
Short-Term Effects of Droughts and Cold Winters on the Growth of Scots Pine at Coastal Sand Dunes around the South Baltic Sea


 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
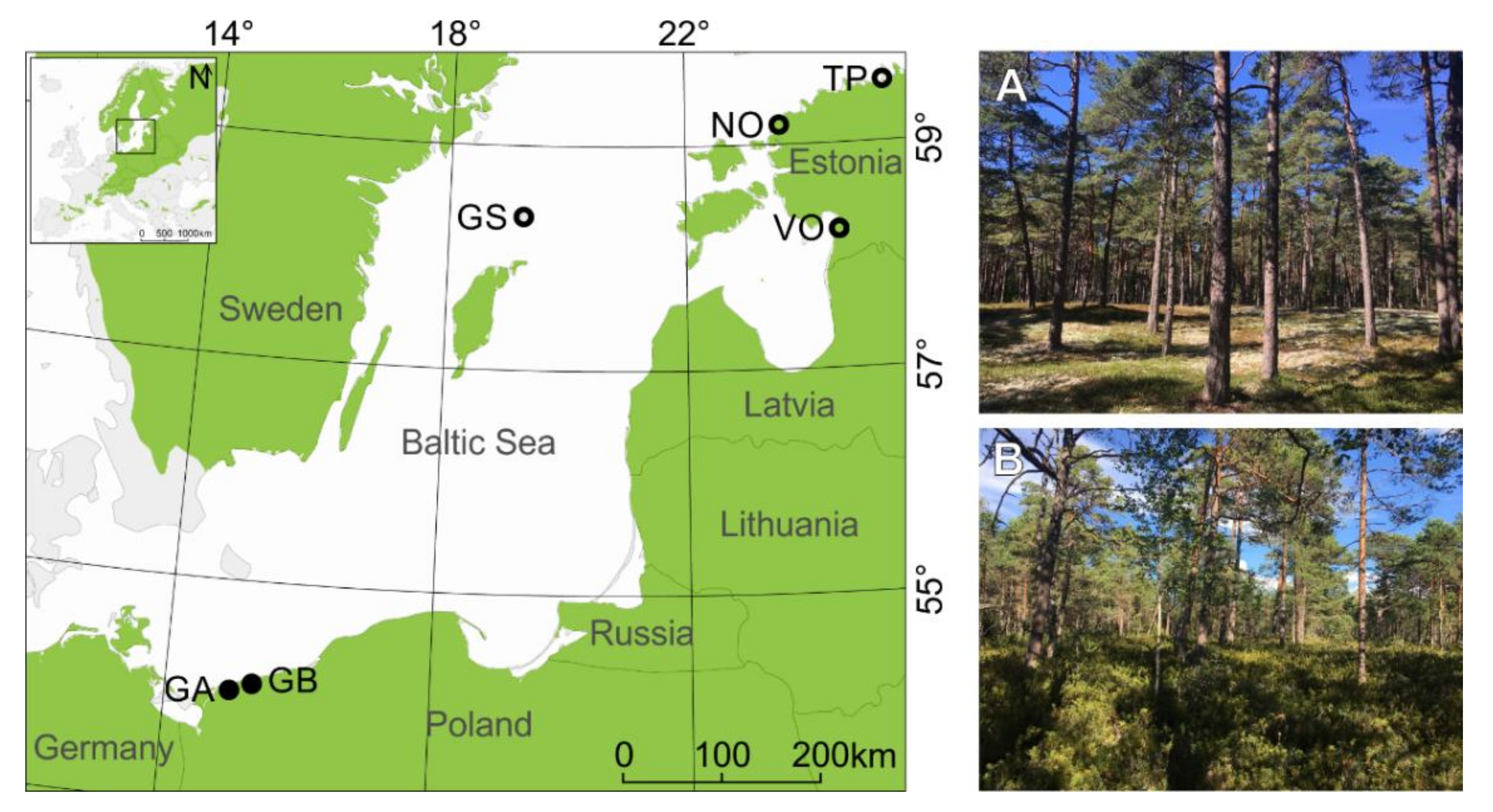
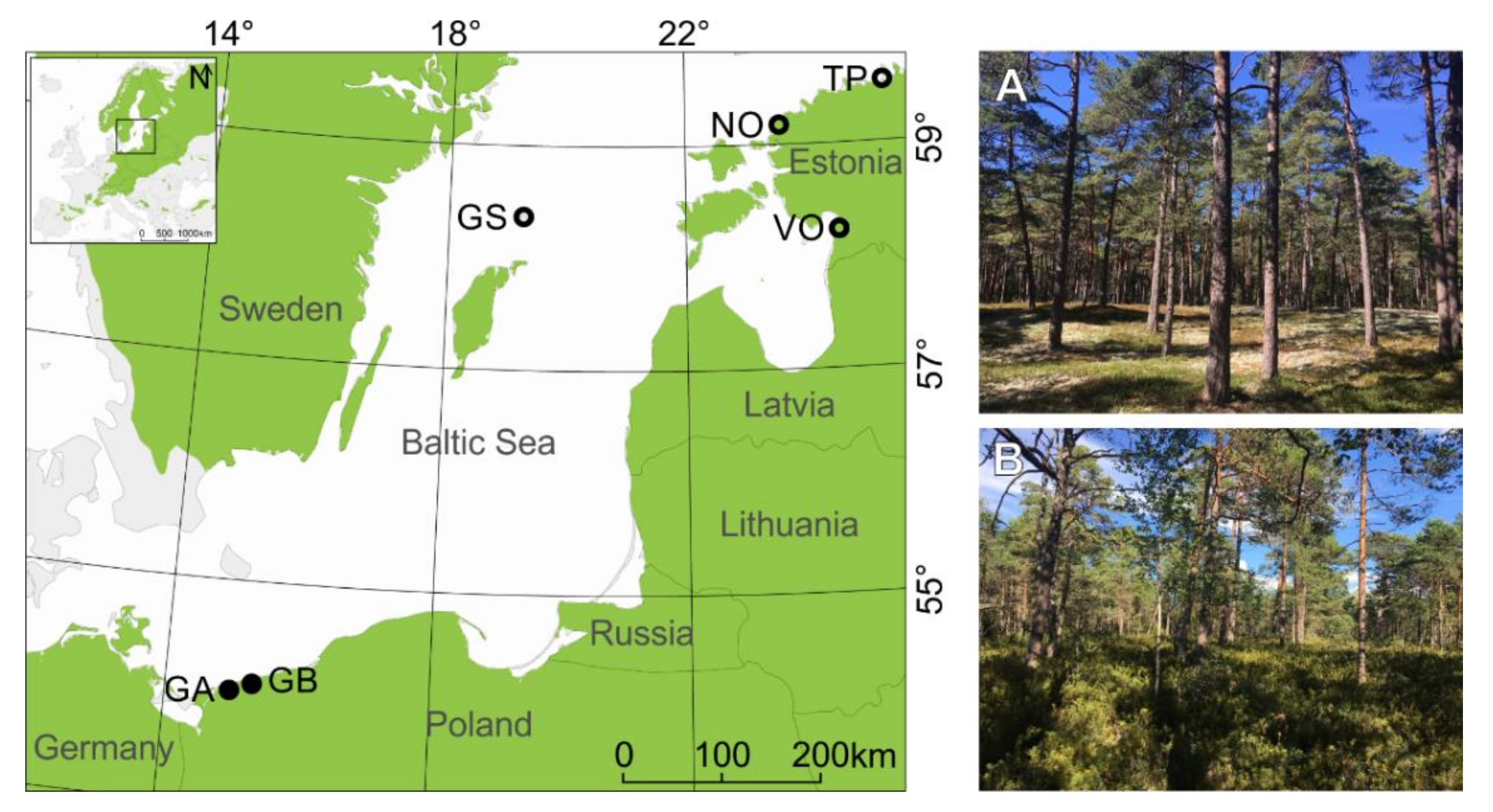
2.1. Research Area and Sampling
2.2. Sample Preparation and Tree-Ring Measurements
2.3. Cross-Dating and Chronology Building
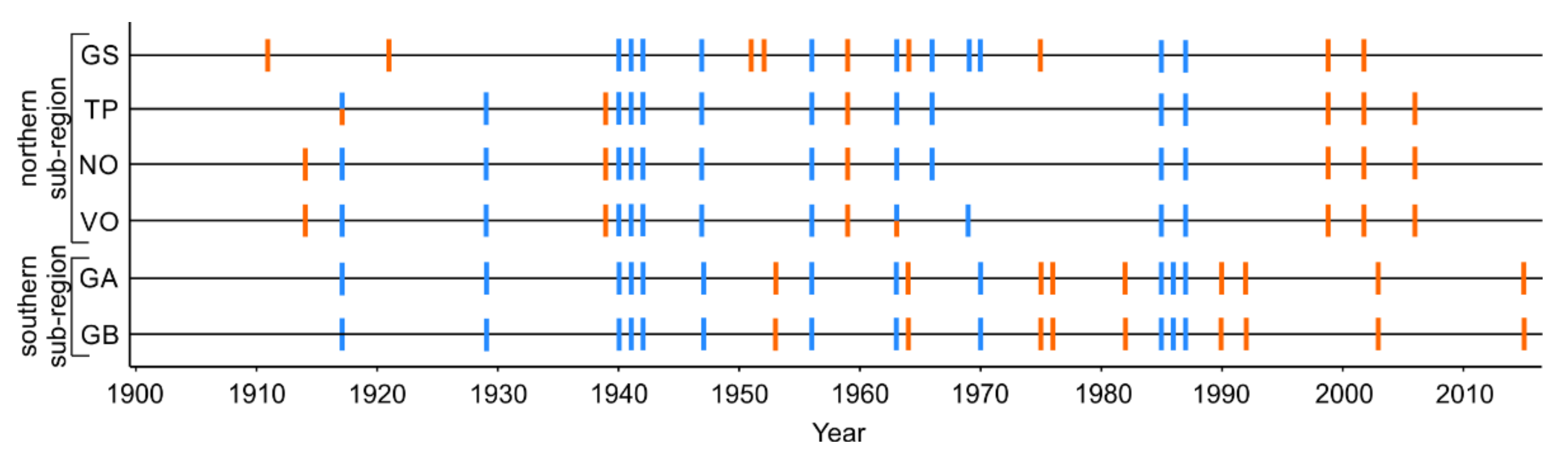
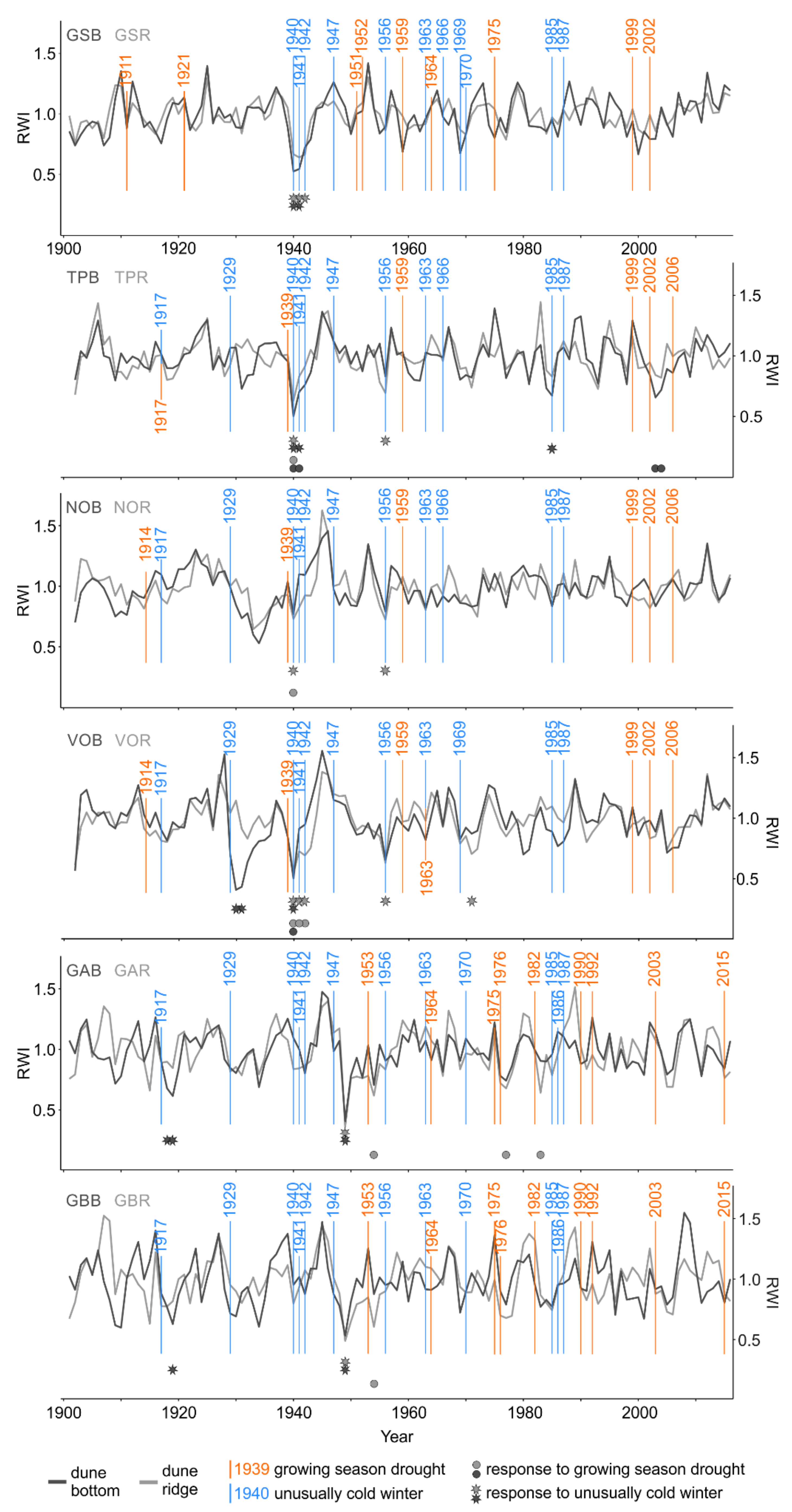
2.4. Identification of Growing Season Droughts and Unusually Cold Winters
2.5. Assessing Scots Pine Responses to Growing Season Droughts and Unusually Cold Winters
- 1.
- At the subregional scale, where we divided microsite-specific chronologies into two subregions (based on [55]): northern and southern. Both subregions were represented by dune ridge and dune bottom microsites. In turn, for each tree-ring parameter, we obtained four datasets of microsite-specific subregional chronologies:
- Dune ridge from the northern subregion (GSR, TPR, NOR, VOR; hereafter called ridge-north);
- Dune bottom from the northern subregion (GSB, TPB, NOB, VOB; hereafter called bottom-north);
- Dune ridge from the southern subregion (GAR, GBR; hereafter called ridge-south);
- Dune bottom from the southern subregion (GAB, GBB; hereafter called bottom-south).
- 2.
- At a local scale, where the SEA was calculated for each single microsite-specific RW and LBI chronologies. Again, three years before and three years after the climate event were considered. In the case of the local-scale SEA, we did not superpose individual years of extreme events but rather analyzed the deviation of growth in each extreme year from the mean of consecutive and following years. By adopting this approach, we highlighted the intensity of each extreme event and tested for its significance.
3. Results
3.1. Extreme Climate Events Dataset and Quality of Tree-Ring Series
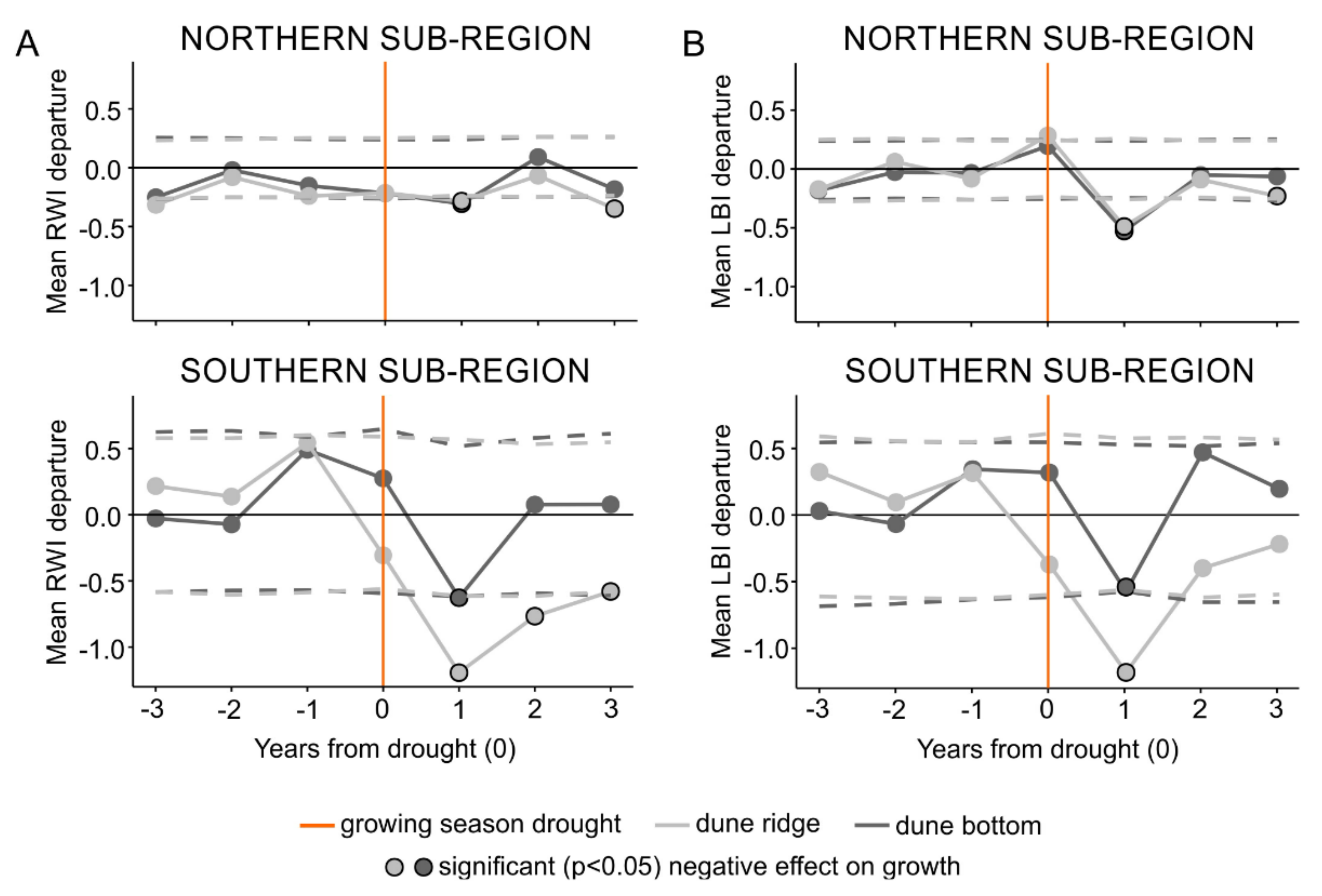
3.2. Ring Width and Latewood Blue Intensity Responses to Growing Season Droughts
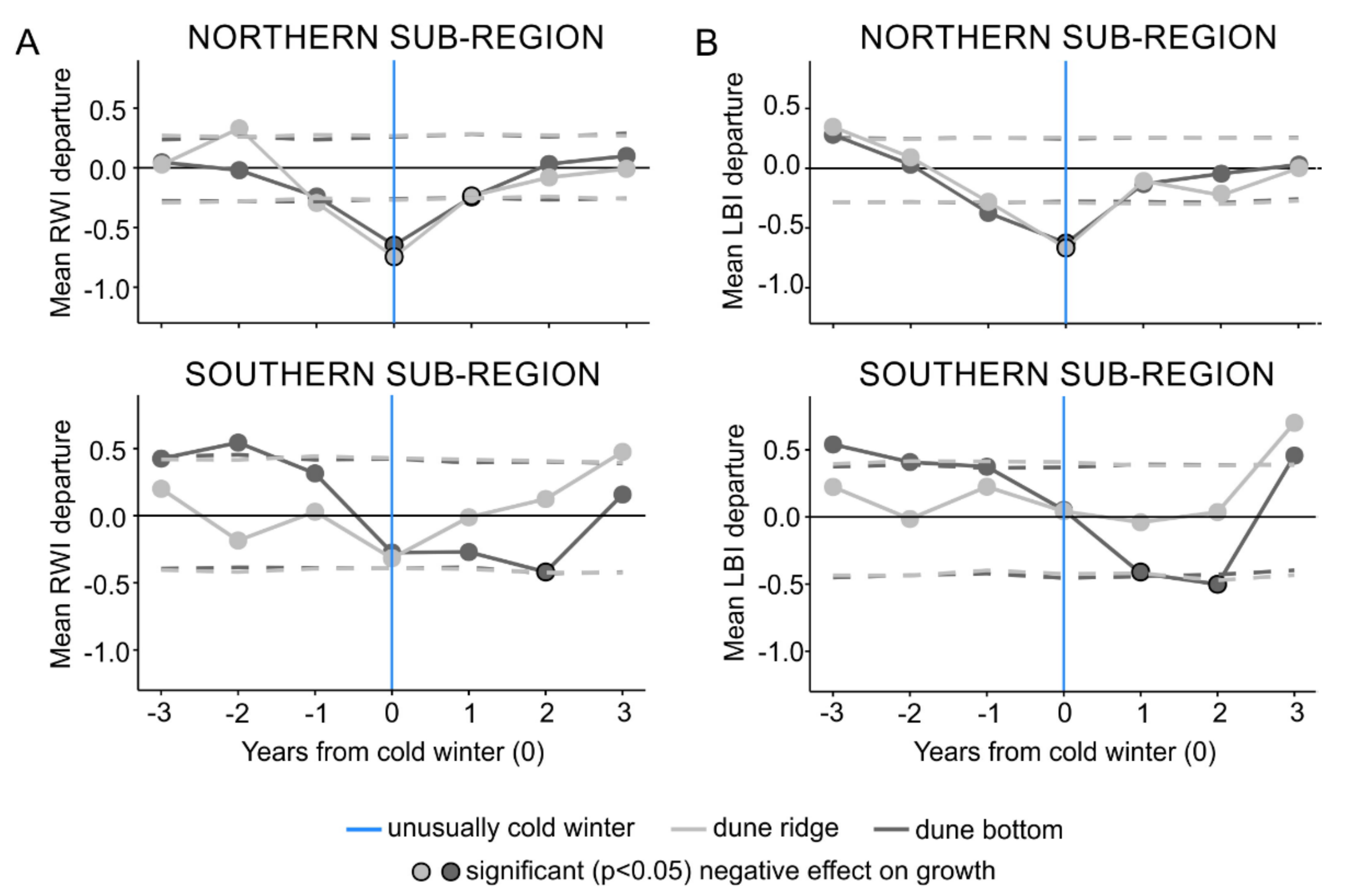
3.3. Ring Width and Latewood Blue Intensity Responses to Unusually Cold Winters
4. Discussion
4.1. Scots Pine Growth Responses to Growing Season Droughts
4.2. Microsite Differences in Scots Pine Growth Responses to Growing Season Droughts
4.3. Scots Pine Growth Responses to Unusually Cold Winters
4.4. Microsite Differences in Scots Pine Growth Responses to Unusually Cold Winters
4.5. Limitations of our Approach
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eilmann, B.; Zweifel, R.; Buchmann, N.; Graf Pannatier, E.; Rigling, A. Drought alters timing, quantity, and quality of wood formation in Scots pine. J. Exp. Bot. 2011, 62, 2763–2771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanoni, M.; Bugmann, H.; Nötzli, M.; Bigler, C. Drought and frost contribute to abrupt growth decreases before tree mortality in nine temperate tree species. For. Ecol. Manag. 2016, 382, 51–63. [Google Scholar] [CrossRef] [Green Version]
- Vanoni, M.; Bugmann, H.; Nötzli, M.; Bigler, C. Quantifying the effects of drought on abrupt growth decreases of major tree species in Switzerland. Ecol. Evol. 2016, 6, 3555–3570. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Sánchez-Salguero, R.; Vicente-Serrano, S.M.; Serra-Maluquer, X.; Gutiérrez, E.; De Luis, M.; Sangüesa-Barreda, G.; Novak, K.; Rozas, V.; et al. Drought legacies are short, prevail in dry conifer forests and depend on growth variability. J. Ecol. 2020, 108, 2473–2484. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Sangüesa-Barreda, G.; Serra-Maluquer, X.; Sánchez-Salguero, R.; Coll, L.; Casals, P. Tree Species Are Differently Impacted by Cumulative Drought Stress and Present Higher Growth Synchrony in Dry Places. Front. For. Glob. Chang. 2020, 3. Available online: https://www.frontiersin.org/articles/10.3389/ffgc.2020.573346/full (accessed on 29 January 2022). [CrossRef]
- Vitas, A. Sensitivity of Scots Pine Trees to Winter Colds and Summer droughts. Balt. For. 2006, 12, 220–226. [Google Scholar]
- Jansons, Ā.; Matisons, R.; Šēnhofa, S.; Katrevičs, J.; Jansons, J. High-frequency variation of tree-ring width of some native and alien tree species in Latvia during the period 1965–2009. Dendrochronologia 2016, 40, 151–158. [Google Scholar] [CrossRef]
- Misi, D.; Nafradi, K. Late Winter—Early Spring Thermal Conditions and their long-term effect on tree-ring growth in Hungary. Balt. For. 2013, 22, 203–211. [Google Scholar]
- IPCC SYR TSU. Climate Change 2014: Synthesis Report; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2015. [Google Scholar]
- Anderegg, W.R.L.; Schwalm, C.R.; Biondi, F.; Camarero, J.J.; Koch, G.W.; Litvak, M.; Ogle, K.; Shaw, J.D.; Shevliakova, E.; Williams, A.P.; et al. Pervasive drought legacies in forest ecosystems and their implications for carbon cycle models. Science 2015, 349, 528–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Büntgen, U.; Urban, O.; Krusic, P.J.; Rybníček, M.; Kolář, T.; Kyncl, T.; Ač, A.; Koňasová, E.; Čáslavský, J.; Esper, J.; et al. Recent European drought extremes beyond Common Era background variability. Nat. Geosci. 2021, 14, 190–196. [Google Scholar] [CrossRef]
- Neuwirth, B.; Esper, J.; Schweingruber, F.H.; Winiger, M. Site ecological differences to the climatic forcing of spruce pointer years from the Lötschental, Switzerland. Dendrochronologia 2004, 21, 69–78. [Google Scholar] [CrossRef]
- Reichstein, M.; Ciais, P.; Papale, D.; Valentini, R.; Running, S.; Viovy, N.; Cramer, W.; Granier, A.; Ogée, J.; Allard, V.; et al. Reduction of ecosystem productivity and respiration during the European summer 2003 climate anomaly: A joint flux tower, remote sensing and modelling analysis. Glob. Chang. Biol. 2007, 13, 634–651. [Google Scholar] [CrossRef]
- Vitasse, Y.; Bottero, A.; Cailleret, M.; Bigler, C.; Fonti, P.; Gessler, A.; Lévesque, M.; Rohner, B.; Weber, P.; Rigling, A.; et al. Contrasting resistance and resilience to extreme drought and late spring frost in five major European tree species. Glob. Chang. Biol. 2019, 25, 3781–3792. [Google Scholar] [CrossRef] [PubMed]
- Gazol, A.; Camarero, J.J.; Vicente-Serrano, S.M.; Sánchez-Salguero, R.; Gutiérrez, E.; de Luis, M.; Sangüesa-Barreda, G.; Novak, K.; Rozas, V.; Tíscar, P.A.; et al. Forest resilience to drought varies across biomes. Glob. Chang. Biol. 2018, 24, 2143–2158. [Google Scholar] [CrossRef]
- Kunstler, G.; Guyennon, A.; Ratcliffe, S.; Rüger, N.; Ruiz-Benito, P.; Childs, D.Z.; Dahlgren, J.; Lehtonen, A.; Thuiller, W.; Wirth, C.; et al. Demographic performance of European tree species at their hot and cold climatic edges. J. Ecol. 2021, 109, 1041–1054. [Google Scholar] [CrossRef]
- Song, L.; Zhu, J.; Zhang, J.; Zhang, T.; Wang, K.; Wang, G.; Liu, J. Effect of Drought and Topographic Position on Depth of Soil Water Extraction of Pinus sylvestris L. var. mongolica Litv. Trees in a Semiarid Sandy Region, Northeast China. Forests 2019, 10, 370. [Google Scholar] [CrossRef] [Green Version]
- Peltier, D.M.P.; Ogle, K. Legacies of more frequent drought in ponderosa pine across the western United States. Glob. Chang. Biol. 2019, 25, 3803–3816. [Google Scholar] [CrossRef]
- Eilmann, B.; Rigling, A. Tree-growth analyses to estimate tree species’ drought tolerance. Tree Physiol. 2012, 32, 178–187. [Google Scholar] [CrossRef] [Green Version]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Cantero, A.; Sánchez-Salguero, R.; Sánchez-Miranda, A.; Granda, E.; Serra-Maluquer, X.; Ibáñez, R. Forest Growth Responses to Drought at Short- and Long-Term Scales in Spain: Squeezing the Stress Memory from Tree Rings. Front. Ecol. Evol. 2018, 6. Available online: https://www.frontiersin.org/articles/10.3389/fevo.2018.00009/full (accessed on 29 January 2022). [CrossRef] [Green Version]
- Sánchez-Salguero, R.; Navarro-Cerrillo, R.M.; Camarero, J.J.; Fernández-Cancio, A. Selective drought-induced decline of pine species in southeastern Spain. Clim. Chang. 2012, 113, 767–785. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J. Functional diversity enhances silver fir growth resilience to an extreme drought. J. Ecol. 2016, 104, 1063–1075. [Google Scholar] [CrossRef]
- Senf, C.; Buras, A.; Zang, C.S.; Rammig, A.; Seidl, R. Excess forest mortality is consistently linked to drought across Europe. Nat. Commun. 2020, 11, 6200. [Google Scholar] [CrossRef]
- Scharnweber, T.; Smiljanic, M.; Cruz-García, R.; Manthey, M.; Wilmking, M. Tree growth at the end of the 21st century—The extreme years 2018/19 as template for future growth conditions. Environ. Res. Lett. 2020, 15, 074022. [Google Scholar] [CrossRef] [Green Version]
- Harvey, J.E.; Smiljanić, M.; Scharnweber, T.; Buras, A.; Cedro, A.; Cruz-García, R.; Drobyshev, I.; Janecka, K.; Jansons, Ā.; Kaczka, R.; et al. Tree growth influenced by warming winter climate and summer moisture availability in northern temperate forests. Glob. Chang. Biol. 2019, 26, 2505–2518. [Google Scholar] [CrossRef] [PubMed]
- Koprowski, M. Spatial distribution of introduced Norway spruce growth in lowland Poland: The influence of changing climate and extreme weather events. Quat. Int. 2013, 283, 139–146. [Google Scholar] [CrossRef]
- Galvez, D.A.; Landhäusser, S.M.; Tyree, M.T. Low root reserve accumulation during drought may lead to winter mortality in poplar seedlings. New Phytol. 2013, 198, 139–148. [Google Scholar] [CrossRef]
- Durrant, T.H.; De Rigo, D.; Caudullo, G. Pinus sylvestris in Europe—Distribution, habitat, usage and threats. Eur. Atlas For. Tree Species 2016, 132–133. Available online: https://forest.jrc.ec.europa.eu/media/atlas/Pinus_sylvestris.pdf (accessed on 29 January 2022).
- Pâques, L.E. Forest Tree Breeding in Europe; Springer: Dordrecht, The Netherlands, 2013. [Google Scholar]
- Klõšeiko, J.; Tilk, M. Drought tolerance of Scots pine in diverse growth conditions on a dune estimated on the basis of carbohydrates and chlorophyll fluorescence in needles. For. Stud. 2008, 49, 25–36. [Google Scholar] [CrossRef]
- Candel-Pérez, D.; Lo, Y.-H.; Blanco, J.A.; Chiu, C.-M.; Camarero, J.J.; De Andrés, E.G.; Imbert, J.B.; Castillo, F.J. Drought-Induced Changes in Wood Density Are Not Prevented by Thinning in Scots Pine Stands. Forests 2018, 9, 4. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Vilalta, J.; Pinol, J. Drought-induced mortality and hydraulic architecture in pine populations of the NE Iberian Peninsula. For. Ecol. Manag. 2002, 161, 247–256. [Google Scholar] [CrossRef]
- Martin-Benito, D.; Beeckman, H.; Cañellas, I. Influence of drought on tree rings and tracheid features of Pinus nigra and Pinus sylvestris in a mesic Mediterranean forest. Eur. J. For. Res. 2013, 132, 33–45. [Google Scholar] [CrossRef]
- Rigling, A.; Bigler, C.; Eilmann, B.; Feldmeyer-Christe, E.; Gimmi, U.; Ginzler, C.; Graf, U.; Mayer, P.; Vacchiano, G.; Weber, P.; et al. Driving factors of a vegetation shift from Scots pine to pubescent oak in dry Alpine forests. Glob. Chang. Biol. 2013, 19, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Salguero, R.; Camarero, J.J.; Rozas, V.; Génova, M.; Olano, J.M.; Arzac, A.; Gazol, A.; Caminero, L.; Tejedor, E.; de Luis, M.; et al. Resist, recover or both? Growth plasticity in response to drought is geographically structured and linked to intraspecific variability in Pinus pinaster. J. Biogeogr. 2018, 45, 1126–1139. [Google Scholar] [CrossRef]
- Taeger, S.; Zang, C.; Liesebach, M.; Schneck, V.; Menzel, A. Impact of climate and drought events on the growth of Scots pine (Pinus sylvestris L.) provenances. For. Ecol. Manag. 2013, 307, 30–42. [Google Scholar] [CrossRef]
- Merlin, M.; Perot, T.; Perret, S.; Korboulewsky, N.; Vallet, P. Effects of stand composition and tree size on resistance and resilience to drought in sessile oak and Scots pine. For. Ecol. Manag. 2015, 339, 22–33. [Google Scholar] [CrossRef] [Green Version]
- Matisons, R.; Jansone, D.; Elferts, D.; Adamovičs, A.; Schneck, V.; Jansons, A. Plasticity of response of tree-ring width of Scots pine provenances to weather extremes in Latvia. Dendrochronologia 2019, 54, 1–10. [Google Scholar] [CrossRef]
- Aldea, J.; Ruiz-Peinado, R.; del Río, M.; Pretzsch, H.; Heym, M.; Brazaitis, G.; Jansons, A.; Metslaid, M.; Barbeito, I.; Bielak, K.; et al. Species stratification and weather conditions drive tree growth in Scots pine and Norway spruce mixed stands along Europe. For. Ecol. Manag. 2021, 481, 118697. [Google Scholar] [CrossRef]
- Stolz, J.; van der Maaten, E.; Kalanke, H.; Martin, J.; Wilmking, M.; van der Maaten-Theunissen, M. Increasing climate sensitivity of beech and pine is not mediated by adaptation and soil characteristics along a precipitation gradient in northeastern Germany. Dendrochronologia 2021, 67, 125834. [Google Scholar] [CrossRef]
- Tuovinen, M.; Jalkanen, R.; McCarroll, D. The effect of severe ground frost on Scots pine (Pinus sylvestris) trees in northern Finland and implications for palaeoclimate reconstructionand implications for paleoclimate reconstruction. Fennia 2005, 183, 109–120. [Google Scholar]
- Zunde, M.; Briede, A.; Elferts, D. Influence of Climatic Factors on the Annual Radial Growth of Scots Pine (Pinus sylvestris L.) in Western Latvia. Proc. Latv. Acad. Sci. Sect. B. Nat. Exact Appl. Sci. 2008, 62, 120–128. [Google Scholar] [CrossRef] [Green Version]
- Martínez, M.L.; Psuty, N.P. Coastal Dunes: Ecology and Conservation; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2008. [Google Scholar]
- Ratas, U.; Rivis, R.; Käärt, K. Changes of coastal dune landscapes in Estonia. For. Stud. 2008, 49, 59–70. [Google Scholar] [CrossRef]
- Łabuz, T.A. Polish coastal dunes—Affecting factors and morphology. Landf. Anal. 2013, 22, 33–59. [Google Scholar] [CrossRef]
- Rutgersson, A.; Jaagus, J.; Schenk, F.; Stendel, M. Observed changes and variability of atmospheric parameters in the Baltic Sea region during the last 200 years. Clim. Res. 2014, 61, 177–190. [Google Scholar] [CrossRef] [Green Version]
- Rimkus, E.; Stonevicius, E.; Kilpys, J.; Maciulyte, V.; Valiukas, D. Drought identification in the eastern Baltic region using NDVI. Earth Syst. Dyn. 2017, 8, 627–637. [Google Scholar] [CrossRef] [Green Version]
- Mandre, M.; Kõresaar, K. Mineral nutrition of natural regeneration of Scots pine on coastal dunes in South-West Estonia. Estonian J. Ecol. 2008, 57, 70. [Google Scholar] [CrossRef] [Green Version]
- Mandre, M.; Lukjanova, A.; Pärn, H.; Kõresaar, K. State of Scots pine (Pinus sylvestris L.) under nutrient and water deficit on coastal dunes of the Baltic Sea. Trees 2010, 24, 1073–1085. [Google Scholar] [CrossRef]
- Maun, M.A. The Biology of Coastal Sand Dunes; Oxford University Press: Oxford, UK; New York, NY, USA, 2009. [Google Scholar]
- Örd, A. Edela-Eesti luitemetsade mullastik. Metsanduslikud Uurim. 1972, 9, 207–221. [Google Scholar]
- Repo, T.; Zhang, G.; Ryyppö, A.; Rikala, R.; Vuorinen, M. The relation between growth cessation and frost hardening in Scots pines of different origins. Trees 2000, 14, 456–464. [Google Scholar] [CrossRef]
- Beck, E.H.; Heim, R.; Hansen, J. Plant resistance to cold stress: Mechanisms and environmental signals triggering frost hardening and dehardening. J. Biosci. 2004, 29, 449–459. [Google Scholar] [CrossRef]
- Avotniece, Z.; Klavins, M.; Rodinovs, V. Changes of Extreme Climate Events in Latvia. Sci. J. Riga Tech. Univ. Environ. Clim. Technol. 2012, 9, 4–11. [Google Scholar] [CrossRef] [Green Version]
- Janecka, K.; Harvey, J.E.; Trouillier, M.; Kaczka, R.J.; Metslaid, S.; Metslaid, M.; Buras, A.; Wilmking, M. Higher Winter-Spring Temperature and Winter-Spring/Summer Moisture Availability Increase Scots Pine Growth on Coastal Dune Microsites Around the South Baltic Sea. Front. For. Glob. Chang. 2020, 3. Available online: https://www.frontiersin.org/articles/10.3389/ffgc.2020.578912/full (accessed on 29 January 2022). [CrossRef]
- Janecka, K.; Kaczka, R.J.; Gärtner, H.; E Harvey, J.; Treydte, K. Compression wood has a minor effect on the climate signal in tree-ring stable isotope records of montane Norway spruce. Tree Physiol. 2020, 40, 1014–1028. [Google Scholar] [CrossRef] [PubMed]
- Speer, J. Fundamentals of Tree-Ring Research; University of Arizona Press: Tucson, AZ, USA, 2010. [Google Scholar]
- Wilson, R.; Rao, R.; Rydval, M.; Wood, C.; Larsson, L.; Luckman, B.H. Blue Intensity for dendroclimatology: The BC blues: A case study from British Columbia, Canada. Holocene 2014, 24, 1428–1438. [Google Scholar] [CrossRef]
- Rydval, M.; Larsson, L.; McGlynn, L.; Gunnarson, B.E.; Loader, N.; Young, G.H.; Wilson, R. Blue intensity for dendroclimatology: Should we have the blues? Experiments from Scotland. Dendrochronologia 2014, 32, 191–204. [Google Scholar] [CrossRef]
- Xu, K.; Wang, X.; Liang, P.; An, H.; Sun, H.; Han, W.; Li, Q. Tree-ring widths are good proxies of annual variation in forest productivity in temperate forests. Sci. Rep. 2017, 7, 1945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaczka, R.J.; Wilson, R. I-BIND: International Blue intensity network development working group. Dendrochronologia 2021, 68, 125859. [Google Scholar] [CrossRef]
- Kaczka, R.J.; Spyt, B.; Janecka, K.; Beil, I.; Büntgen, U.; Scharnweber, T.; Nievergelt, D.; Wilmking, M. Different maximum latewood density and blue intensity measurements techniques reveal similar results. Dendrochronologia 2018, 49, 94–101. [Google Scholar] [CrossRef]
- Larsson, L. CDendro: Cybis Dendro Dating Program; Cybis Elektronik & Data AB: Saltsjöbaden, Sweden, 2003. [Google Scholar]
- Holmes, R.L. Quality control of crossdating and measuring. Users manual for computer program COFECHA. In Tree-Ring Chronologies of Western North America: California, Eastern Oregon and Northern Great Basin with Procedures Used in the Chronology Development Work Including Users Manuals for Computer Programs COFECHA and ARSTAN; Laboratory of Tree-Ring Research, University of Arizona: Tucson, AZ, USA, 1986. [Google Scholar]
- Bunn, A. A dendrochronology program library in R (dplR). Dendrochronologia 2008, 26, 115–124. [Google Scholar] [CrossRef]
- Cook, E.R.; Peters, K. Calculating unbiased tree-ring indices for the study of climatic and environmental change. Holocene 1997, 7, 361–370. [Google Scholar] [CrossRef]
- Helama, S.; Lindholm, M.; Timonen, M.; Eronen, M. Detection of climate signal in dendrochronological data analysis: A comparison of tree-ring standardization methods. Theor. Appl. Climatol. 2004, 79, 239–254. [Google Scholar] [CrossRef]
- Cook, E.R.; Briffa, K.; Shiyatov, S.; Mazepa, V. Tree-ring standardization and growth trend estimation. In Methods of Dendrochronology: Applications in the Environmental Sciences; Kluwer: Dordrecht, The Netherlands, 1990; pp. 104–123. [Google Scholar]
- Wigley, T.M.L.; Briffa, K.R.; Jones, P.D. On the Average Value of Correlated Time Series, with Applications in Dendroclimatology and Hydrometeorology. J. Appl. Meteorol. Climatol. 1984, 23, 201–213. [Google Scholar] [CrossRef]
- Buras, A. A comment on the expressed population signal. Dendrochronologia 2017, 44, 130–132. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A Multiscalar Drought Index Sensitive to Global Warming: The Standardized Precipitation Evapotranspiration Index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef] [Green Version]
- Tammets, T.; Jaagus, J. Climatology of precipitation extremes in Estonia using the method of moving precipitation totals. Arch. Meteorol. Geophys. Bioclimatol. Ser. B 2013, 111, 623–639. [Google Scholar] [CrossRef]
- Li, B.; Zhou, W.; Zhao, Y.; Ju, Q.; Yu, Z.; Liang, Z.; Acharya, K. Using the SPEI to Assess Recent Climate Change in the Yarlung Zangbo River Basin, South Tibet. Water 2015, 7, 5474–5486. [Google Scholar] [CrossRef] [Green Version]
- Orwig, D.A.; Abrams, M.D. Variation in radial growth responses to drought among species, site, and canopy strata. Trees 1997, 11, 474–484. [Google Scholar] [CrossRef]
- Pasho, E.; Camarero, J.J.; de Luis, M.; Vicente-Serrano, S.M. Impacts of drought at different time scales on forest growth across a wide climatic gradient in north-eastern Spain. Agric. For. Meteorol. 2011, 151, 1800–1811. [Google Scholar] [CrossRef]
- Lévesque, M.; Saurer, M.; Siegwolf, R.T.W.; Eilmann, B.; Brang, P.; Bugmann, H.; Rigling, A. Drought response of five conifer species under contrasting water availability suggests high vulnerability of Norway spruce and European larch. Glob. Chang. Biol. 2013, 19, 3184–3199. [Google Scholar] [CrossRef]
- Eilmann, B.; Zweifel, R.; Buchmann, N.; Fonti, P.; Rigling, A. Drought-induced adaptation of the xylem in Scots pine and pubescent oak. Tree Physiol. 2009, 29, 1011–1020. [Google Scholar] [CrossRef] [Green Version]
- Hacke, U.G.; Sperry, J.S.; Pockman, W.T.; Davis, S.D.; McCulloh, K.A. Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure. Oecologia 2001, 126, 457–461. [Google Scholar] [CrossRef]
- Pittermann, J.; Sperry, J.S.; Wheeler, J.K.; Hacke, U.G.; Sikkema, E.H. Mechanical reinforcement of tracheids compromises the hydraulic efficiency of conifer xylem. Plant Cell Environ. 2006, 29, 1618–1628. [Google Scholar] [CrossRef] [Green Version]
- Tyree, M.T.; Zimmermann, M.T. Xylem Structure and the Ascent of Sap; Springer: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- Högberg, P.; Nordgren, A.; Buchmann, N.; Taylor, A.F.S.; Ekblad, A.; Högberg, M.N.; Nyberg, G.; Ottosson-Löfvenius, M.; Read, D.J. Large-scale forest girdling shows that current photosynthesis drives soil respiration. Nature 2001, 411, 789–792. [Google Scholar] [CrossRef] [PubMed]
- Allen, S.T.; Kirchner, J.W.; Braun, S.; Siegwolf, R.T.W.; Goldsmith, G.R. Seasonal origins of soil water used by trees. Hydrol. Earth Syst. Sci. 2019, 23, 1199–1210. [Google Scholar] [CrossRef] [Green Version]
- Mandre, M. Conditions for mineral nutrition and content of nutrients in scots pine on dunes in Southwest Estonia. For. Ecosyst. Coast. Dunes Southwest Est. 2003, 39, 32–42. [Google Scholar]
- Abrams, M.D.; Ruffner, C.M.; Morgan, T.A. Tree-Ring Responses to Drought Across Species and Contrasting Sites in the Ridge and Valley of Central Pennsylvania. For. Sci. 1998, 44, 550–558. [Google Scholar]
- Vicente-Serrano, S.M.; Lasanta, T.; Gracia, C. Aridification determines changes in forest growth in Pinus halepensis forests under semiarid Mediterranean climate conditions. Agric. For. Meteorol. 2010, 150, 614–628. [Google Scholar] [CrossRef]
- Rigling, A.; Brühlhart, H.; Bräker, O.U.; Forster, T.; Schweingruber, F.H. Effects of irrigation on diameter growth and vertical resin duct production in Pinus sylvestris L. on dry sites in the central Alps, Switzerland. For. Ecol. Manag. 2003, 175, 285–296. [Google Scholar] [CrossRef]
- Tumajer, J.; Treml, V. Response of floodplain pedunculate oak (Quercus robur L.) tree-ring width and vessel anatomy to climatic trends and extreme hydroclimatic events. For. Ecol. Manag. 2016, 379, 185–194. [Google Scholar] [CrossRef]
- Fritts, H. Tree Rings and Climate; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Tyree, M.T.; Sperry, J.S. Vulnerability of Xylem to Cavitation and Embolism. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 1989, 40, 19–36. [Google Scholar] [CrossRef]
- Perry, T.O. Dormancy of trees in winter. Science 1971, 171, 29–36. [Google Scholar] [CrossRef]
- Kozlowski, T.T.; Kramer, P.J.; Pallardy, S.G. The Physiological Ecology of Woody Plants; Academic Press: Cambridge, MS, USA, 2012. [Google Scholar]
- Pederson, N.; Cook, E.R.; Jacoby, G.C.; Peteet, D.M.; Griffin, K.L. The influence of winter temperatures on the annual radial growth of six northern range margin tree species. Dendrochronologia 2004, 22, 7–29. [Google Scholar] [CrossRef]
- Sperry, J.S.; Sullivan, J.E.M. Xylem Embolism in Response to Freeze-Thaw Cycles and Water Stress in Ring-Porous, Diffuse-Porous, and Conifer Species. Plant Physiol. 1992, 100, 605–613. [Google Scholar] [CrossRef] [PubMed]
- Tierney, G.L.; Fahey, T.J.; Groffman, P.; Hardy, J.P.; Fitzhugh, R.D.; Driscoll, C.T. Soil freezing alters fine root dynamics in a northern hardwood forest. Biogeochemistry 2001, 56, 175–190. [Google Scholar] [CrossRef]
- Peterson, D.W.; Ettl, G.J.; Peterson, D.L. Growth responses of subalpine fir to climatic variability in the Pacific Northwest. Can. J. For. Res. 2002, 32, 1503–1517. [Google Scholar] [CrossRef]
- Peterson, D.W.; Peterson, D.L. Mountain hemlock growth responds to climatic variability at annual and decadal time scales. Ecology 2001, 82, 3330–3345. [Google Scholar] [CrossRef]
- Niklasson, M. Skogshistoria och Bränder på Gotska Sandön; Länsstyrelsen: Visby, Sweden, 2015; p. 58. [Google Scholar]
- Brienen, R.J.; Gloor, E.; Zuidema, P.A. Detecting Evidence for CO2 Fertilization from Tree Ring Studies: The Potential Role of Sampling Biases. Glob. Biogeochem. Cycles Int. J. Glob. Change 2012, 26, GB1025. Available online: https://agupubs.onlinelibrary.wiley.com/doi/full/10.1029/2011GB004143 (accessed on 29 January 2022).
- Rao, M.P.; Cook, E.R.; Cook, B.I.; Anchukaitis, K.J.; D’Arrigo, R.D.; Krusic, P.J.; LeGrande, A.N. A double bootstrap approach to Superposed Epoch Analysis to evaluate response uncertainty. Dendrochronologia 2019, 55, 119–124. [Google Scholar] [CrossRef]
- Lévesque, M.; Rigling, A.; Bugmann, H.; Weber, P.; Brang, P. Growth response of five co-occurring conifers to drought across a wide climatic gradient in Central Europe. Agric. For. Meteorol. 2014, 197, 1–12. [Google Scholar] [CrossRef]
- Doody, J.P. Sand Dune Conservation, Management and Restoration; Springer: Dordrecht, The Netherlands, 2013. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subregion | Country | Site | Microsite | Lat | Lon | Number of Analyzed Trees | Tree Height (m) | Organic Layer (cm) | Forest Site Type | |
|---|---|---|---|---|---|---|---|---|---|---|
| RW | LBI | |||||||||
| northern | Sweden | Gotska Sandön (GS) | bottom (GSB) | 58.38° | 19.20° | 43 | 40 | 15.3 ** | 10.2 ** | CA |
| ridge (GSR) | 58.37° | 19.20° | 64 | 52 | 17.2 ** | 5.1 ** | CL | |||
| Estonia | Tapurla (TP) | bottom (TPB) | 59.59° | 25.55° | 50 | 48 | 28.5 *** | 11.9 *** | VU | |
| ridge (TPR) | 59.60° | 25.55° | 47 | 44 | 19.2 *** | 7 *** | CL | |||
| Nõva (NO) | bottom (NOB) | 59.21° | 23.62° | 43 | 39 | 13.3 *** | 11.5 *** | VU | ||
| ridge (NOR) | 59.22° | 23.61° | 50 | 43 | 15.8 *** | 5.5 *** | CL | |||
| Võiste (VO) | bottom (VOB) | 58.20° | 24.49° | 49 | 36 | 23.4 | 27.2 *** | VU | ||
| ridge (VOR) | 58.20° | 24.49° | 50 | 47 | 22.7 | 12.4 *** | CL | |||
| southern | Poland | Gryfice (GA) | bottom (GAB) | 54.11° | 15.20° | 34 | 34 | 14.9 | 11 *** | CA |
| ridge (GAR) | 54.11° | 15.20° | 36 | 33 | 16.3 | 8.6 *** | CL | |||
| Gryfice (GB) | bottom (GBB) | 54.11° | 15.20° | 35 | 34 | 17.6 *** | 11.4 *** | CA | ||
| ridge (GBR) | 54.11° | 15.20° | 35 | 34 | 12.7 *** | 7.3 *** | CL | |||
| total | 536 | 484 | ||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janecka, K.; Metslaid, S.; Metslaid, M.; Harvey, J.E.; Wilmking, M. Short-Term Effects of Droughts and Cold Winters on the Growth of Scots Pine at Coastal Sand Dunes around the South Baltic Sea. Forests 2022, 13, 477. https://doi.org/10.3390/f13030477
Janecka K, Metslaid S, Metslaid M, Harvey JE, Wilmking M. Short-Term Effects of Droughts and Cold Winters on the Growth of Scots Pine at Coastal Sand Dunes around the South Baltic Sea. Forests. 2022; 13(3):477. https://doi.org/10.3390/f13030477
Chicago/Turabian StyleJanecka, Karolina, Sandra Metslaid, Marek Metslaid, Jill E. Harvey, and Martin Wilmking. 2022. "Short-Term Effects of Droughts and Cold Winters on the Growth of Scots Pine at Coastal Sand Dunes around the South Baltic Sea" Forests 13, no. 3: 477. https://doi.org/10.3390/f13030477
APA StyleJanecka, K., Metslaid, S., Metslaid, M., Harvey, J. E., & Wilmking, M. (2022). Short-Term Effects of Droughts and Cold Winters on the Growth of Scots Pine at Coastal Sand Dunes around the South Baltic Sea. Forests, 13(3), 477. https://doi.org/10.3390/f13030477