
Rhizosphere Fungal Dynamics in Sugarcane during Different Growth Stages
 , , ,
, , ,
Abstract
1. Introduction
2. Results
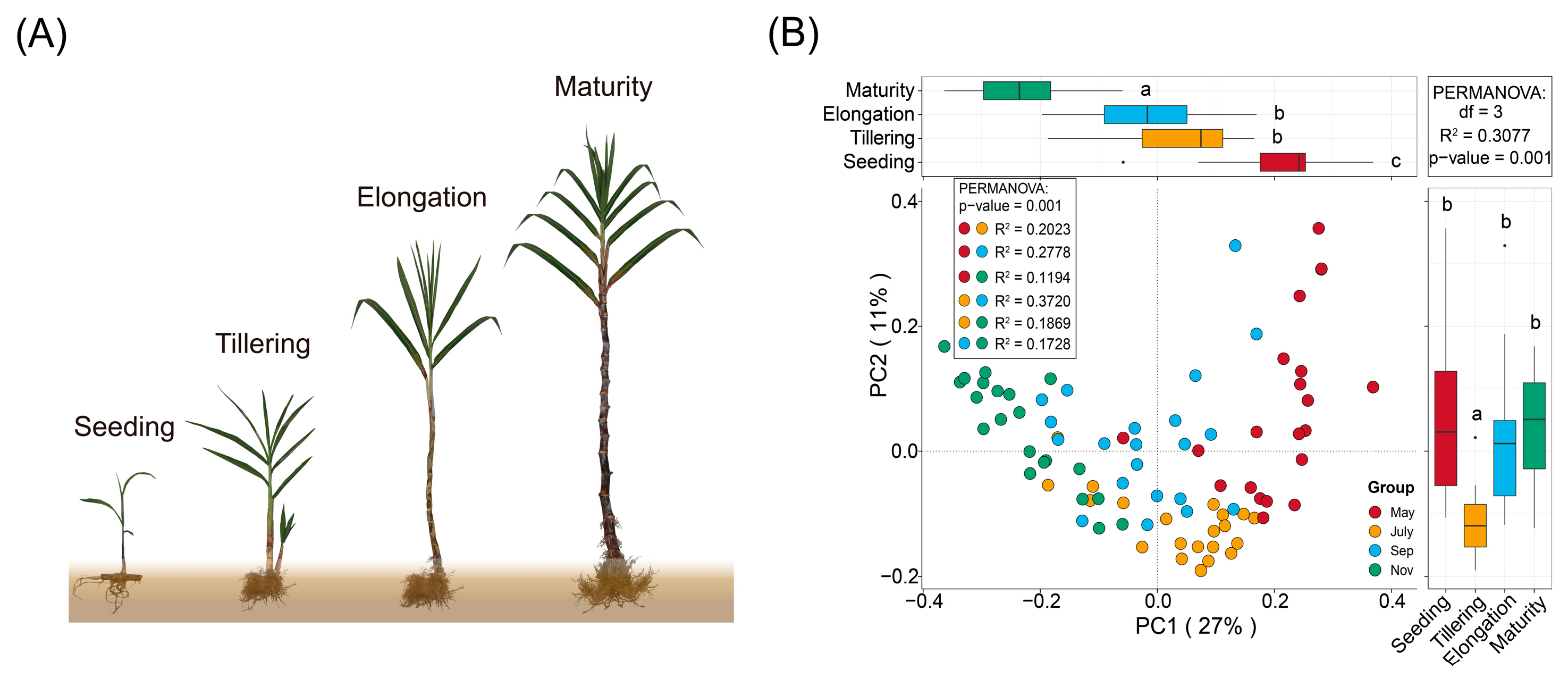
2.1. Sugarcane Rhizosphere Nutrients Produce Differential Changes at Different Stages
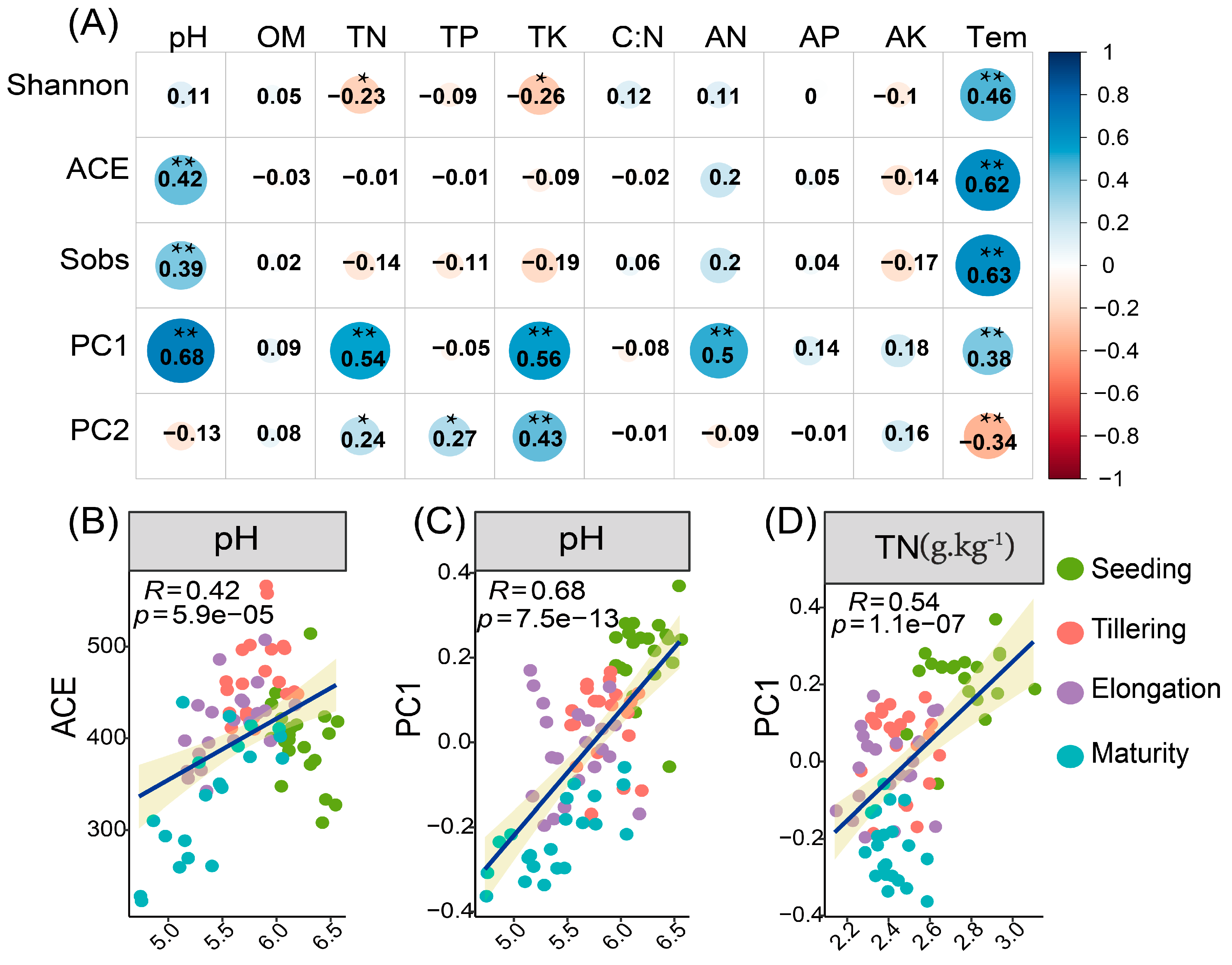
2.2. Rhizosphere Fungal Diversity Differs between Different Stages and Correlates with Soil Properties
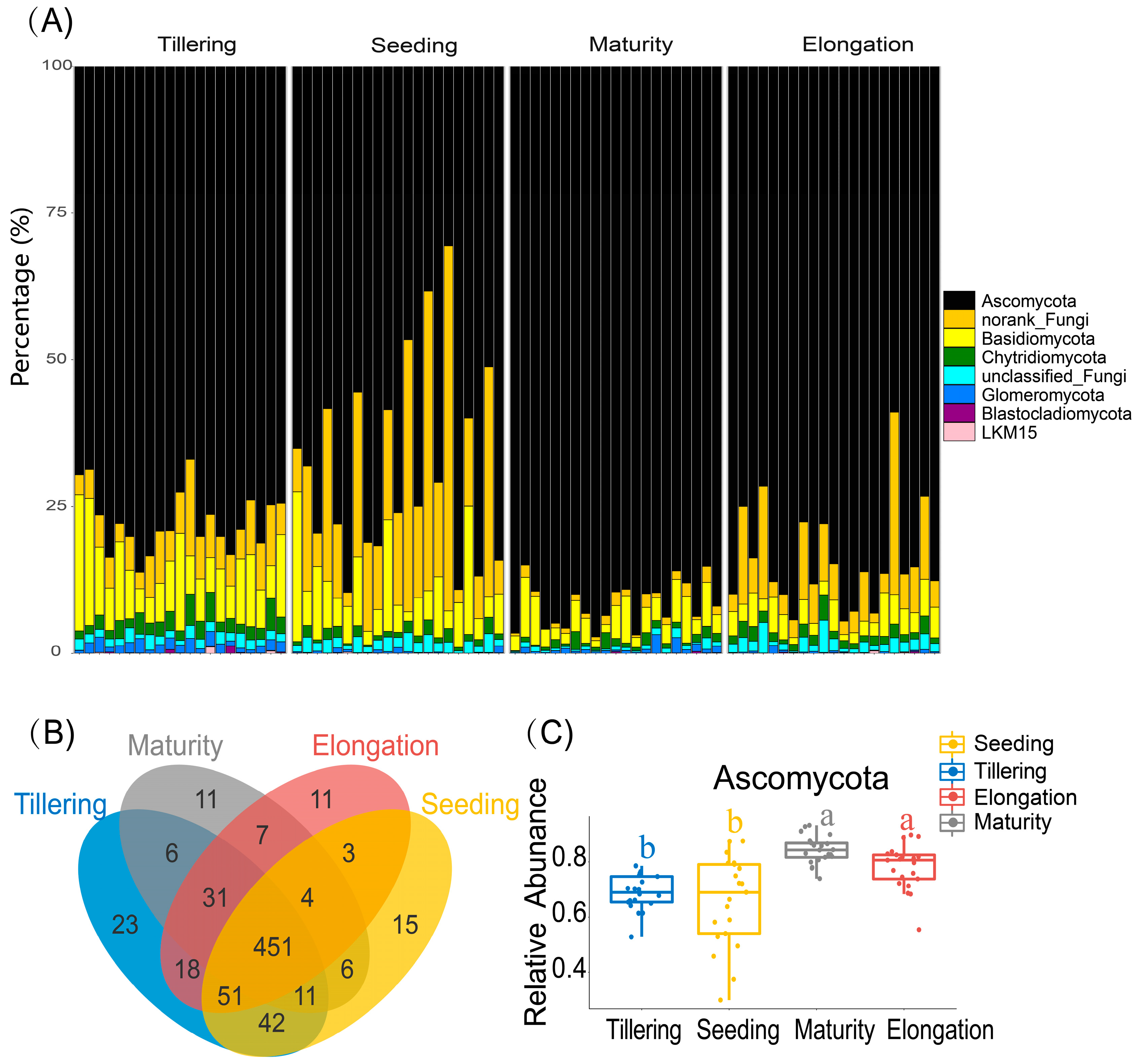
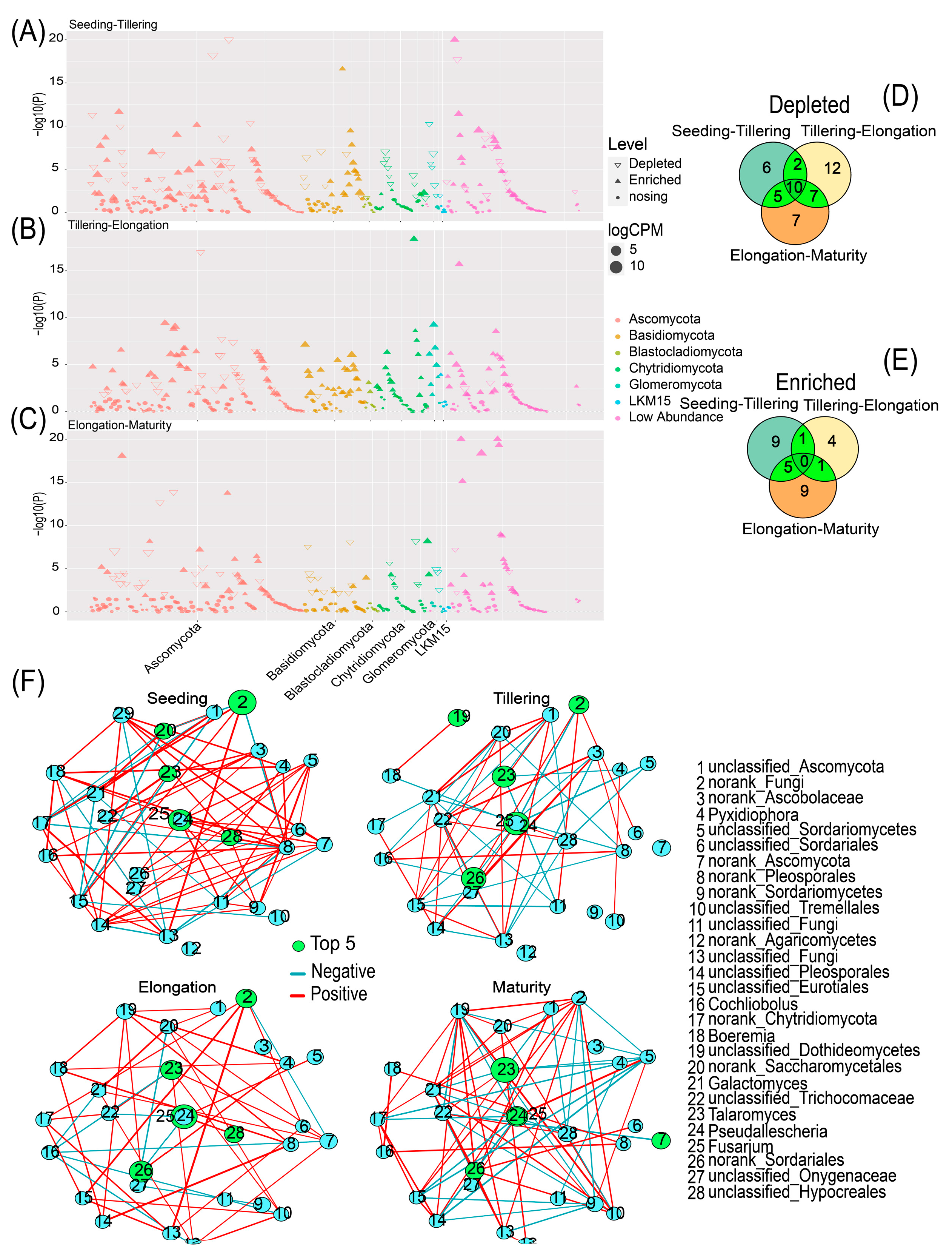
2.3. Variability in Fungal Community Composition at Different Growth Stages
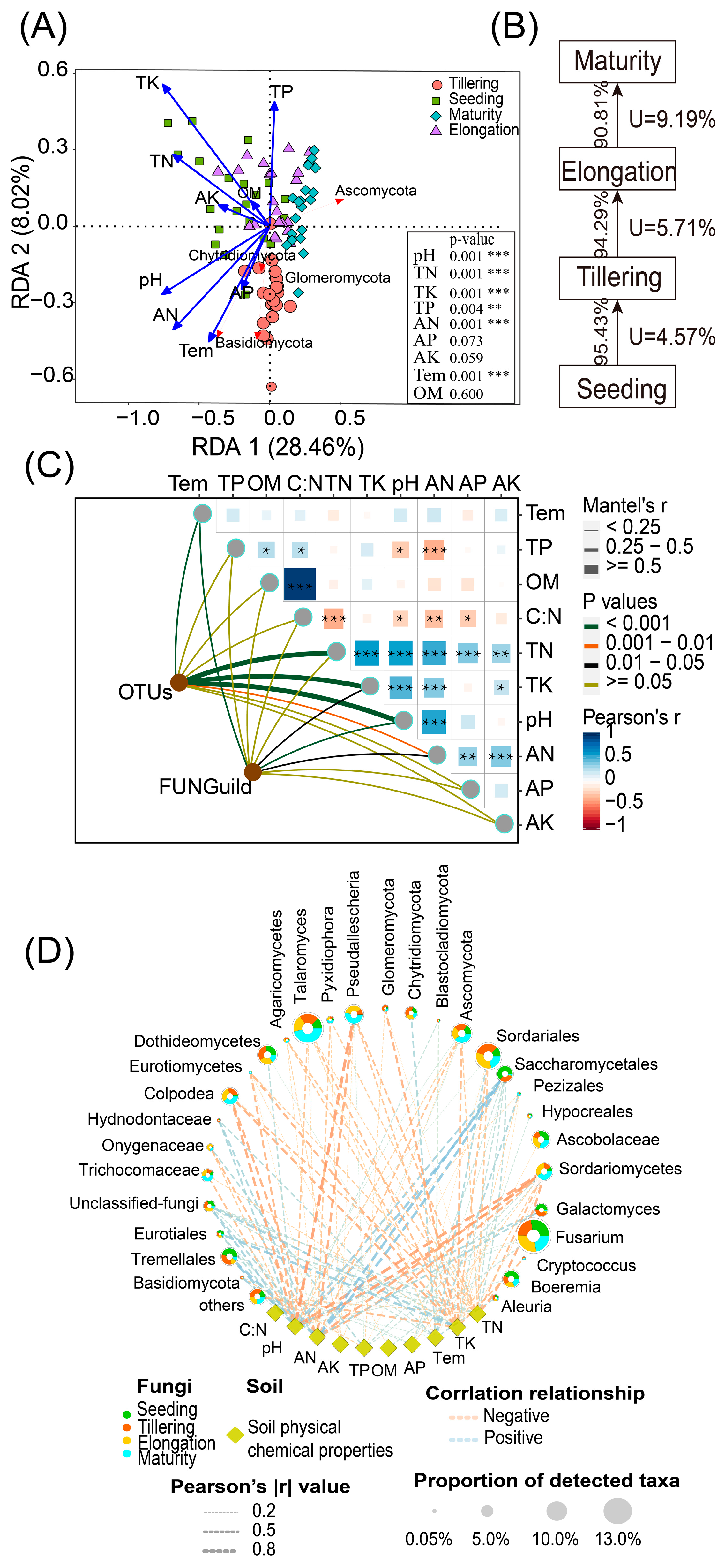
2.4. Fungal Community Composition, FUNGuild, and Soil Physicochemical Properties Are Interrelated
2.5. Fungal Communities Differ in Co-Occurrence Networks with Changes in Growth Periods
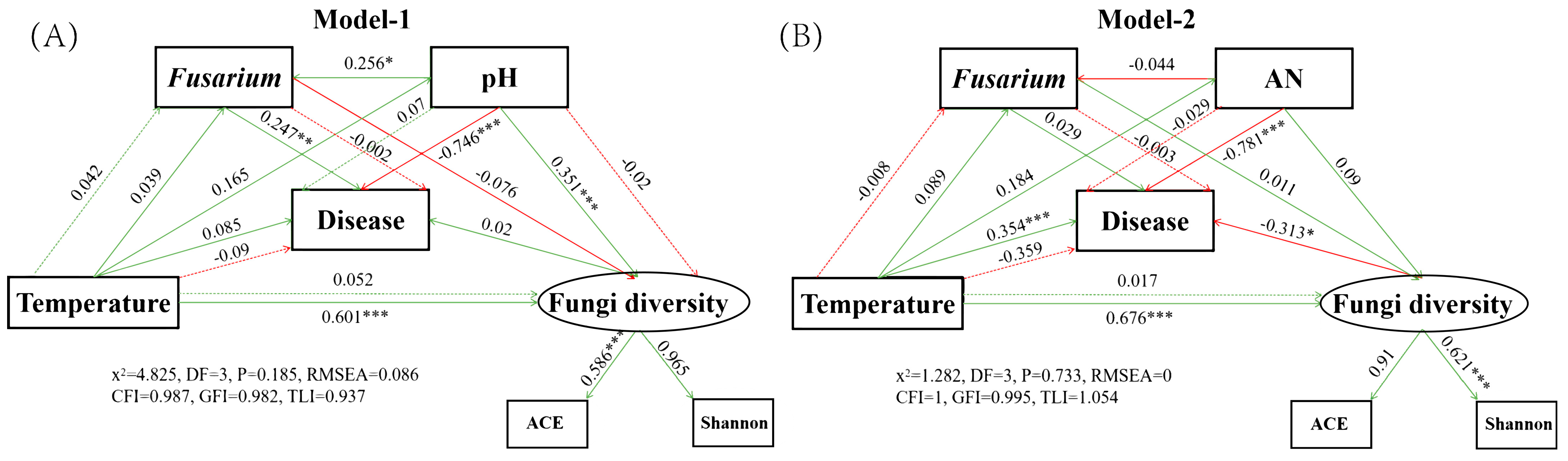
2.6. Construction of Structural Equation Models Related to Pokkah Boeng Disease of Sugarcane
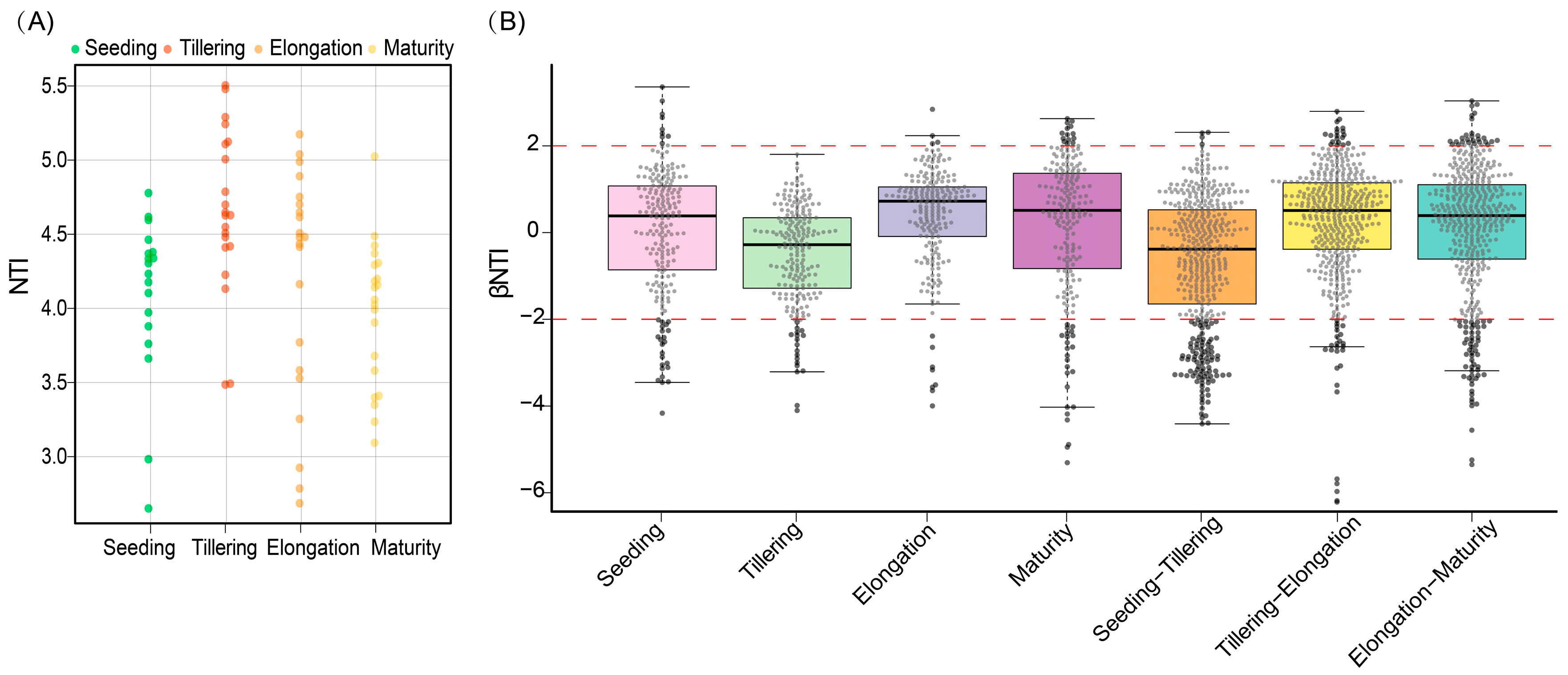
2.7. The Assembly Process of Fungal Communities Is Influenced by Changes in the Growth Period
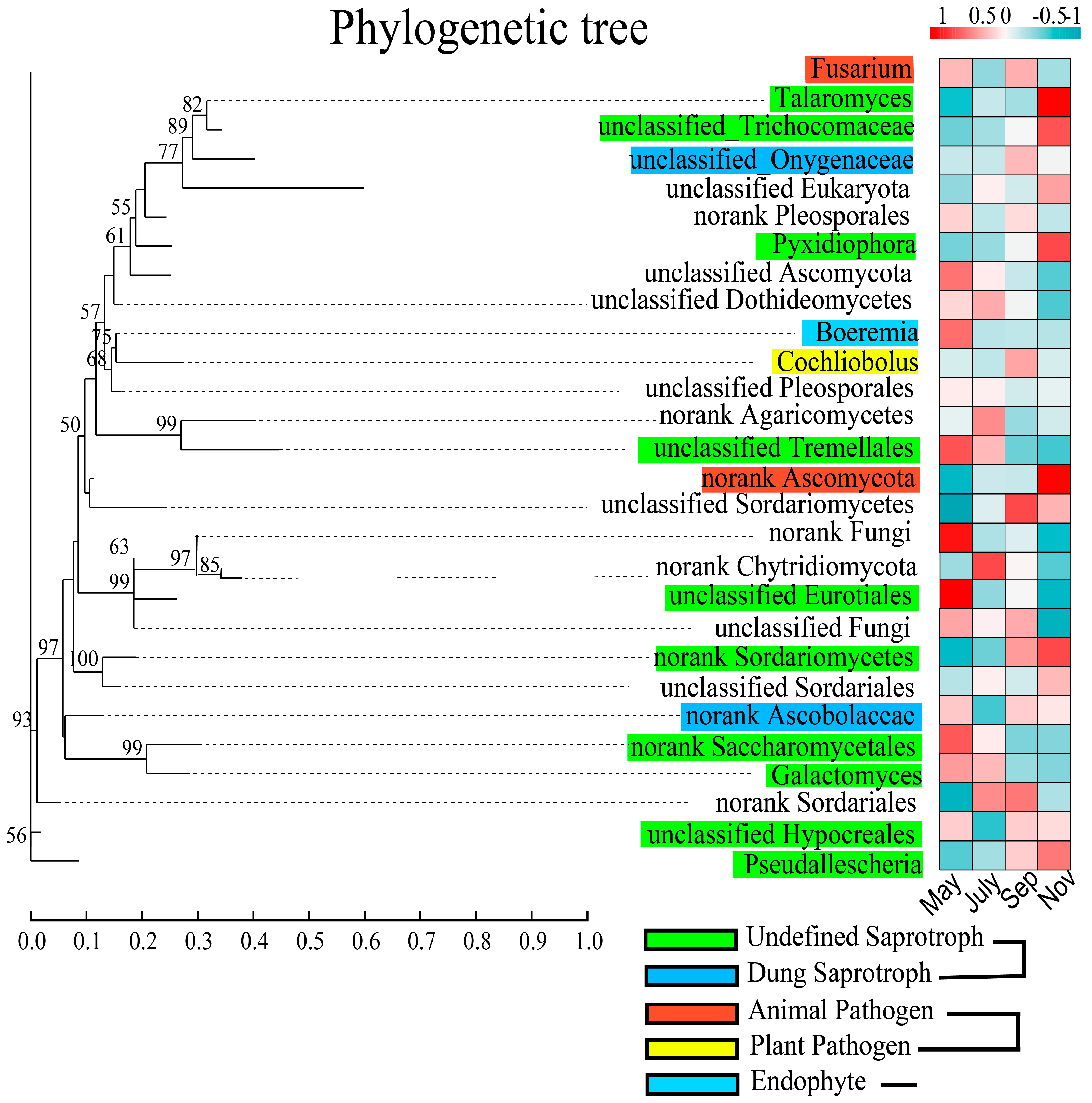
2.8. Functional and Evolutionary Relationships among the Major Fungal Genera of Rhizosphere Soil
3. Discussion
3.1. Changes in Sugarcane Growth Periods Alter the Composition and Diversity of Soil Properties and Fungal Communities
3.2. Fungal Taxonomy, Function, and Soil Traits Were Interrelated, and the Transfer of Fungal Communities between Changes in Sugarcane Growth Periods Showed Regularity
3.3. Structural Equation Modeling Demonstrates a Correlation between Key Fungi, Soil Properties, and Sugarcane Disease Rates
3.4. Rhizosphere Fungal Community Assembly Is Mainly Influenced by Stochastic Factors
4. Materials and Methods
4.1. Sugarcane Planting
4.2. Sample Collection
4.3. Determination of Pokkah Boeng Disease of Sugarcane and Soil Physio-Chemical Properties
4.4. DNA Extraction and PCR Amplification
4.5. Data Quality Control and Filtering
4.6. Sequences and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Breidenbach, B.; Pump, J.; Dumont, M.G. Microbial Community Structure in the Rhizosphere of Rice Plants. Front. Microbiol. 2016, 6, 1537. [Google Scholar] [CrossRef]
- Hilbig, B.E.; Allen, E.B. Plant-soil feedbacks and competitive interactions between invasive Bromus diandrus and native forb species. Plant Soil 2015, 392, 191–203. [Google Scholar] [CrossRef]
- Qiao, Y.; Miao, S.; Han, X.; Xing, B. Root and Microbial Contributions to Soil Respiration during a Soybean Growing Season. Commun. Soil Sci. Plant Anal. 2009, 40, 1658–1673. [Google Scholar] [CrossRef]
- Mijangos, I.; Becerril, J.M.; Albizu, I.; Epelde, L.; Garbisu, C. Effects of glyphosate on rhizosphere soil microbial communities under two different plant compositions by cultivation-dependent and-independent methodologies. Soil Biol. Biochem. 2009, 41, 505–513. [Google Scholar] [CrossRef]
- Broeckling, C.D.; Broz, A.K.; Bergelson, J.; Manter, D.K.; Vivanco, J.M. Root Exudates Regulate Soil Fungal Community Composition and Diversity. Appl. Environ. Microbiol. 2008, 74, 738–744. [Google Scholar] [CrossRef]
- Aoki, Y.; Fujita, K.; Shima, H.; Suzuki, S. Survey of Annual and Seasonal Fungal Communities in Japanese Prunus mume Orchard Soil by Next-Generation Sequencing. Adv. Microbiol. 2015, 5, 817–824. [Google Scholar] [CrossRef]
- Dong, C.; Wang, W.; Liu, H.; Xu, X.; Chen, X.; Zeng, H. Comparison of soil microbial responses to nitrogen addition between ex-arable grassland and natural grassland. J. Soils Sediments 2021, 21, 1371–1384. [Google Scholar] [CrossRef]
- Pandey, S.S.; Singh, S.; Babu, C.S.V.; Shanker, K.; Srivastava, N.K.; Kalra, A. Endophytes of opium poppy differentially modulate host plant productivity and genes for the biosynthetic pathway of benzylisoquinoline alkaloids. Planta 2016, 243, 1097–1114. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.; Wolverton, B. Plants+soil/wetland microbes: Food crop systems that also clean air and water. Adv. Space Res. 2011, 47, 582–590. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, N.; Liu, Y.-X.; Zhang, X.; Hu, B.; Qin, Y.; Xu, H.; Wang, H.; Guo, X.; Qian, J.; et al. Root microbiota shift in rice correlates with resident time in the field and developmental stage. Sci. China Life Sci. 2018, 61, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.L.; Han, F.; Xiao, Z.; Ren, M.B.; Hu, K.Z. Ariation of Rhizosphere Fungal of Gardenia at Different Growth Stage Based on High-Throughput Sequencing. Southwest Agric. J. 2020, 33, 2628–2634. [Google Scholar] [CrossRef]
- Prasara-A, J.; Gheewala, S.H. An assessment of social sustainability of sugarcane and cassava cultivation in Thailand. Sustain. Prod. Consum. 2021, 27, 372–382. [Google Scholar] [CrossRef]
- Lam, E.; Shine, J.; Da Silva, J.; Lawton, M.; Bonos, S.; Calvino, M.; Carrer, H.; Silva-Filho, M.; Glynn, N.; Helsel, Z.; et al. Improving sugarcane for biofuel: Engineering for an even better feedstock. GCB Bioenergy 2009, 1, 251–255. [Google Scholar] [CrossRef]
- Naylor, D.; Degraaf, S.; Purdom, E.; Coleman-Derr, D. Drought and host selection influence bacterial community dynamics in the grass root microbiome. ISME J. 2017, 11, 2691–2704. [Google Scholar] [CrossRef]
- Hamonts, K.; Trivedi, P.; Garg, A.; Janitz, C.; Grinyer, J.; Holford, P.; Botha, F.C.; Anderson, I.C.; Singh, B.K. Field study reveals core plant microbiota and relative importance of their drivers. Environ. Microbiol. 2017, 20, 124–140. [Google Scholar] [CrossRef] [PubMed]
- Da Vitória, E.L.; Rodrigues, J.P. Emergy efficiency analysis of sugarcane as a raw material for ethanol production. Rev. Ciência Agronômica 2016, 4, 47. [Google Scholar] [CrossRef]
- Singh, J.; Singh, R.D.; Anwar, S.I.; Solomon, S. Alternative Sweeteners Production from Sugarcane in India: Lump Sugar (Jaggery). Sugar Tech 2011, 13, 366–371. [Google Scholar] [CrossRef]
- Xie, L.; Long, Y.-Y.; Zhang, Y.; Chen, Y.-L.; Zhang, W.-L. Serendipita sacchari sp. nov. from a sugarcane rhizosphere in southern China. Mycotaxon 2020, 135, 579–587. [Google Scholar] [CrossRef]
- Hartoyo, B.; Trisilawati, O. Diversity of Arbuscular Mycorrhiza Fungi (AMF) in the rhizosphere of sugarcane. IOP Conf. Series Earth Environ. Sci. 2021, 653, 012066. [Google Scholar] [CrossRef]
- Safirzadeh, S.; Chorom, M.; Enayatizamir, N. Speciation and Fractionation of Phosphorus Affected by Enterobacter cloacae in the Rhizosphere of Sugarcane (Saccharum officinarum L.). J. Soil Sci. Plant Nutr. 2020, 21, 187–199. [Google Scholar] [CrossRef]
- Setiawati, T.C.; Mutmainnah, L. Solubilization of Potassium Containing Mineral by Microorganisms From Sugarcane Rhizosphere. Agric. Agric. Sci. Procedia 2016, 9, 108–117. [Google Scholar] [CrossRef]
- Juma, E.O.A.; Musyimi, D.M.; George, O. Antagonistic Potential Of Selected Fungal And Bacterial Isolates From Rhizosphere Of Sugarcane Variety Co 421 Against Sporisorium Scitamineum In Kibos, Kisumu County, Kenya. Appl. Sci. Rep. 2018, 1, 7–15. [Google Scholar]
- She, J.; Wang, J.; Wei, X.; Zhang, Q.; Xie, Z.; Beiyuan, J.; Xiao, E.; Yang, X.; Liu, J.; Zhou, Y.; et al. Survival strategies and dominant phylotypes of maize-rhizosphere microorganisms under metal (loid) s contamination. Sci. Total Environ. 2021, 774, 145143. [Google Scholar] [CrossRef]
- Sindhu, S.S.; Rakshiya, Y.S.; Verma, M.K. Biological Control of Termites by Antagonistic Soil Microorganisms; Springer: Berlin/Heidelberg, Germany, 2011; pp. 261–309. [Google Scholar]
- Bitas, V.; Kim, H.-S.; Bennett, J.W.; Kang, S. Sniffing on Microbes: Diverse Roles of Microbial Volatile Organic Compounds in Plant Health. Mol. Plant-Microbe Interact. 2013, 26, 835–843. [Google Scholar] [CrossRef] [PubMed]
- Köberl, M.; Müller, H.; Ramadan, E.M.; Berg, G. Desert Farming Benefits from Microbial Potential in Arid Soils and Promotes Diversity and Plant Health. PLoS ONE 2011, 6, e24452. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.; Wen, T.; Zhang, H.; Zhao, M.; Penton, C.R.; Thomashow, L.S.; Shen, Q. Predicting disease occurrence with high accuracy based on soil macroecological patterns of Fusarium wilt. ISME J. 2020, 14, 2936–2950. [Google Scholar] [CrossRef] [PubMed]
- Edwards, J.A.; Santos-Medellin, C.M.; Liechty, Z.S.; Nguyen, B.; Lurie, E.; Eason, S.; Phillips, G.; Sundaresan, V. Compositional shifts in root-associated bacterial and archaeal microbiota track the plant life cycle in field-grown rice. PLoS Biol. 2018, 16, e2003862. [Google Scholar] [CrossRef]
- Vannier, N.; Bittebiere, A.; Mony, C.; Vandenkoornhuyse, P. Root endophytic fungi impact host plant biomass and respond to plant composition at varying spatio-temporal scales. Fungal Ecol. 2020, 44, 100907. [Google Scholar] [CrossRef]
- Sugiyama, A.; Ueda, Y.; Zushi, T.; Takase, H.; Yazaki, K. Changes in the Bacterial Community of Soybean Rhizospheres during Growth in the Field. PLoS ONE 2014, 9, e100709. [Google Scholar] [CrossRef]
- Dombrowski, N.; Schlaeppi, K.; Agler, M.T.; Hacquard, S.; Kemen, E.; Garrido-Oter, R.; Wunder, J.; Coupland, G.; Schulze-Lefert, P. Root microbiota dynamics of perennial Arabis alpina are dependent on soil residence time but independent of flowering time. ISME J. 2016, 11, 43–55. [Google Scholar] [CrossRef]
- Wang, X.L.; Yang, D.T. Absorption and distribution of nitrogen, phosphorus and potassium in different growth periods of sugarcane. Chin. J. Soil Sci. 1994, 2, 224–226. [Google Scholar] [CrossRef]
- Zeng, Q.; Liu, Y.; Xiao, L.; An, S. Climate and soil properties regulate soil fungal communities on the Loess Plateau. Pedobiologia 2020, 81–82, 150668. [Google Scholar] [CrossRef]
- Štursová, M.; Žifčáková, L.; Leigh, M.B.; Burgess, R.; Baldrian, P. Cellulose utilization in forest litter and soil: Identification of bacterial and fungal decomposers. Fems Microbiol. Ecol. 2012, 80, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Jia, R.; Yu, L. Diversity and distribution of soil fungal communities associated with biological soil crusts in the southeastern Tengger Desert (China) as revealed by 454 pyrosequencing. Fungal. Ecol. 2016, 23, 156–163. [Google Scholar] [CrossRef]
- Da Silva, K.J.; de Armas, R.D.; Soares, C.R.F.S.; Ogliari, J.B. Communities of endophytic microorganisms in different developmental stages from a local variety as well as transgenic and conventional isogenic hybrids of maize. World J. Microbiol. Biotechnol. 2016, 32, 189. [Google Scholar] [CrossRef]
- Wang, Z.; Wang, T.; Zhu, H.; Pan, H.; Yu, X. Diversity and dynamics of microbial communities in brown planthopper at different developmental stages revealed by high-throughput amplicon sequencing. Insect Sci. 2019, 27, 883–894. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, F.; Vale, F.X.R.; Datnoff, L.E.; Prabhu, A.S.; Korndörfer, G.H. Effect of Rice Growth Stages and Silicon on Sheath Blight Development. Phytopathology® 2003, 93, 256–261. [Google Scholar] [CrossRef]
- Poli, A.; Lazzari, A.; Prigione, V.; Voyron, S.; Spadaro, D.; Varese, G.C. Influence of plant genotype on the cultivable fungi associated to tomato rhizosphere and roots in different soils. Fungal Biol. 2016, 120, 862–872. [Google Scholar] [CrossRef]
- Hu, X.; Li, M.; Chen, H. Community structure of gut fungi during different developmental stages of the Chinese white pine beetle (Dendroctonus armandi). Sci. Rep. 2015, 5, srep08411. [Google Scholar] [CrossRef]
- Babu, A.G.; Kim, S.W.; Yadhav, D.R.; Adhikari, M.; Kim, C.; Lee, H.B.; Lee, Y.S. A New Record of Pseudallescheria boydii Isolated from Crop Field Soil in Korea. Mycobiology 2014, 42, 397–400. [Google Scholar] [CrossRef]
- Felix, G.; Josep, C.; Josepa, G.; Josep, G. Molecular Phylogeny of the Pseudallescheria boydii Species Complex: Proposal of Two New Species. J. Clin. Microbiol. 2005, 43, 4930–4942. [Google Scholar]
- Rainer, J.; De Hoog, G.S.; Wedde, M.; Gräser, Y.; Gilges, S.; Centraalbureau Voor Schimmelcultures, B.N. Molecular Variability of Pseudallescheria boydii, a Neurotropic Opportunist. J. Clin. Microbiol. 2000, 38, 3267–3273. [Google Scholar] [CrossRef]
- Lombard, L.; van der Merwe, N.A.; Groenewald, J.Z.; Crous, P.W. Generic concepts in Nectriaceae. Stud. Mycol. 2015, 80, 189–245. [Google Scholar] [CrossRef]
- Meng, Z. Relation between the Quantity of Soil Microbe and Soil Factor in the Second Rotation Chinese Fir Plantation. Sci. Silvae Sin. 2007, 43, 7–12. [Google Scholar]
- Hüttl, R.; Frielinghaus, M. Soil fertility problems—An agriculture and forestry perspective. Sci. Total Environ. 1994, 143, 63–74. [Google Scholar] [CrossRef]
- Lin, Z.; Wang, J.; Bao, Y.; Guo, Q.; Powell, C.A.; Xu, S.; Chen, B.; Zhang, M. Deciphering the transcriptomic response of Fusarium verticillioides in relation to nitrogen availability and the development of sugarcane pokkah boeng disease. Sci. Rep. 2016, 6, 29692. [Google Scholar] [CrossRef]
- Bao, Y.; Pan, K.; Khan, M.T.; Chen, B.; Zhang, M. High-Quality Genome Sequence Resource forFusarium andiyazi Causing Pokkah Boeng Disease of Sugarcane in China. Mol. Plant-Microbe Interact. 2021, 34, 973–976. [Google Scholar] [CrossRef]
- Jia, X.; Dini-Andreote, F.; Salles, J.F. Community Assembly Processes of the Microbial Rare Biosphere. Trends Microbiol. 2018, 26, 738–747. [Google Scholar] [CrossRef]
- Dini-Andreote, F.; Stegen, J.C.; van Elsas, J.D.; Salles, J.F. Disentangling mechanisms that mediate the balance between stochastic and deterministic processes in microbial succession. Proc. Natl. Acad. Sci. USA 2015, 112, E1326–E1332. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, Z.; Li, T.; Zhao, D.; Han, J.; Liao, Y. Wheat rhizosphere fungal community is affected by tillage and plant growth. Agric. Ecosyst. Environ. 2021, 317, 107475. [Google Scholar] [CrossRef]
- Xu, J. Sugarcane planting techniques and field management methods. Rural. Technol. 2020, 11, 103–104. [Google Scholar]
- Sun, W.; Chen, S.; Liu, X.; Wang, K.; Yang, Y. Effect of soil sampling designs on soil microbial alpha diversity. Microbiol. China 2020, 47, 1699–1708. [Google Scholar] [CrossRef]
- Riley, D.; Barber, S.A. Bicarbonate Accumulation and pH Changes at the Soybean (Glycine max (L.) Merr.) Root-Soil Interface. Soil Sci. Soc. Am. J. 1969, 33, 905–908. [Google Scholar] [CrossRef]
- Liu, X.; Li, Z.; Liu, R.; Li, L.; Wang, W. Changes of bacterial flora structure in rhizosphere soil of soybean at different growth stages. Guangxi Plant 2018, 38, 1363–1370. [Google Scholar] [CrossRef]
- Bao, S.D. Soil and Agricultural Chemistry Analysis; China Agriculture Press: Beijing, China, 2000. [Google Scholar]
- Tong, J.; Xie, C.M. Analysis of Influencing Factors for Determination of Soil Hydrolyzable Nitrogen by Alkaline Diffusion Method. Ningxia Agric. For. Sci. Technol. 2011, 52, 61–71. [Google Scholar]
- Qin, L.; Huang, S.Q.; Zhong, L.L.; Zhou, H.; Zhao, S.; Xiang, B.; Lei, S.R. Comparison of Dumas combustion and Kjeldahl methods for determining total nitrogen content in soil. Soil Fertil. Sci. China 2020, 5, 258–265. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Du, Q.W.; Liu, Y.; Han, G.; Wang, W. Comparison of Three Commonly Used Methods for Measuring Soil Organic Matter. Heilongjiang Agric. Sci. 2014, 12, 163. [Google Scholar]
- Wang, F. Study on the Potassium Determination in Soil by Automatic Digestion Equipment-Flame Photometry Method. Ningxia Agric. For. Sci. Technol. 2017, 58, 38–39. [Google Scholar]
- Ye, X.S.; Wu, X.R.; Zhao, Z.Q. Comparison on the Determination of Soil Total Phosphorus Concentration by Continuous Flow Injection Analyzer and Traditional Analysis Methods. Lab. Res. Explor. 2010, 29, 50–53. [Google Scholar]
- Guo, Y.F. Discussion on the Determination of Quick-acting Potassium in Soil by Using Ammonium Acetate Flame Photometry and M3 Method and the Correlation Study. Shanxi Sci. Technol. 2014, 29, 54–56. [Google Scholar]
- Bu, Y.S.; Magdoff, F. A comparison of ten methods for determination of available phosphorus in soils. Acta Pedol. Sin. 2003, 40, 140–146. [Google Scholar] [CrossRef]
- He, L. The overview of plant diseases and insect pests of sugarcane and the pest control technique in Guangxi. J. Guangxi Agric. 2017, 32, 4. [Google Scholar]
- Rousk, J.; Bååth, E.; Brookes, P.C.; Lauber, C.L.; Lozupone, C.; Caporaso, J.G.; Knight, R.; Fierer, N. Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 2010, 4, 1340–1351. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Tang, Q.; Zhang, C. Data Processing System (DPS) software with experimental design, statistical analysis and data mining developed for use in entomological research. Insect. Sci. 2013, 20, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Wade, J.; Waterhouse, H.; Roche, L.M.; Horwath, W.R. Structural equation modeling reveals iron (hydr)oxides as a strong mediator of N mineralization in California agricultural soils. Geoderma 2018, 315, 120–129. [Google Scholar] [CrossRef]
- Kembel, S.W. Disentangling niche and neutral influences on community assembly: Assessing the performance of community phylogenetic structure tests. Ecol. Lett. 2009, 12, 949–960. [Google Scholar] [CrossRef]
- Webb, C.O.; Ackerly, D.D.; Kembel, S.W. Phylocom: Software for the analysis of phylogenetic community structure and trait evolution. Bioinformatics 2008, 24, 2098–2100. [Google Scholar] [CrossRef] [PubMed]
- Borgatti, S.P.; Everett, M.G.; Freeman, L.C. UCINET for Windows: Software for social network analysis. Harv. MA Anal. Technol. 2002, 6, 12–15. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Height (cm) | Stem Diameter (mm) | Total Number/ha | Valid Number/ha | Effective Ratio (%) | Disease Ratio (%) | |
|---|---|---|---|---|---|---|
| Seedling | 42.49 ± 1.37 c | 14.70 ± 0.80 c | 3270 | 3185 | 97.40 | 1.4 |
| Tillering | 56.30 ± 2.10 c | 23.70 ± 1.49 b | 3473 | 3307 | 95.22 | 5.1 |
| Elongation | 122.90 ± 4.54 b | 27.90 ± 1.25 b | 2940 | 2103 | 71.52 | 19.8 |
| Maturity | 219.67 ± 5.98 a | 36.67 ± 1.01 a | 2950 | 2528 | 85.70 | 15.1 |
| Seedling | Tillering | Elongation | Maturity | |
|---|---|---|---|---|
| pH | 6.22 ± 0.04 a | 5.83 ± 0.05 b | 5.55 ± 0.06 c | 5.38 ± 0.09 c |
| OM/(g kg−1) | 26.78 ± 0.89 ab | 25.03 ± 0.54 b | 30.80 ± 1.91 a | 26.71 ± 1.19 ab |
| TN/(g kg−1) | 2.74 ± 0.04 a | 2.45 ± 0.02 b | 2.39 ± 0.03 b | 2.40 ± 0.02 b |
| TP/(g kg−1) | 0.80 ± 0.01 b | 0.78 ± 0.01 b | 0.92 ± 0.02 a | 0.82 ± 0.01 b |
| TK/(g kg−1) | 23.25 ± 0.20 a | 16.58 ± 0.44 c | 19.65 ± 0.24 b | 17.18 ± 0.28 c |
| AN/(mg kg−1) | 182.06 ± 5.73 a | 168.75 ± 3.12 a | 127.01 ± 1.67 b | 125.04 ± 3.46 b |
| AP/(mg kg−1) | 78.61 ± 1.87 ab | 82.08 ± 1.19 a | 65.47 ± 1.48 c | 75.46 ± 2.39 b |
| AK/(mg kg−1) | 155.99 ± 4.08 a | 149.97 ± 3.49 ab | 136.11 ± 5.70 ab | 125.11 ± 14.72 b |
| C:N | 9.84 ± 0.35 b | 10.23 ± 0.27 b | 12.89 ± 0.80 a | 11.15 ± 0.52 ab |
| Tem | 24.49 ± 0.04 b | 30.36 ± 0.06 a | 30.22 ± 0.04 a | 19.37 ± 0.03 c |
| Seedling | Tillering | Elongation | Maturity | |
|---|---|---|---|---|
| Sobs | 310.81 ± 6.34 b | 388.48 ± 6.79 a | 328.62 ± 6.46 b | 277.67 ± 12.15 c |
| ACE | 396.43 ± 9.88 b | 466.04 ± 9.49 a | 415.22 ± 9.08 b | 338.92 ± 14.47 c |
| Shannon | 3.36 ± 0.08 b | 3.83 ± 0.04 a | 3.52 ± 0.06 b | 3.35 ± 0.05 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Q.; Pang, Z.; Liu, Y.; Fallah, N.; Hu, C.; Lin, W.; Yuan, Z. Rhizosphere Fungal Dynamics in Sugarcane during Different Growth Stages. Int. J. Mol. Sci. 2023, 24, 5701. https://doi.org/10.3390/ijms24065701
Liu Q, Pang Z, Liu Y, Fallah N, Hu C, Lin W, Yuan Z. Rhizosphere Fungal Dynamics in Sugarcane during Different Growth Stages. International Journal of Molecular Sciences. 2023; 24(6):5701. https://doi.org/10.3390/ijms24065701
Chicago/Turabian StyleLiu, Qiang, Ziqin Pang, Yueming Liu, Nyumah Fallah, Chaohua Hu, Wenxiong Lin, and Zhaonian Yuan. 2023. "Rhizosphere Fungal Dynamics in Sugarcane during Different Growth Stages" International Journal of Molecular Sciences 24, no. 6: 5701. https://doi.org/10.3390/ijms24065701
APA StyleLiu, Q., Pang, Z., Liu, Y., Fallah, N., Hu, C., Lin, W., & Yuan, Z. (2023). Rhizosphere Fungal Dynamics in Sugarcane during Different Growth Stages. International Journal of Molecular Sciences, 24(6), 5701. https://doi.org/10.3390/ijms24065701