
Distinct Patterns of GR Transcriptional Regulation in Liver and Muscle of LPS-Challenged Weaning Piglets


Abstract
1. Introduction
2. Results
2.1. Distinct Inflammatory Responses Caused by LPS Challenge in Liver and Muscle
2.2. Cortisol Content and GR Expression in Liver and Muscle in Response to LPS Challenge
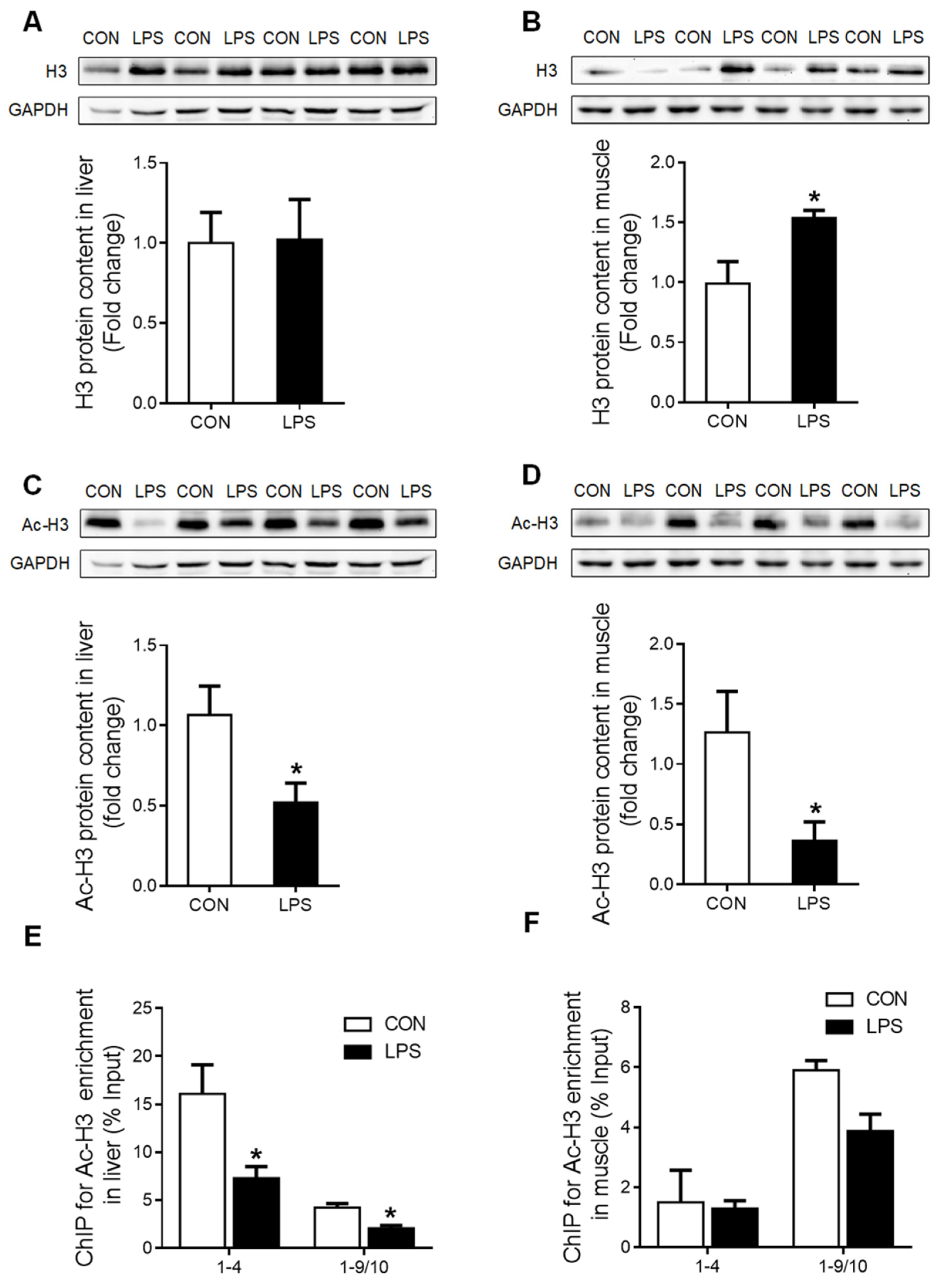
2.3. Transcriptional Regulation of GR in Liver and Muscle under LPS Treatment
2.4. GR Binding to TNF-α and IL-6 Gene Promoters in Liver and Muscle
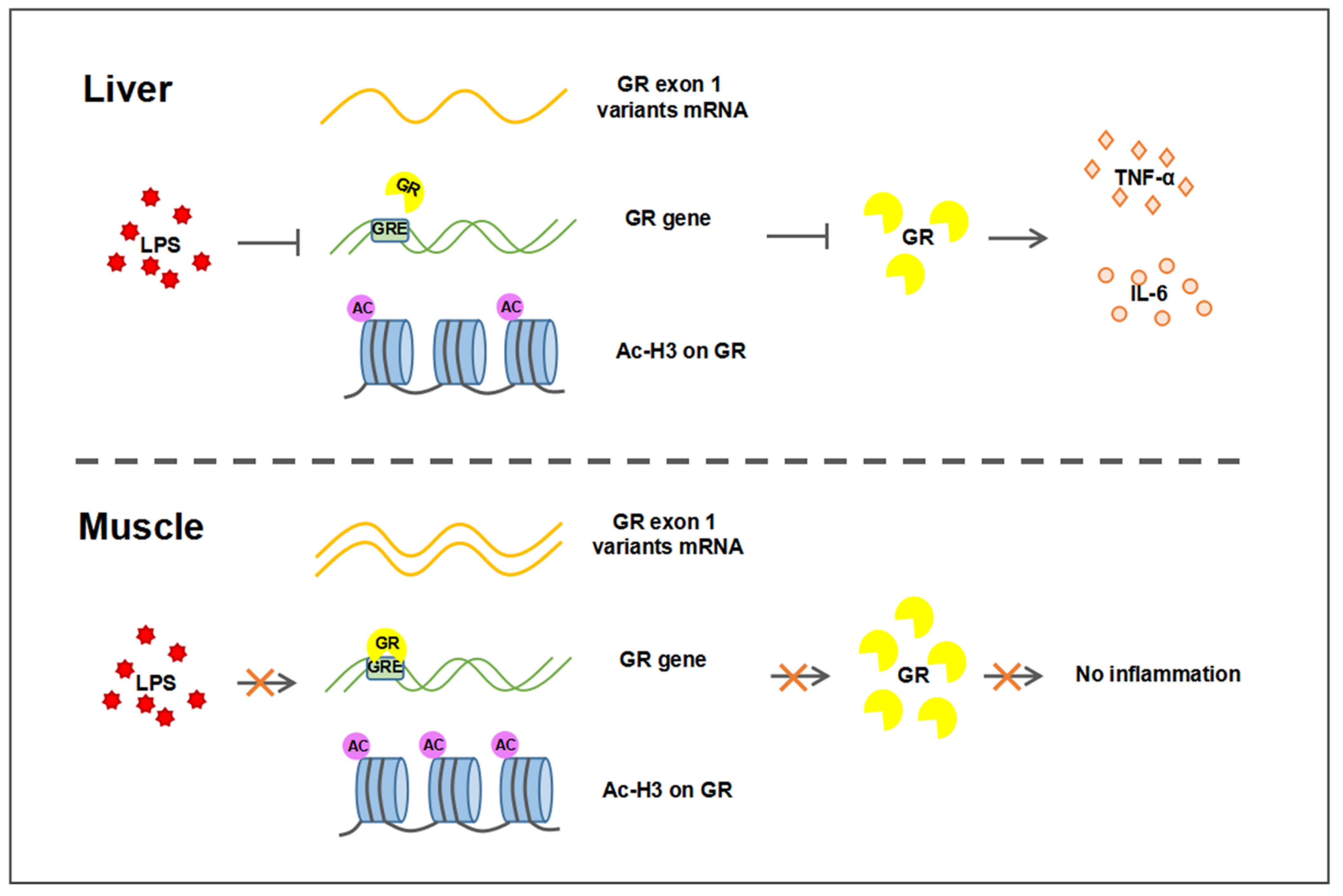
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Animals and Treatment
4.3. Liver and Muscle Cortisol Assay
4.4. Determination of Liver and Muscle Concentrations of IL-6 and TNF-α
4.5. Total RNA Isolation and Real-Time PCR
4.6. Total Cytosolic Protein Extraction and Western Blotting
4.7. Chromatin Immunoprecipitation (ChIP) Assay
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Oakley, R.H.; Cidlowski, J.A. The biology of the glucocorticoid receptor: New signaling mechanisms in health and disease. J. Allergy Clin. Immunol. 2013, 132, 1033–1044. [Google Scholar] [CrossRef] [PubMed]
- Baschant, U.; Tuckermann, J. The role of the glucocorticoid receptor in inflammation and immunity. J. Steroid Biochem. Mol. Biol. 2010, 120, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Meers, G.K.; Bohnenberger, H.; Reichardt, H.M.; Luhder, F.; Reichardt, S.D. Impaired resolution of DSS-induced colitis in mice lacking the glucocorticoid receptor in myeloid cells. PLoS ONE 2018, 13, e0190846. [Google Scholar] [CrossRef]
- Carrillo-de Sauvage, M.A.; Maatouk, L.; Arnoux, I.; Pasco, M.; Sanz Diez, A.; Delahaye, M.; Herrero, M.T.; Newman, T.A.; Calvo, C.F.; Audinat, E.; et al. Potent and multiple regulatory actions of microglial glucocorticoid receptors during CNS inflammation. Cell Death Differ. 2013, 20, 1546–1557. [Google Scholar] [CrossRef] [PubMed]
- Cain, D.W.; Cidlowski, J.A. Specificity and sensitivity of glucocorticoid signaling in health and disease. Best Pract. Res. Clin. Endocrinol. Metab. 2015, 29, 545–556. [Google Scholar] [CrossRef] [PubMed]
- Donham, K.J. Association of environmental air contaminants with disease and productivity in swine. Am. J. Vet. Res. 1991, 52, 1723–1730. [Google Scholar] [PubMed]
- Patil, Y.; Gooneratne, R.; Ju, X.H. Interactions between host and gut microbiota in domestic pigs: A review. Gut Microbes 2019, 11, 310–334. [Google Scholar] [CrossRef]
- Zhang, G.; Meredith, T.C.; Kahne, D. On the essentiality of lipopolysaccharide to Gram-negative bacteria. Curr. Opin. Microbiol. 2013, 16, 779–785. [Google Scholar] [CrossRef]
- Chow, J.C.; Young, D.W.; Golenbock, D.T.; Christ, W.J.; Gusovsky, F. Toll-like receptor-4 mediates lipopolysaccharide-induced signal transduction. J. Biol. Chem. 1999, 274, 10689–10692. [Google Scholar] [CrossRef]
- Yucel, G.; Zhao, Z.; El-Battrawy, I.; Lan, H.; Lang, S.; Li, X.; Buljubasic, F.; Zimmermann, W.H.; Cyganek, L.; Utikal, J.; et al. Lipopolysaccharides induced inflammatory responses and electrophysiological dysfunctions in human-induced pluripotent stem cell derived cardiomyocytes. Sci. Rep. 2017, 7, 2935. [Google Scholar] [CrossRef]
- McGinnis, R.; Walker, J.; Margules, D.; Aird, F.; Redei, E. Dysregulation of the Hypothalamus-Pituitary-Adrenal Axis in Male and Female, Genetically Obese (ob/ob) Mice. J. Neuroendocrinol. 1992, 4, 765–771. [Google Scholar] [CrossRef] [PubMed]
- Rhen, T.; Cidlowski, J.A. Antiinflammatory action of glucocorticoids—New mechanisms for old drugs. N. Engl. J. Med. 2005, 353, 1711–1723. [Google Scholar] [CrossRef]
- Da, J.; Chen, L.; Hedenstierna, G. Nitric oxide up-regulates the glucocorticoid receptor and blunts the inflammatory reaction in porcine endotoxin sepsis. Crit. Care Med. 2007, 35, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Zou, H.; Jiang, Z.; Li, R.; Jia, Y.; Yang, X.; Ni, Y.; Zhao, R. p53 cooperates with Sp1 to regulate breed-dependent expression of glucocorticoid receptor in the liver of preweaning piglets. PLoS ONE 2013, 8, e70494. [Google Scholar] [CrossRef] [PubMed]
- Geng, C.D.; Vedeckis, W.V. Steroid-responsive sequences in the human glucocorticoid receptor gene 1A promoter. Mol. Endocrinol. 2004, 18, 912–924. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Zong, J.; Ashraf, J.; Thompson, E.B. The promoter and first, untranslated exon of the human glucocorticoid receptor gene are GC rich but lack consensus glucocorticoid receptor element sites. Mol. Cell. Biol. 1990, 10, 5580–5585. [Google Scholar] [CrossRef]
- Lu, Y.; Xiong, X.; Wang, X.; Zhang, Z.; Li, J.; Shi, G.; Yang, J.; Zhang, H.; Ning, G.; Li, X. Yin Yang 1 promotes hepatic gluconeogenesis through upregulation of glucocorticoid receptor. Diabetes 2013, 62, 1064–1073. [Google Scholar] [CrossRef]
- Ratman, D.; Vanden Berghe, W.; Dejager, L.; Libert, C.; Tavernier, J.; Beck, I.M.; De Bosscher, K. How glucocorticoid receptors modulate the activity of other transcription factors: A scope beyond tethering. Mol. Cell. Endocrinol. 2013, 380, 41–54. [Google Scholar] [CrossRef]
- Presul, E.; Schmidt, S.; Kofler, R.; Helmberg, A. Identification, tissue expression, and glucocorticoid responsiveness of alternative first exons of the human glucocorticoid receptor. J. Mol. Endocrinol. 2007, 38, 79–90. [Google Scholar] [CrossRef][Green Version]
- Jiang, Z.; Qian, L.; Zou, H.; Jia, Y.; Ni, Y.; Yang, X.; Jiang, Z.; Zhao, R. Porcine glucocorticoid receptor (NR3C1) gene: Tissue-specificity of transcriptional strength and glucocorticoid responsiveness of alternative promoters. J. Steroid Biochem. Mol. Biol. 2014, 141, 87–93. [Google Scholar] [CrossRef]
- Zhang, Y.; Guo, F.; Ni, Y.; Zhao, R. LPS-induced inflammation in the chicken is associated with CCAAT/enhancer binding protein beta-mediated fat mass and obesity associated gene down-regulation in the liver but not hypothalamus. BMC Vet. Res. 2013, 9, 257. [Google Scholar] [CrossRef] [PubMed]
- Bode, J.G.; Albrecht, U.; Haussinger, D.; Heinrich, P.C.; Schaper, F. Hepatic acute phase proteins—Regulation by IL-6- and IL-1-type cytokines involving STAT3 and its crosstalk with NF-kappaB-dependent signaling. Eur. J. Cell Biol. 2012, 91, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Boaru, S.G.; Borkham-Kamphorst, E.; Tihaa, L.; Haas, U.; Weiskirchen, R. Expression analysis of inflammasomes in experimental models of inflammatory and fibrotic liver disease. J. Inflamm. 2012, 9, 49. [Google Scholar] [CrossRef] [PubMed]
- Labbus, K.; Henning, M.; Borkham-Kamphorst, E.; Geisler, C.; Berger, T.; Mak, T.W.; Knuchel, R.; Meyer, H.E.; Weiskirchen, R.; Henkel, C. Proteomic profiling in Lipocalin 2 deficient mice under normal and inflammatory conditions. J. Proteom. 2013, 78, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Borkham-Kamphorst, E.; van de Leur, E.; Zimmermann, H.W.; Karlmark, K.R.; Tihaa, L.; Haas, U.; Tacke, F.; Berger, T.; Mak, T.W.; Weiskirchen, R. Protective effects of lipocalin-2 (LCN2) in acute liver injury suggest a novel function in liver homeostasis. Biochim. Biophys. Acta 2013, 1832, 660–673. [Google Scholar] [CrossRef] [PubMed]
- Frost, R.A.; Nystrom, G.J.; Lang, C.H. Lipopolysaccharide regulates proinflammatory cytokine expression in mouse myoblasts and skeletal muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 283, R698–R709. [Google Scholar] [CrossRef]
- Smoak, K.A.; Cidlowski, J.A. Mechanisms of glucocorticoid receptor signaling during inflammation. Mech. Ageing Dev. 2004, 125, 697–706. [Google Scholar] [CrossRef]
- Whirledge, S.; DeFranco, D.B. Glucocorticoid Signaling in Health and Disease: Insights from Tissue-Specific GR Knockout Mice. Endocrinology 2018, 159, 46–64. [Google Scholar] [CrossRef]
- Dostal, C.R.; Gamsby, N.S.; Lawson, M.A.; McCusker, R.H. Glia- and tissue-specific changes in the Kynurenine Pathway after treatment of mice with lipopolysaccharide and dexamethasone. Brain Behav. Immun. 2018, 69, 321–335. [Google Scholar] [CrossRef]
- McCormick, J.A.; Lyons, V.; Jacobson, M.D.; Noble, J.; Diorio, J.; Nyirenda, M.; Weaver, S.; Ester, W.; Yau, J.L.; Meaney, M.J.; et al. 5′-heterogeneity of glucocorticoid receptor messenger RNA is tissue specific: Differential regulation of variant transcripts by early-life events. Mol. Endocrinol. 2000, 14, 506–517. [Google Scholar] [CrossRef][Green Version]
- Strahle, U.; Schmidt, A.; Kelsey, G.; Stewart, A.F.; Cole, T.J.; Schmid, W.; Schutz, G. At least three promoters direct expression of the mouse glucocorticoid receptor gene. Proc. Natl. Acad. Sci. USA 1992, 89, 6731–6735. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Watson, C.S.; Gametchu, B. Multiple glucocorticoid receptor transcripts in membrane glucocorticoid receptor-enriched S-49 mouse lymphoma cells. J. Cell. Biochem. 1999, 74, 418–429. [Google Scholar] [CrossRef]
- Turner, J.D.; Muller, C.P. Structure of the glucocorticoid receptor (NR3C1) gene 5′ untranslated region: Identification, and tissue distribution of multiple new human exon 1. J. Mol. Endocrinol. 2005, 35, 283–292. [Google Scholar] [CrossRef]
- Turner, J.D.; Schote, A.B.; Macedo, J.A.; Pelascini, L.P.; Muller, C.P. Tissue specific glucocorticoid receptor expression, a role for alternative first exon usage? Biochem. Pharmacol. 2006, 72, 1529–1537. [Google Scholar] [CrossRef] [PubMed]
- Eisen, L.P.; Elsasser, M.S.; Harmon, J.M. Positive regulation of the glucocorticoid receptor in human T-cells sensitive to the cytolytic effects of glucocorticoids. J. Biol. Chem. 1988, 263, 12044–12048. [Google Scholar] [CrossRef]
- Ramdas, J.; Liu, W.; Harmon, J.M. Glucocorticoid-induced cell death requires autoinduction of glucocorticoid receptor expression in human leukemic T cells. Cancer Res. 1999, 59, 1378–1385. [Google Scholar] [PubMed]
- Sapolsky, R.M.; Krey, L.C.; McEwen, B.S. Stress down-regulates corticosterone receptors in a site-specific manner in the brain. Endocrinology 1984, 114, 287–292. [Google Scholar] [CrossRef]
- Vandevyver, S.; Dejager, L.; Libert, C. Comprehensive overview of the structure and regulation of the glucocorticoid receptor. Endocr. Rev. 2014, 35, 671–693. [Google Scholar] [CrossRef]
- Wang, Z.; Frederick, J.; Garabedian, M.J. Deciphering the phosphorylation “code” of the glucocorticoid receptor in vivo. J. Biol. Chem. 2002, 277, 26573–26580. [Google Scholar] [CrossRef]
- Rogatsky, I.; Waase, C.L.; Garabedian, M.J. Phosphorylation and inhibition of rat glucocorticoid receptor transcriptional activation by glycogen synthase kinase-3 (GSK-3). Species-specific differences between human and rat glucocorticoid receptor signaling as revealed through GSK-3 phosphorylation. J. Biol. Chem. 1998, 273, 14315–14321. [Google Scholar] [CrossRef]
- Rollins, D.A.; Kharlyngdoh, J.B.; Coppo, M.; Tharmalingam, B.; Mimouna, S.; Guo, Z.; Sacta, M.A.; Pufall, M.A.; Fisher, R.P.; Hu, X.; et al. Glucocorticoid-induced phosphorylation by CDK9 modulates the coactivator functions of transcriptional cofactor GRIP1 in macrophages. Nat. Commun. 2017, 8, 1739. [Google Scholar] [CrossRef] [PubMed]
- Beck, I.M.; Vanden Berghe, W.; Vermeulen, L.; Yamamoto, K.R.; Haegeman, G.; De Bosscher, K. Crosstalk in inflammation: The interplay of glucocorticoid receptor-based mechanisms and kinases and phosphatases. Endocr. Rev. 2009, 30, 830–882. [Google Scholar] [CrossRef] [PubMed]
- Govindan, M.V. Recruitment of cAMP-response element-binding protein and histone deacetylase has opposite effects on glucocorticoid receptor gene transcription. J. Biol. Chem. 2010, 285, 4489–4510. [Google Scholar] [CrossRef] [PubMed]
- Aung, H.T.; Schroder, K.; Himes, S.R.; Brion, K.; van Zuylen, W.; Trieu, A.; Suzuki, H.; Hayashizaki, Y.; Hume, D.A.; Sweet, M.J.; et al. LPS regulates proinflammatory gene expression in macrophages by altering histone deacetylase expression. FASEB J. 2006, 20, 1315–1327. [Google Scholar] [CrossRef]
- Sailhamer, E.A.; Li, Y.; Smith, E.J.; Shuja, F.; Shults, C.; Liu, B.; Soupir, C.; deMoya, M.; Velmahos, G.; Alam, H.B. Acetylation: A novel method for modulation of the immune response following trauma/hemorrhage and inflammatory second hit in animals and humans. Surgery 2008, 144, 204–216. [Google Scholar] [CrossRef]
- Yamamoto, K.R. Multilayered control of intracellular receptor function. Harvey Lect. 1995, 91, 1–19. [Google Scholar]
- Kumar, R.; Thompson, E.B. Gene regulation by the glucocorticoid receptor: Structure:function relationship. J. Steroid Biochem. Mol. Biol. 2005, 94, 383–394. [Google Scholar] [CrossRef]
- Meagher, L.C.; Cousin, J.M.; Seckl, J.R.; Haslett, C. Opposing effects of glucocorticoids on the rate of apoptosis in neutrophilic and eosinophilic granulocytes. J. Immunol. 1996, 156, 4422–4428. [Google Scholar]
- Vandevyver, S.; Dejager, L.; Van Bogaert, T.; Kleyman, A.; Liu, Y.; Tuckermann, J.; Libert, C. Glucocorticoid receptor dimerization induces MKP1 to protect against TNF-induced inflammation. J. Clin. Investig. 2012, 122, 2130–2140. [Google Scholar] [CrossRef]
- Ronchetti, S.; Migliorati, G.; Riccardi, C. GILZ as a Mediator of the Anti-Inflammatory Effects of Glucocorticoids. Front. Endocrinol. 2015, 6, 170. [Google Scholar] [CrossRef]
- Chinenov, Y.; Coppo, M.; Gupte, R.; Sacta, M.A.; Rogatsky, I. Glucocorticoid receptor coordinates transcription factor-dominated regulatory network in macrophages. BMC Genom. 2014, 15, 656. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Sun, Q.; Li, X.; Wang, M.; Cai, D.; Li, X.; Zhao, R. In Ovo injection of betaine affects hepatic cholesterol metabolism through epigenetic gene regulation in newly hatched chicks. PLoS ONE 2015, 10, e0122643. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Cong, R.; Li, R.; Yang, X.; Sun, Q.; Parvizi, N.; Zhao, R. Maternal low-protein diet induces gender-dependent changes in epigenetic regulation of the glucose-6-phosphatase gene in newborn piglet liver. J. Nutr. 2012, 142, 1659–1665. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Genes | Primer Sequences (5′ to 3′) | Used For | |
|---|---|---|---|
| GR exon1–4 | F: ATCGTAATATGTGCGGCC | R: TTGCTTCCTGAGCCTTTT | Real-time PCR |
| GR exon1–5 | F: AACTTGGATGCGGGCCC | R: TTGCTTCCTGAGCCTTTT | Real-time PCR |
| GR exon1–6 | F: ATGCGGGGGAGGGGGACC | R: ACGCTGCTGGGGATTTC | Real-time PCR |
| GR exon1–7 | F: GCGGCGAAGAGAAACTAGAG | R: ACGCTGCTGGGGATTTC | Real-time PCR |
| GR exon1–9/10 | F: CCTGCTTTCACACGCTAA | R: ACGCTGCTGGGGATTTC | Real-time PCR |
| GR exon1–11 | F: AACTTGGATGCGGGCC | R: TTGCTTCCTGAGCCTTTT | Real-time PCR |
| Total GR | F: CCAAGGAATCGCTGACCCAG | R: ATTGCTTCCTGAGCCTTTTGG | Real-time PCR |
| TNF-α Fragment 1 | F: CGCTCAGAAGTCCGTGTGC | R: GTCATAAGGGAGGTGAAAAGGG | ChIP PCR |
| TNF-α Fragment 2 | F: GCCTTCTCCCCTTTTCACCT | R: CAGATCCTCAGCAGAGCATCC | ChIP PCR |
| TNF-α Fragment 3 | F: TGAGGGACTTTGAACGGATGA | R: CAGATACGAGCAGACAGCATTTC | ChIP PCR |
| IL-6 Fragment 1 | F: TTGTTCAGTGCTGGAGGGTTG | R: TGCTTGGGTTTGCTTCGC | ChIP PCR |
| IL-6 Fragment 2 | F: CAAACCCAAGCAGATTGTGATG | R: CAGTTGCCCAGACACCCG | ChIP PCR |
| GR exon1–4 Fragment | F: AGACTTAGGTGTGATTCTGCGGA | R: CATTGATATGACTGACGGTGGC | ChIP PCR |
| GR exon1–9/10 Fragment | F: CTGAGATACCGCCATCCTAAAGT | R: GGGCTCTGTCCACAAACCATT | ChIP PCR |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Jiang, Y.; Jiang, Z.; Feng, Y.; Zhao, R. Distinct Patterns of GR Transcriptional Regulation in Liver and Muscle of LPS-Challenged Weaning Piglets. Int. J. Mol. Sci. 2022, 23, 8072. https://doi.org/10.3390/ijms23158072
Liu J, Jiang Y, Jiang Z, Feng Y, Zhao R. Distinct Patterns of GR Transcriptional Regulation in Liver and Muscle of LPS-Challenged Weaning Piglets. International Journal of Molecular Sciences. 2022; 23(15):8072. https://doi.org/10.3390/ijms23158072
Chicago/Turabian StyleLiu, Jie, Yidan Jiang, Zheng Jiang, Yue Feng, and Ruqian Zhao. 2022. "Distinct Patterns of GR Transcriptional Regulation in Liver and Muscle of LPS-Challenged Weaning Piglets" International Journal of Molecular Sciences 23, no. 15: 8072. https://doi.org/10.3390/ijms23158072
APA StyleLiu, J., Jiang, Y., Jiang, Z., Feng, Y., & Zhao, R. (2022). Distinct Patterns of GR Transcriptional Regulation in Liver and Muscle of LPS-Challenged Weaning Piglets. International Journal of Molecular Sciences, 23(15), 8072. https://doi.org/10.3390/ijms23158072