
Utilization of Aloe Compounds in Combatting Viral Diseases


Abstract
:
1. Introduction
2. Methodology
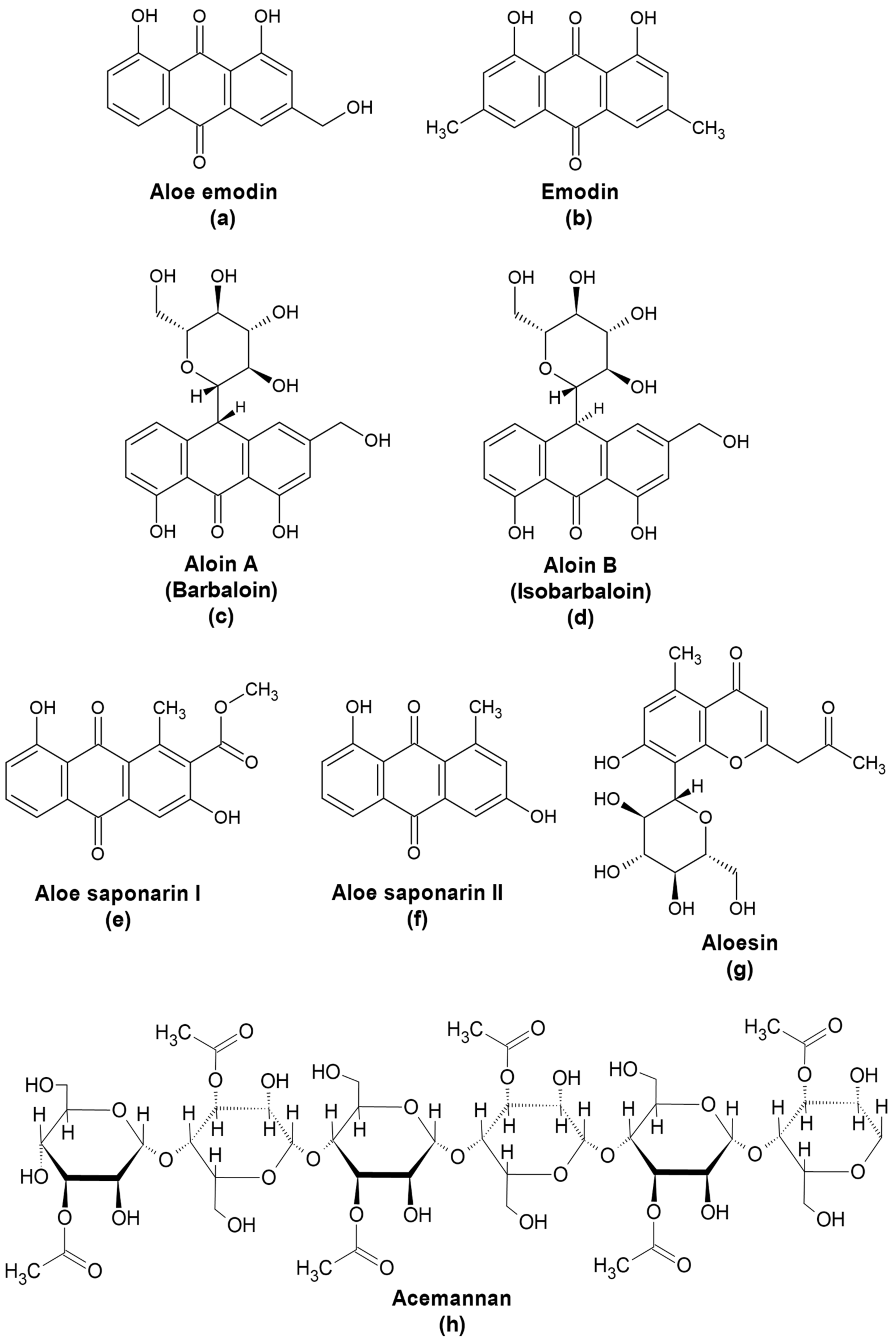
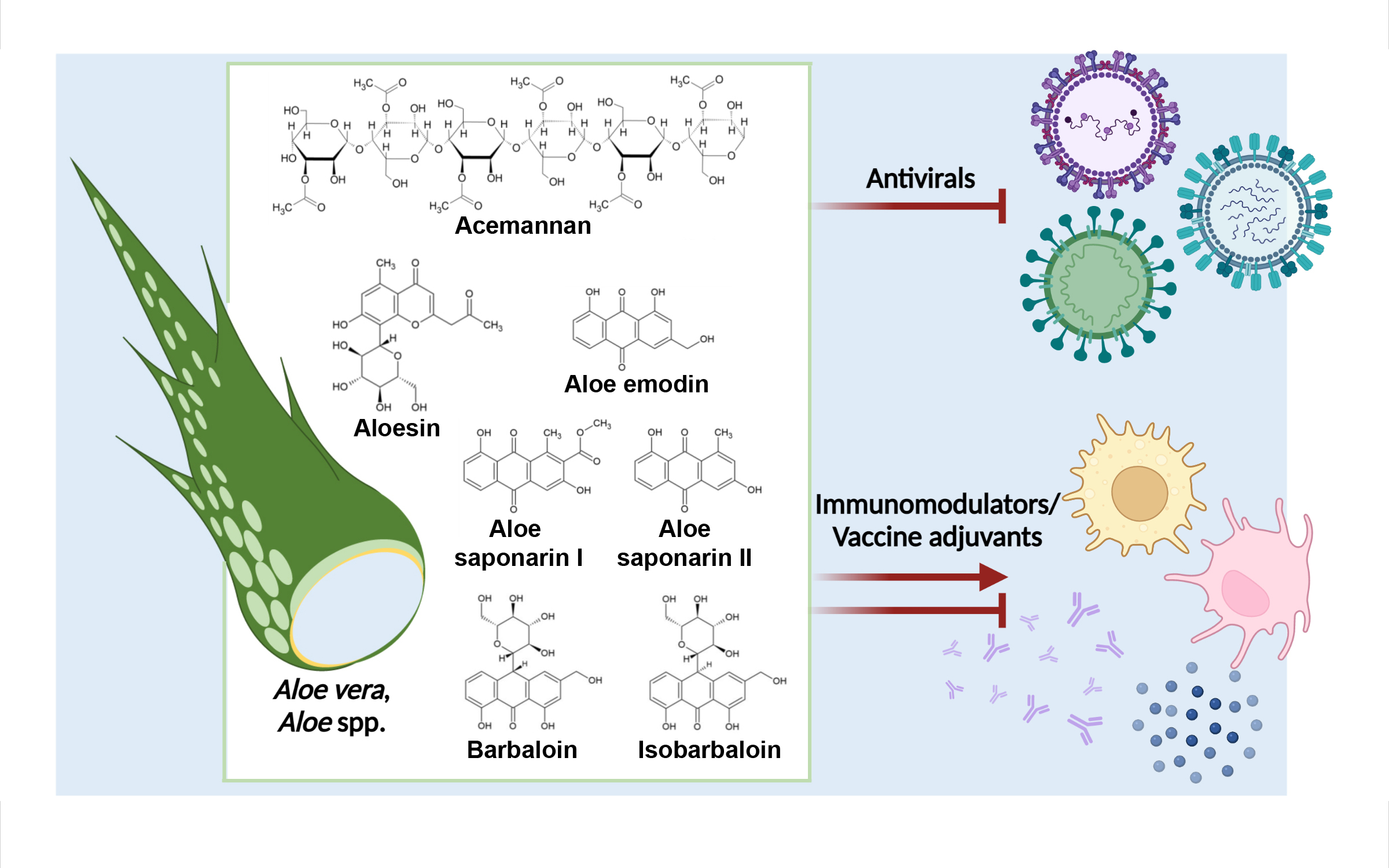
3. Aloe Compounds as Antivirals
3.1. Aloe Extracts Demonstrate Antiviral Activity
3.2. Antiviral Activities of Phenolic Compounds from A. vera
3.2.1. Aloe Emodin
3.2.2. Aloin
3.2.3. Other Phenolic Compounds from Aloe
3.3. Polysaccharides from Aloe
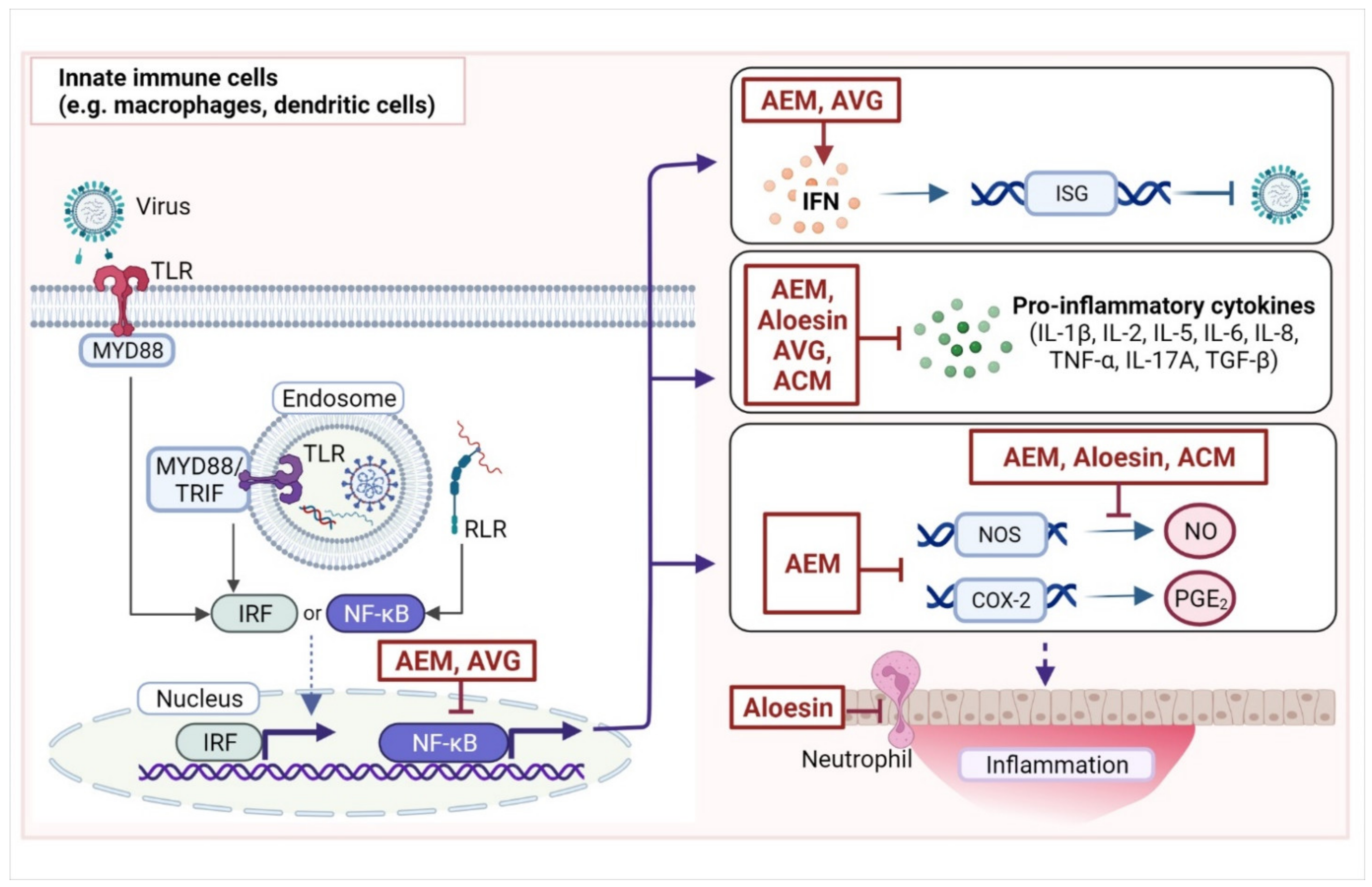
4. Aloe Compounds for the Regulation of Immune Responses to Viral Infection
4.1. Aloe Phenolics as Immunomodulatory Agents
4.1.1. Immunomodulatory Effects of Aloe Emodin
4.1.2. Immunomodulatory Effects of Aloin
4.1.3. Anti-Inflammatory Effects of Aloesin
4.2. Immunomodulatory Effects of A. vera Gel and Its Components
4.2.1. Immunomodulatory Effects of A. vera Gel
4.2.2. Immunomodulatory Effects of Acemannan and A. vera Polysaccharides
4.2.3. Aloe Compounds as Viral Vaccine Adjuvants
5. Hurdles to Applying Aloe Compounds in Preventing and Managing Viral Diseases
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meganck, R.M.; Baric, R.S. Developing therapeutic approaches for twenty-first-century emerging infectious viral diseases. Nat. Med. 2021, 27, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Chitalia, V.C.; Munawar, A.H. A painful lesson from the COVID-19 pandemic: The need for broad-spectrum, host-directed antivirals. J. Transl. Med. 2020, 18, 390. [Google Scholar] [CrossRef] [PubMed]
- Lautié, E.; Russo, O.; Ducrot, P.; Boutin, J.A. Unraveling Plant Natural Chemical Diversity for Drug Discovery Purposes. Front. Pharmacol. 2020, 11, 397. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.I.; Sheikh, W.M.; Rather, M.A.; Venkatesalu, V.; Muzamil Bashir, S.; Nabi, S.U. Medicinal plants: Treasure for antiviral drug discovery. Phytother. Res. 2021, 35, 3447–3483. [Google Scholar] [CrossRef] [PubMed]
- Maan, A.A.; Nazir, A.; Khan, M.K.I.; Ahmad, T.; Zia, R.; Murid, M.; Abrar, M. The therapeutic properties and applications of Aloe vera: A review. J. Herb. Med. 2018, 12, 1–10. [Google Scholar] [CrossRef]
- Surjushe, A.; Vasani, R.; Saple, D.G. Aloe vera: A short review. Indian J. Dermatol. 2008, 53, 163–166. [Google Scholar] [CrossRef]
- Radha, M.H.; Laxmipriya, N.P. Evaluation of biological properties and clinical effectiveness of Aloe vera: A systematic review. J. Tradit. Complement. Med. 2015, 5, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Saoo, K.; Miki, H.; Ohmori, M.; Winters, W.D. Antiviral Activity of Aloe Extracts against Cytomegalovirus. Phytother. Res. 1996, 10, 348–350. [Google Scholar] [CrossRef]
- Sydiskis, R.J.; Owen, D.G.; Lohr, J.L.; Rosler, K.H.; Blomster, R.N. Inactivation of enveloped viruses by anthraquinones extracted from plants. Antimicrob. Agents Chemother. 1991, 35, 2463–2466. [Google Scholar] [CrossRef] [Green Version]
- Zandi, K.; Zadeh, M.; Sartavi, K.; Rastian, Z. Antiviral activity of Aloe vera against herpes simplex virus type 2: An in vitro study. Afr. J. Biotechnol. 2007, 6, 1770–1773. [Google Scholar] [CrossRef]
- Waihenya, R.K.; Mtambo, M.M.A.; Nkwengulila, G. Evaluation of the efficacy of the crude extract of Aloe secundiflora in chickens experimentally infected with Newcastle disease virus. J. Ethnopharmacol. 2002, 79, 299–304. [Google Scholar] [CrossRef]
- Xu, Z.; Liu, Y.; Peng, P.; Liu, Y.; Huang, M.; Ma, Y.; Xue, C.; Cao, Y. Aloe extract inhibits porcine epidemic diarrhea virus in vitro and in vivo. Vet. Microbiol. 2020, 249, 108849. [Google Scholar] [CrossRef] [PubMed]
- Ng, Y.C.; Kim, Y.W.; Ryu, S.; Lee, A.; Lee, J.-S.; Song, M.J. Suppression of norovirus by natural phytochemicals from Aloe vera and Eriobotryae Folium. Food Control 2017, 73, 1362–1370. [Google Scholar] [CrossRef]
- Abd-Alla, H.I.; Abu-Gabal, N.S.; Hassan, A.Z.; El-Safty, M.M.; Shalaby, N.M. Antiviral activity of Aloe hijazensis against some haemagglutinating viruses infection and its phytoconstituents. Arch. Pharmacal Res. 2012, 35, 1347–1354. [Google Scholar] [CrossRef]
- Glatthaar-Saalmüller, B.; Fal, A.M.; Schönknecht, K.; Conrad, F.; Sievers, H.; Saalmüller, A. Antiviral activity of an aqueous extract derived from Aloe arborescens Mill. against a broad panel of viruses causing infections of the upper respiratory tract. Phytomedicine 2015, 22, 911–920. [Google Scholar] [CrossRef]
- Bastian, P.; Fal, A.M.; Jambor, J.; Michalak, A.; Noster, B.; Sievers, H.; Steuber, A.; Walas-Marcinek, N. Candelabra Aloe (Aloe arborescens) in the therapy and prophylaxis of upper respiratory tract infections: Traditional use and recent research results. Wien. Med. Wochenschr. 2013, 163, 73–79. [Google Scholar] [CrossRef]
- Tutin, F.; Clewer, H.W.B. XCIX.—The constituents of rhubarb. J. Chem. Soc. Trans. 1911, 99, 946–967. [Google Scholar] [CrossRef] [Green Version]
- Chihara, T.; Shimpo, K.; Beppu, H.; Yamamoto, N.; Kaneko, T.; Wakamatsu, K.; Sonoda, S. Effects of Aloe-emodin and Emodin on Proliferation of the MKN45 Human Gastric Cancer Cell Line. Asian Pac. J. Cancer Prev. 2015, 16, 3887–3891. [Google Scholar] [CrossRef] [Green Version]
- Andersen, D.O.; Weber, N.D.; Wood, S.G.; Hughes, B.G.; Murray, B.K.; North, J.A. In vitro virucidal activity of selected anthraquinones and anthraquinone derivatives. Antivir. Res. 1991, 16, 185–196. [Google Scholar] [CrossRef]
- Lin, C.-W.; Wu, C.-F.; Hsiao, N.-W.; Chang, C.-Y.; Li, S.-W.; Wan, L.; Lin, Y.-J.; Lin, W.-Y. Aloe-emodin is an interferon-inducing agent with antiviral activity against Japanese encephalitis virus and enterovirus 71. Int. J. Antimicrob. Agents 2008, 32, 355–359. [Google Scholar] [CrossRef]
- Bhimaneni, S.; Kumar, A. Abscisic acid and aloe-emodin against NS2B-NS3A protease of Japanese encephalitis virus. Environ. Sci. Pollut. Res. 2022, 29, 8759–8766. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-W.; Tsai, F.-J.; Tsai, C.-H.; Lai, C.-C.; Wan, L.; Ho, T.-Y.; Hsieh, C.-C.; Chao, P.-D.L. Anti-SARS coronavirus 3C-like protease effects of Isatis indigotica root and plant-derived phenolic compounds. Antivir. Res. 2005, 68, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Parvez, M.K.; Al-Dosari, M.S.; Alam, P.; Rehman, M.; Alajmi, M.F.; Alqahtani, A.S. The anti-hepatitis B virus therapeutic potential of anthraquinones derived from Aloe vera. Phytother. Res. 2019, 33, 2960–2970. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-T.; Hung, C.-Y.; Hseih, Y.-C.; Chang, C.-S.; Velu, A.B.; He, Y.-C.; Huang, Y.-L.; Chen, T.-A.; Chen, T.-C.; Lin, C.-Y.; et al. Effect of aloin on viral neuraminidase and hemagglutinin-specific T cell immunity in acute influenza. Phytomedicine 2019, 64, 152904. [Google Scholar] [CrossRef] [PubMed]
- Alves, D.S.; Pérez-Fons, L.; Estepa, A.; Micol, V. Membrane-related effects underlying the biological activity of the anthraquinones emodin and barbaloin. Biochem. Pharmacol. 2004, 68, 549–561. [Google Scholar] [CrossRef]
- Borges-Argáez, R.; Chan-Balan, R.; Cetina-Montejo, L.; Ayora-Talavera, G.; Sansores-Peraza, P.; Gómez-Carballo, J.; Cáceres-Farfán, M. In vitro evaluation of anthraquinones from Aloe vera (Aloe barbadensis Miller) roots and several derivatives against strains of influenza virus. Ind. Crops Prod. 2019, 132, 468–475. [Google Scholar] [CrossRef]
- Sun, Z.; Yu, C.; Wang, W.; Yu, G.; Zhang, T.; Zhang, L.; Zhang, J.; Wei, K. Aloe Polysaccharides Inhibit Influenza A Virus Infection—A Promising Natural Anti-flu Drug. Front. Microbiol. 2018, 9, 2338. [Google Scholar] [CrossRef]
- Kahlon, J.B.; Kemp, M.C.; Yawei, N.; Carpenter, R.H.; Shannon, W.M.; McAnalley, B.H. In vitro evaluation of the synergistic antiviral effects of acemannan in combination with azidothymidine and acyclovir. Mol. Biother. 1991, 3, 214–223. [Google Scholar]
- Yates, K.M.; Rosenberg, L.J.; Harris, C.K.; Bronstad, D.C.; King, G.K.; Biehle, G.A.; Walker, B.; Ford, C.R.; Hall, J.E.; Tizard, I.R. Pilot study of the effect of acemannan in cats infected with feline immunodeficiency virus. Vet. Immunol. Immunopathol. 1992, 35, 177–189. [Google Scholar] [CrossRef]
- Sheets, M.A.; Unger, B.A.; Giggleman, G.F., Jr.; Tizard, I.R. Studies of the effect of acemannan on retrovirus infections: Clinical stabilization of feline leukemia virus-infected cats. Mol. Biother. 1991, 3, 41–45. [Google Scholar]
- Rolta, R.; Yadav, R.; Salaria, D.; Trivedi, S.; Imran, M.; Sourirajan, A.; Baumler, D.J.; Dev, K. In silico screening of hundred phytocompounds of ten medicinal plants as potential inhibitors of nucleocapsid phosphoprotein of COVID-19: An approach to prevent virus assembly. J. Biomol. Struct. Dyn. 2021, 39, 7017–7034. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Zeng, Y.; Liu, Y.; You, L.; Yin, X.; Fu, J.; Ni, J. Aloe-emodin: A review of its pharmacology, toxicity, and pharmacokinetics. Phytother. Res. 2020, 34, 270–281. [Google Scholar] [CrossRef] [PubMed]
- Şeker Karatoprak, G.; Küpeli Akkol, E.; Yücel, Ç.; Bahadır Acıkara, Ö.; Sobarzo-Sánchez, E. Advances in Understanding the Role of Aloe Emodin and Targeted Drug Delivery Systems in Cancer. Oxidative Med. Cell. Longev. 2022, 2022, 7928200. [Google Scholar] [CrossRef] [PubMed]
- Groom, Q.J.; Reynolds, T. Barbaloin in aloe species. Planta Med. 1987, 53, 345–348. [Google Scholar] [CrossRef] [PubMed]
- Lewis, D.S.M.; Ho, J.; Wills, S.; Kawall, A.; Sharma, A.; Chavada, K.; Ebert, M.C.C.J.C.; Evoli, S.; Singh, A.; Rayalam, S.; et al. Aloin isoforms (A and B) selectively inhibits proteolytic and deubiquitinating activity of papain like protease (PLpro) of SARS-CoV-2 in vitro. Sci. Rep. 2022, 12, 2145. [Google Scholar] [CrossRef]
- Park, M.-Y.; Kwon, H.-J.; Sung, M.-K. Plasma, tissue and urinary levels of aloin in rats after the administration of pure aloin. Nutr. Res. Pract. 2008, 2, 17–21. [Google Scholar] [CrossRef]
- Akao, T.; Che, Q.M.; Kobashi, K.; Hattori, M.; Namba, T. A purgative action of barbaloin is induced by Eubacterium sp. strain BAR, a human intestinal anaerobe, capable of transforming barbaloin to aloe-emodin anthrone. Biol. Pharm. Bull. 1996, 19, 136–138. [Google Scholar] [CrossRef] [Green Version]
- Final Report on the Safety Assessment of Aloe andongensis Extract, Aloe andongensis Leaf Juice, Aloe arborescens Leaf Extract, Aloe arborescens Leaf Juice, Aloe arborescens Leaf Protoplasts, Aloe barbadensis Flower Extract, Aloe barbadensis Leaf, Aloe barbadensis Leaf Extract, Aloe barbadensis Leaf Juice, Aloe barbadensis Leaf Polysaccharides, Aloe barbadensis Leaf Water, Aloe ferox Leaf Extract, Aloe ferox Leaf Juice, and Aloe ferox Leaf Juice Extract. Int. J. Toxicol. 2007, 26, 1–50. [CrossRef]
- Guo, X.; Mei, N. Aloe vera: A review of toxicity and adverse clinical effects. J. Environ. Sci. Health Part C 2016, 34, 77–96. [Google Scholar] [CrossRef]
- Ni, Y.; Yates, K.M.; Tizard, I.R. Aloe polysaccharides. In Aloes: The Genus Aloe; Reynolds, T., Ed.; CRC Press LLC: Boca Raton, FL, USA, 2004. [Google Scholar]
- Hamman, J.H. Composition and applications of Aloe vera leaf gel. Molecules 2008, 13, 1599–1616. [Google Scholar] [CrossRef] [Green Version]
- Rai, K.R.; Shrestha, P.; Yang, B.; Chen, Y.; Liu, S.; Maarouf, M.; Chen, J.-L. Acute Infection of Viral Pathogens and Their Innate Immune Escape. Front. Microbiol. 2021, 12, 672026. [Google Scholar] [CrossRef] [PubMed]
- Rouse, B.T.; Sehrawat, S. Immunity and immunopathology to viruses: What decides the outcome? Nat. Rev. Immunol. 2010, 10, 514–526. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.N.; Rouse, B.T. Immune responses to viruses. Clin. Immunol. 2008, 421–431. [Google Scholar] [CrossRef]
- Chen, R.; Lan, Z.; Ye, J.; Pang, L.; Liu, Y.; Wu, W.; Qin, X.; Guo, Y.; Zhang, P. Cytokine Storm: The Primary Determinant for the Pathophysiological Evolution of COVID-19 Deterioration. Front. Immunol. 2021, 12, 589095. [Google Scholar] [CrossRef]
- Ciancanelli, M.J.; Huang, S.X.L.; Luthra, P.; Garner, H.; Itan, Y.; Volpi, S.; Lafaille, F.G.; Trouillet, C.; Schmolke, M.; Albrecht, R.A.; et al. Infectious disease. Life-threatening influenza and impaired interferon amplification in human IRF7 deficiency. Science 2015, 348, 448–453. [Google Scholar] [CrossRef] [Green Version]
- Stertz, S.; Hale, B.G. Interferon system deficiencies exacerbating severe pandemic virus infections. Trends Microbiol. 2021, 29, 973–982. [Google Scholar] [CrossRef]
- Yu, C.S.; Yu, F.S.; Chan, J.K.; Li, T.M.; Lin, S.S.; Chen, S.C.; Hsia, T.C.; Chang, Y.H.; Chung, J.G. Aloe-emodin affects the levels of cytokines and functions of leukocytes from Sprague-Dawley rats. In Vivo 2006, 20, 505–509. [Google Scholar]
- Park, M.-Y.; Kwon, H.-J.; Sung, M.-K. Evaluation of Aloin and Aloe-Emodin as Anti-Inflammatory Agents in Aloe by Using Murine Macrophages. Biosci. Biotechnol. Biochem. 2009, 73, 828–832. [Google Scholar] [CrossRef]
- Kshirsagar, A.D.; Panchal, P.V.; Harle, U.N.; Nanda, R.K.; Shaikh, H.M. Anti-Inflammatory and Antiarthritic Activity of Anthraquinone Derivatives in Rodents. Int. J. Inflam. 2014, 2014, 690596. [Google Scholar] [CrossRef] [Green Version]
- Xian, M.; Cai, J.; Zheng, K.; Liu, Q.; Liu, Y.; Lin, H.; Liang, S.; Wang, S. Aloe-emodin prevents nerve injury and neuroinflammation caused by ischemic stroke via the PI3K/AKT/mTOR and NF-κB pathway. Food Funct. 2021, 12, 8056–8067. [Google Scholar] [CrossRef]
- Li, S.-W.; Yang, T.-C.; Lai, C.-C.; Huang, S.-H.; Liao, J.-M.; Wan, L.; Lin, Y.-J.; Lin, C.-W. Antiviral activity of aloe-emodin against influenza A virus via galectin-3 up-regulation. Eur. J. Pharmacol. 2014, 738, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Park, M.-Y.; Kwon, H.-J.; Sung, M.-K. Dietary aloin, aloesin, or aloe-gel exerts anti-inflammatory activity in a rat colitis model. Life Sci. 2011, 88, 486–492. [Google Scholar] [CrossRef] [PubMed]
- Langmead, L.; Makins, R.J.; Rampton, D.S. Anti-inflammatory effects of Aloe vera gel in human colorectal mucosa in vitro. Aliment. Pharmacol. Ther. 2004, 19, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Yazdani, N.; Hossini, S.E.; Edalatmanesh, M.A. Anti-inflammatory Effect of Aloe vera Extract on Inflammatory Cytokines of Rats Fed with a High-Fat Diet (HFD). Jundishapur J. Nat. Pharm. Prod. 2021, 17, e114323. [Google Scholar] [CrossRef]
- Budai, M.M.; Varga, A.; Milesz, S.; Tőzsér, J.; Benkő, S. Aloe vera downregulates LPS-induced inflammatory cytokine production and expression of NLRP3 inflammasome in human macrophages. Mol. Immunol. 2013, 56, 471–479. [Google Scholar] [CrossRef] [Green Version]
- Yun, N.; Lee, C.H.; Lee, S.M. Protective effect of Aloe vera on polymicrobial sepsis in mice. Food Chem. Toxicol. 2009, 47, 1341–1348. [Google Scholar] [CrossRef]
- Paul, S.; Modak, D.; Chattaraj, S.; Nandi, D.; Sarkar, A.; Roy, J.; Chaudhuri, T.K.; Bhattacharjee, S. Aloe vera gel homogenate shows anti-inflammatory activity through lysosomal membrane stabilization and downregulation of TNF-α and Cox-2 gene expressions in inflammatory arthritic animals. Future J. Pharm. Sci. 2021, 7, 12. [Google Scholar] [CrossRef]
- Na, K.; Lkhagva-Yondon, E.; Kim, M.; Lim, Y.R.; Shin, E.; Lee, C.K.; Jeon, M.S. Oral treatment with Aloe polysaccharide ameliorates ovalbumin-induced atopic dermatitis by restoring tight junctions in skin. Scand. J. Immunol. 2020, 91, e12856. [Google Scholar] [CrossRef]
- López, Z.; Femenia, A.; Núñez-Jinez, G.; Salazar Zúñiga, M.N.; Cano, M.E.; Espino, T.; Knauth, P. In vitro Immunomodulatory Effect of Food Supplement from Aloe vera. Evid. Based Complement. Alternat. Med. 2019, 2019, 5961742. [Google Scholar] [CrossRef] [Green Version]
- Ahluwalia, B.; Magnusson, M.K.; Isaksson, S.; Larsson, F.; Öhman, L. Effects of Aloe barbadensis Mill. extract (AVH200®) on human blood T cell activity in vitro. J. Ethnopharmacol. 2016, 179, 301–309. [Google Scholar] [CrossRef]
- Womble, D.; Helderman, J.H. The Impact of Acemannan on the Generation and Function of Cytotoxic T-Lymphocytes. Immunopharmacol. Immunotoxicol. 1992, 14, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Tizard, I.R. Activation of a mouse macrophage cell line by acemannan: The major carbohydrate fraction from Aloe vera gel. Immunopharmacology 1996, 35, 119–128. [Google Scholar] [CrossRef]
- Karaca, K.; Sharma, J.M.; Nordgren, R. Nitric oxide production by chicken macrophages activated by Acemannan, a complex carbohydrate extracted from Aloe vera. Int. J. Immunopharmacol. 1995, 17, 183–188. [Google Scholar] [CrossRef]
- Lee, J.K.; Lee, M.K.; Yun, Y.-P.; Kim, Y.; Kim, J.S.; Kim, Y.S.; Kim, K.; Han, S.S.; Lee, C.-K. Acemannan purified from Aloe vera induces phenotypic and functional maturation of immature dendritic cells. Int. Immunopharmacol. 2001, 1, 1275–1284. [Google Scholar] [CrossRef]
- Jia, D.; Rahbar, R.; Chan, R.W.; Lee, S.M.; Chan, M.C.; Wang, B.X.; Baker, D.P.; Sun, B.; Peiris, J.S.; Nicholls, J.M.; et al. Influenza virus non-structural protein 1 (NS1) disrupts interferon signaling. PLoS ONE 2010, 5, e13927. [Google Scholar] [CrossRef] [Green Version]
- Jeon, S.B.; Yoon, H.J.; Chang, C.Y.; Koh, H.S.; Jeon, S.H.; Park, E.J. Galectin-3 exerts cytokine-like regulatory actions through the JAK-STAT pathway. J. Immunol. 2010, 185, 7037–7046. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Ren, A.; Wang, X.; Fan, X.; Zhao, Y.; Gao George, F.; Cleary, P.; Wang, B. Influenza viral neuraminidase primes bacterial coinfection through TGF-β–mediated expression of host cell receptors. Proc. Natl. Acad. Sci. USA 2015, 112, 238–243. [Google Scholar] [CrossRef] [Green Version]
- Denney, L.; Branchett, W.; Gregory, L.G.; Oliver, R.A.; Lloyd, C.M. Epithelial-derived TGF-β1 acts as a pro-viral factor in the lung during influenza A infection. Mucosal Immunol. 2018, 11, 523–535. [Google Scholar] [CrossRef] [Green Version]
- Seo, S.H.; Webster, R.G. Tumor necrosis factor alpha exerts powerful anti-influenza virus effects in lung epithelial cells. J. Virol. 2002, 76, 1071–1076. [Google Scholar] [CrossRef] [Green Version]
- Le Bel, M.; Brunet, A.; Gosselin, J. Leukotriene B4, an Endogenous Stimulator of the Innate Immune Response against Pathogens. J. Innate Immun. 2014, 6, 159–168. [Google Scholar] [CrossRef]
- Kumar, S.; Singh, B.K.; Pandey, A.K.; Kumar, A.; Sharma, S.K.; Raj, H.G.; Prasad, A.K.; Eycken, E.V.D.; Parmar, V.S.; Ghosh, B. A chromone analog inhibits TNF-α induced expression of cell adhesion molecules on human endothelial cells via blocking NF-κB activation. Bioorg. Med. Chem. 2007, 15, 2952–2962. [Google Scholar] [CrossRef] [PubMed]
- Eamlamnam, K.; Patumraj, S.; Visedopas, N.; Thong-Ngam, D. Effects of Aloe vera and sucralfate on gastric microcirculatory changes, cytokine levels and gastric ulcer healing in rats. World J. Gastroenterol. 2006, 12, 2034–2039. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.K.; Kumar, A.; Pereira, M.D.; Siddiqi, N.J.; Sharma, B. Anti-Inflammatory and Antioxidative Potential of Aloe vera on the Cartap and Malathion Mediated Toxicity in Wistar Rats. Int. J. Environ. Res. Public Health 2020, 17, 5177. [Google Scholar] [CrossRef]
- Ahluwalia, B.; Magnusson, M.K.; Isaksson, S.; Larsson, F.; Öhman, L. Aloe barbadensis Mill. extract improves symptoms in IBS patients with diarrhoea: Post hoc analysis of two randomized double-blind controlled studies. Ther. Adv. Gastroenterol. 2021, 14, 17562848211048133. [Google Scholar] [CrossRef] [PubMed]
- Im, S.-A.; Oh, S.-T.; Song, S.; Kim, M.-R.; Kim, D.-S.; Woo, S.-S.; Jo, T.H.; Park, Y.I.; Lee, C.-K. Identification of optimal molecular size of modified Aloe polysaccharides with maximum immunomodulatory activity. Int. Immunopharmacol. 2005, 5, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, L.; Tizard, I.R. Induction of Apoptosis in a Macrophage Cell Line RAW 264.7 by Acemannan, a β-(1,4)-Acetylated Mannan. Mol. Pharmacol. 1998, 53, 415. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Simmonds, M.S.J. Pharmacodynamics of Aloe vera and acemannan in therapeutic applications for skin, digestion, and immunomodulation. Phytother. Res. 2021, 35, 6572–6584. [Google Scholar] [CrossRef]
- Chinnah, A.D.; Baig, M.A.; Tizard, I.R.; Kemp, M.C. Antigen dependent adjuvant activity of a polydispersed β-(1,4)-linked acetylated mannan (acemannan). Vaccine 1992, 10, 551–557. [Google Scholar] [CrossRef]
- Gauntt, C.J.; Wood, H.J.; McDaniel, H.R.; McAnalley, B.H. Aloe polymannose enhances anti-coxsackievirus antibody titres in mice. Phytother. Res. 2000, 14, 261–266. [Google Scholar] [CrossRef]
- Song, E.-J.; Españo, E.; Nam, J.-H.; Kim, J.; Shim, K.-S.; Shin, E.; Park, Y.I.; Lee, C.-K.; Kim, J.-K. Adjuvanticity of Processed Aloe vera gel for Influenza Vaccination in Mice. Immune Netw. 2020, 20, e31. [Google Scholar] [CrossRef]
- Vahedi, G.; Taghavi, M.; Maleki, A.K.; Habibian, R. The effect of Aloe vera extract on humoral and cellular immune response in rabbit. Afr. J. Biotechnol. Afr. J. Biotechnol. 2011, 10, 5225–5228. [Google Scholar]
- Nikookalam, M.; Salehian, Z.; Mirzaee, S.; Ajideh, R.; Mahdavi, M.; Yazdi, M. Aloe Vera Extracted Polysaccharides Shift the Immune Responses of Tumor Bearing Mice Toward Th1 Pattern: Animal Study. Biomed. J. Sci. Tech. Res. 2019, 16, 12148–12156. [Google Scholar] [CrossRef]
- Hwang, J.H.; Oh, M.R.; Hwang, J.H.; Choi, E.K.; Jung, S.J.; Song, E.J.; Españo, E.; Webby, R.J.; Webster, R.G.; Kim, J.K.; et al. Effect of processed Aloe vera gel on immunogenicity in inactivated quadrivalent influenza vaccine and upper respiratory tract infection in healthy adults: A randomized double-blind placebo-controlled trial. Phytomedicine 2021, 91, 153668. [Google Scholar] [CrossRef] [PubMed]
- Bałan, B.J.; Niemcewicz, M.; Kocik, J.; Jung, L.; Skopińska-Różewska, E.; Skopiński, P. Oral administration of Aloe vera gel, anti-microbial and anti-inflammatory herbal remedy, stimulates cell-mediated immunity and antibody production in a mouse model. Cent. Eur. J. Immunol. 2014, 39, 125–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plotkin, S.A. Correlates of protection induced by vaccination. Clin. Vaccine Immunol. 2010, 17, 1055–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sridhar, S. Heterosubtypic T-Cell Immunity to Influenza in Humans: Challenges for Universal T-Cell Influenza Vaccines. Front. Immunol. 2016, 7, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baruah, A.; Bordoloi, M.; Deka Baruah, H.P. Aloe vera: A multipurpose industrial crop. Ind. Crops Prod. 2016, 94, 951–963. [Google Scholar] [CrossRef]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Orhan, I.E.; Banach, M.; Rollinger, J.M.; Barreca, D.; Weckwerth, W.; Bauer, R.; Bayer, E.A.; et al. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug. Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Source | Extract | Virus | Observation/Target | Ref |
|---|---|---|---|---|
| A. barbadensis (A. vera) | Freeze-dried and ethanol-treated extracts from leaf gel filet | Human cytomegalovirus | Middle stages of infection (DNA synthesis) | [8] |
| Glycerin extract from leaves | Herpes simplex virus 1 | Inhibited infection in vitro | [9] | |
| Glycerin extract from leaves Hot glycerin extract from leaf gel | Herpes simplex virus 2 | Inhibited infection in vitro; Inhibited pre-entry, entry, post-attachment stages | [9,10] [8,11] | |
| Commercial freeze-dried powder | Porcine epidemic diarrhea virus | Inhibited replication Inhibited post-entry stages; Protected piglets from intestinal pathology | [12] | |
| Distilled water precipitate | Murine norovirus 1 | Virucidal effects | [13] | |
| A. secundiflora | Distilled water leaf exudate | Newcastle disease virus | Reduced disease severity | [11] |
| A. hijanensis | Fractionated leaf homogenate | Newcastle disease virus | Reduced egg infectivity | [14] |
| Fractionated leaf homogenate | Influenza A H5N1 virus | Reduced egg infectivity | [14] | |
| Fractionated leaf homogenate | Egg drop syndrome virus | Reduced egg infectivity | [14] | |
| Fractionated leaf homogenate | Avian paramyxovirus type 1 | Reduced egg infectivity | [14] | |
| A. arborescens | Commercial aqueous extract | Human rhinovirus 14 | Inhibited infection/production | [15] |
| Commercial aqueous extract | Influenza A and B viruses | Inhibited infection/production | [15] | |
| Commercial aqueous extract | Parainfluenza virus 3 | Modest inhibition of infection | [15] | |
| Commercial aqueous extract | Respiratory syncytial virus | Modest inhibition of infection | [15] | |
| Commercial aqueous extract | Upper respiratory tract viruses | Reduced recurrence of upper respiratory tract infections in children | [16] |
| Compound | Virus | Observation/Mode of Action | Ref |
|---|---|---|---|
| Aloe emodin | Herpes simplex virus (HSV)1 | Inhibited infection in vitro; disrupted the viral envelope | [9] |
| HSV2 | Inhibited infection in vitro | [9] | |
| Varicella zoster virus | Inhibited infection in vitro | [9] | |
| Pseudorabies virus | Inhibited infection in vitro | [9] | |
| Influenza A virus | Inhibited infection in vitro | [9] | |
| Enterovirus 71 | Reduced virus production in vitro | [20] | |
| Japanese encephalitis virus (JEV) | Reduced virus production in vitro; Inhibited JEV NS2B-NS3A protease | [20] | |
| Severe acute respiratory syndrome coronavirus | Inhibited 3C-like protease | [22] | |
| Hepatitis B virus (HBV) | Reduced production of HBV S and E antigens; May bind HBV polymerase | [23] | |
| Aloin | Influenza A and B viruses | Inhibited infection in vitro; Improved mouse survival after influenza A H1N1 (PR8) challenge; Accelerated viral clearance in mice | [24] |
| HBV | Reduced production of HBV S and E antigens | [23] | |
| Hemorrhagic septicemia rhabdovirus | Inhibited viral infection in vitro; May disrupt the viral envelope | [25] | |
| Aloesaponarin II | Influenza A virus | Inhibited infection of oseltamivir-susceptible influenza A virus in vitro | [26] |
| Aloe polysaccharides | Influenza A H1N1 (PR8) | Inhibited virus production in vitro; Protected mice from virus-induced pathology; Caused irregularities in viral particle shape | [27] |
| Acemannan | Human immunodeficiency virus | Inhibited viral replication; Synergistic effects with azithymidine in vitro | [28] |
| Feline immunodeficiency virus | Stable clinical states | [29] | |
| Feline leukemia virus | Improved clinical signs | [30] |
| Component | Model | Condition/Stimulation | Observed Effects | Ref |
|---|---|---|---|---|
| Aloe emodin | Rat leukocytes | ↓ NK activity, macrophage phagocytosis ↑ IL-1β, TNF-α in leukocytes | [48] | |
| Mouse macrophages | LPS | ↓ Nos, Cox mRNA ↓ NO, PGE2 | [49] | |
| Rat | Arthritis | ↓ NO in rat paw edema model | [50] | |
| Rat | Cerebral occlusion reperfusion | ↓ TNF-α in serum of rat model ↓ IL-6, TNF-α, NO in microglial BV2 cells | [51] | |
| TE-671 cells, HL-CZ cells | ↑ IFN-α, ISG promoters (ISRE, GAS) ↑ ISG (PKR, OAS) mRNA levels | [20] | ||
| MDCK cells | Influenza A virus/ Influenza NS1 protein | ↑ Galectin-3, IFN-β, IFNγ ↑ pSTAT1, ISGs (PKR, OAS) | [52] | |
| Aloin | Mouse | Influenza A virus | ↑ CD4+, CD8+ T cells in lungs ↑ IFN-γ, TNF-α in T cells ↓ TGF-β in T cells | [24] |
| Aloesin | Rat | Colitis | ↓ Granulocyte infiltration in rat colon ↓ TNF-α, PGE2, LTB4 in serum ↓Tnfa, Il1b mRNA | [53] |
| A. vera gel | Human biopsies, Caco-2 cells | Ulcerative colitis | ↓ PGE2 in biopsies from patients with active ulcerative colitis ↓ IL-8 in Caco-2 cells | [54] |
| Rat | High-fat diet | ↓ TNF-α, TGF-β, and IL-6 | [55] | |
| Macrophages (Primary and cell line) | LPS | ↓ IL-6, IL-8, IL-1β, TNF-α, NLRP3, P2X7 ↓ NF-κB, and MAPK pathway kinases in THP-1 cells and primary macrophages | [56] | |
| Mouse | Sepsis | ↓ Multiorgan dysfunction ↑ Bacterial clearance, survival ↓ TNF-α, IL-6, IL-1β in serum | [57] | |
| Rat | Arthritis | ↓ Paw swelling ↓ Tnfa, Cox2 mRNA | [58] | |
| Mouse | Atopic dermatitis | ↓ Histopathological markers ↓ IFN-γ, IL-4, IL-17A in skin lesions | [59] | |
| THP-1 cells | LPS | ↓ TNF-α, IL-1β ↑ IL-10 | [60] | |
| Acemannan | T cells from human PBMC | ↓ T cell activation and proliferation ↓ IL-2, IL-5, IL-17A | [61] | |
| T cells from human PBMC | Alloantigen | ↑ Cytotoxic T cell generation ↑ Response to alloantigen | [62] | |
| Macrophages | ↑ Macrophage activation upon IFN-γ stimulation; ↑ IL-6, TNF-α, NO in macrophages | [63] | ||
| Chicken | ↑ NO in splenocytes and macrophages | [64] | ||
| Mouse | ↑ Mitogenesis of splenocytes ↑ DC maturation and differentiation | [65] |
| Virus/Antigen | Adjuvant Form (Administration) | Organism | Observed Effects | Ref |
|---|---|---|---|---|
| Newcastle disease virus | Acemannan (s.c.) | Chicken | ↑ Protective antibody titers | [79] |
| Coxsackievirus B3 | Acemannan (i.p.) | Mouse | ↑ Antibody titers | [80] |
| Pandemic AH1N1 antigen, Quadrivalent influenza vaccine | Processed Aloe gel (p.o.) | Mouse | ↑ Hemagglutinin and neutralizing antibody titers ↑ Survival after homologous and heterologous challenge | [81] |
| Myxomatosis virus | Aloe polysaccharides (p.o.) | Rabbit | ↑ IgA, IgG, IgM ↑ CD4+, CD8+ T cells | [82] |
| Human papillomavirus 16 E7d | A. vera gel (p.o.) | Mouse | ↑ IFN-γ, IL-4 (with alum) ↑ IgG2 (with Montanide) | [83] |
| Quadrivalent influenza vaccine | Processed Aloe gel (p.o.) | Human | ↑ Geometric mean titer increase, geometric mean fold increase ↓ Incidence of upper respiratory tract infection | [84] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Españo, E.; Kim, J.; Kim, J.-K. Utilization of Aloe Compounds in Combatting Viral Diseases. Pharmaceuticals 2022, 15, 599. https://doi.org/10.3390/ph15050599
Españo E, Kim J, Kim J-K. Utilization of Aloe Compounds in Combatting Viral Diseases. Pharmaceuticals. 2022; 15(5):599. https://doi.org/10.3390/ph15050599
Chicago/Turabian StyleEspaño, Erica, Jiyeon Kim, and Jeong-Ki Kim. 2022. "Utilization of Aloe Compounds in Combatting Viral Diseases" Pharmaceuticals 15, no. 5: 599. https://doi.org/10.3390/ph15050599
APA StyleEspaño, E., Kim, J., & Kim, J.-K. (2022). Utilization of Aloe Compounds in Combatting Viral Diseases. Pharmaceuticals, 15(5), 599. https://doi.org/10.3390/ph15050599