


Vesicular Stomatitis Virus Elicits Early Transcriptome Response in Culicoides sonorensis Cells

Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Virus
2.2. Time Course Infection of W8 Cells with VSNJV
2.3. VSNJV Quantification
2.4. W8 RNA Extraction, Sequencing, and QC
2.5. De Novo Assembly, Reduction, and Annotation from W8 Cells
2.6. Differential Expression from W8 Cells
2.7. Statistical Analysis
3. Results
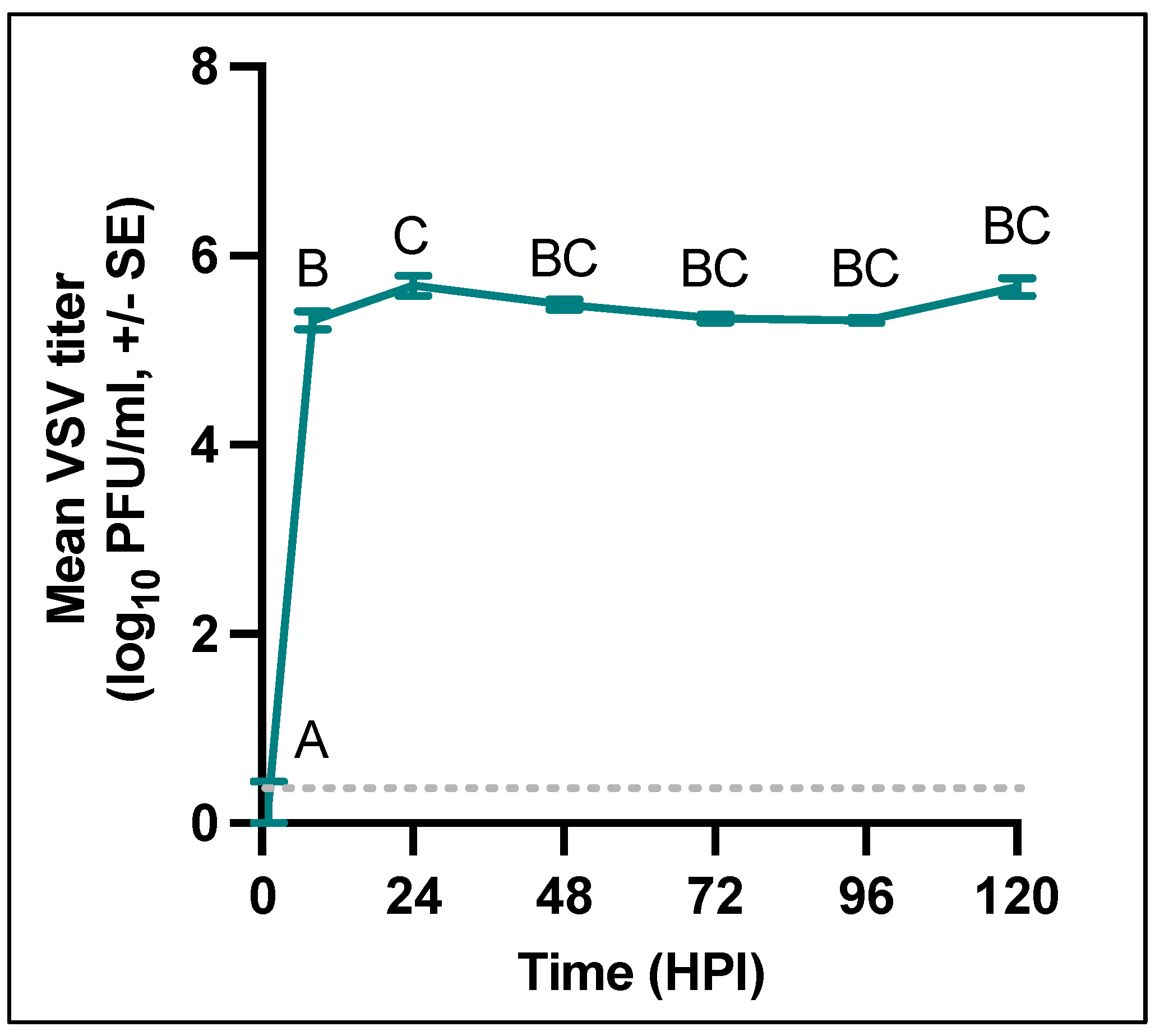
3.1. VSNJV Rapidly Produces Infectious Virus W8 Cells
3.2. W8 Transcriptome Response to VSNJV Infection
3.3. VSNJV Transcriptome from W8 Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McCombs, R.M.; Benyesh-Melnick, M.; Brunschwig, J.P. Biophysical studies of vesicular stomatitis virus. J. Bacteriol. 1966, 91, 803–812. [Google Scholar] [CrossRef] [PubMed]
- Rozo-Lopez, P.; Drolet, B.S.; Londoño-Renteria, B. Vesicular stomatitis virus transmission: A comparison of incriminated vectors. Insects 2018, 9, 190. [Google Scholar] [CrossRef] [PubMed]
- Letchworth, G.; Rodriguez, L.; Del Cbarrera, J. Vesicular stomatitis. Vet. J. 1999, 157, 239–260. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, L.L. Emergence and re-emergence of vesicular stomatitis in the United States. Virus Res. 2002, 85, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Pelzel-McClusky, A.M. Vesicular stomatitis in large animals. In Merck Veterinary Manual, Online Edition; Merck & Co., Inc.: Kenilworth, NJ, USA, 2022. [Google Scholar]
- Alderink, F.J. Vesicular stomatitis epidemic in Colorado: Clinical observations and financial losses reported by dairymen. Prev. Vet. Med. 1984, 3, 29–44. [Google Scholar] [CrossRef]
- Goodger, W.; Thurmond, M.; Nehay, J.; Mitchell, J.; Smith, P. Economic impact of an epizootic of bovine vesicular stomatitis in California. J. Am. Vet. Med. Assoc. 1985, 186, 370–373. [Google Scholar]
- Hayek, A.; McCluskey, B.; Chavez, G.; Salman, M. Financial impact of the 1995 outbreak of vesicular stomatitis on 16 beef ranches in Colorado. J. Am. Vet. Med. Assoc. 1998, 212, 820–823. [Google Scholar]
- Drolet, B.S.; Campbell, C.L.; Stuart, M.A.; Wilson, W.C. Vector competence of Culicoides sonorensis (Diptera: Ceratopogonidae) for vesicular stomatitis virus. J. Med. Entomol. 2005, 42, 409–418. [Google Scholar] [CrossRef]
- De León, A.A.P.; O’Toole, D.; Tabachnick, W.J. Infection of guinea pigs with vesicular stomatitis New Jersey virus transmitted by Culicoides sonorensis (Diptera: Ceratopogonidae). J. Med. Entomol. 2006, 43, 568–573. [Google Scholar] [CrossRef]
- De Leon, A.A.P.; Tabachnick, W.J. Transmission of vesicular stomatitis New Jersey virus to cattle by the biting midge Culicoides sonorensis (Diptera: Ceratopogonidae). J. Med. Entomol. 2006, 43, 323–329. [Google Scholar] [CrossRef]
- Rozo-Lopez, P.; Londono-Renteria, B.; Drolet, B.S. Venereal transmission of vesicular stomatitis virus by Culicoides sonorensis midges. Pathogens 2020, 9, 316. [Google Scholar] [CrossRef]
- Mullens, B.A.; Schmidtmann, E.T. The gonotrophic cycle of Culicoides variipennis (Diptera: Ceratopogonidae) and its implications in age-grading field populations in New York State, USA. J. Med. Entomol. 1982, 19, 340–349. [Google Scholar] [CrossRef]
- Nayduch, D.; Lee, M.B.; Saski, C.A. Gene discovery and differential expression analysis of humoral immune response elements in female Culicoides sonorensis (Diptera: Ceratopogonidae). Parasites Vectors 2014, 7, 388. [Google Scholar] [CrossRef]
- Sim, S.; Dimopoulos, G. Dengue virus inhibits immune responses in Aedes aegypti cells. PLoS ONE 2010, 5, e10678. [Google Scholar] [CrossRef]
- Schnettler, E.; Ratinier, M.; Watson, M.; Shaw, A.E.; McFarlane, M.; Varela, M.; Elliott, R.M.; Palmarini, M.; Kohl, A. RNA interference targets arbovirus replication in Culicoides cells. J. Virol. 2013, 87, 2441–2454. [Google Scholar] [CrossRef]
- Mills, M.K.; Nayduch, D.; Michel, K. Inducing RNA interference in the arbovirus vector, Culicoides sonorensis. Insect Mol. Biol. 2015, 24, 105–114. [Google Scholar] [CrossRef]
- Cheng, G.; Liu, Y.; Wang, P.; Xiao, X. Mosquito defense strategies against viral infection. Trends Parasitol. 2016, 32, 177–186. [Google Scholar] [CrossRef]
- Hajdušek, O.; Šíma, R.; Ayllón, N.; Jalovecká, M.; Perner, J.; De La Fuente, J.; Kopáček, P. Interaction of the tick immune system with transmitted pathogens. Front. Cell. Infect. Microbiol. 2013, 3, 26. [Google Scholar] [CrossRef] [PubMed]
- Nayduch, D.; Shankar, V.; Mills, M.K.; Robl, T.; Drolet, B.S.; Ruder, M.G.; Scully, E.D.; Saski, C.A. Transcriptome response of female Culicoides sonorensis biting midges (Diptera: Ceratopogonidae) to early infection with epizootic hemorrhagic disease virus (EHDV-2). Viruses 2019, 11, 473. [Google Scholar] [CrossRef] [PubMed]
- McHolland, L.E.; Mecham, J.O. Characterization of cell lines developed from field populations of culicoides sonorensis (Diptera: Ceratopogonidae). J. Med. Entomol. 2003, 40, 348–351. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ghosh, A.; Jasperson, D.; Cohnstaedt, L.W.; Brelsfoard, C.L. Transfection of Culicoides sonorensis biting midge cell lines with Wolbachia pipientis. Parasites Vectors 2019, 12, 483. [Google Scholar] [CrossRef] [PubMed]
- Chen, S. Ultrafast one-pass FASTQ data preprocessing, quality control, and deduplication using fastp. iMeta 2023, 2, e107. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, D. Gene-omes built from mRNA seq not genome DNA. In Proceedings of the 7th Annual Arthropod Genomics Symposium, Notre Dame, IN, USA, 12 June 2023. [Google Scholar]
- Wood, D.E.; Lu, J.; Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Rincon, N.; Wood, D.E.; Breitwieser, F.P.; Pockrandt, C.; Langmead, B.; Salzberg, S.L.; Steinegger, M. Metagenome analysis using the Kraken software suite. Nat. Protoc. 2022, 17, 2815–2839. [Google Scholar] [CrossRef]
- Bryant, D.M.; Johnson, K.; DiTommaso, T.; Tickle, T.; Couger, M.B.; Payzin-Dogru, D.; Lee, T.J.; Leigh, N.D.; Kuo, T.-H.; Davis, F.G. A tissue-mapped axolotl de novo transcriptome enables identification of limb regeneration factors. Cell Rep. 2017, 18, 762–776. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.; Tosatto, S.C.; Paladin, L.; Raj, S.; Richardson, L.J. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Boutet, E.; Lieberherr, D.; Tognolli, M.; Schneider, M.; Bairoch, A. UniProtKB/Swiss-Prot: The manually annotated section of the UniProt KnowledgeBase. In Plant Bioinformatics: Methods and Protocols; Springer: Berlin, Germany, 2007; pp. 89–112. [Google Scholar]
- UniProtConsortium. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar] [CrossRef]
- Patro, R.; Duggal, G.; Love, M.I.; Irizarry, R.A.; Kingsford, C. Salmon provides fast and bias-aware quantification of transcript expression. Nat. Methods 2017, 14, 417–419. [Google Scholar] [CrossRef]
- RCoreTeam. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Soneson, C.; Love, M.I.; Robinson, M.D. Differential analyses for RNA-seq: Transcript-level estimates improve gene-level inferences. F1000Research 2015, 4, 1521. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Van Gelder, R.N.; Bae, H.; Palazzolo, M.J.; Krasnow, M.A. Extent and character of circadian gene expression in Drosophila melanogaster: Identification of twenty oscillating mRNAs in the fly head. Curr. Biol. 1995, 5, 1424–1436. [Google Scholar] [CrossRef] [PubMed]
- Van Gelder, R.N.; Krasnow, M.A. A novel circadianly expressed Drosophila melanogaster gene dependent on the period gene for its rhythmic expression. EMBO J. 1996, 15, 1625–1631. [Google Scholar] [CrossRef]
- Guo, S.; Yang, P.; Liang, B.; Zhou, F.; Hou, L.; Kang, L.; Wang, X. Aging features of the migratory locust at physiological and transcriptional levels. BMC Genom. 2021, 22, 257. [Google Scholar] [CrossRef]
- Choi, K.C.; Lee, Y.S.; Lim, S.; Choi, H.K.; Lee, C.H.; Lee, E.K.; Hong, S.; Kim, I.H.; Kim, S.J.; Park, S.H. Smad6 negatively regulates interleukin 1-receptor-Toll-like receptor signaling through direct interaction with the adaptor Pellino-1. Nat. Immunol. 2006, 7, 1057–1065. [Google Scholar] [CrossRef]
- Nakamoto, M.; Moy, R.H.; Xu, J.; Bambina, S.; Yasunaga, A.; Shelly, S.S.; Gold, B.; Cherry, S. Virus recognition by Toll-7 activates antiviral autophagy in Drosophila. Immunity 2012, 36, 658–667. [Google Scholar] [CrossRef]
- Wang, J.; Dupuis, C.; Tyring, S.K.; Underbrink, M.P. Sterile alpha Motif Domain Containing 9 Is a Novel Cellular Interacting Partner to Low-Risk Type Human Papillomavirus E6 Proteins. PLoS ONE 2016, 11, e0149859. [Google Scholar]
- Jouvenet, N.; Monaghan, P.; Way, M.; Wileman, T. Transport of African swine fever virus from assembly sites to the plasma membrane is dependent on microtubules and conventional kinesin. J. Virol. 2004, 78, 7990–8001. [Google Scholar] [CrossRef]
- Strunze, S.; Engelke, M.F.; Wang, I.H.; Puntener, D.; Boucke, K.; Schleich, S.; Way, M.; Schoenenberger, P.; Burckhardt, C.J.; Greber, U.F. Kinesin-1-mediated capsid disassembly and disruption of the nuclear pore complex promote virus infection. Cell Host Microbe 2011, 10, 210–223. [Google Scholar] [CrossRef]
- Hombria, J.C.; Sotillos, S. JAK/STAT signalling: STAT cannot play with Ken and Barbie. Curr. Biol. 2006, 16, R98–R100. [Google Scholar] [CrossRef] [PubMed]
- Arbouzova, N.I.; Bach, E.A.; Zeidler, M.P. Ken & barbie selectively regulates the expression of a subset of Jak/STAT pathway target genes. Curr. Biol. 2006, 16, 80–88. [Google Scholar] [PubMed]
- Orvedahl, A.; MacPherson, S.; Sumpter, R.; Tallóczy, Z.; Zou, Z.; Levine, B. Autophagy protects against Sindbis virus infection of the central nervous system. Cell Host Microbe 2010, 7, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Brackney, D.E.; Correa, M.A.; Cozens, D.W. The impact of autophagy on arbovirus infection of mosquito cells. PLoS Negl. Trop. Dis. 2020, 14, e0007754. [Google Scholar] [CrossRef]
- Ruan, X.; Zhang, R.; Li, R.; Zhu, H.; Wang, Z.; Wang, C.; Cheng, Z.; Peng, H. The research progress in physiological and pathological functions of TRAF4. Front. Oncol. 2022, 12, 842072. [Google Scholar] [CrossRef]
- Ma, T.H.; Benzie, J.A.; He, J.-G.; Sun, C.-B.; Chan, S.F. PmPPAF is a pro-phenoloxidase activating factor involved in innate immunity response of the shrimp Penaeus monodon. Dev. Comp. Immunol. 2014, 44, 163–172. [Google Scholar] [CrossRef]
- Sanfilippo, C.; Nunnari, G.; Calcagno, A.; Malaguarnera, L.; Blennow, K.; Zetterberg, H.; Di Rosa, M. The chitinases expression is related to Simian Immunodeficiency Virus Encephalitis (SIVE) and in HIV encephalitis (HIVE). Virus Res. 2017, 227, 220–230. [Google Scholar] [CrossRef][Green Version]
- Lee, C.G.; Da Silva, C.A.; Dela Cruz, C.S.; Ahangari, F.; Ma, B.; Kang, M.J.; He, C.H.; Takyar, S.; Elias, J.A. Role of chitin and chitinase/chitinase-like proteins in inflammation, tissue remodeling, and injury. Annu. Rev. Physiol. 2011, 73, 479–501. [Google Scholar] [CrossRef]
- Willecke, M.; Hamaratoglu, F.; Kango-Singh, M.; Udan, R.; Chen, C.-L.; Tao, C.; Zhang, X.; Halder, G. The fat cadherin acts through the hippo tumor-suppressor pathway to regulate tissue size. Curr. Biol. 2006, 16, 2090–2100. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhou, R.; Xu, P. The Hippo pathway in innate anti-microbial immunity and anti-tumor immunity. Front. Immunol. 2020, 11, 1473. [Google Scholar] [CrossRef]
- Kumar, V.; Garg, S.; Gupta, L.; Gupta, K.; Diagne, C.T.; Misse, D.; Pompon, J.; Kumar, S.; Saxena, V. Delineating the role of Aedes aegypti ABC transporter gene family during mosquito development and arboviral infection via transcriptome analyses. Pathogens 2021, 10, 1127. [Google Scholar] [CrossRef] [PubMed]
- Accaoui, M.J.; Enoiu, M.; Mergny, M.; Masson, C.; Dominici, S.; Wellman, M.; Visvikis, A. Gamma-glutamyltranspeptidase-dependent glutathione catabolism results in activation of NF-kB. Biochem. Biophys. Res. Commun. 2000, 276, 1062–1067. [Google Scholar] [CrossRef] [PubMed]
- Ball, L.A.; White, C.N. Order of transcription of genes of vesicular stomatitis virus. Proc. Natl. Acad. Sci. USA 1976, 73, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Abraham, G.; Banerjee, A.K. Sequential transcription of the genes of vesicular stomatitis virus. Proc. Natl. Acad. Sci. USA 1976, 73, 1504–1508. [Google Scholar] [CrossRef]
- Iverson, L.E.; Rose, J.K. Localized attenuation and discontinuous synthesis during vesicular stomatitis virus transcription. Cell 1981, 23, 477–484. [Google Scholar] [CrossRef]
- Lichty, B.D.; Power, A.T.; Stojdl, D.F.; Bell, J.C. Vesicular stomatitis virus: Re-inventing the bullet. Trends Mol. Med. 2004, 10, 210–216. [Google Scholar] [CrossRef]
- Stojdl, D.F.; Lichty, B.D.; Paterson, J.M.; Power, A.T.; Knowles, S.; Marius, R.; Reynard, J.; Poliquin, L.; Atkins, H.; Brown, E.G. VSV strains with defects in their ability to shutdown innate immunity are potent systemic anti-cancer agents. Cancer Cell 2003, 4, 263–275. [Google Scholar] [CrossRef]
- Von Kobbe, C.; Van Deursen, J.M.; Rodrigues, J.P.; Sitterlin, D.; Bachi, A.; Wu, X.; Wilm, M.; Carmo-Fonseca, M.; Izaurralde, E. Vesicular stomatitis virus matrix protein inhibits host cell gene expression by targeting the nucleoporin Nup98. Mol. Cell 2000, 6, 1243–1252. [Google Scholar] [CrossRef]
- Petersen, J.M.; Her, L.-S.; Varvel, V.; Lund, E.; Dahlberg, J.E. The matrix protein of vesicular stomatitis virus inhibits nucleocytoplasmic transport when it is in the nucleus and associated with nuclear pore complexes. Mol. Cell. Biol. 2000, 20, 8590–8601. [Google Scholar] [CrossRef]
- Petersen, J.M.; Her, L.-S.; Dahlberg, J.E. Multiple vesiculoviral matrix proteins inhibit both nuclear export and import. Proc. Natl. Acad. Sci. USA 2001, 98, 8590–8595. [Google Scholar] [CrossRef]
- Black, B.L.; Lyles, D.S. Vesicular stomatitis virus matrix protein inhibits host cell-directed transcription of target genes in vivo. J. Virol. 1992, 66, 4058–4064. [Google Scholar] [CrossRef] [PubMed]
- Black, B.L.; Rhodes, R.; McKenzie, M.; Lyles, D. The role of vesicular stomatitis virus matrix protein in inhibition of host-directed gene expression is genetically separable from its function in virus assembly. J. Virol. 1993, 67, 4814–4821. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Lyles, D.S. Effect of vesicular stomatitis virus matrix protein on transcription directed by host RNA polymerases I, II, and III. J. Virol. 1998, 72, 8413–8419. [Google Scholar] [CrossRef] [PubMed]
- Neidermyer, W.J., Jr.; Whelan, S.P. Global analysis of polysome-associated mRNA in vesicular stomatitis virus infected cells. PLoS Pathog. 2019, 15, e1007875. [Google Scholar] [CrossRef] [PubMed]
- Etebari, K.; Hegde, S.; Saldaña, M.A.; Widen, S.G.; Wood, T.G.; Asgari, S.; Hughes, G.L. Global transcriptome analysis of Aedes aegypti mosquitoes in response to Zika virus infection. MSphere 2017, 2, e00456-17. [Google Scholar] [CrossRef] [PubMed]
- Velikkakath, A.K.G.; Nishimura, T.; Oita, E.; Ishihara, N.; Mizushima, N. Mammalian Atg2 proteins are essential for autophagosome formation and important for regulation of size and distribution of lipid droplets. Mol. Biol. Cell. 2012, 23, 896–909. [Google Scholar] [CrossRef]
- Imamura, T.; Takase, M.; Nishihara, A.; Oeda, E.; Hanai, J.-i.; Kawabata, M.; Miyazono, K. Smad6 inhibits signalling by the TGF-β superfamily. Nature 1997, 389, 622–626. [Google Scholar] [CrossRef]
- Takeshita, F.; Ishii, K.J.; Kobiyama, K.; Kojima, Y.; Coban, C.; Sasaki, S.; Ishii, N.; Klinman, D.M.; Okuda, K.; Akira, S. TRAF4 acts as a silencer in TLR-mediated signaling through the association with TRAF6 and TRIF. Eur. J. Immunol. 2005, 35, 2477–2485. [Google Scholar] [CrossRef]
- Marinis, J.M.; Homer, C.R.; McDonald, C.; Abbott, D.W. A novel motif in the Crohn’s disease susceptibility protein, NOD2, allows TRAF4 to down-regulate innate immune responses. J. Biol. Chem. 2011, 286, 1938–1950. [Google Scholar] [CrossRef]
- Marinis, J.M.; Hutti, J.E.; Homer, C.R.; Cobb, B.A.; Cantley, L.C.; McDonald, C.; Abbott, D.W. IκB kinase α phosphorylation of TRAF4 downregulates innate immune signaling. Mol. Cell. Biol. 2012, 32, 2479–2489. [Google Scholar] [CrossRef]
- DuRaine, G.; Wisner, T.W.; Howard, P.; Johnson, D.C. Kinesin-1 proteins KIF5A, -5B, and -5C promote anterograde transport of herpes simplex virus enveloped virions in axons. J. Virol. 2018, 92, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Das, S.C.; Nayak, D.; Zhou, Y.; Pattnaik, A.K. Visualization of intracellular transport of vesicular stomatitis virus nucleocapsids in living cells. J. Virol. 2006, 80, 6368–6377. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Annotation | Name | Log2 FC | FDR p-Value | Possible Immune or Proviral Function |
|---|---|---|---|---|
| REG5 | Rhythmically expressed gene 5 | −2.41 | 4.72 × 10−65 | Unknown |
| SMAD6 | Mothers against decapentaplegic homolog 6 | −2.27 | 5.23 × 10−266 | Immune [40] |
| DSX | Protein doublesex | −1.77 | 4.88 × 10−8 | Unknown |
| CUD2 | Endocuticle structural glycoprotein | −1.55 | 1.23 × 10−48 | Unknown |
| LIPT | Lipoyltransferae 1 * | −1.35, −1.36, −1.35 | 1.15 × 10−15, 0.007, 8.35 × 10−24 | Unknown |
| TOLL7 | Toll-like receptor 7 * | −1.02, −0.95 | 1.64 × 10−17, 5.17 × 10−7 | Immune [41] |
| SAM11 | Sterile alpha motif domain-containing protein 11 | −0.93 | 2.54 × 10−4 | Immune [42] |
| KI26L | Kinesin-like protein * | −0.82, −0.85 | 3.00 × 10−13, 5.10 × 10−5 | Proviral [43,44] |
| KEN1 | Transcription factor Ken1 | −0.81 | 0.02 | Immune [45,46] |
| ATG2B | Autophagy-related protein 2 homolog B | 0.83 | 7.75 × 10−13 | Immune, proviral [47,48] |
| TRAF4 | TNF receptor-associated factor 4 | 0.84 | 6.59 × 10−4 | Immune [49] |
| NAAT1 | Sodium-dependent nutrient amino acid transporter 1 | 1.03 | 2.53 × 10−20 | Unknown |
| PPAF3 | Phenoloxidase-activating factor 3 | 1.30 | 1.73 × 10−7 | Immune [50] |
| CHIT1 | Chitotriosidase 1 | 1.55 | 3.03 × 10−8 | Immune [51,52] |
| FAT | Cadherin-related tumor suppressor/FAT tumor suppressor homolog 1 | 1.78 | 6.51 × 10−6 | Immune [53,54] |
| CP4CU | Cytochrome P450 4c21 | 1.93 | 0.005 | Unknown |
| SCX | Basic helix-loop-helix transcription factor scleraxis | 1.97 | 0.009 | Unknown |
| ABCA3 | Phospholipid-transporting ATPase ABCA3 | 2.44, 1.43 | 0.02, 3.72 × 10−6 | Immune [55] |
| GGT | Gamma-glutamytranspeptidase 1 | 2.47 | 0.02 | Immune [56] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scroggs, S.L.P.; Bird, E.J.; Molik, D.C.; Nayduch, D. Vesicular Stomatitis Virus Elicits Early Transcriptome Response in Culicoides sonorensis Cells. Viruses 2023, 15, 2108. https://doi.org/10.3390/v15102108
Scroggs SLP, Bird EJ, Molik DC, Nayduch D. Vesicular Stomatitis Virus Elicits Early Transcriptome Response in Culicoides sonorensis Cells. Viruses. 2023; 15(10):2108. https://doi.org/10.3390/v15102108
Chicago/Turabian StyleScroggs, Stacey L. P., Edward J. Bird, David C. Molik, and Dana Nayduch. 2023. "Vesicular Stomatitis Virus Elicits Early Transcriptome Response in Culicoides sonorensis Cells" Viruses 15, no. 10: 2108. https://doi.org/10.3390/v15102108
APA StyleScroggs, S. L. P., Bird, E. J., Molik, D. C., & Nayduch, D. (2023). Vesicular Stomatitis Virus Elicits Early Transcriptome Response in Culicoides sonorensis Cells. Viruses, 15(10), 2108. https://doi.org/10.3390/v15102108