |
| Breakfast Fat (g) | 17.48 (5.12) | 21.98 (7.12) | −4.49 (7.36) | 0.016 (0.31) |
| Breakfast protein (g) | 25.93 (7.61) | 25.65 (8.04) | 0.28 (6.82) | 0.700 (0.05) |
| Lunch EI (kcal) | 1002.35 (144.21) | 1004.34 (155.65) | −1.99 (236.57) | 0.989 (0.02) |
| Lunch CHO (g) | 116.32 (16.29) | 120.92 (21.93) | −4.60 (29.32) | 0.242 (0.15) |
| Lunch Fat (g) | 39.55 (7.09) | 36.83 (10.22) | 2.72 (13.67) | 0.169 (0.15) |
| Lunch protein (g) | 45.28 (11.51) | 47.30 (12.39) | −2.02 (9.95) | 0.219 (0.10) |
| Dinner EI (kcal) | 1001.04 (116.30) | 959.73 (71.56) | 41.32 (146.50) | 0.432 (0.14) |
| Dinner CHO (g) | 99.05 (13.77) | 98.94 (12.42) | 0.11 (21.18) | 0.988 (0.06) |
| Dinner Fat (g) | 48.37 (7.14) | 45.00 (7.52) | 3.37 (10.79) | 0.170 (0.28) |
| Dinner protein (g) | 42.38 (10.09) | 39.76 (9.13) | 2.63 (7.77) | 0.204 (0.15) |
| Snacking EI (kcal) | 268.47 (59.69) | 233.19 (80.74) | 35.28 (94.34) | 0.002 (0.38) |
| Snacking CHO (g) | 35.75 (11.88) | 29.45 (10.66) | 6.30 (13.52) | 0.027 (0.24) |
| Snacking Fat (g) | 8.85 (5.16) | 8.38 (4.25) | 0.47 (6.22) | 0.944 (0.003) |
| Snacking protein (g) | 11.46 (8.02) | 10.00 (7.09) | 1.46 (10.38) | 0.406 (0.11) |
| HPLC (n = 24) | LPHC (n = 24) | HPLC vs. LPHC | ||
|---|---|---|---|---|
| Mean (SD) | Mean (SD) | Delta HPLC-LPHC (SD) | p-Value (ES) of Differences | |
| Time spent on SB (min) | 485 (30) | 482 (24) | 3 (20) | 0.533 (0.06) |
| Time spent on LPA (min) | 436 (41) | 437 (32) | −1 (20) | 0.553 (0.05) |
| Time spent on MPA (min) | 35 (16) | 40 (17) | −5 (15) | 0.117 (0.13) |
| Time spent on VPA (min) | 7 (6) | 8 (5) | −1 (5) | 0.637 (0.10) |
| Time spent on MVPA (min) | 42 (19) | 48 (20) | −6 (15) | 0.074 (0.16) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saidi, O.; Rochette, E.; Del Sordo, G.; Peyrel, P.; Salles, J.; Doré, E.; Merlin, E.; Walrand, S.; Duché, P. Isocaloric Diets with Different Protein-Carbohydrate Ratios: The Effect on Sleep, Melatonin Secretion and Subsequent Nutritional Response in Healthy Young Men. Nutrients 2022, 14, 5299. https://doi.org/10.3390/nu14245299
Saidi O, Rochette E, Del Sordo G, Peyrel P, Salles J, Doré E, Merlin E, Walrand S, Duché P. Isocaloric Diets with Different Protein-Carbohydrate Ratios: The Effect on Sleep, Melatonin Secretion and Subsequent Nutritional Response in Healthy Young Men. Nutrients. 2022; 14(24):5299. https://doi.org/10.3390/nu14245299
Chicago/Turabian StyleSaidi, Oussama, Emmanuelle Rochette, Giovanna Del Sordo, Paul Peyrel, Jérôme Salles, Eric Doré, Etienne Merlin, Stéphane Walrand, and Pascale Duché. 2022. "Isocaloric Diets with Different Protein-Carbohydrate Ratios: The Effect on Sleep, Melatonin Secretion and Subsequent Nutritional Response in Healthy Young Men" Nutrients 14, no. 24: 5299. https://doi.org/10.3390/nu14245299
APA StyleSaidi, O., Rochette, E., Del Sordo, G., Peyrel, P., Salles, J., Doré, E., Merlin, E., Walrand, S., & Duché, P. (2022). Isocaloric Diets with Different Protein-Carbohydrate Ratios: The Effect on Sleep, Melatonin Secretion and Subsequent Nutritional Response in Healthy Young Men. Nutrients, 14(24), 5299. https://doi.org/10.3390/nu14245299