Flavonoids, Potential Bioactive Compounds, and Non-Shivering Thermogenesis


Abstract
1. Introduction
2. Effects of Flavonoids on Non-Shivering Thermogenesis
2.1. Flavones
2.1.1. Primary Food Sources and Daily Intake
2.1.2. Effects of Flavones on Non-Shivering Thermogenesis
2.2. Flavonols
2.2.1. Major Food Sources and Daily Intake
2.2.2. Effect of Flavonols on Non-Shivering Thermogenesis
2.3. Flavanones
2.3.1. Major Food Sources and Daily Intake
2.3.2. Effect of Flavanones on Non-Shivering Thermogenesis
2.4. Flavanols
2.4.1. Major Food Sources and Daily Intake
2.4.2. Effects of Flavanols on Non-Shivering Thermogenesis
2.5. Anthocyanins
2.5.1. Major Food Sources and Daily Intake
2.5.2. Effect of Anthocyanins on Non-Shivering Thermogenesis
2.6. Isoflavones
2.6.1. Major Food Sources and Daily Intake
2.6.2. Effect of Isoflavones on Non-Shivering Thermogenesis
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| ACC | acetyl-coenzyme A carboxylase |
| ACO | acyl-coenzyme A oxidase |
| AMPK | AMP-activated protein kinase |
| AP2 | adipocyte protein 2 |
| ATP5α | ATP synthase F1 subunit alpha |
| BAT | brown adipose tissue |
| β-AR | beta-adrenergic receptor |
| BMP7 | bone morphogenetic protein 7 |
| C/EBPβ | CCAAT/enhancer-binding protein beta |
| CD137/ TNFRSF9 | tumor necrosis factor receptor superfamily member 9 |
| CIDEA | cell death-inducing DNA fragmentation factor alpha (DFFA)-like effector a |
| CITED | cbp/p300-interacting transactivator |
| COMT | catechol-O-methyltransferase |
| CPT1α | carnitine palmitoyltransferase 1 alpha |
| DHEA | dehydroepiandrosterone |
| DIO2 | deiodinase 2 |
| EG | epicatechin gallate |
| EGC | epigallocatechin |
| EGCG | epigallocatechin gallate |
| ELOVL3 | elongation of very-long chain fatty acids-like 3 |
| EWAT | epididymal white adipose tissue |
| FAS | fatty acid synthase |
| FGF 21 | fibroblast growth factor 21 |
| HFD | high-fat diet |
| HSL | hormone sensitive lipase |
| IWAT | inguinal white adipose tissue |
| LDL | low density lipoprotein |
| MAPK | mitogen-activated protein kinase |
| MCAFA | medium-chain acyl-coenzyme A |
| ME | mulberry extract |
| Myf5 | myogenic factor 5 |
| NHANES | National Health and Nutrition Examination Survey |
| NRF1 | nuclear respiratory factor 1 |
| NDUFB8 | 246 NADH Dehydrogenase (Ubiquinone) 1 Beta Subcomplex, 8 |
| OLE | olive leaf extract |
| OPE | onion peel extract |
| PGC1α | peroxisome proliferator-activated receptor gamma coactivator 1 alpha |
| PLIN | perilipin |
| PKA | protein kinase A |
| PKC | protein kinase C |
| PPARα | peroxisome proliferator-activated receptor alpha |
| PPARβ | peroxisome proliferator-activated receptor beta |
| PPARγ | peroxisome proliferator-activated receptor gamma |
| PRDM16 | positive regulatory domain containing 16 |
| PCOS | polycystic ovary syndrome |
| RWAT | retroperitoneal white adipose tissue |
| SWAT | subcutaneous white adipose tissue |
| SIRT1 | silent mating type information regulation 2 homolog 1 |
| SDHB | succinate dehydrogenase complex iron sulfur subunit B |
| TAG | triacylglycerol |
| TBX1 | T-box transcription factor 1 |
| TFAM | mitochondrial transcription factor A |
| TMEM 26 | transmembrane protein 26 |
| UCP1 | uncoupling protein 1 |
| UCP2 | uncoupling protein 2 |
| UCP3 | uncoupling protein 3 |
| UQCRC2 | ubiquinol-cytochrome c reductase core protein 2 |
| VLDL | very-low density lipoprotein |
| WAT | white adipose tissue |
| WNT | wingless type |
| WNT10b | wingless type MMTV integration site family, member 10b |
References
- Grundy, S.M. Metabolic complications of obesity. Endocrine 2000, 13, 155–165. [Google Scholar] [CrossRef]
- Fruhbeck, G.; Becerril, S.; Sainz, N.; Garrastachu, P.; Garcia-Velloso, M.J. Bat: A new target for human obesity? Trends Pharmacol. Sci. 2009, 30, 387–396. [Google Scholar] [CrossRef] [PubMed]
- Kajimura, S.; Saito, M. A new era in brown adipose tissue biology: Molecular control of brown fat development and energy homeostasis. Annu. Rev. Physiol. 2014, 76, 225–249. [Google Scholar] [CrossRef] [PubMed]
- Cypess, A.M.; Lehman, S.; Williams, G.; Tal, I.; Rodman, D.; Goldfine, A.B.; Kuo, F.C.; Palmer, E.L.; Tseng, Y.H.; Doria, A.; et al. Identification and importance of brown adipose tissue in adult humans. N. Engl. J. Med. 2009, 360, 1509–1517. [Google Scholar] [CrossRef] [PubMed]
- Van Marken Lichtenbelt, W.D.; Vanhommerig, J.W.; Smulders, N.M.; Drossaerts, J.M.; Kemerink, G.J.; Bouvy, N.D.; Schrauwen, P.; Teule, G.J. Cold-activated brown adipose tissue in healthy men. N. Engl. J. Med. 2009, 360, 1500–1508. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, K.A.; Lidell, M.E.; Orava, J.; Heglind, M.; Westergren, R.; Niemi, T.; Taittonen, M.; Laine, J.; Savisto, N.J.; Enerbäck, S.; et al. Functional brown adipose tissue in healthy adults. N. Engl. J. Med. 2009, 360, 1518–1525. [Google Scholar] [CrossRef] [PubMed]
- Bartelt, A.; Bruns, O.T.; Reimer, R.; Hohenberg, H.; Ittrich, H.; Peldschus, K.; Kaul, M.G.; Tromsdorf, U.I.; Weller, H.; Waurisch, C.; et al. Brown adipose tissue activity controls triglyceride clearance. Nat. Med. 2011, 17, 200–205. [Google Scholar] [CrossRef] [PubMed]
- Timmons, J.A.; Wennmalm, K.; Larsson, O.; Walden, T.B.; Lassmann, T.; Petrovic, N.; Hamilton, D.L.; Gimeno, R.E.; Wahlestedt, C.; Baar, K.; et al. Myogenic gene expression signature establishes that brown and white adipocytes originate from distinct cell lineages. Proc. Natl. Acad. Sci. USA 2007, 104, 4401–4406. [Google Scholar] [CrossRef] [PubMed]
- Seale, P.; Bjork, B.; Yang, W.; Kajimura, S.; Chin, S.; Kuang, S.; Scime, A.; Devarakonda, S.; Conroe, H.M.; Erdjument-Bromage, H.; et al. Prdm16 controls a brown fat/skeletal muscle switch. Nature 2008, 454, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Brand, M.D.; Esteves, T.C. Physiological functions of the mitochondrial uncoupling proteins UCP2 and UCP3. Cell Metab. 2005, 2, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Seale, P. Transcriptional regulatory circuits controlling brown fat development and activation. Diabetes 2015, 64, 2369–2375. [Google Scholar] [CrossRef] [PubMed]
- Hondares, E.; Rosell, M.; Diaz-Delfin, J.; Olmos, Y.; Monsalve, M.; Iglesias, R.; Villarroya, F.; Giralt, M. PPARα induces pgc-1α gene expression and contributes to the thermogenic activation of brown fat; involvement of prdm16. J. Biol. Chem. 2011, 286, 43112–43122. [Google Scholar] [CrossRef] [PubMed]
- Hanssen, M.J.; Broeders, E.; Samms, R.J.; Vosselman, M.J.; Van Der Lans, A.A.; Cheng, C.C.; Adams, A.C.; van Marken Lichtenbelt, W.D.; Schrauwen, P. Serum fgf21 levels are associated with brown adipose tissue activity in humans. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Zhou, L.; Chen, C.; Gong, J.; Xu, L.; Ye, J.; Li, P. Cidea controls lipid droplet fusion and lipid storage in brown and white adipose tissue. Sci. China Life Sci. 2014, 57, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Barneda, D.; Planas-Iglesias, J.; Gaspar, M.L.; Mohammadyani, D.; Prasannan, S.; Dormann, D.; Han, G.-S.; Jesch, S.A.; Carman, G.M.; Kagan, V.; et al. The brown adipocyte protein cidea promotes lipid droplet fusion via a phosphatidic acid-binding amphipathic helix. eLife 2015, 4, e07485. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Yon Toh, S.; Chen, Z.; Guo, K.; Peng Ng, C.; Ponniah, S.; Lin, S.-C.; Hong, W.; Li, P. Cidea-deficient mice have lean phenotype and are resistant to obesity. Nat. Genet. 2003, 35, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Cohen, P.; Spiegelman, B.M. Adaptive thermogenesis in adipocytes: Is beige the new brown? Genes Dev. 2013, 27, 234–250. [Google Scholar] [CrossRef] [PubMed]
- Harms, M.; Seale, P. Brown and beige fat: Development, function and therapeutic potential. Nat. Med. 2013, 19, 1252–1263. [Google Scholar] [CrossRef] [PubMed]
- Garcia, R.A.; Roemmich, J.N.; Claycombe, K.J. Evaluation of markers of beige adipocytes in white adipose tissue of the mouse. J. Nutr. Metab. 2016, 13, 24. [Google Scholar] [CrossRef] [PubMed]
- Cannon, B.; Nedergaard, J. Brown adipose tissue: Function and physiological significance. Physiol. Rev. 2004, 84, 277–359. [Google Scholar] [CrossRef] [PubMed]
- Weiner, J.; Kranz, M.; Klöting, N.; Kunath, A.; Steinhoff, K.; Rijntjes, E.; Köhrle, J.; Zeisig, V.; Hankir, M.; Gebhardt, C.; et al. Thyroid hormone status defines brown adipose tissue activity and browning of white adipose tissues in mice. Sci. Rep. 2016, 6, 38124. [Google Scholar] [CrossRef] [PubMed]
- Bianco, A.C.; McAninch, E.A. The role of thyroid hormone and brown adipose tissue in energy homoeostasis. Lancet Diabetes Endocrinol. 2013, 1, 250–258. [Google Scholar] [CrossRef]
- Cantó, C.; Auwerx, J. Pgc-1alpha, sirt1 and ampk, an energy sensing network that controls energy expenditure. Curr. Opin. Lipidol. 2009, 20, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Takahashi, N.; Goto, T.; Kawada, T. Dietary factors evoke thermogenesis in adipose tissues. Obes. Res. Clin. Pract. 2014, 8, e533–e539. [Google Scholar] [CrossRef] [PubMed]
- Castrejon-Tellez, V.; Rodriguez-Perez, J.M.; Perez-Torres, I.; Perez-Hernandez, N.; Cruz-Lagunas, A.; Guarner-Lans, V.; Vargas-Alarcon, G.; Rubio-Ruiz, M.E. The effect of resveratrol and quercetin treatment on ppar mediated uncoupling protein (ucp-) 1, 2, and 3 expression in visceral white adipose tissue from metabolic syndrome rats. Int. J. Mol. Sci. 2016, 17, 1069. [Google Scholar] [CrossRef] [PubMed]
- Lone, J.; Choi, J.H.; Kim, S.W.; Yun, J.W. Curcumin induces brown fat-like phenotype in 3t3-l1 and primary white adipocytes. J. Nutr. Biochem. 2016, 27, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Kawser Hossain, M.; Abdal Dayem, A.; Han, J.; Yin, Y.; Kim, K.; Kumar Saha, S.; Yang, G.M.; Yeon Choi, H.; Cho, S.G. Molecular mechanisms of the anti-obesity and anti-diabetic properties of flavonoids. Int. J. Mol. Sci. 2016, 17, 569. [Google Scholar] [CrossRef] [PubMed]
- Vernarelli, J.A.; Lambert, J.D. Flavonoid intake is inversely associated with obesity and c-reactive protein, a marker for inflammation, in us adults. Nutr. Diabetes 2017, 7, e276. [Google Scholar] [CrossRef] [PubMed]
- Woodman, O.L.; Meeker, W.F.; Boujaoude, M. Vasorelaxant and antioxidant activity of flavonols and flavones: Structure-activity relationships. J. Cardiovasc. Pharmacol. 2005, 46, 302–309. [Google Scholar] [CrossRef] [PubMed]
- Miean, K.H.; Mohamed, S. Flavonoid (myricetin, quercetin, kaempferol, luteolin, and apigenin) content of edible tropical plants. J. Agric. Food Chem. 2001, 49, 3106–3112. [Google Scholar] [CrossRef] [PubMed]
- Chun, O.K.; Chung, S.J.; Song, W.O. Estimated dietary flavonoid intake and major food sources of U.S. Adults. J. Nutr. 2007, 137, 1244–1252. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Lazaro, M. Distribution and biological activities of the flavonoid luteolin. Mini Rev. Med. Chem. 2009, 9, 31–59. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, Q.X.; Wang, X.; Zhang, L.; Qu, W.; Bao, B.; Liu, C.A.; Liu, J. Dietary luteolin activates browning and thermogenesis in mice through an AMPK/PGC1 α pathway-mediated mechanism. Int. J. Obes. 2016, 40, 1841–1849. [Google Scholar] [CrossRef] [PubMed]
- Mulligan, J.D.; Gonzalez, A.A.; Stewart, A.M.; Carey, H.V.; Saupe, K.W. Upregulation of AMPK during cold exposure occurs via distinct mechanisms in brown and white adipose tissue of the mouse. J. Physiol. 2007, 580, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Van Dam, A.D.; Kooijman, S.; Schilperoort, M.; Rensen, P.C.; Boon, M.R. Regulation of brown fat by amp-activated protein kinase. Trends Mol. Med. 2015, 21, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Premalatha, M.; Parameswari, C. Renoprotective effect of chrysin (5, 7 dihydroxy flavone) in streptozotocin induced diabetic nephropathy in rats. Int. J. Pharm. Pharm. Sci. 2012, 4, 241–247. [Google Scholar]
- Hougee, S.; Sanders, A.; Faber, J.; Graus, Y.M.; van den Berg, W.B.; Garssen, J.; Smit, H.F.; Hoijer, M.A. Decreased pro-inflammatory cytokine production by LPS-stimulated PBMC upon in vitro incubation with the flavonoids apigenin, luteolin or chrysin, due to selective elimination of monocytes/macrophages. Biochem. Pharmacol. 2005, 69, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Khoo, B.Y.; Chua, S.L.; Balaram, P. Apoptotic effects of chrysin in human cancer cell lines. Int J. Mol. Sci. 2010, 11, 2188–2199. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.H.; Yun, J.W. Chrysin induces brown fat-like phenotype and enhances lipid metabolism in 3t3-l1 adipocytes. Nutrition 2016, 32, 1002–1010. [Google Scholar] [CrossRef] [PubMed]
- Asano, H.; Kanamori, Y.; Higurashi, S.; Nara, T.; Kato, K.; Matsui, T.; Funaba, M. Induction of beige-like adipocytes in 3T3-L1 cells. J. Vet. Med. Sci. 2014, 76, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Karamanlidis, G.; Karamitri, A.; Docherty, K.; Hazlerigg, D.G.; Lomax, M.A. C/EBPbeta reprograms white 3T3-L1 preadipocytes to a brown adipocyte pattern of gene expression. J. Biol. Chem. 2007, 282, 24660–24669. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.P.; Ferreira, I.C.; Marcelino, F.; Valentão, P.; Andrade, P.B.; Seabra, R.; Estevinho, L.; Bento, A.; Pereira, J.A. Phenolic compounds and antimicrobial activity of olive (Olea europaea L. Cv. Cobrançosa) leaves. Molecules 2007, 12, 1153–1162. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Song, S.J.; Keum, N.; Park, T. Olive leaf extract attenuates obesity in high-fat diet-fed mice by modulating the expression of molecules involved in adipogenesis and thermogenesis. Evid. Based Complement. Alternat. Med. 2014, 2014, 971890. [Google Scholar] [CrossRef] [PubMed]
- Lo, K.A.; Ng, P.Y.; Kabiri, Z.; Virshup, D.; Sun, L. Wnt inhibition enhances browning of mouse primary white adipocytes. Adipocyte 2016, 5, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-H.; Jung, E.-A.; Sohng, I.-S.; Han, J.-A.; Kim, T.-H.; Han, M.J. Intestinal bacterial metabolism of flavonoids and its relation to some biological activities. Arch. Pharm. Res. 1998, 21, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Itav, S.; Rothschild, D.; Meijer, M.; Levy, M.; Moresi, C.; Dohnalova, L.; Braverman, S.; Rozin, S.; Malitsky, S.; et al. Persistent microbiome alterations modulate the rate of post-dieting weight regain. Nature 2016, 540, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Suarez-Zamorano, N.; Fabbiano, S.; Chevalier, C.; Stojanovic, O.; Colin, D.J.; Stevanovic, A.; Veyrat-Durebex, C.; Tarallo, V.; Rigo, D.; Germain, S.; et al. Microbiota depletion promotes browning of white adipose tissue and reduces obesity. Nat. Med. 2015, 21, 1497–1501. [Google Scholar] [CrossRef] [PubMed]
- Braune, A.; Blaut, M. Bacterial species involved in the conversion of dietary flavonoids in the human gut. Gut Microbes 2016, 7, 216–234. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Takaishi, Y.; Tanaka, N.; Tsuchiya, K.; Shibata, H.; Higuti, T. Chemical constituents from the peels of citrus sudachi. J. Nat. Prod. 2006, 69, 1177–1179. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumi, R.; Yoshida, T.; Nii, Y.; Okahisa, N.; Iwata, S.; Tsukayama, M.; Hashimoto, R.; Taniguchi, Y.; Sakaue, H.; Hosaka, T.; et al. Sudachitin, a polymethoxylated flavone, improves glucose and lipid metabolism by increasing mitochondrial biogenesis in skeletal muscle. Nutr. Metab. 2014, 11, 32. [Google Scholar] [CrossRef] [PubMed]
- Bhagwat, S.; Haytowitz, D.B.; Wasswa-Kintu, S.I.; Holden, J.M. USDA develops a database for flavonoids to assess dietary intakes. Proc. Food Sci. 2013, 2, 81–86. [Google Scholar] [CrossRef]
- Moon, J.; Do, H.J.; Kim, O.Y.; Shin, M.J. Antiobesity effects of quercetin-rich onion peel extract on the differentiation of 3T3-L1 preadipocytes and the adipogenesis in high fat-fed rats. Food Chem. Toxicol. 2013, 58, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Zhang, X.; Zhang, L.; Bian, H.X.; Xu, N.; Bao, B.; Liu, J. Quercetin reduces obesity-associated atm infiltration and inflammation in mice: A mechanism including ampkalpha1/sirt1. J. Lipid Res. 2014, 55, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.G.; Parks, J.S.; Kang, H.W. Quercetin, a functional compound of onion peel, remodels white adipocytes to brown-like adipocytes. J. Nutr. Biochem. 2017, 42, 62–71. [Google Scholar] [CrossRef] [PubMed]
- Stewart, L.K.; Soileau, J.L.; Ribnicky, D.; Wang, Z.Q.; Raskin, I.; Poulev, A.; Majewski, M.; Cefalu, W.T.; Gettys, T.W. Quercetin transiently increases energy expenditure but persistently decreases circulating markers of inflammation in C57BL/6J mice fed a high-fat diet. Metabolism 2008, 57, S39–S46. [Google Scholar] [CrossRef] [PubMed]
- Henagan, T.; Cefalu, W.; Ribnicky, D.; Noland, R.; Dunville, K.; Campbell, W.; Stewart, L.; Forney, L.; Gettys, T.; Chang, J. In vivo effects of dietary quercetin and quercetin-rich red onion extract on skeletal muscle mitochondria, metabolism, and insulin sensitivity. Genes Nutr. 2015, 10, 2. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Wei, G.; You, Y.; Huang, Y.; Lee, H.J.; Dong, M.; Lin, J.; Hu, T.; Zhang, H.; Zhang, C.; et al. Rutin ameliorates obesity through brown fat activation. FASEB J. 2017, 31, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Ward, W.F. PGC-1alpha: A key regulator of energy metabolism. Adv. Physiol. Educ. 2006, 30, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.I.; Chen, Y.C.; Chen, L.H.; Juan, C.C.; Ku, H.H.; Wang, S.T.; Chiou, S.H.; Chiou, G.Y.; Chi, C.W.; Hsu, C.C.; et al. PGC-1alpha mediates differentiation of mesenchymal stem cells to brown adipose cells. J. Atheroscler. Thromb. 2011, 18, 966–980. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Yuan, X.; Ye, R.; Zhou, H.; Lin, J.; Zhang, C.; Zhang, H.; Wei, G.; Dong, M.; Huang, Y.; et al. Brown adipose tissue activation by rutin ameliorates polycystic ovary syndrome in rat. J. Nutr. Biochem. 2017, 47, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.; Yuan, X.; Wei, G.; Luo, H.; Lee, H.J.; Jin, W. Myricetin-induced brown adipose tissue activation prevents obesity and insulin resistance in db/db mice. Eur J. Nutr. 2017, 57, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Da-Silva, W.S.; Harney, J.W.; Kim, B.W.; Li, J.; Bianco, S.D.; Crescenzi, A.; Christoffolete, M.A.; Huang, S.A.; Bianco, A.C. The small polyphenolic molecule kaempferol increases cellular energy expenditure and thyroid hormone activation. Diabetes 2007, 56, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Vatkar, B.; Pratapwar, A.; Tapas, A.; Butle, S.; Tiwari, B. Synthesis and antimicrobial activity of some flavanone derivatives. Int. J. ChemTech Res. 2010, 2, 504–508. [Google Scholar]
- Klimczak, I.; Małecka, M.; Szlachta, M.; Gliszczyńska-Świgło, A. Effect of storage on the content of polyphenols, vitamin c and the antioxidant activity of orange juices. J. Food Compos. Anal. 2007, 20, 313–322. [Google Scholar] [CrossRef]
- Brett, G.M.; Hollands, W.; Needs, P.W.; Teucher, B.; Dainty, J.R.; Davis, B.D.; Brodbelt, J.S.; Kroon, P.A. Absorption, metabolism and excretion of flavanones from single portions of orange fruit and juice and effects of anthropometric variables and contraceptive pill use on flavanone excretion. Br. J. Nutr. 2008, 101, 664–675. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.-J.; Lee, O.-H.; Choi, H.-S.; Lee, B.-Y. Extract from edible red seaweed (gelidium amansi) inhibits lipid accumulation and GOS production during differentiation in 3T3-L1 cells. Prev. Nutr. Food Sci. 2012, 17, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.-H.; Yao, H.-T.; Chiang, M.-T. Red algae (gelidium amansii) reduces adiposity via activation of lipolysis in rats with diabetes induced by streptozotocin-nicotinamide. J. Food Drug Anal. 2015, 23, 758–765. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Kim, K.J.; Koh, E.J.; Lee, B.Y. Gelidium elegans regulates the ampk-prdm16-ucp-1 pathway and has a synergistic effect with orlistat on obesity-associated features in mice fed a high-fat diet. Nutrients 2017, 9, 342. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Kim, K.-J.; Koh, E.-J.; Lee, B.-Y. Altered gelidium elegans extract-stimulated beige-like phenotype attenuates adipogenesis in 3T3-L1 cells. J. Food Nutr. Res. 2016, 4, 448–453. [Google Scholar]
- Shen, J.; Nakamura, H.; Fujisaki, Y.; Tanida, M.; Horii, Y.; Fuyuki, R.; Takumi, H.; Shiraishi, K.; Kometani, T.; Nagai, K. Effect of 4G-alpha-glucopyranosyl hesperidin on brown fat adipose tissue- and cutaneous-sympathetic nerve activity and peripheral body temperature. Neurosci. Lett. 2009, 461, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Chetia, D.; Rudrapal, M. Structurally Diverse Bioflavonoids as Potential Source of Antimalarial Lead Molecules, 2016. Available online: http://www.old.iitbhu.ac.in/phe/pharmsociety/issue_2015/7.Chetia_and_Rudrapal.pdf (accessed on 30 May 2018).
- Song, W.O.; Chun, O.K. Tea is the major source of flavan-3-ol and flavonol in the us diet. J. Nutr. 2008, 138, 1543S–1547S. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Williamson, G. Dietary intake and bioavailability of polyphenols. J. Nutr. 2000, 130, 2073S–2085S. [Google Scholar] [CrossRef] [PubMed]
- Tsao, R. Chemistry and biochemistry of dietary polyphenols. Nutrients 2010, 2, 1231–1246. [Google Scholar] [CrossRef] [PubMed]
- Dulloo, A.G.; Seydoux, J.; Girardier, L.; Chantre, P.; Vandermander, J. Green tea and thermogenesis: Interactions between catechin-polyphenols, caffeine and sympathetic activity. Int J. Obes. Relat. Metab. Disord. 2000, 24, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Choo, J.J. Green tea reduces body fat accretion caused by high-fat diet in rats through beta-adrenoceptor activation of thermogenesis in brown adipose tissue. J. Nutr. Biochem. 2003, 14, 671–676. [Google Scholar] [CrossRef] [PubMed]
- Nomura, S.; Ichinose, T.; Jinde, M.; Kawashima, Y.; Tachiyashiki, K.; Imaizumi, K. Tea catechins enhance the mrna expression of uncoupling protein 1 in rat brown adipose tissue. J. Nutr. Biochem. 2008, 19, 840–847. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Zhao, Y.; Zhao, B. Green tea catechins prevent obesity through modulation of peroxisome proliferator-activated receptors. Sci. China Life Sci. 2013, 56, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Collins, S.; Surwit, R.S. The beta-adrenergic receptors and the control of adipose tissue metabolism and thermogenesis. Recent Prog. Horm. Res. 2001, 56, 309–328. [Google Scholar] [CrossRef] [PubMed]
- Gogos, J.A.; Morgan, M.; Luine, V.; Santha, M.; Ogawa, S.; Pfaff, D.; Karayiorgou, M. Catechol-o-methyltransferase-deficient mice exhibit sexually dimorphic changes in catecholamine levels and behavior. Proc. Natl. Acad. Sci. USA 1998, 95, 9991–9996. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Meng, X.; Yang, C.S. Enzymology of methylation of tea catechins and inhibition of catechol-o-methyltransferase by (−)-epigallocatechin gallate. Drug Metab. Dispos. 2003, 31, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, Y.; Wang, L.; Wang, L.; Tanaka, Y.; Zhang, T.; Ashida, H. Oolong, black and pu-erh tea suppresses adiposity in mice via activation of AMP-activated protein kinase. Food Funct. 2014, 5, 2420–2429. [Google Scholar] [CrossRef] [PubMed]
- Dulloo, A.G.; Duret, C.; Rohrer, D.; Girardier, L.; Mensi, N.; Fathi, M.; Chantre, P.; Vandermander, J. Efficacy of a green tea extract rich in catechin polyphenols and caffeine in increasing 24-h energy expenditure and fat oxidation in humans. Am. J. Clin. Nutr. 1999, 70, 1040–1045. [Google Scholar] [CrossRef] [PubMed]
- Nirengi, S.; Amagasa, S.; Homma, T.; Yoneshiro, T.; Matsumiya, S.; Kurosawa, Y.; Sakane, N.; Ebi, K.; Saito, M.; Hamaoka, T. Daily ingestion of catechin-rich beverage increases brown adipose tissue density and decreases extramyocellular lipids in healthy young women. Springerplus 2016, 5, 1363. [Google Scholar] [CrossRef] [PubMed]
- Gosselin, C.; Haman, F. Effects of green tea extracts on non-shivering thermogenesis during mild cold exposure in young men. Br. J. Nutr. 2013, 110, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Matsumura, Y.; Nakagawa, Y.; Mikome, K.; Yamamoto, H.; Osakabe, N. Enhancement of energy expenditure following a single oral dose of flavan-3-ols associated with an increase in catecholamine secretion. PLoS ONE 2014, 9, e112180. [Google Scholar] [CrossRef] [PubMed]
- Kamio, N.; Suzuki, T.; Watanabe, Y.; Suhara, Y.; Osakabe, N. A single oral dose of flavan-3-ols enhances energy expenditure by sympathetic nerve stimulation in mice. Free Radic. Biol. Med. 2016, 91, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Rabadan-Chávez, G.; Quevedo-Corona, L.; Garcia, A.M.; Reyes-Maldonado, E.; Jaramillo-Flores, M.E. Cocoa powder, cocoa extract and epicatechin attenuate hypercaloric diet-induced obesity through enhanced β-oxidation and energy expenditure in white adipose tissue. J. Funct. Food 2016, 20, 54–67. [Google Scholar] [CrossRef]
- Gutierrez-Salmean, G.; Ortiz-Vilchis, P.; Vacaseydel, C.M.; Garduno-Siciliano, L.; Chamorro-Cevallos, G.; Meaney, E.; Villafana, S.; Villarreal, F.; Ceballos, G.; Ramirez-Sanchez, I. Effects of (−)-epicatechin on a diet-induced rat model of cardiometabolic risk factors. Eur. J. Pharmacol. 2014, 728, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.W.; Gong, C.C.; Song, H.F.; Cui, Y.Y. Effects of anthocyanins on the prevention and treatment of cancer. Brit. J. Pharmacol. 2017, 174, 1226–1243. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Kou, X.; Fugal, K.; McLaughlin, J. Comparison of HPLC methods for determination of anthocyanins and anthocyanidins in bilberry extracts. J. Agric. Food Chem. 2004, 52, 688–691. [Google Scholar] [CrossRef] [PubMed]
- Yoshitama, K.; Hisada, M.; Ishikura, N. Distribution pattern of anthocyanins in the polygonaceae. J. Plan. Res. 1984, 97, 31–38. [Google Scholar] [CrossRef]
- You, Y.; Yuan, X.; Lee, H.J.; Huang, W.; Jin, W.; Zhan, J. Mulberry and mulberry wine extract increase the number of mitochondria during brown adipogenesis. Food Funct. 2015, 6, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.; Medvedev, A.V.; Daniel, K.W.; Collins, S. Beta-adrenergic activation of p38 MAP kinase in adipocytes: cAMP induction of the uncoupling protein 1 (UCP1) gene requires p38 map kinase. J. Biol. Chem. 2001, 276, 27077–27082. [Google Scholar] [CrossRef] [PubMed]
- Takikawa, M.; Inoue, S.; Horio, F.; Tsuda, T. Dietary anthocyanin-rich bilberry extract ameliorates hyperglycemia and insulin sensitivity via activation of amp-activated protein kinase in diabetic mice. J. Nutr. 2010, 140, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Matsukawa, T.; Villareal, M.O.; Motojima, H.; Isoda, H. Increasing camp levels of preadipocytes by cyanidin-3-glucoside treatment induces the formation of beige phenotypes in 3T3-L1 adipocytes. J. Nutr. Biochem. 2017, 40, 77–85. [Google Scholar] [CrossRef] [PubMed]
- You, Y.; Yuan, X.; Liu, X.; Liang, C.; Meng, M.; Huang, Y.; Han, X.; Guo, J.; Guo, Y.; Ren, C.; et al. Cyanidin-3-glucoside increases whole body energy metabolism by upregulating brown adipose tissue mitochondrial function. Mol. Nutr. Food Res. 2017, 61. [Google Scholar]
- Davis, S.R.; Dalais, F.S.; Simpson, E.R.; Murkies, A.L. Phytoestrogens in health and disease. Recent prog. Horm. Res. 1999, 54, 185–210. [Google Scholar] [PubMed]
- Tapas, A.R.; Sakarkar, D.; Kakde, R. Flavonoids as nutraceuticals: A review. Trop J. Pharm. Res. 2008, 7, 1089–1099. [Google Scholar] [CrossRef]
- Murphy, P.A.; Song, T.; Buseman, G.; Barua, K.; Beecher, G.R.; Trainer, D.; Holden, J. Isoflavones in retail and institutional soy foods. J. Agric. Food Chem. 1999, 47, 2697–2704. [Google Scholar] [CrossRef] [PubMed]
- Rogers, N.H.; Perfield, J.W.; Strissel, K.J.; Obin, M.S.; Greenberg, A.S. Reduced energy expenditure and increased inflammation are early events in the development of ovariectomy-induced obesity. Endocrinology 2009, 150, 2161–2168. [Google Scholar] [CrossRef] [PubMed]
- Mauvais-Jarvis, F.; Clegg, D.J.; Hevener, A.L. The role of estrogens in control of energy balance and glucose homeostasis. Endocr. Rev. 2013, 34, 309–338. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, T.; Nagamine, R.; Sameshima-Kamiya, M.; Tsubata, M.; Ikeguchi, M.; Takagaki, K. The isoflavone-rich fraction of the crude extract of the puerariae flower increases oxygen consumption and bat UCP1 expression in high-fat diet-fed mice. Glob. J. Health Sci. 2012, 4, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Lephart, E.D.; Porter, J.P.; Lund, T.D.; Bu, L.; Setchell, K.D.; Ramoz, G.; Crowley, W.R. Dietary isoflavones alter regulatory behaviors, metabolic hormones and neuroendocrine function in long-Evans male rats. Nutr. Metab. (Lond) 2004, 1, 16. [Google Scholar] [CrossRef] [PubMed]
- Crespillo, A.; Alonso, M.; Vida, M.; Pavon, F.J.; Serrano, A.; Rivera, P.; Romero-Zerbo, Y.; Fernandez-Llebrez, P.; Martinez, A.; Perez-Valero, V.; et al. Reduction of body weight, liver steatosis and expression of stearoyl-coa desaturase 1 by the isoflavone daidzein in diet-induced obesity. Br. J. Pharmacol. 2011, 164, 1899–1915. [Google Scholar] [CrossRef] [PubMed]
- Aziz, S.A.; Wakeling, L.A.; Miwa, S.; Alberdi, G.; Hesketh, J.E.; Ford, D. Metabolic programming of a beige adipocyte phenotype by genistein. Mol. Nutr. Food Res. 2017, 61, 1600574. [Google Scholar] [CrossRef] [PubMed]
- Gautam, J.; Khedgikar, V.; Kushwaha, P.; Choudhary, D.; Nagar, G.K.; Dev, K.; Dixit, P.; Singh, D.; Maurya, R.; Trivedi, R. Formononetin, an isoflavone, activates AMP-activated protein kinase/β-catenin signalling to inhibit adipogenesis and rescues C57BL/6 mice from high-fat diet-induced obesity and bone loss. Br. J. Nutr. 2017, 117, 645–661. [Google Scholar] [CrossRef] [PubMed]
- Fiorani, M.; Guidarelli, A.; Blasa, M.; Azzolini, C.; Candiracci, M.; Piatti, E.; Cantoni, O. Mitochondria accumulate large amounts of quercetin: Prevention of mitochondrial damage and release upon oxidation of the extramitochondrial fraction of the flavonoid. J. Nutr. Biochem. 2010, 21, 397–404. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.M.; Barger, J.L.; Edwards, M.G.; Braun, K.H.; O’Connor, C.E.; Prolla, T.A.; Weindruch, R. Dynamic regulation of PGC-1α localization and turnover implicates mitochondrial adaptation in calorie restriction and the stress response. Aging Cell 2008, 7, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Bai, W.; Zhang, X. Nucleus or cytoplasm? The mysterious case of sirt1′s subcellular localization. Cell Cycle 2016, 15, 3337–3338. [Google Scholar] [CrossRef] [PubMed]
- Qiang, L.; Wang, L.; Kon, N.; Zhao, W.; Lee, S.; Zhang, Y.; Rosenbaum, M.; Zhao, Y.; Gu, W.; Farmer, S.R.; et al. Brown remodeling of white adipose tissue by SirT1-dependent deacetylation of Pparγ. Cell 2012, 150, 620–632. [Google Scholar] [CrossRef] [PubMed]
- Grundlingh, J.; Dargan, P.I.; El-Zanfaly, M.; Wood, D.M. 2, 4-dinitrophenol (DNP): A weight loss agent with significant acute toxicity and risk of death. J. Med. Toxicol. 2011, 7, 205. [Google Scholar] [CrossRef] [PubMed]
- Law, J.; Morris, D.E.; Izzi-Engbeaya, C.; Salem, V.; Coello, C.; Robinson, L.; Jayasinghe, M.; Scott, R.; Gunn, R.; Rabiner, E.; et al. Thermal imaging is a noninvasive alternative to PET/CT for measurement of brown adipose tissue activity in humans. J. Nucl. Med. 2018, 59, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, A.P.; Gee, J.M.; Dupont, M.S.; Needs, P.W.; Mellon, F.A.; Williamson, G.; Johnson, I.T. Hydrolysis by lactase phlorizin hydrolase is the first step in the uptake of daidzein glucosides by rat small intestine in vitro. Xenobiotica 2003, 33, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Day, A.J.; Cañada, F.J.; Dı́az, J.C.; Kroon, P.A.; Mclauchlan, R.; Faulds, C.B.; Plumb, G.W.; Morgan, M.R.; Williamson, G. Dietary flavonoid and isoflavone glycosides are hydrolysed by the lactase site of lactase phlorizin hydrolase. FEBS Lett. 2000, 468, 166–170. [Google Scholar] [CrossRef]
- Zou, T.-B.; Feng, D.; Song, G.; Li, H.-W.; Tang, H.-W.; Ling, W.-H. The role of sodium-dependent glucose transporter 1 and glucose transporter 2 in the absorption of cyanidin-3-o-β-glucoside in caco-2 cells. Nutrients 2014, 6, 4165–4177. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-H.; Hsu, H.-J.; Huang, Y.-J.; Lin, C.-J. Interaction of flavonoids and intestinal facilitated glucose transporters. Planta Med. 2007, 73, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; You, L.; Zeng, S. Studies on the flavonoid substrates of human UDP-glucuronosyl transferase (UGT) 2B7. Die Pharmazie 2007, 62, 625–629. [Google Scholar] [PubMed]
- Zhang, L.; Zuo, Z.; Lin, G. Intestinal and hepatic glucuronidation of flavonoids. Mol. Pharm. 2007, 4, 833–845. [Google Scholar] [CrossRef] [PubMed]
- Mullen, W.; Edwards, C.A.; Crozier, A. Absorption, excretion and metabolite profiling of methyl-, glucuronyl-, glucosyl- and sulpho-conjugates of quercetin in human plasma and urine after ingestion of onions. Brit. J. Nutr. 2006, 96, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Strobel, P.; Allard, C.; Perez-Acle, T.; Calderon, R.; Aldunate, R.; Leighton, F. Myricetin, quercetin and catechin-gallate inhibit glucose uptake in isolated rat adipocytes. Biochem. J. 2005, 386, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Li, Y.; Brobbey Oppong, M.; Qiu, F. Insights into the intestinal bacterial metabolism of flavonoids and the bioactivities of their microbe-derived ring cleavage metabolites. Drug Metab. Rev. 2018, 16, 1–14. [Google Scholar] [CrossRef] [PubMed]



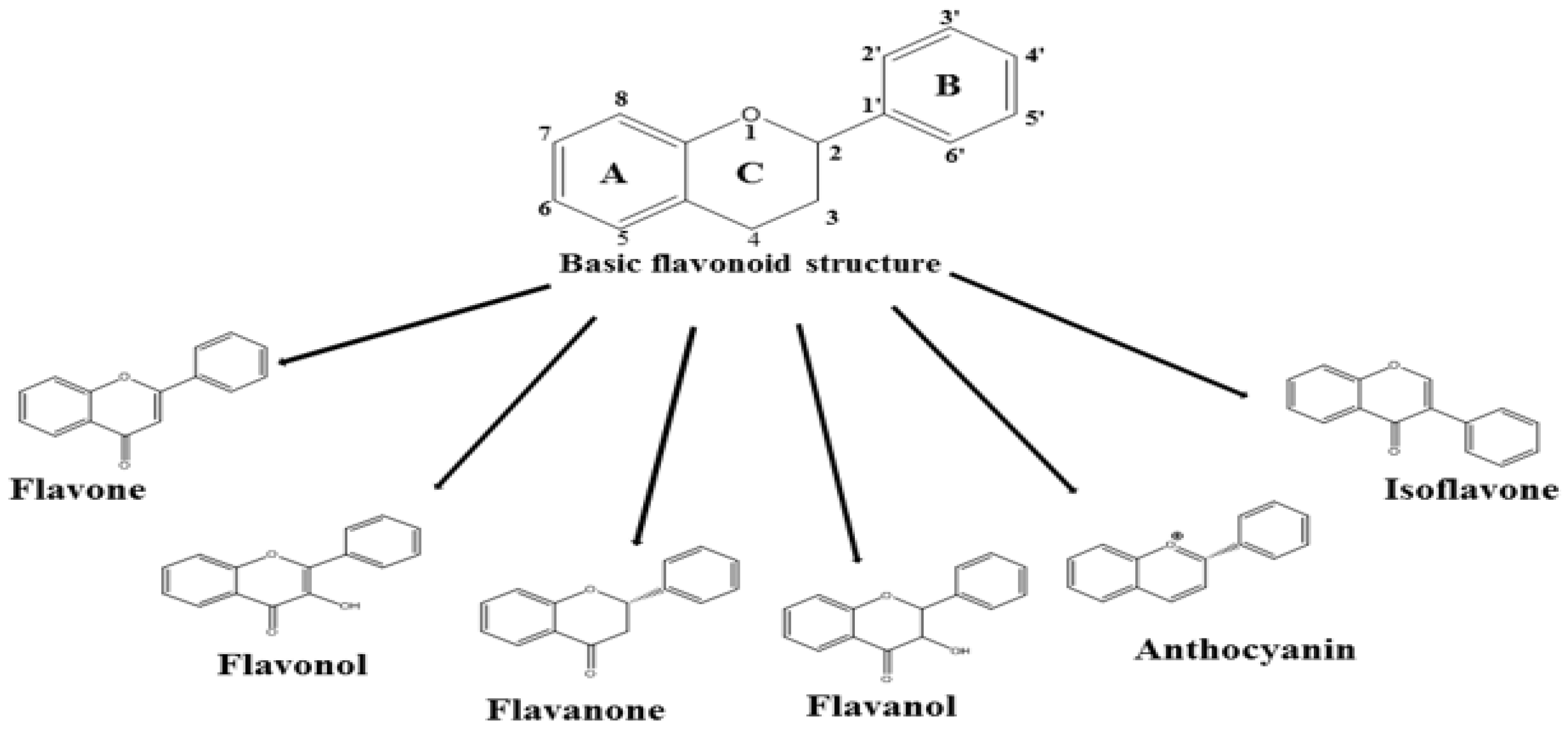{kind=link}
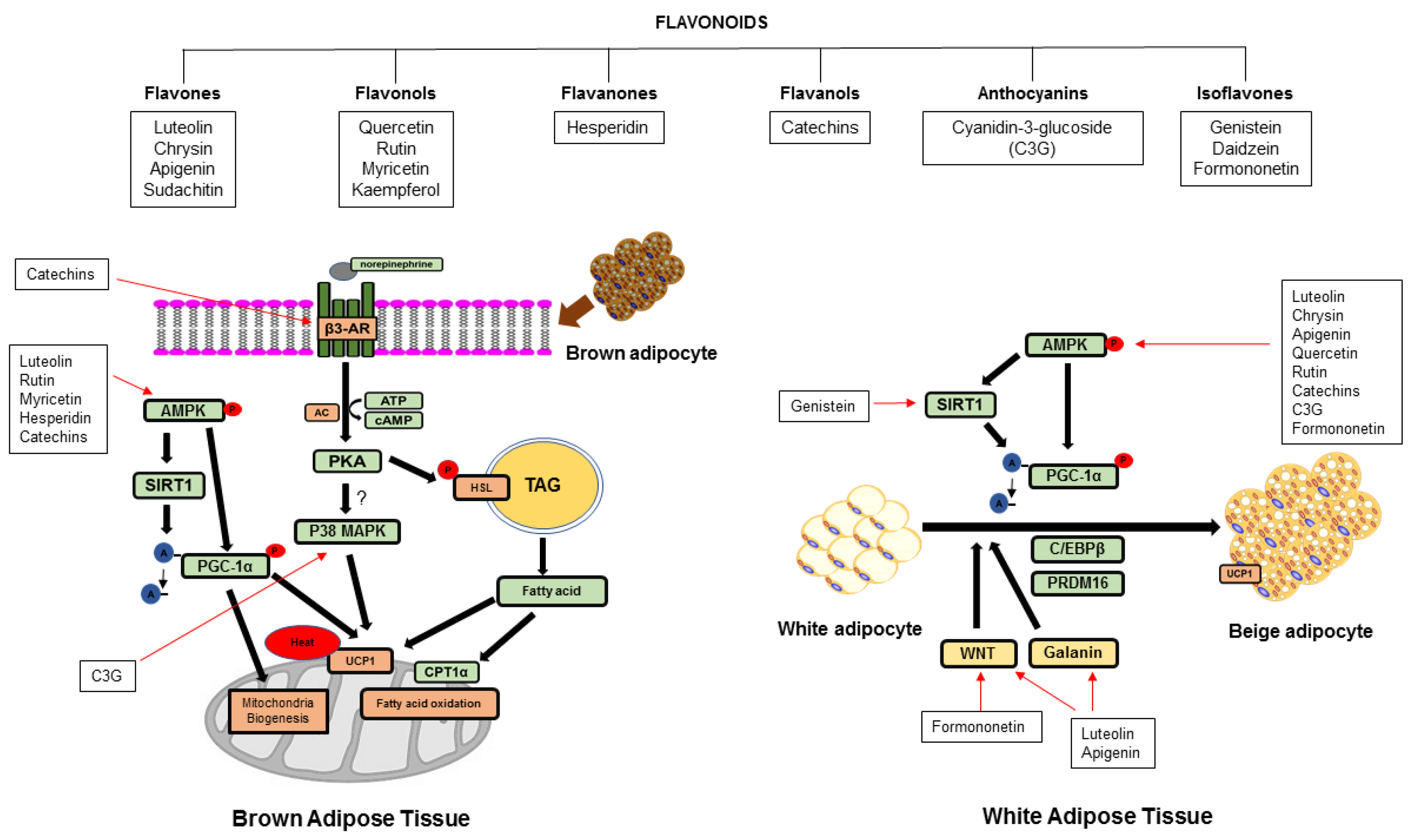{kind=link}
| Groups | Dietary Flavonoid Intakes (mean ± SD) a |
|---|---|
| Flavone | 1.6 ± 0.2 mg/day |
| Flavonol | 12.9 ± 0.4 mg/day |
| Flavanol | 156.5 ± 11.3 mg/day |
| Flavanone | 14.4 ± 0.6 mg/day |
| Anthocyanin | 3.1 ± 0.5 mg/day |
| Isoflavone | 1.2 ± 0.2 mg/day |
| Flavonoids | Subjects | Treatments | Outcomes | Authors | |
|---|---|---|---|---|---|
| Flavone | Luteolin | Primary adipocytes from BAT and SWAT | 100 nM | ↑ Ucp1, Pgc1α and Sirt1 in BAT and SWAT ↑ AMPK phosphorylation in BAT and SWAT | Zhang et al. [33] |
| Chrysin | 3T3-L1 | 50 µM | ↑ UCP1, Ucp1, PGC1α, Pgc1α PRDM16, Prdm16, FGF21, Fgf21, TBX1, Tbx1, TMEM26, Tmem26, CIDEA, Cidea and CITED 1, Cited1 ↑ AMPK phosphorylation | Choi et al. [39] | |
| Sudachitin | Primary myoblasts | 30 mM | ↑ Sirt1, Pgc1α, and Ucp1 | Tsutsumi et al. [50] | |
| Flavonol | Onion peel (quercetin) | 3T3-L1 | 25-100 µg/mL | ↑ Cpt1α, Fabp4 | Moon et al. [52] |
| Quercetin | 3T3-L1 | 25–100 µM | ↑ Ucp1, Cpt1α, Tbx1, Pgc1α, Pparγ, and Prdm16 | Lee et al. [54] | |
| Rutin | C3H10T1/2 cells | 0.1–100 µM | ↑ UCP1, Ucp1, Prdm16, Pgc1α, Tfam, Nrf1, and Nrf2 ↑ PGC1α deacetylation by stabilizing SIRT1 | Yuan et al. [57] | |
| Myricetin | C3H10T1/2 cells | 0.001–10 µM | ↑ Ucp1, UCP1, PGC1α, SIRT1, and adiponectin | Hu et al. [61] | |
| Flavanone | Gelidium elegans (Hesperidin) | 3T3-L1 | 12.5 and 50 µg/mL | ↑ UCP1 and PRDM16 | Choi et al. [69] |
| Anthocyanin | Mulberry extract (ME), mulberry wine extract (MWE) and cyanidin-3-glucoside (C3G) | C3H10T1/2 cells | ME and MWE (10 µg/mL) of C3G (1–100 µM) | ↑ UCP1, Ucp1, Pgc1α, Cpt1α, and Prdm16 by ME and MWE ↑ Phosphorylation of p38 MAPK by ME ↑ UCP1, Ucp1, Pgc1α, Pgc1β, Prdm16, Nrf1, mitochondrial copy number, and cellular oxygen respiration by C3G | You et al. [94] |
| C3G | 3T3-L1 | 50 or 100 µM | ↑ Cellular cAMP concentration ↑ AMPK phosphorylation ↑ FABP4, UCP1, PGC1α expression ↑ Mitochondrial biogenesis ↑ C/ebpβ, Tbx1 and Cited 1 | Matsukawa et al. [97] | |
| Isoflavone | Genistein | 3T3-L1 | 100 µM | ↑ Ucp1, Pgc1α, and Sirt1 ↑ Oxygen consumption | Aziz et al. [107] |
| Formononetin | 3T3-L1 | 10 nM | ↑ AMPK phosphorylation and β-catenin expression | Gautam et al. [108] | |
| Flavonoids | Subjects | Treatments | Outcomes | Authors | |
|---|---|---|---|---|---|
| Flavone | Luteolin | Male C57BL/6 mice | HFD with 0.01% luteolin for 12 weeks | ↑ O2 consumption and CO2 production ↑ BAT activity ↑ SWAT browning ↑ AMPK/PGC1α signaling | Zhang et al. [33] |
| Olive leaf extract (luteolin) | Male C57BL/6N mice | HFD with 0.15% olive leaf extract for 8 weeks | ↓ Body weight and fat pad weight ↑ Browning and mitochondrial biogenesis in EWAT | Shen et al. [43] | |
| Apigenin mixed with naringenin | Male C57BL/6 mice | Apigenin/naringenin (80 mg/kg) for 2 weeks | ↑ UCP1 in BAT | Thaiss et al. [46] | |
| Sudachitin | C57BL/6 J mice and db/db mice | HFD with 5 mg/kg sudachitin for 12 weeks | ↓ Body weight, subcutaneous and visceral fat contents ↑ O2 consumption and energy expenditure ↑ Ucp1 in SWAT | Tsutsumi et al. [50] | |
| Flavonol | Onion peel Extract (quercetin) | Male SD rats | HFD with 0.36 and 0.72% onion peel extract for 8 weeks | ↓ Body weight and weights of total visceral, retroperitoneal, and mesenteric fat fads ↑ Ucp1 and Cpt1α in EWAT | Moon et al. [52] |
| Onion peel extract (quercetin) | Male C57BL/6 mice | HFD with 0.5% onion peel for 8 weeks | ↑ Adipocyte browning in RWAT and SWAT | Lee et al. [54] | |
| Quercetin | Male C57BL/6 mice | HFD with 0.1% Quercetin for 12 weeks | ↓ Body weight and weights of EWAT and SWAT ↑ AMPK phosphorylation, and SIRT1 expression in EWAT ↑ Ucp1 in BAT | Dong et al. [53] | |
| Rutin | Male C57BL/6 J mice and C57BLKS/J-(db/db) mice | HFD with rutin (1 mg/mL) in drinking water for 10 weeks | ↑ Mitochondria biogenesis and whole-body energy expenditure ↑ BAT activity and SWAT browning | Yuan et al. [57] | |
| Rutin | Female polycystic ovary syndrome SD-rats | Rutin (100 mg/kg) in drinking water for 3 weeks | ↑ UCP1, Ucp1, Pparα, Pgc1α, Pgc1β, and Cpt1α in BAT, ↑ Body temperature | Hu et al. [60] | |
| Myricetin | Male C57BLKS/J-(db/db) mice | HFD with myricetin (400 mg/kg) in drinking water for 14 weeks | ↓ Body weight, fat mass, and blood glucose ↑ Body temperature and oxygen consumption ↑ BAT activity ↑ IWAT browning and mitochondrial biogenesis | Hu et al. [61] | |
| Gelidium elegans (hesperidin rich) | Male ICR mice | HFD with Gelidium elegans extract (50, 200 mg/kg/day) for 7 weeks | ↓ Body weight, fat mass, plasma insulin, and TG level ↑ AMPK phosphorylation in BAT and BAT activity | Choi et al. [68] | |
| G-hesperidin | Male Wistar rats | 60 mg of G-hesperidin by acute oral administration | ↑ BAT-sympathetic nerve activity ↑ Body temperature ↓ Cutaneous sympathetic nerve activity | Shen et al. [70] | |
| Flavanal | Green tea extract (catechin and EGCG) | Male SD rats | Chow diet with catechin and EGCG (0–200 µM) in drinking water | ↑ BAT activity and O2 uptake rate | Dulloo et al. [75] |
| Green tea (EGCG) | Male SD rats | HFD with green tea extract (20 g/kg) | ↓ Body weight, digestibility ↑ Energy expenditure ↑ BAT density and β-adrenoceptor activity | Choo et al. [76] | |
| Tea catechins (TC) | Male SD rats | Low fat diet (LFD) and HFD with 0.5% TC for 5 weeks | ↑ Ucp1 in BAT of LFD with TC group - No significant difference between HF and HF with TC | Nomura et al. [77] | |
| Green tea catechins | Male SD rats | LFD and HFD with green tea catechins (100 mg/kg) for 5 weeks | ↑ PPARδ, UCP1, Ucp1, and CPT1α in WAT and BAT | Yan et al. [78] | |
| Oolong, black, and pure teas | Male ICR mice | 7 days consumption with tea boiled with 2 g tea leaves in 100 mL | ↓ Weight of WAT, ↑ AMPK phosphorylation in WAT and BAT, ↑ UCP1 in WAT | Yamashita et al. [82] | |
| Catechin | Healthy young women | 540 mg/day; catechin for 12 weeks | ↑ BAT density | Nirengi et al. [84] | |
| Epigalo catechin gallate (EGCG) | Healthy young men | Cold exposure for 3 h after 1600 mg of EGCG and 600 mg of caffeine intake | ↑ Energy expenditure ↓ Shivering thermogenesis | Gosselin et al. [85] | |
| Cocoa flavanols | Male ICR mice | 10 mg/kg cocoa flavonoid fraction or epicatechin. | ↑ BAT activity ↑ AMPK phosphorylation in BAT ↑ Plasma catecholamine level | Matsumura et al. [85] | |
| Cocoa flavanols | Male Wistar rat | HFD with cocoa powder 1 g/kg, cocoa extract 100 mg/kg and (-)-epicatechin 10 mg/kg for 8 weeks | ↑ Ucp1, Pparγ, Pparα, Sirt1, and Pgc1α in BAT ↑ AMPK phosphorylation in BAT ↑ Plasma catecholamine level | Rabadan-Chávez et al. [88] | |
| Epicatechin | Male Wistar rats | HFD for 5 weeks with (-)-epicatechin (1 mg/kg) for additional 2 weeks | ↑ EWAT browning ↓ Body weight | Gutiérrez-Salmeán et al. [89] | |
| Anthocyanin | Bilberry Extract | Male KK-Ay mice | 27 g/kg diet for 5weeks | ↑ AMPK in SWAT and skeletal muscle | Takikawa et al. [96] |
| Cyanidin-3-glucoside (C3G) | Male C57BLKS/J-(db/db) mice | C3G dissolved in drinking water (1 mg/mL) for 16 weeks | ↑ Energy expenditure representing oxygen consumption ↑ BAT activity ↑ Body temperature and mitochondrial biogenesis in BAT ↑ SWAT browning ↓ Body weight gain, and weight of EWAT and SWAT | You et al. [98] | |
| Isoflavone | Puerariae flower (PFE) and isoflavone fraction (PF) | Male C57BL/6J mice | HFD with 5% PFE and PF isoflavone fraction for 6 weeks | ↑ Energy expenditure representing oxygen consumption ↑ UCP1-positive area in BAT ↓ Body weight gain, and weight of EWAT and SWAT | Kamiya et al. [104] |
| Isoflavone mixture | Long-Evans male and female rats | 600 µg of phytoestrogens/g of diet | ↑ Core body temperature during light cycle ↑ Ucp1 in BAT | Lephart et al. [105] | |
| Daidzein | Male Wistar rats | LFD and HFD with 50 mg/kg for 2 weeks | ↑ Ucp1 in BAT in HFD | Crespillo et al. [106] | |
| Formononetin | Male C57BL/6J mice | HFD with 0.1, 1, and 10 mg of formononetin | ↑ SWAT browning ↑ Small and multilocular lipid droplets in SWAT | Gautam et al. [108] | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, H.W.; Lee, S.G.; Otieno, D.; Ha, K. Flavonoids, Potential Bioactive Compounds, and Non-Shivering Thermogenesis. Nutrients 2018, 10, 1168. https://doi.org/10.3390/nu10091168
Kang HW, Lee SG, Otieno D, Ha K. Flavonoids, Potential Bioactive Compounds, and Non-Shivering Thermogenesis. Nutrients. 2018; 10(9):1168. https://doi.org/10.3390/nu10091168
Chicago/Turabian StyleKang, Hye Won, Sang Gil Lee, Dammah Otieno, and Kyoungsoo Ha. 2018. "Flavonoids, Potential Bioactive Compounds, and Non-Shivering Thermogenesis" Nutrients 10, no. 9: 1168. https://doi.org/10.3390/nu10091168
APA StyleKang, H. W., Lee, S. G., Otieno, D., & Ha, K. (2018). Flavonoids, Potential Bioactive Compounds, and Non-Shivering Thermogenesis. Nutrients, 10(9), 1168. https://doi.org/10.3390/nu10091168