
The Association of the Polymorphisms in the FUT8-Related Locus with the Plasma Glycosylation in Post-Traumatic Stress Disorder
 , , , , , and
, , , , , and 
Abstract
1. Introduction
2. Results
2.1. Association of the FUT8-Related Polymorphisms with PTSD
2.2. Differences in the N-Glycome between the PTSD and the Control Group
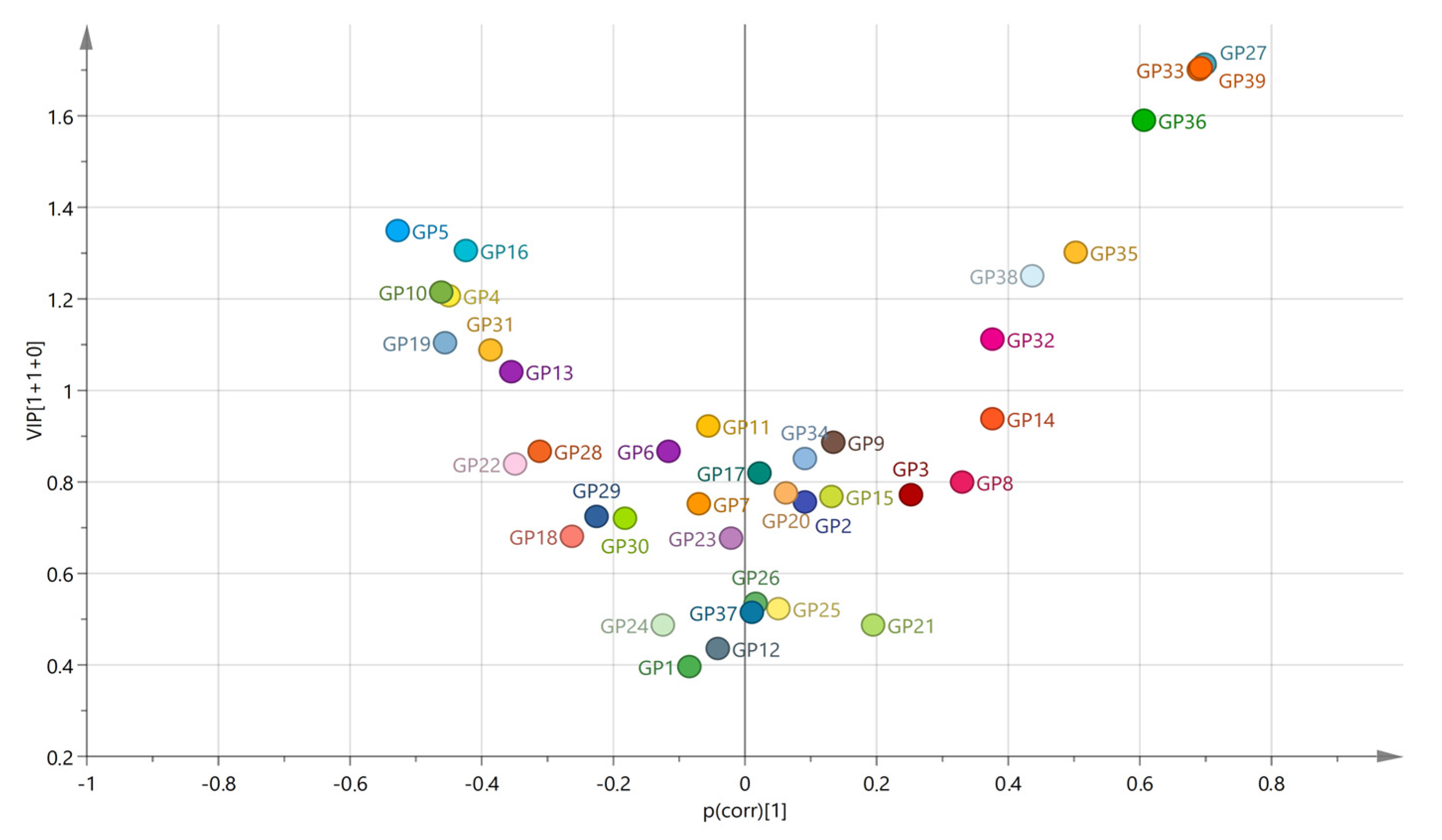
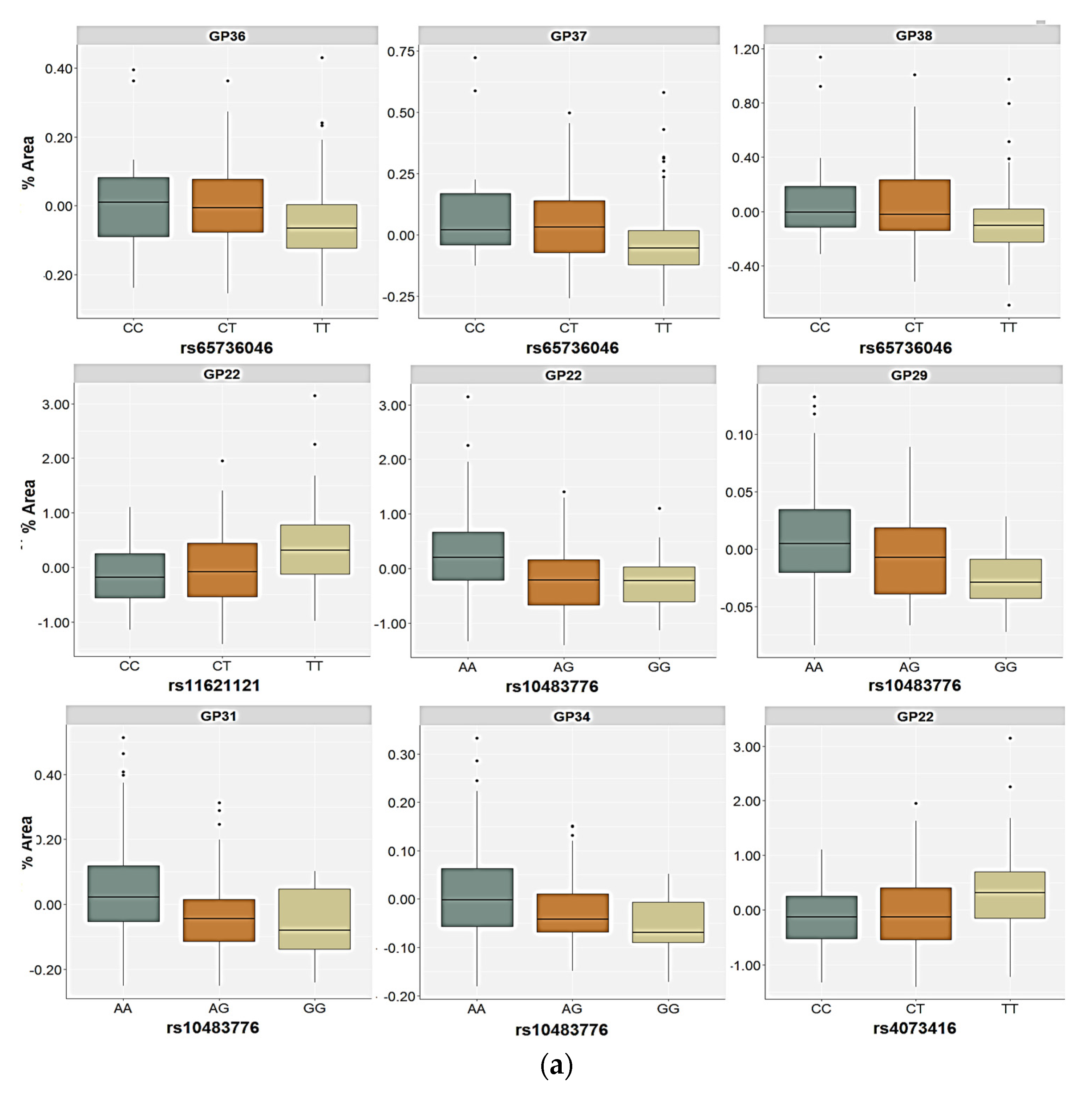
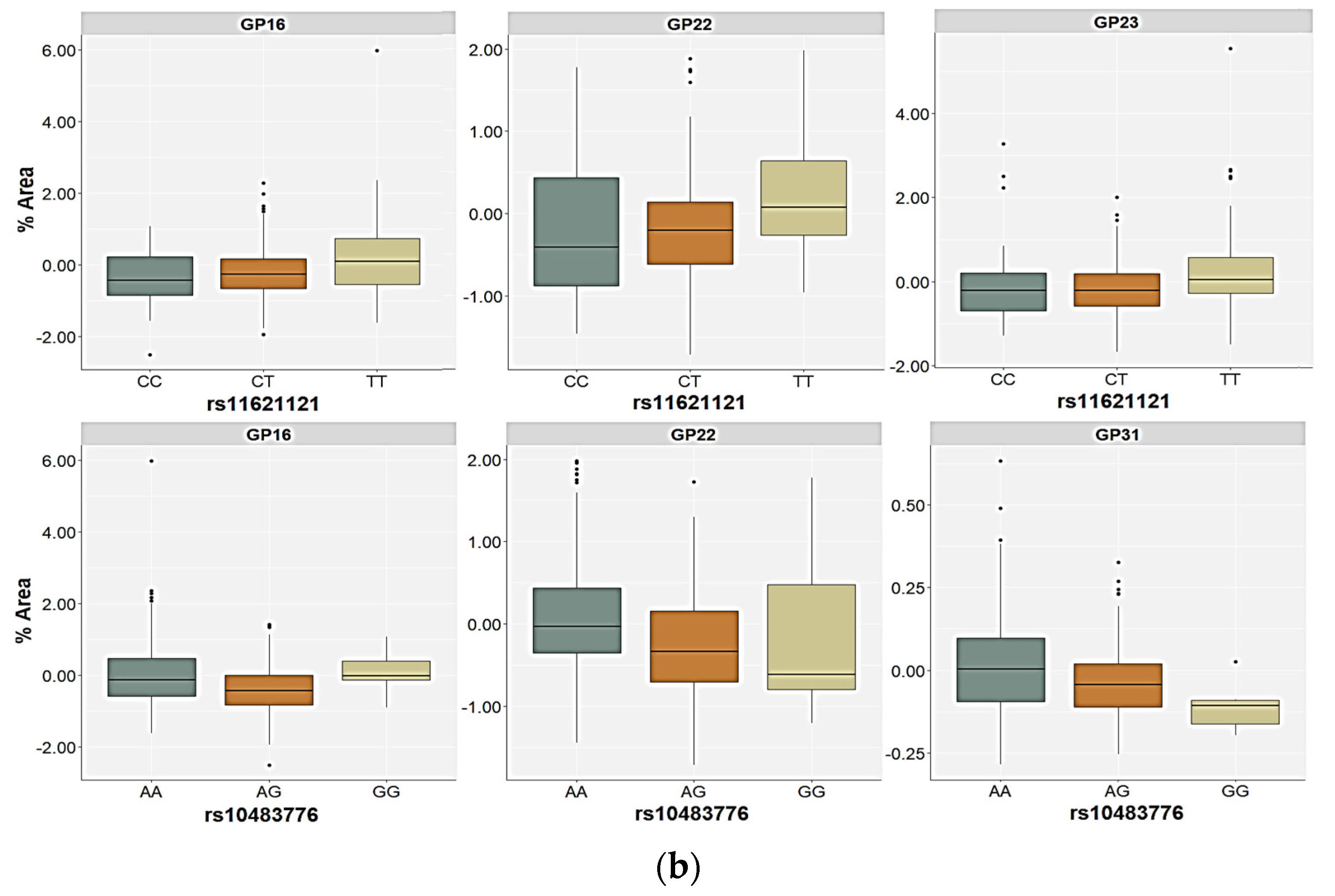
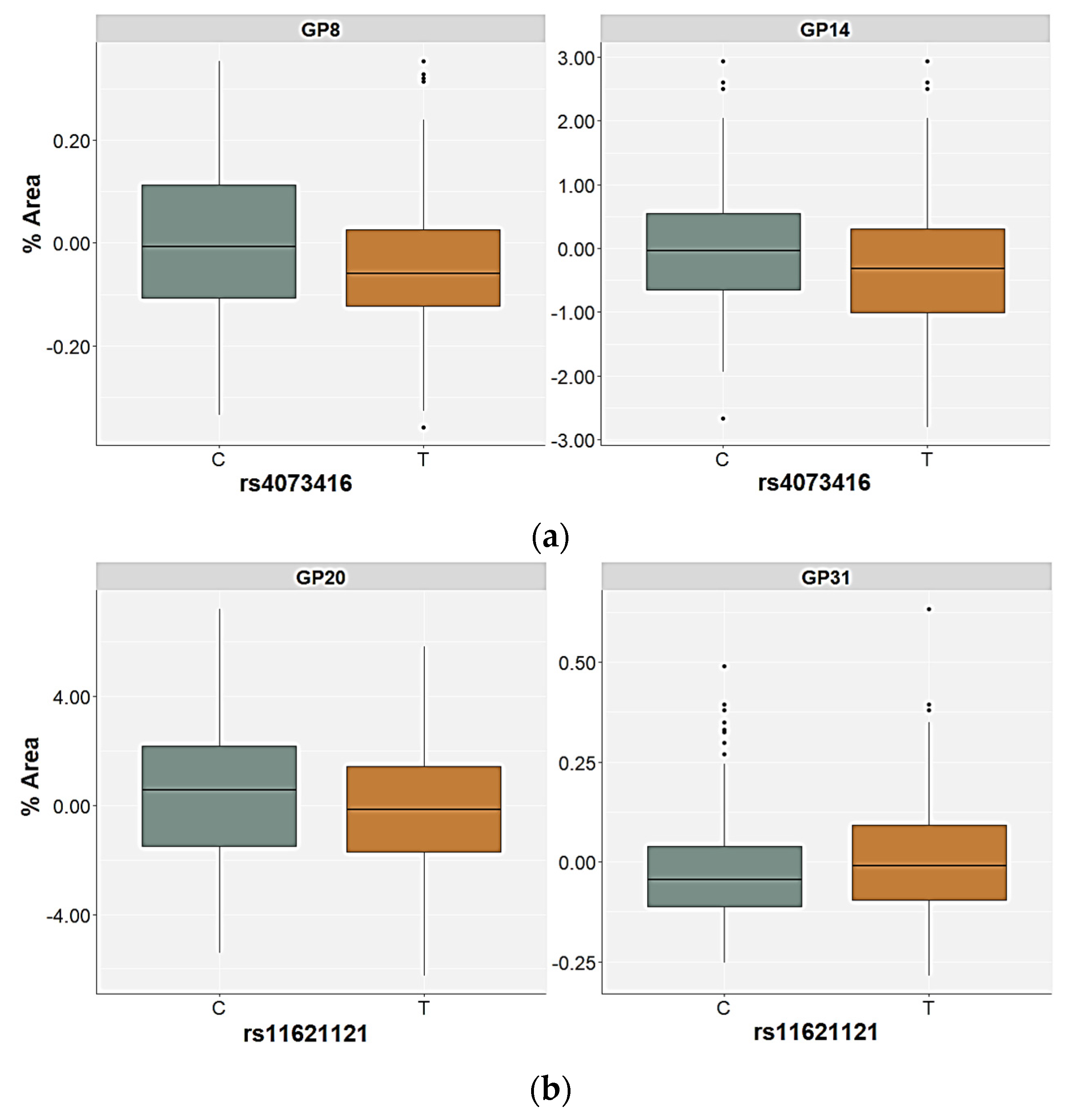
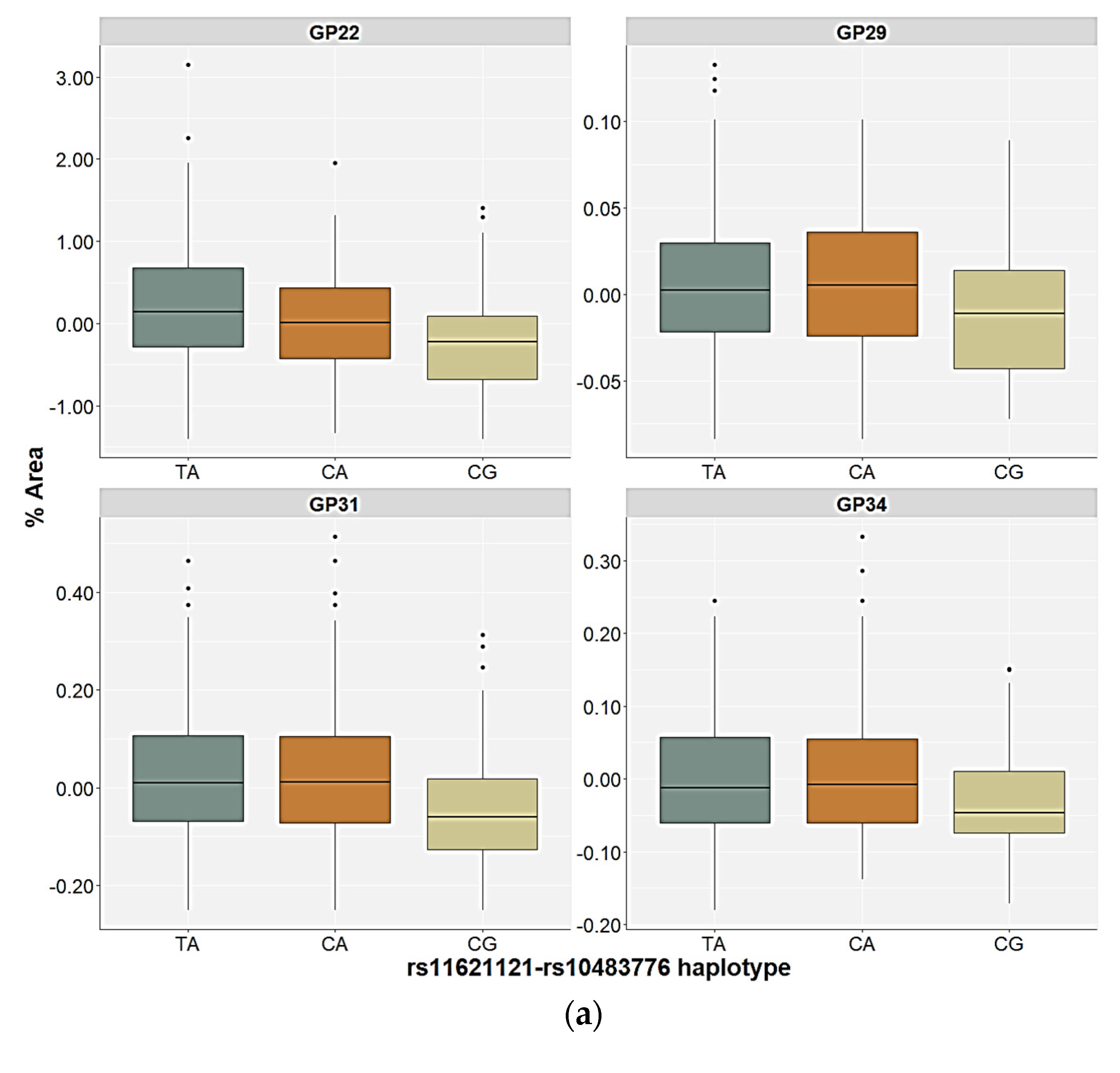
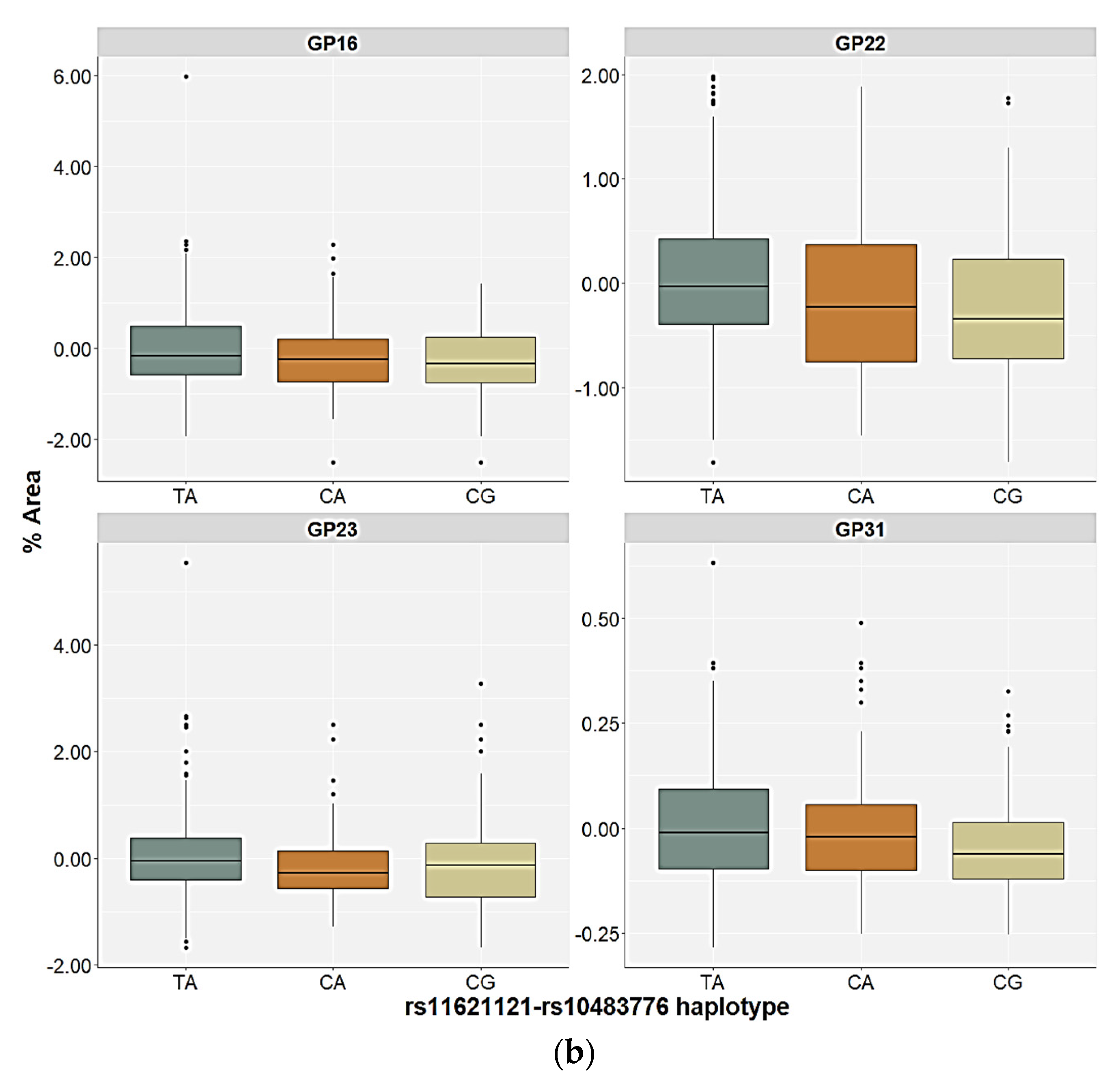
2.3. Association of the FUT8-Related Polymorphisms with the N-Glycan Levels
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. Blood Processing
4.3. Determination of N-Glycan Levels in the Plasma
4.4. Genotyping
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Britvić, D.; Antičević, V.; Kaliterna, M.; Lušić, L.; Beg, A.; Brajević-Gizdić, I.; Kudrić, M.; Stupalo, Ž.; Krolo, V.; Pivac, N. Comorbidities with Posttraumatic Stress Disorder (PTSD) among combat veterans: 15 years postwar analysis. Int. J. Clin. Health Psychol. 2015, 15, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Dedert, E.A.; Calhoun, P.S.; Watkins, L.L.; Sherwood, A.; Beckham, J.C. Posttraumatic stress disorder, cardiovascular, and metabolic disease: A review of the evidence. Ann. Behav. Med. 2010, 39, 61–78. [Google Scholar] [CrossRef] [PubMed]
- Barone, R.; Sturiale, L.; Palmigiano, A.; Zappia, M.; Garozzo, D. Glycomics of pediatric and adulthood diseases of the central nervous system. J. Proteom. 2012, 75, 5123–5139. [Google Scholar] [CrossRef] [PubMed]
- Stanta, J.L.; Saldova, R.; Struwe, W.B.; Byrne, J.C.; Leweke, F.M.; Rothermund, M.; Rahmoune, H.; Levin, Y.; Guest, P.C.; Bahn, S.; et al. Identification of N-glycosylation changes in the CSF and serum in patients with schizophrenia. J. Proteome Res. 2010, 9, 4476–4489. [Google Scholar] [CrossRef]
- Park, D.I.; Štambuk, J.; Razdorov, G.; Pučić-Baković, M.; Martins-De-Souza, D.; Lauc, G.; Turck, C.W. Blood plasma/IgG N-glycome biosignatures associated with major depressive disorder symptom severity and the antidepressant response. Sci. Rep. 2018, 8, 179. [Google Scholar] [CrossRef]
- Boeck, C.; Pfister, S.; Bürkle, A.; Vanhooren, V.; Libert, C.; Salinas-Manrique, J.; Dietrich, D.E.; Kolassa, I.T.; Karabatsiakis, A. Alterations of the serum N-glycan profile in female patients with Major Depressive Disorder. J. Affect. Disord. 2018, 234, 139–147. [Google Scholar] [CrossRef]
- Moreno-Villanueva, M.; Morath, J.; Vanhooren, V.; Elbert, T.; Kolassa, S.; Libert, C.; Bürkle, A.; Kolassa, I.-T. N-glycosylation profiling of plasma provides evidence for accelerated physiological aging in post-traumatic stress disorder. Transl. Psychiatry 2013, 3, e320. [Google Scholar] [CrossRef]
- Tudor, L.; Nedic Erjavec, G.; Nikolac Perkovic, M.; Konjevod, M.; Svob Strac, D.; Uzun, S.; Kozumplik, O.; Jovanovic, T.; Lauc, G.; Pivac, N. N-glycomic profile in combat related post-traumatic stress disorder. Biomolecules 2019, 9, 834. [Google Scholar] [CrossRef]
- Gudelj, I.; Lauc, G.; Pezer, M. Immunoglobulin G glycosylation in aging and diseases. Cell. Immunol. 2018, 333, 65–79. [Google Scholar] [CrossRef]
- Reily, C.; Stewart, T.J.; Renfrow, M.B.; Novak, J. Glycosylation in health and disease. Nat. Rev. Nephrol. 2019, 15, 346–366. [Google Scholar] [CrossRef]
- Moremen, K.W.; Tiemeyer, M.; Nairn, A.V. Vertebrate protein glycosylation: Diversity, synthesis and function. Nat. Rev. Mol. Cell Biol. 2012, 13, 448–462. [Google Scholar] [CrossRef] [PubMed]
- Ohtsubo, K.; Marth, J.D. Glycosylation in Cellular Mechanisms of Health and Disease. Cell 2006, 126, 855–867. [Google Scholar] [CrossRef] [PubMed]
- Peschke, B.; Keller, C.W.; Weber, P.; Quast, I.; Lünemann, J.D. Fc-galactosylation of human immunoglobulin gamma isotypes improves C1q binding and enhances complement-dependent cytotoxicity. Front. Immunol. 2017, 8, 646. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, Y.; Nimmerjahn, F.; Ravetch, J.V. Anti-inflammatory activity of immunoglobulin G resulting from Fc sialylation. Science 2006, 313, 670–673. [Google Scholar] [CrossRef]
- Markina, Y.V.; Gerasimova, E.V.; Markin, A.M.; Glanz, V.Y.; Wu, W.K.; Sobenin, I.A.; Orekhov, A.N. Sialylated immunoglobulins for the treatment of immuno-inflammatory diseases. Int. J. Mol. Sci. 2020, 21, 5472. [Google Scholar] [CrossRef] [PubMed]
- Mitoma, J.; Bao, X.; Petryanik, B.; Schaerli, P.; Gauguet, J.M.; Yu, S.Y.; Kawashima, H.; Saito, H.; Ohtsubo, K.; Marth, J.D.; et al. Critical functions of N-glycans in L-selectin-mediated lymphocyte homing and recruitment. Nat. Immunol. 2007, 8, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Ma, B.; Simala-Grant, J.L.; Taylor, D.E. Fucosylation in prokaryotes and eukaryotes. Glycobiology 2006, 16, 158R–184R. [Google Scholar] [CrossRef]
- Freeze, H.H. Genetic defects in the human glycome. Nat. Rev. Genet. 2006, 7, 537–551. [Google Scholar] [CrossRef]
- Becker, D.J.; Lowe, J.B. Fucose: Biosynthesis and biological function in mammals. Glycobiology 2003, 13, 41R–53R. [Google Scholar] [CrossRef]
- Gornik, O.; Lauc, G. Glycosylation of serum proteins in inflammatory diseases. Dis. Markers 2008, 25, 267–278. [Google Scholar] [CrossRef]
- Miyoshi, E.; Moriwaki, K.; Nakagawa, T. Biological function of fucosylation in cancer biology. J. Biochem. 2008, 143, 725–729. [Google Scholar] [CrossRef] [PubMed]
- Bastian, K.; Scott, E.; Elliott, D.J.; Munkley, J. Fut8 alpha-(1,6)-fucosyltransferase in cancer. Int. J. Mol. Sci. 2021, 22, 455. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Yokote, H.; Arao, T.; Maegawa, M.; Tanaka, K.; Fujita, Y.; Shimizu, C.; Hanafusa, T.; Fujiwara, Y.; Nishio, K. N-Glycan fucosylation of epidermal growth factor receptor modulates receptor activity and sensitivity to epidermal growth factor receptor tyrosine kinase inhibitor. Cancer Sci. 2008, 99, 1611–1617. [Google Scholar] [CrossRef]
- Okada, M.; Chikuma, S.; Kondo, T.; Hibino, S.; Machiyama, H.; Yokosuka, T.; Nakano, M.; Yoshimura, A. Blockage of Core Fucosylation Reduces Cell-Surface Expression of PD-1 and Promotes Anti-tumor Immune Responses of T Cells. Cell Rep. 2017, 20, 1017–1028. [Google Scholar] [CrossRef] [PubMed]
- García-García, A.; Serna, S.; Yang, Z.; Delso, I.; Taleb, V.; Hicks, T.; Artschwager, R.; Vakhrushev, S.Y.; Clausen, H.; Angulo, J.; et al. FUT8-Directed Core Fucosylation of N-glycans Is Regulated by the Glycan Structure and Protein Environment. ACS Catal. 2021, 11, 9052–9065. [Google Scholar] [CrossRef]
- Ng, B.G.; Xu, G.; Chandy, N.; Steyermark, J.; Shinde, D.N.; Radtke, K.; Raymond, K.; Lebrilla, C.B.; AlAsmari, A.; Suchy, S.F.; et al. Biallelic Mutations in FUT8 Cause a Congenital Disorder of Glycosylation with Defective Fucosylation. Am. J. Hum. Genet. 2018, 102, 188–195. [Google Scholar] [CrossRef]
- Wang, X.; Inoue, S.; Gu, J.; Miyoshi, E.; Noda, K.; Li, W.; Mizuno-Horikawa, Y.; Nakano, M.; Asahi, M.; Takahashi, M.; et al. Dysregulation of TGF-β1 receptor activation leads to abnormal lung development and emphysema-like phenotype in core fucose-deficient mice. Proc. Natl. Acad. Sci. USA 2005, 102, 15791–15796. [Google Scholar] [CrossRef]
- Fukuda, T.; Hashimoto, H.; Okayasu, N.; Kameyama, A.; Onogi, H.; Nakagawasai, O.; Nakazawa, T.; Kurosawa, T.; Hao, Y.; Isaji, T.; et al. α1,6-Fucosyltransferase-deficient Mice Exhibit Multiple Behavioral Abnormalities Associated with a Schizophrenia-like Phenotype. J. Biol. Chem. 2011, 286, 18434–18443. [Google Scholar] [CrossRef]
- Fukuda, T.; Gu, W.; Isaji, T.; Taniguchi, N.; Gu, J. Mice lacking fucosyltransferase 8 exhibit abnormal behavioral abnormalities associated with a schizophrenia-like phenotype. Glycoconj. J. 2013, 30, 441. [Google Scholar]
- Lauc, G.; Rudan, I.; Campbell, H.; Rudd, P.M. Complex genetic regulation of protein glycosylation. Mol. Biosyst. 2010, 6, 329–335. [Google Scholar] [CrossRef]
- Neelamegham, S.; Mahal, L.K. Multi-level regulation of cellular glycosylation: From genes to transcript to enzyme to structure. Curr. Opin. Struct. Biol. 2016, 40, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Zaytseva, O.O.; Freidin, M.B.; Keser, T.; Štambuk, J.; Ugrina, I.; Šimurina, M.; Vilaj, M.; Štambuk, T.; Trbojević-Akmačić, I.; Pučić-Baković, M.; et al. Heritability of Human Plasma N-Glycome. J. Proteome Res. 2020, 19, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Knežević, A.; Polašek, O.; Gornik, O.; Rudan, I.; Campbell, H.; Hayward, C.; Wright, A.; Kolčić, I.; O’Donoghue, N.; Bones, J.; et al. Variability, heritability and environmental determinants of human plasma n-glycome. J. Proteome Res. 2009, 8, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Lauc, G.; Essafi, A.; Huffman, J.E.; Hayward, C.; Knežević, A.; Kattla, J.J.; Polašek, O.; Gornik, O.; Vitart, V.; Abrahams, J.L.; et al. Genomics meets glycomics-the first gwas study of human N-glycome identifies HNF1A as a master regulator of plasma protein fucosylation. PLoS Genet. 2010, 6, e1001256. [Google Scholar] [CrossRef] [PubMed]
- Huffman, J.E.; Knežević, A.; Vitart, V.; Kattla, J.; Adamczyk, B.; Novokmet, M.; Igl, W.; Pučić, M.; Zgaga, L.; Johannson, Å.; et al. Polymorphisms in B3GAT1, SLC9A9 and MGAT5 are associated with variation within the human plasma N-glycome of 3533 European adults. Hum. Mol. Genet. 2011, 20, 5000–5011. [Google Scholar] [CrossRef] [PubMed]
- Sharapov, S.Z.; Tsepilov, Y.A.; Klaric, L.; Mangino, M.; Thareja, G.; Shadrina, A.S.; Simurina, M.; Dagostino, C.; Dmitrieva, J.; Vilaj, M.; et al. Defining the genetic control of human blood plasma N-glycome using genome-wide association study. Hum. Mol. Genet. 2019, 28, 2062–2077. [Google Scholar] [CrossRef]
- Tudor, L.; Konjevod, M.; Nedic Erjavec, G.; Nikolac Perkovic, M.; Uzun, S.; Kozumplik, O.; Zoldos, V.; Lauc, G.; Svob Strac, D.; Pivac, N. Genetic and Epigenetic Association of Hepatocyte Nuclear Factor-1α with Glycosylation in Post-Traumatic Stress Disorder. Genes 2022, 13, 1063. [Google Scholar] [CrossRef]
- Zoldoš, V.; Horvat, T.; Novokmet, M.; Cuenin, C.; Mužinić, A.; Pučić, M.; Hufman, J.E.; Gornik, O.; Polašek, O.; Campbell, H.; et al. Epigenetic silencing of HNF1A associates with changes in the composition of the human plasma N-glycome. Epigenetics 2012, 7, 164–172. [Google Scholar] [CrossRef]
- Phan, L.; Jin, Y.; Zhang, H.; Qiang, W.; Shektman, E.; Shao, D.; Revoe, D.; Villamarin, R.; Ivanchenko, E.; Kimura, M.; et al. ALFA: Allele Frequency Aggregator; National Center for Biotechnology Information, US National Library of Medicine: Bethesda, MD, USA, 2020. Available online: www.ncbi.nlm.nih.gov/snp/docs/gsr/alfa/ (accessed on 25 January 2023).
- Vanhooren, V.; Desmyter, L.; Liu, X.-E.; Cardelli, M.; Franceschi, C.; Federico, A.; Libert, C.; Laroy, W.; Dewaele, S.; Contreras, R.; et al. N-Glycomic Changes in Serum Proteins During Human Aging. Rejuvenation Res. 2007, 10, 521–531a. [Google Scholar] [CrossRef]
- Vanhooren, V.; Dewaele, S.; Libert, C.; Engelborghs, S.; Paul, P.; Deyn, D.; Toussaint, O.; Debacq-chainiaux, F.; Poulain, M.; Glupczynski, Y.; et al. Serum N-glycan profile shift during human ageing. Exp. Geronotol. 2010, 45, 738–743. [Google Scholar] [CrossRef]
- Clerc, F.; Reiding, K.R.; Jansen, B.C.; Kammeijer, G.S.M.; Bondt, A.; Wuhrer, M. Human plasma protein N-glycosylation. Glycoconj. J. 2016, 33, 309–343. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, G.; Bentlage, A.E.H.; Plomp, R.; Visser, R.; Koeleman, C.A.M.; Beentjes, A.; Mok, J.Y.; van Esch, W.J.E.; Wuhrer, M.; Rispens, T.; et al. Conserved FcγR- glycan discriminates between fucosylated and afucosylated IgG in humans and mice. Mol. Immunol. 2018, 94, 54–60. [Google Scholar] [CrossRef] [PubMed]
- Niwa, R.; Hatanaka, S.; Shoji-Hosaka, E.; Sakurada, M.; Kobayashi, Y.; Uehara, A.; Yokoi, H.; Nakamura, K.; Shitara, K. Enhancement of the antibody-dependent cellular cytotoxicity of low-fucose IgG1 is independent of FcγRIIIa functional polymorphism. Clin. Cancer Res. 2004, 10, 6248–6255. [Google Scholar] [CrossRef] [PubMed]
- Masuda, K.; Kubota, T.; Kaneko, E.; Iida, S.; Wakitani, M.; Kobayashi-Natsume, Y.; Kubota, A.; Shitara, K.; Nakamura, K. Enhanced binding affinity for FcγRIIIa of fucose-negative antibody is sufficient to induce maximal antibody-dependent cellular cytotoxicity. Mol. Immunol. 2007, 44, 3122–3131. [Google Scholar] [CrossRef]
- Gudelj, I.; Baciarello, M.; Ugrina, I.; De Gregori, M.; Napolioni, V.; Ingelmo, P.M.; Bugada, D.; De Gregori, S.; Derek, L.; Pučić-Baković, M.; et al. Changes in total plasma and serum N-glycome composition and patient-controlled analgesia after major abdominal surgery. Sci. Rep. 2016, 6, 31234. [Google Scholar] [CrossRef]
- Dotz, V.; Lemmers, R.F.H.; Reiding, K.R.; Hipgrave Ederveen, A.L.; Lieverse, A.G.; Mulder, M.T.; Sijbrands, E.J.G.; Wuhrer, M.; van Hoek, M. Plasma protein N-glycan signatures of type 2 diabetes. Biochim. Biophys. Acta-Gen. Subj. 2018, 1862, 2613–2622. [Google Scholar] [CrossRef]
- Ikeda, M.; Takahashi, A.; Kamatani, Y.; Momozawa, Y.; Saito, T.; Kondo, K.; Shimasaki, A.; Kawase, K.; Sakusabe, T.; Iwayama, Y.; et al. Genome-wide association study detected novel susceptibility genes for schizophrenia and shared trans-populations/diseases genetic effect. Schizophr. Bull. 2019, 45, 824–834. [Google Scholar] [CrossRef]
- Li, Z.; Chen, J.; Yu, H.; He, L.; Xu, Y.; Zhang, D.; Yi, Q.; Li, C.; Li, X.; Shen, J.; et al. Genome-wide association analysis identifies 30 new susceptibility loci for schizophrenia. Nat. Genet. 2017, 49, 824–834. [Google Scholar] [CrossRef]
- Clements, C.C.; Karlsson, R.; Lu, Y.; Juréus, A.; Rück, C.; Andersson, E.; Boberg, J.; Pedersen, N.L.; Bulik, C.M.; Nordenskjöld, A.; et al. Genome-wide association study of patients with a severe major depressive episode treated with electroconvulsive therapy. Mol. Psychiatry 2021, 26, 2429–2439. [Google Scholar] [CrossRef]
- Sherva, R.; Gross, A.; Mukherjee, S.; Koesterer, R.; Amouyel, P.; Bellenguez, C.; Dufouil, C.; Bennett, D.A.; Chibnik, L.; Cruchaga, C.; et al. Genome-wide association study of rate of cognitive decline in Alzheimer’s disease patients identifies novel genes and pathways. Alzheimer’s Dement. 2020, 16, 1134–1145. [Google Scholar] [CrossRef]
- Baranzini, S.E.; Srinivasan, R.; Khankhanian, P.; Okuda, D.T.; Nelson, S.J.; Matthews, P.M.; Hauser, S.L.; Oksenberg, J.R.; Pelletier, D. Genetic variation influences glutamate concentrations in brains of patients with multiple sclerosis. Brain 2010, 133, 2603–2611. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.J.; Peloso, G.M.; Yu, H.; Butterworth, A.S.; Wang, X.; Mahajan, A.; Saleheen, D.; Emdin, C.; Alam, D.; Alves, A.C.; et al. Exome-wide association study of plasma lipids in >300,000 individuals. Nat. Genet. 2017, 49, 1758–1766. [Google Scholar] [CrossRef] [PubMed]
- Graham, S.E.; Clarke, S.L.; Wu, K.H.H.; Kanoni, S.; Zajac, G.J.M.; Ramdas, S.; Surakka, I.; Ntalla, I.; Vedantam, S.; Winkler, T.W.; et al. The power of genetic diversity in genome-wide association studies of lipids. Nature 2021, 600, 675–679. [Google Scholar] [CrossRef]
- Comuzzie, A.G.; Cole, S.A.; Laston, S.L.; Voruganti, V.S.; Haack, K.; Gibbs, R.A.; Butte, N.F. Novel Genetic Loci Identified for the Pathophysiology of Childhood Obesity in the Hispanic Population. PLoS ONE 2012, 7, e51954. [Google Scholar] [CrossRef]
- Zhu, Z.; Wang, X.; Li, X.; Lin, Y.; Shen, S.; Liu, C.L.; Hobbs, B.D.; Hasegawa, K.; Liang, L.; Boezen, H.M.; et al. Genetic overlap of chronic obstructive pulmonary disease and cardiovascular disease-related traits: A large-scale genome-wide cross-trait analysis. Respir. Res. 2019, 20, 64. [Google Scholar] [CrossRef]
- Kim, D.K.; Cho, M.H.; Hersh, C.P.; Lomas, D.A.; Miller, B.E.; Kong, X.; Bakke, P.; Gulsvik, A.; Agustí, A.; Wouters, E.; et al. Genome-wide association analysis of blood biomarkers in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2012, 186, 1238–1247. [Google Scholar] [CrossRef] [PubMed]
- Blain, P.; Mucklow, J.; Rawlins, M.; Roberts, D.; Routledge, P.; Shand, D. Determinants of plasma alpha 1-acid glycoprotein (AAG) concentrations in health. Br. J. Clin. Pharmacol. 1985, 20, 500–502. [Google Scholar] [CrossRef]
- Williams, J.P.; Weiser, M.R.; Pechet, T.T.V.; Kobzik, L.; Moore, F.D.; Hechtman, H.B. α1-acid glycoprotein reduces local and remote injuries after intestinal ischemia in the rat. Am. J. Physiol.-Gastrointest. Liver Physiol. 1997, 273, G1031–G1035. [Google Scholar] [CrossRef]
- Liu, X.-E.; Desmyter, L.; Gao, C.-F.; Laroy, W.; Dewaele, S.; Vanhooren, V.; Wang, L.; Zhuang, H.; Callewaert, N.; Libert, C.; et al. N-glycomic changes in hepatocellular carcinoma patients with liver cirrhosis induced by hepatitis B virus. Hepatology 2007, 46, 1426–1435. [Google Scholar] [CrossRef]
- Higai, K.; Aoki, Y.; Azuma, Y.; Matsumoto, K. Glycosylation of site-specific glycans of α1-acid glycoprotein and alterations in acute and chronic inflammation. Biochim. Biophys. Acta-Gen. Subj. 2005, 1725, 128–135. [Google Scholar] [CrossRef]
- Menni, C.; Keser, T.; Mangino, M.; Bell, J.T.; Erte, I.; Akmačić, I.; Baković, M.P.; Gornik, O.; Mccarthy, M.I.; Zoldoš, V.; et al. Glycosylation of Immunoglobulin G: Role of Genetic and Epigenetic Influences. PLoS ONE 2013, 8, e82558. [Google Scholar] [CrossRef] [PubMed]
- Heydarzadeh, S.; Ranjbar, M.; Karimi, F.; Seif, F.; Alivand, M.R. Overview of host miRNA properties and their association with epigenetics, long non-coding RNAs, and Xeno-infectious factors. Cell Biosci. 2021, 11, 43. [Google Scholar] [CrossRef]
- Breen, L.D.; Pučić-Baković, M.; Vučković, F.; Reiding, K.; Trbojević-Akmačić, I.; Gajdošik, M.Š.; Cook, M.I.; Lopez, M.J.; Wuhrer, M.; Camara, L.M.; et al. IgG and IgM glycosylation patterns in patients undergoing image-guided tumor ablation. Biochim. Biophys. Acta-Gen. Subj. 2016, 1860, 1786–1794. [Google Scholar] [CrossRef] [PubMed]
- Preissner, K.T.; Reuning, U. Vitronectin in vascular context: Facets of a multitalented matricellular protein. Semin. Thromb. Hemost. 2011, 37, 408–424. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.; Lee, J.Y.; Lee, H.K.; Park, G.W.; Jeong, H.K.; Moon, M.H.; Kim, J.Y.; Yoo, J.S. In-depth analysis of site-specific N-glycosylation in vitronectin from human plasma by tandem mass spectrometry with immunoprecipitation. Anal. Bioanal. Chem. 2014, 406, 7999–8011. [Google Scholar] [CrossRef]
- Quadrelli, S.; Tosh, N.; Urquhart, A.; Trickey, K.; Tremewan, R.; Galloway, G.; Rich, L.; Lea, R.; Malycha, P.; Mountford, C. Post-traumatic stress disorder affects fucose-α(1–2)-glycans in the human brain: Preliminary findings of neuro deregulation using in vivo two-dimensional neuro MR spectroscopy. Transl. Psychiatry 2019, 9, 27. [Google Scholar] [CrossRef] [PubMed]
- Sawa, M.; Hsu, T.L.; Itoh, T.; Sugiyama, M.; Hanson, S.R.; Vogt, P.K.; Wong, C.H. Glycoproteomic probes for fluorescent imaging of fucosylated glycans in vivo. Proc. Natl. Acad. Sci. USA 2006, 103, 12371–12376. [Google Scholar] [CrossRef]
- Wibowo, A.; Peters, E.C.; Hsieh-Wilson, L.C. Photoactivatable glycopolymers for the proteome-wide identification of fucose-∝(1-2)-galactose binding proteins. J. Am. Chem. Soc. 2014, 136, 9528–9531. [Google Scholar] [CrossRef]
- Mellon, S.H.; Gautam, A.; Hammamieh, R.; Jett, M.; Wolkowitz, O.M. Metabolism, Metabolomics, and Inflammation in Posttraumatic Stress Disorder. Biol. Psychiatry 2018, 83, 866–875. [Google Scholar] [CrossRef]
- Konjevod, M.; Tudor, L.; Svob Strac, D.; Nedic Erjavec, G.; Barbas, C.; Zarkovic, N.; Nikolac Perkovic, M.; Uzun, S.; Kozumplik, O.; Lauc, G.; et al. Metabolomic and glycomic findings in posttraumatic stress disorder. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2019, 88, 181–193. [Google Scholar] [CrossRef]
- Raposo, C.D.; Canelas, A.B.; Barros, M.T. Human lectins, their carbohydrate affinities and where to find them. Biomolecules 2021, 11, 188. [Google Scholar] [CrossRef] [PubMed]
- Doran, R.C.; Tatsuno, G.P.; O’Rourke, S.M.; Yu, B.; Alexander, D.L.; Mesa, K.A.; Berman, P.W. Glycan modifications to the gp120 immunogens used in the RV144 vaccine trial improve binding to broadly neutralizing antibodies. PLoS ONE 2018, 13, e0196370. [Google Scholar] [CrossRef] [PubMed]
- Yates, N.L.; deCamp, A.C.; Korber, B.T.; Liao, H.-X.; Irene, C.; Pinter, A.; Peacock, J.; Harris, L.J.; Sawant, S.; Hraber, P.; et al. HIV-1 Envelope Glycoproteins from Diverse Clades Differentiate Antibody Responses and Durability among Vaccinees. J. Virol. 2018, 92, e01843-17. [Google Scholar] [CrossRef] [PubMed]
- Anthony, R.M.; Ravetch, J.V. A novel role for the IgG Fc glycan: The anti-inflammatory activity of sialylated IgG Fcs. J. Clin. Immunol. 2010, 30, 9–14. [Google Scholar] [CrossRef]
- Yu, X.; Marshall, M.J.E.; Cragg, M.S.; Crispin, M. Improving Antibody-Based Cancer Therapeutics Through Glycan Engineering. BioDrugs 2017, 31, 151–166. [Google Scholar] [CrossRef]
- Barth, C.; Villringer, A.; Sacher, J. Sex hormones affect neurotransmitters and shape the adult female brain during hormonal transition periods. Front. Neurosci. 2015, 9, 37. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Publishing: Washington, DC, USA, 2013; ISBN 9780890425541. [Google Scholar]
- Weathers, F.W.; Keane, T.M.; Davidson, J.R.T. Clinician-administered PTSD scale: A review of the first ten years of research. Depress. Anxiety 2001, 13, 132–156. [Google Scholar] [CrossRef]
- Nedic Erjavec, G.; Bektic Hodzic, J.; Repovecki, S.; Nikolac Perkovic, M.; Uzun, S.; Kozumplik, O.; Tudor, L.; Mimica, N.; Svob Strac, D.; Pivac, N. Alcohol-related phenotypes and platelet serotonin concentration. Alcohol 2021, 97, 41–49. [Google Scholar] [CrossRef]
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef]
- Trbojević Akmačić, I.; Ugrina, I.; Štambuk, J.; Gudelj, I.; Vučković, F.; Lauc, G.; Pučić-Baković, M. High-throughput glycomics: Optimization of sample preparation. Biochemistry 2015, 80, 934–942. [Google Scholar] [CrossRef]
- Saldova, R.; Asadi Shehni, A.; Haakensen, V.D.; Steinfeld, I.; Hilliard, M.; Kifer, I.; Helland, Å.; Yakhini, Z.; Børresen-Dale, A.L.; Rudd, P.M. Association of N-glycosylation with breast carcinoma and systemic features using high-resolution quantitative UPLC. J. Proteome Res. 2014, 13, 2314–2327. [Google Scholar] [CrossRef] [PubMed]
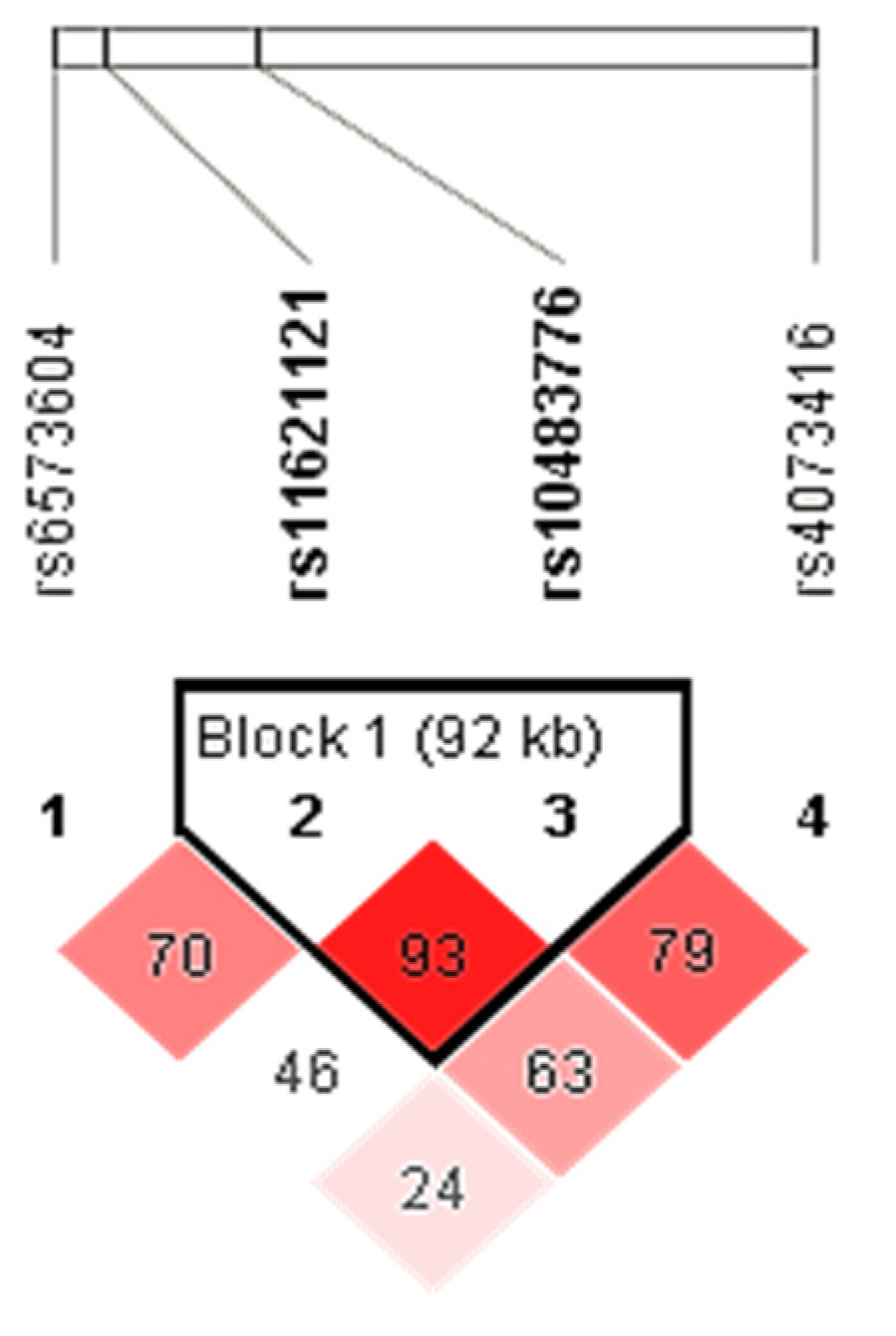
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2005, 21, 263–265. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, S.B.; Schaffner, S.F.; Nguyen, H.; Moore, J.M.; Roy, J.; Blumenstiel, B.; Higgins, J.; DeFelice, M.; Lochner, A.; Faggart, M.; et al. The structure of haplotype blocks in the human genome. Science 2002, 296, 2225–2229. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; De Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A tool set for whole-genome association and population-based linkage analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- Unwin, A. Discovering Statistics Using R by Andy Field, Jeremy Miles, Zoë Field. Int. Stat. Rev. 2013, 81, 169–170. [Google Scholar] [CrossRef]
- Pučić, M.; Mužinić, A.; Novokmet, M.; Škledar, M.; Pivac, N.; Lauc, G.; Gornik, O. Changes in plasma and IgG N-glycome during childhood and adolescence. Glycobiology 2012, 22, 975–982. [Google Scholar] [CrossRef] [PubMed]
- Konjevod, M.; Nedic Erjavec, G.; Nikolac Perkovic, M.; Sáiz, J.; Tudor, L.; Uzun, S.; Kozumplik, O.; Svob Strac, D.; Zarkovic, N.; Pivac, N. Metabolomics in posttraumatic stress disorder: Untargeted metabolomic analysis of plasma samples from Croatian war veterans. Free Radic. Biol. Med. 2021, 162, 636–641. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| SNP | Position (bp) | Associated Gene | MAF (EU) | MAF | HWE |
|---|---|---|---|---|---|
| rs6573604 | 14:65321223 | PTBP1P, MIR4708 | C = 0.185 | C = 0.190 | p = 0.003 |
| rs11621121 | 14:65355775 | MIR4708, FUT8 | C = 0.410 | C = 0.440 | p = 0.467 |
| rs10483776 | 14:65448149 | FUT8 | G = 0.176 | G = 0.207 | p = 0.487 |
| rs4073416 | 14:65792676 | NCOA4P1, FUT8 | C = 0.419 | C = 0.416 | p = 0.516 |
| SNP | Genotype/Allele | Control Participants | Patients with PTSD | Statistics | ||
|---|---|---|---|---|---|---|
| N | % | N | % | |||
| rs6573604 | CC | 20 | 8.1% | 11 | 3.7% | χ2 = 5.861; df = 2; p = 0.053 |
| CT | 69 | 28.1% | 75 | 25.4% | ||
| TT | 157 | 63.8% | 209 | 70.8% | ||
| C | 109 | 22.2% | 97 | 16.4% | χ2 = 5.682; df = 1; p = 0.017 | |
| T | 383 | 77.8% | 493 | 83.6% | ||
| rs11621121 | CC | 51 | 20.7% | 58 | 19.7% | χ2 = 2.571; df = 2; p = 0.277 |
| CT | 124 | 50.4% | 133 | 45.1% | ||
| TT | 71 | 28.9% | 104 | 35.3% | ||
| C | 226 | 45.9% | 249 | 42.2% | χ2 = 1.517; df = 1; p = 0.218 | |
| T | 266 | 54.1% | 341 | 57.8% | ||
| rs10483776 | AA | 160 | 65.0% | 179 | 60.7% | χ2 = 2.473; df = 2; p = 0.290 |
| AG | 75 | 30.5% | 107 | 36.3% | ||
| GG | 11 | 4.5% | 9 | 3.1% | ||
| A | 395 | 80.3% | 465 | 78.8% | χ2 = 0.356; df = 1; p = 0.551 | |
| G | 97 | 19.7% | 125 | 21.2% | ||
| rs4073416 | CC | 46 | 18.7% | 43 | 14.6% | χ2 = 2.958; df = 2; p = 0.228 |
| CT | 114 | 46.3% | 157 | 53.2% | ||
| TT | 86 | 35.0% | 95 | 32.2% | ||
| C | 206 | 41.9% | 243 | 41.2% | χ2 = 0.052; df = 1; p = 0.820 | |
| T | 286 | 58.1% | 347 | 58.8% | ||
| Haplotype rs11621121–rs10483776 | All Participants | Control Participants | Patients with PTSD | |||
|---|---|---|---|---|---|---|
| N | % | N | % | N | % | |
| TA | 601 | 55.5% | 264 | 53.5% | 337 | 57.1% |
| CA | 259 | 23.9% | 131 | 26.6% | 128 | 21.7% |
| CG | 216 | 20.0% | 95 | 19.3% | 121 | 20.5% |
| TG | 6 | 0.5% | 2 | 0.4% | 4 | 0.7% |
| Statistics | χ2 = 3.853; df = 3; p = 0.278 | |||||
| Glycan Peak | Model | rs6573604 | rs11621121 | rs10483776 | rs4073416 | Haplo-Type | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| H/U * | p | H/U * | p | H/U * | p | H/U * | p | |||
| GP08 (A2G2) | Genetic | 0.06 | 0.999 | 9.06 | 0.215 | 2.95 | 0.593 | 10.26 | 0.117 | H = 26.75; p = 0.101 |
| Allelic | 20754.5 | 0.978 | 25454.0 | 0.059 | 17330.5 | 0.333 | 24352.0 | 0.020 | ||
| GP14 (A2G2S1) | Genetic | 0.62 | 0.893 | 4.61 | 0.650 | 5.24 | 0.475 | 9.85 | 0.091 | H = 21.39; p = 0.351 |
| Allelic | 19956.5 | 0.726 | 26820.0 | 0.254 | 16446.5 | 0.202 | 24472 | 0.013 | ||
| GP22 (FA2G2S2) | Genetic | 1.56 | 0.744 | 21.00 | 0.001 | 22.05 | <0.001 | 14.61 | 0.039 | H = 31.41; p < 0.001 |
| Allelic | 19856.5 | 0.710 | 23408.0 | 0.001 | 13506.5 | <0.001 | 23952 | 0.016 | ||
| GP29 (FA3G3S3) | Genetic | 3.99 | 0.530 | 4.07 | 0.464 | 12.85 | 0.020 | 6.99 | 0.234 | H = 28.37; p = 0.020 |
| Allelic | 18026.5 | 0.167 | 27664.0 | 0.333 | 14608.5 | 0.003 | 25248 | 0.068 | ||
| GP31 (FA3G3S3) | Genetic | 3.94 | 0.493 | 8.69 | 0.169 | 24.36 | <0.001 | 1.29 | 0.757 | H = 41.19; p < 0.001 |
| Allelic | 18042.5 | 0.151 | 25588.0 | 0.052 | 13052.5 | <0.001 | 27646 | 0.397 | ||
| GP34 (A4G4S3) | Genetic | 6.66 | 0.281 | 2.57 | 0.540 | 13.53 | 0.013 | 1.38 | 0.755 | H = 14.39; p = 0.013 |
| Allelic | 17340.5 | 0.068 | 27578.0 | 0.345 | 14452.5 | 0.002 | 28230 | 0.559 | ||
| GP36 (A4G4S4) | Genetic | 15.13 | 0.020 | 4.58 | 0.563 | 1.44 | 0.702 | 0.41 | 0.881 | H = 23.30; p = 0.256 |
| Allelic | 16010.5 | 0.003 | 26752.0 | 0.341 | 18606.5 | 0.781 | 28718 | 0.706 | ||
| GP37 (A4G4S4) | Genetic | 15.14 | 0.013 | 0.10 | 0.978 | 3.50 | 0.522 | 0.18 | 0.912 | H = 35.03; p = 0.316 |
| Allelic | 15514.5 | 0.001 | 29586.0 | 0.876 | 16828.5 | 0.273 | 28868 | 0.743 | ||
| GP38 (A4G4S4) | Genetic | 17.40 | 0.007 | 0.85 | 0.728 | 1.62 | 0.666 | 0.23 | 0.914 | H = 33.42; p = 0.440 |
| Allelic | 15342.5 | 0.001 | 28706.0 | 0.585 | 17988.5 | 0.622 | 29370 | 0.980 | ||
| Glycan Peak | Model | rs6573604 | rs11621121 | rs10483776 | rs4073416 | Haplo-Type | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| H/U * | p | H/U * | p | H/U * | p | H/U * | p | |||
| GP16 (FA2G2S1) | Genetic | 0.18 | 0.991 | 11.51 | 0.039 | 15.50 | 0.017 | 6.85 | 0.322 | H = 21.10; p = 0.029 |
| Allelic | 23364.5 | 0.908 | 35666.5 | 0.020 | 24838.5 | 0.169 | 37340.5 | 0.176 | ||
| GP20 (A2G2S2) | Genetic | 2.82 | 0.683 | 7.82 | 0.156 | 6.19 | 0.160 | 3.92 | 0.687 | H = 20.13; p = 0.109 |
| Allelic | 22042.5 | 0.794 | 36544.5 | 0.039 | 26016.5 | 0.255 | 38474.5 | 0.341 | ||
| GP22 (FA2G2S2) | Genetic | 0.05 | 1.000 | 19.34 | 0.002 | 12.92 | 0.026 | 11.30 | 0.156 | H = 29.04; p = 0.002 |
| Allelic | 23552.5 | 0.936 | 33526.5 | 0.001 | 23454.5 | 0.039 | 36856.5 | 0.351 | ||
| GP23 (FA2BG2S2) | Genetic | 0.94 | 0.938 | 12.78 | 0.039 | 9.19 | 0.065 | 7.69 | 0.273 | H = 21.73; p = 0.026 |
| Allelic | 23606.5 | 0.865 | 35466.5 | 0.013 | 26536.5 | 0.351 | 37268.5 | 0.208 | ||
| GP31 (FA3G3S3) | Genetic | 2.34 | 0.674 | 6.57 | 0.160 | 13.75 | 0.020 | 7.99 | 0.351 | H = 20.77; p = 0.020 |
| Allelic | 22074.5 | 0.754 | 37016.5 | 0.045 | 23276.5 | 0.020 | 37588.5 | 0.195 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tudor, L.; Nedic Erjavec, G.; Nikolac Perkovic, M.; Konjevod, M.; Uzun, S.; Kozumplik, O.; Mimica, N.; Lauc, G.; Svob Strac, D.; Pivac, N. The Association of the Polymorphisms in the FUT8-Related Locus with the Plasma Glycosylation in Post-Traumatic Stress Disorder. Int. J. Mol. Sci. 2023, 24, 5706. https://doi.org/10.3390/ijms24065706
Tudor L, Nedic Erjavec G, Nikolac Perkovic M, Konjevod M, Uzun S, Kozumplik O, Mimica N, Lauc G, Svob Strac D, Pivac N. The Association of the Polymorphisms in the FUT8-Related Locus with the Plasma Glycosylation in Post-Traumatic Stress Disorder. International Journal of Molecular Sciences. 2023; 24(6):5706. https://doi.org/10.3390/ijms24065706
Chicago/Turabian StyleTudor, Lucija, Gordana Nedic Erjavec, Matea Nikolac Perkovic, Marcela Konjevod, Suzana Uzun, Oliver Kozumplik, Ninoslav Mimica, Gordan Lauc, Dubravka Svob Strac, and Nela Pivac. 2023. "The Association of the Polymorphisms in the FUT8-Related Locus with the Plasma Glycosylation in Post-Traumatic Stress Disorder" International Journal of Molecular Sciences 24, no. 6: 5706. https://doi.org/10.3390/ijms24065706
APA StyleTudor, L., Nedic Erjavec, G., Nikolac Perkovic, M., Konjevod, M., Uzun, S., Kozumplik, O., Mimica, N., Lauc, G., Svob Strac, D., & Pivac, N. (2023). The Association of the Polymorphisms in the FUT8-Related Locus with the Plasma Glycosylation in Post-Traumatic Stress Disorder. International Journal of Molecular Sciences, 24(6), 5706. https://doi.org/10.3390/ijms24065706