
Isomeric 3-Pyridinylmethylcoumarins Differ in Erk1/2-Inhibition and Modulation of BV2 Microglia-Mediated Neuroinflammation

 , , , and
, , , and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
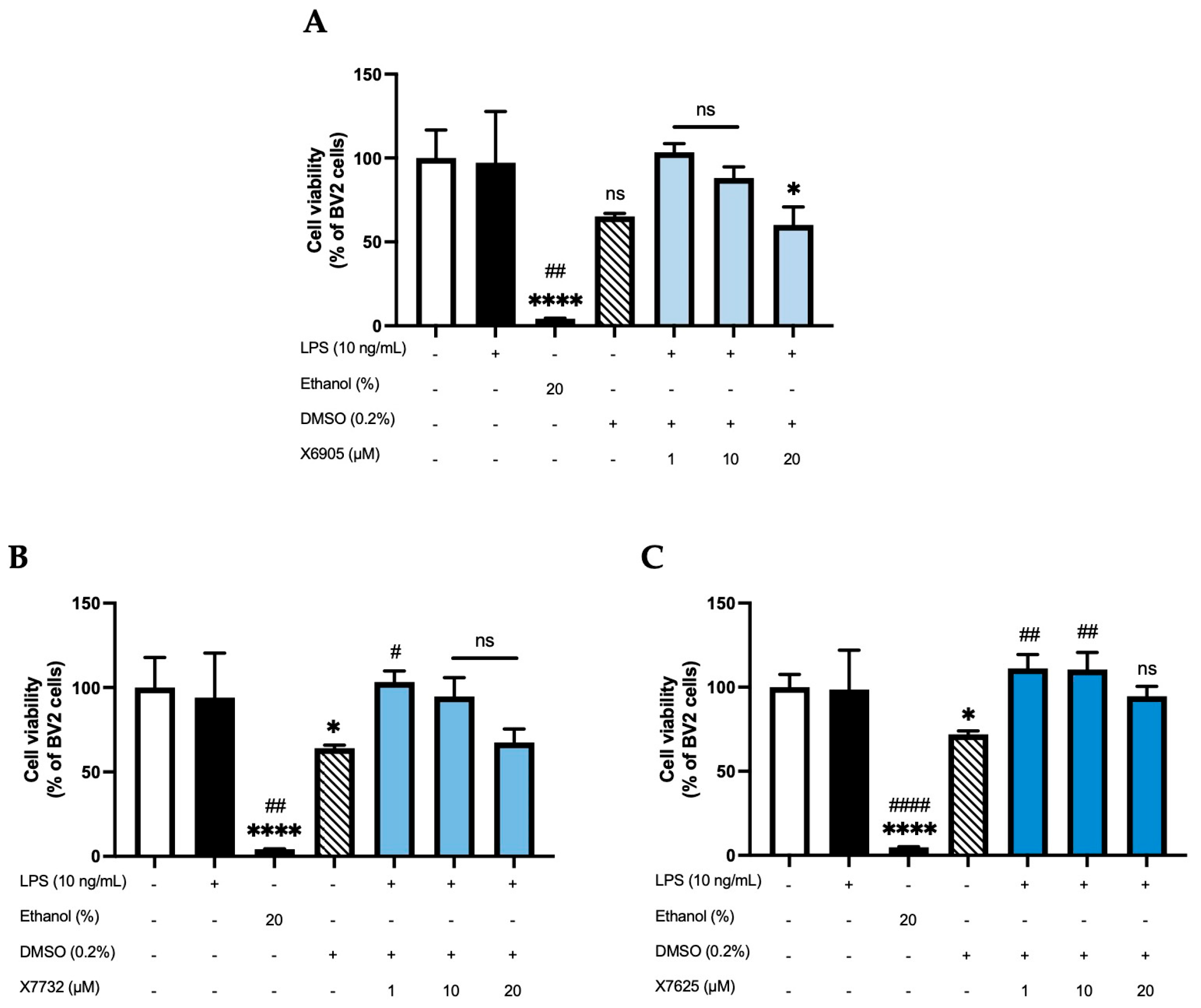
2.1. Effects of X6905, X7732, and X7625 on Cell Viability (ATP -Assay)
2.2. Effects of X6905, X7732, and X7625 on LPS-Induced Cytokine Release
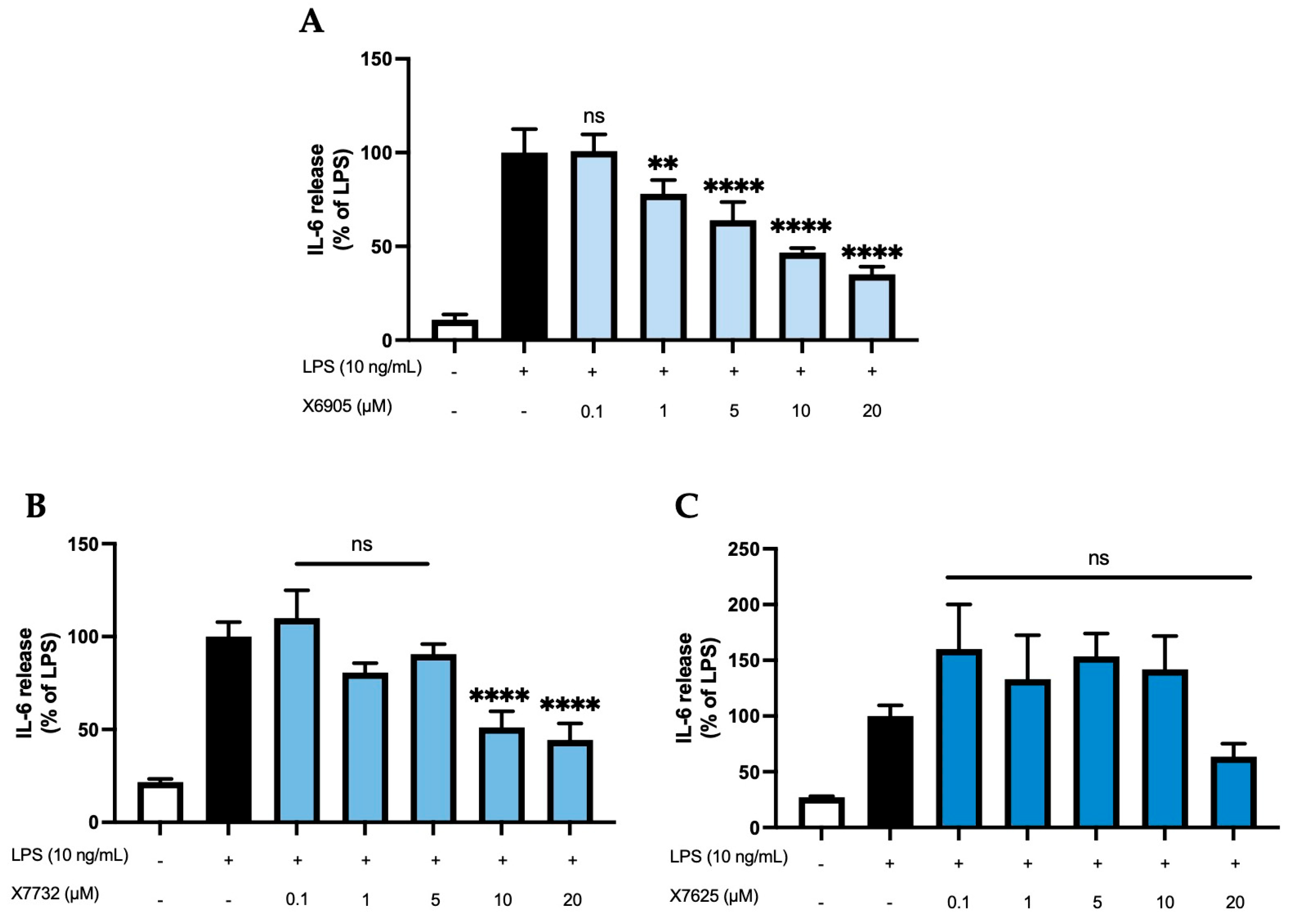
2.2.1. IL-6 Release
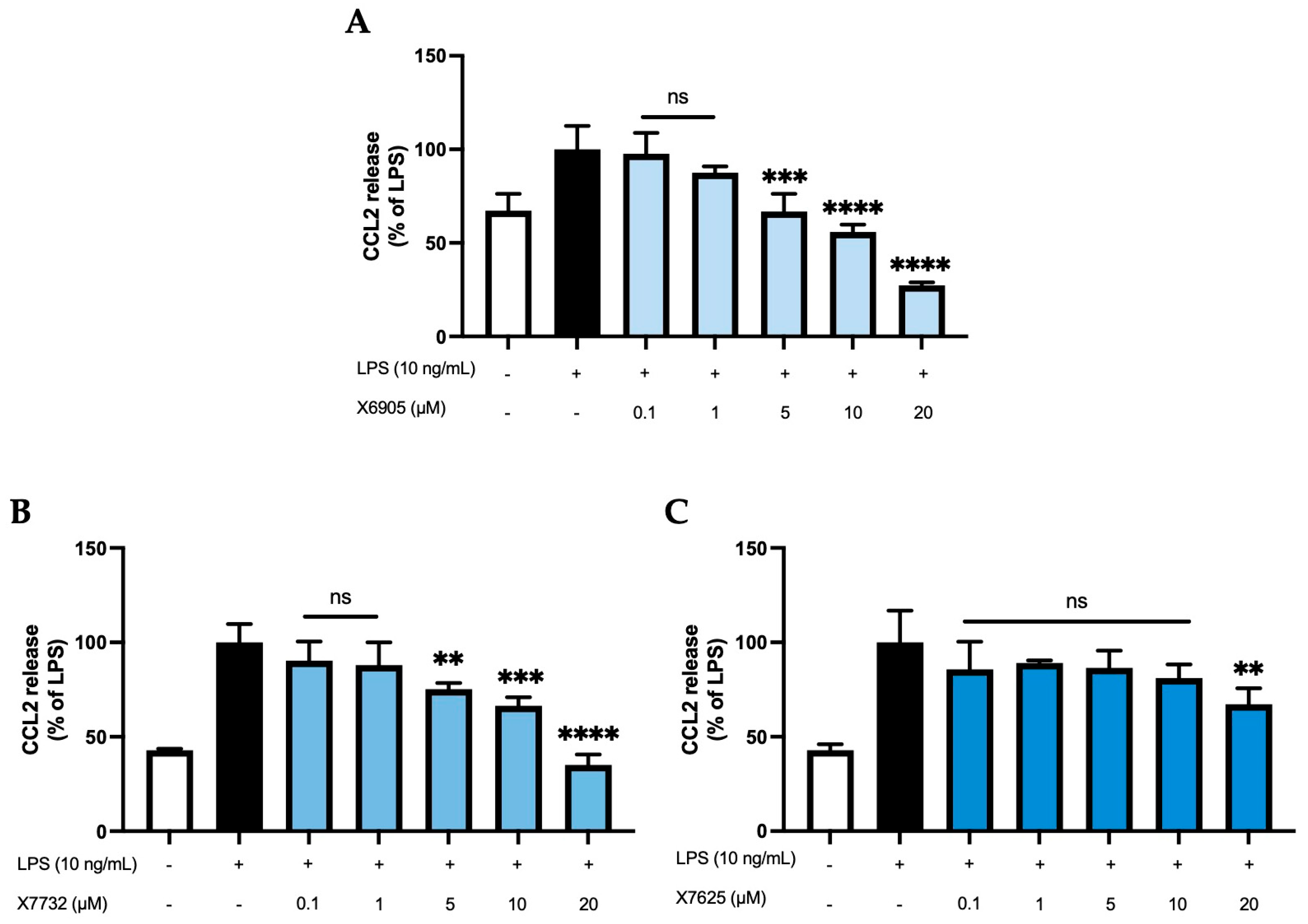
2.2.2. CCL2 Release
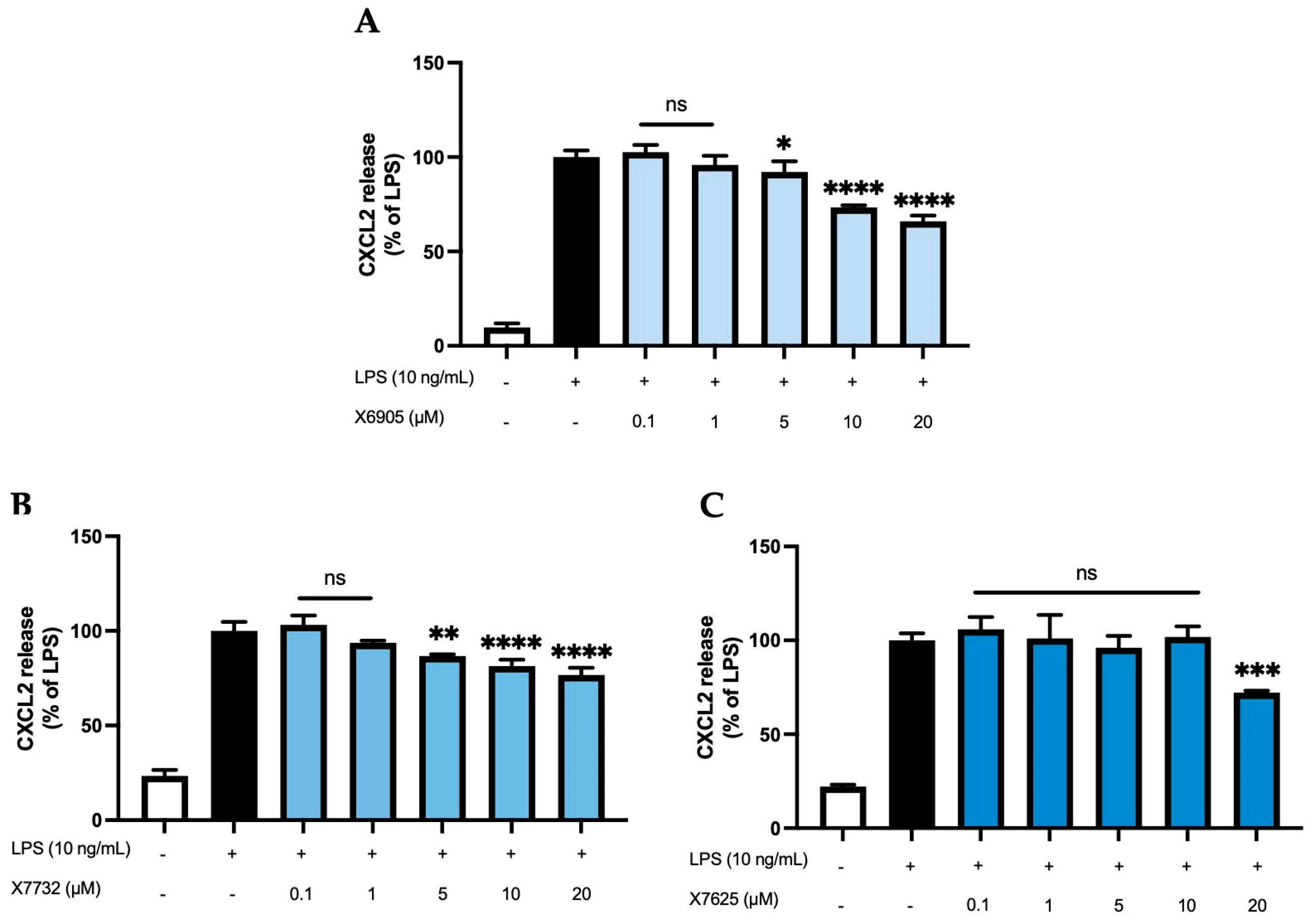
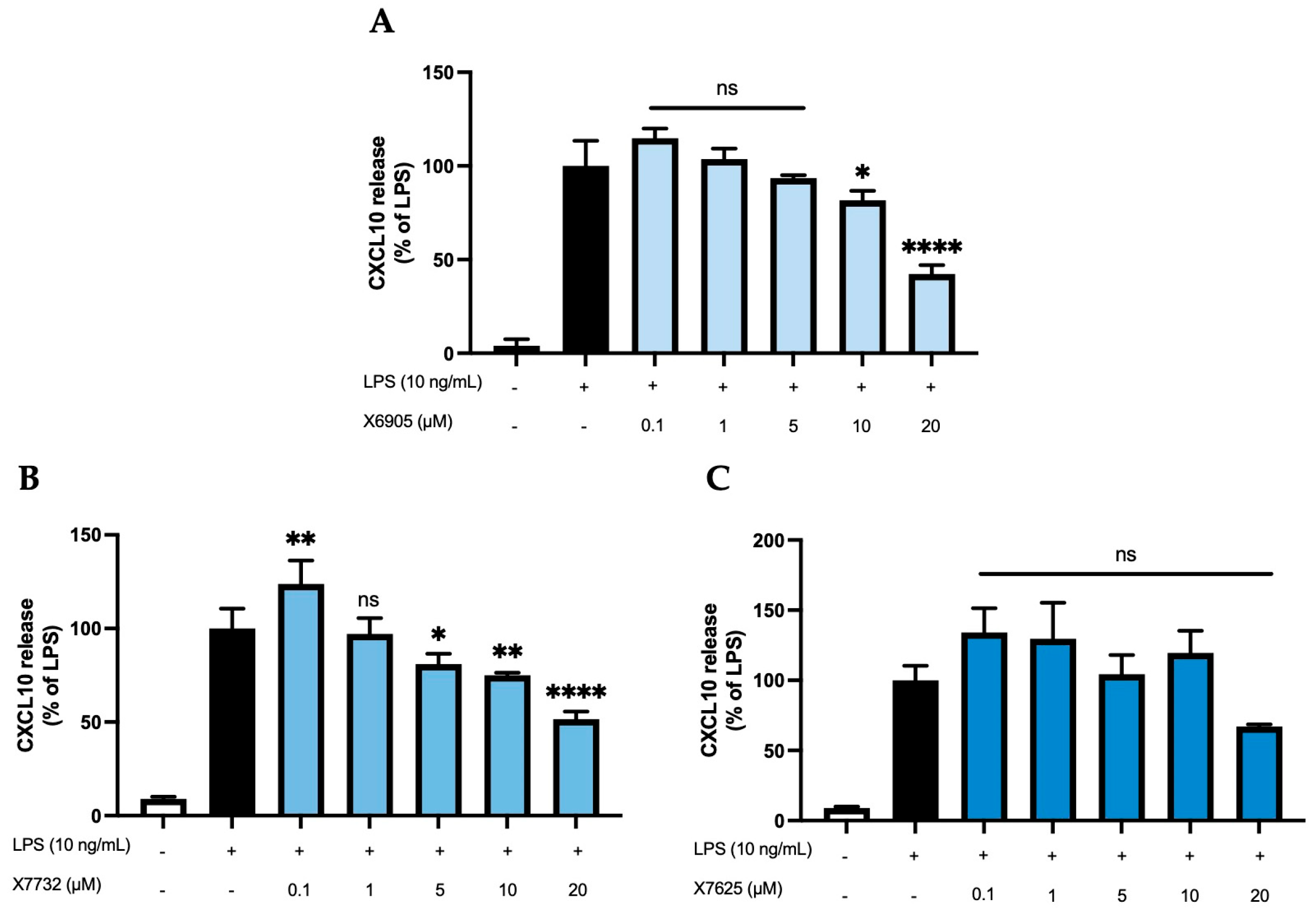
2.2.3. CXCL2 and CXCL10 Release
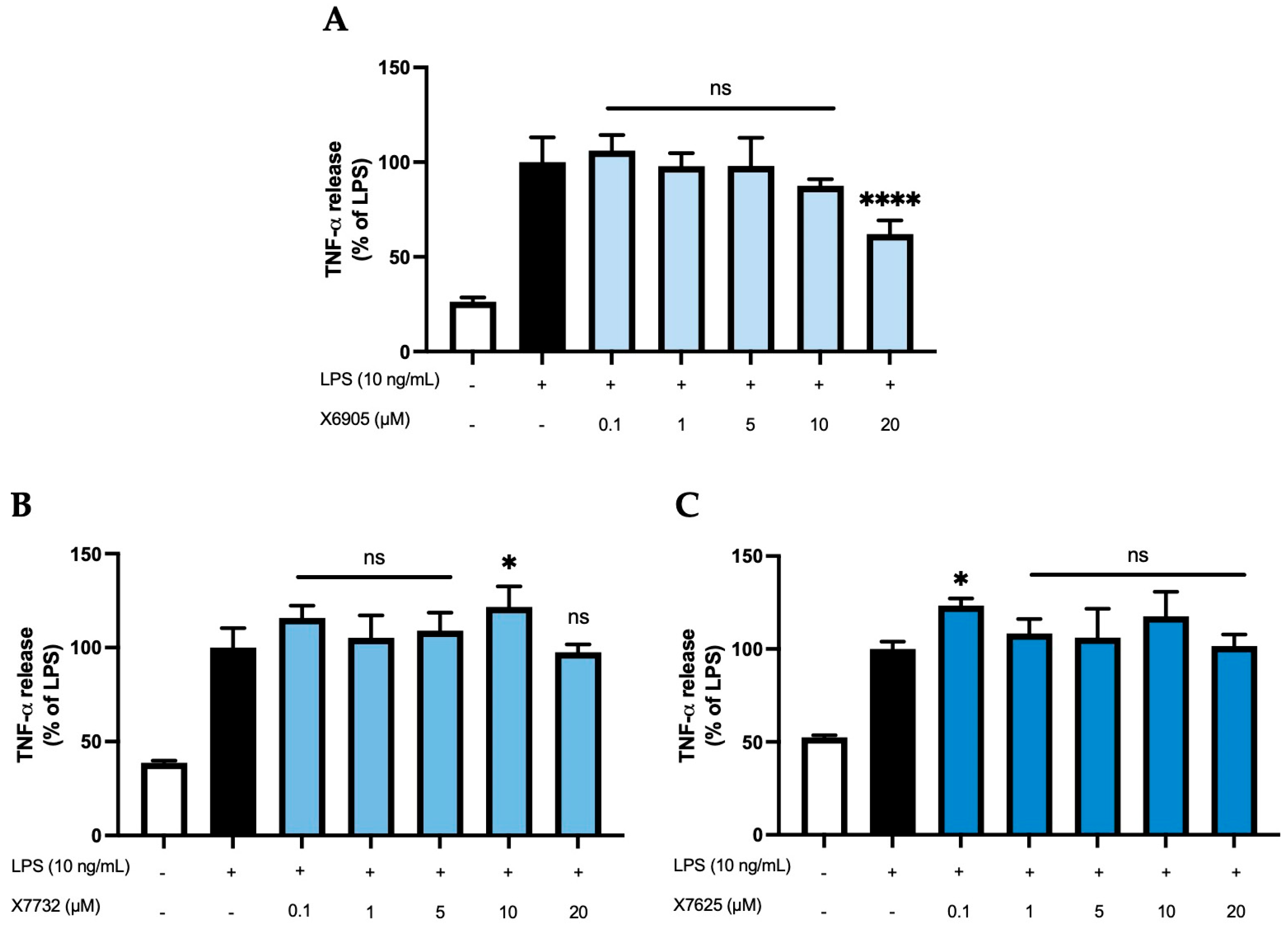
2.2.4. TNF-α Release
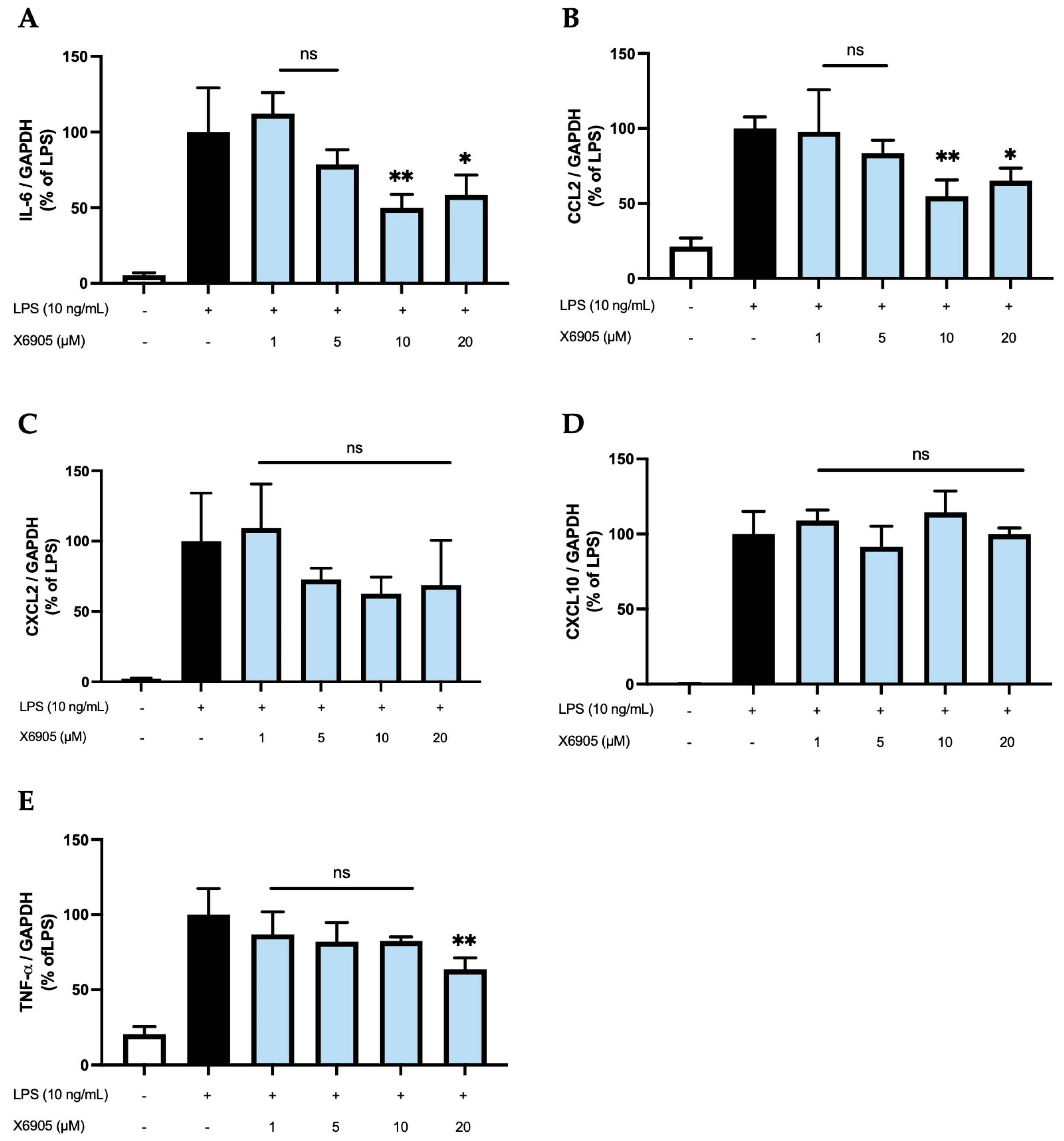
2.3. Effects of X6905 on LPS-Induced Cytokine Expression
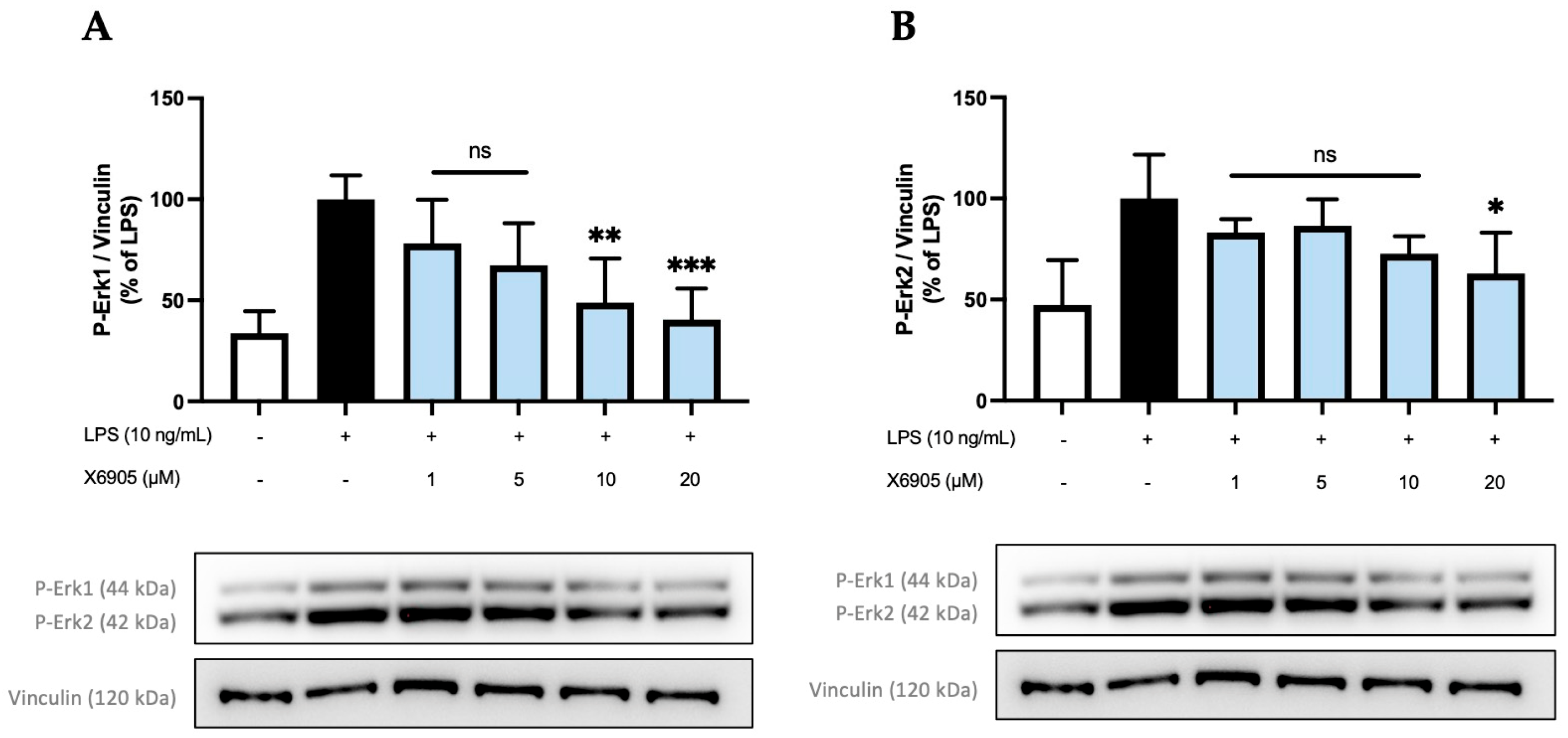
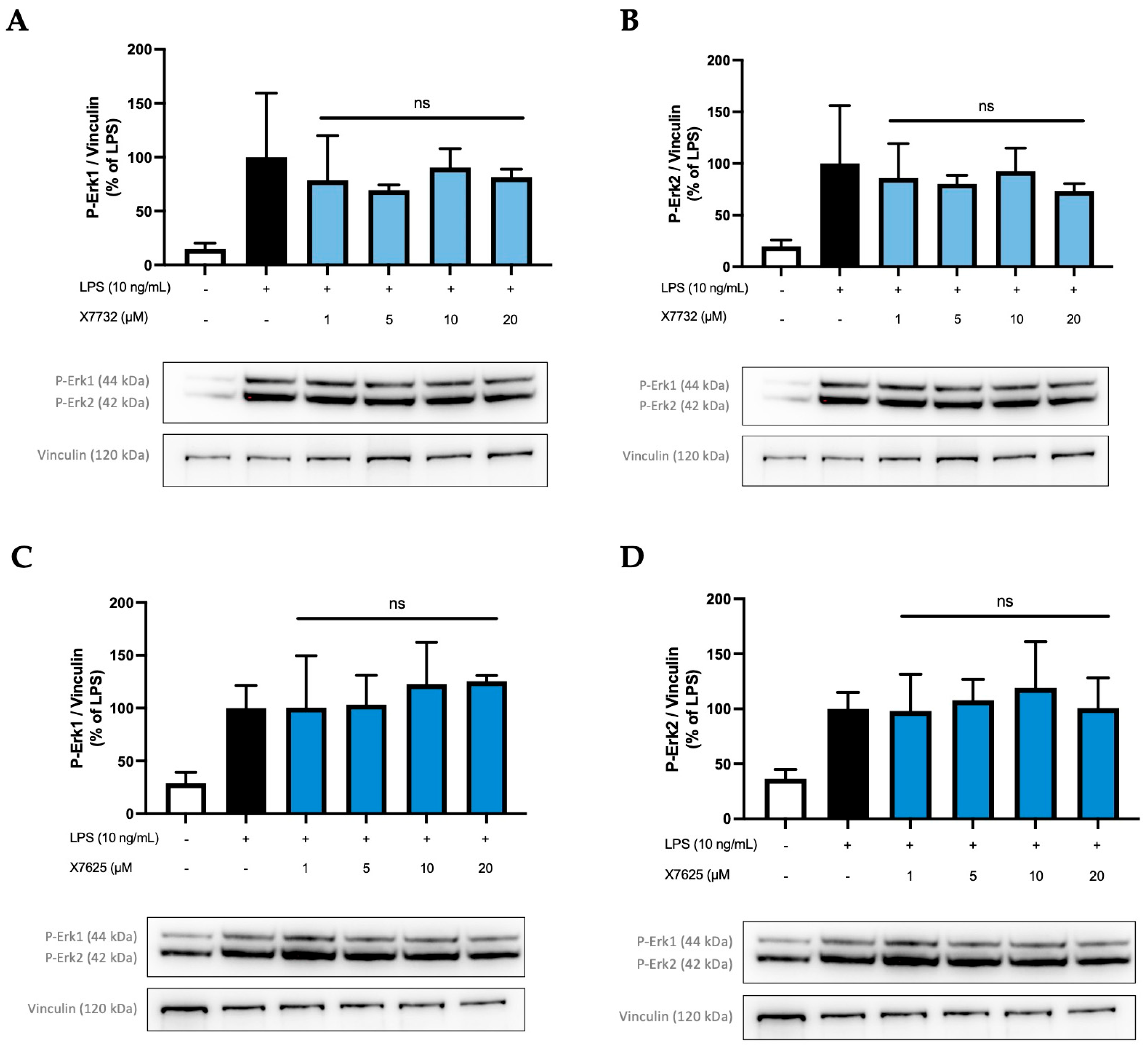
2.4. Effects of X6905, X7732, and X7625 on LPS-Mediated Phosphorylation of Erk1/2
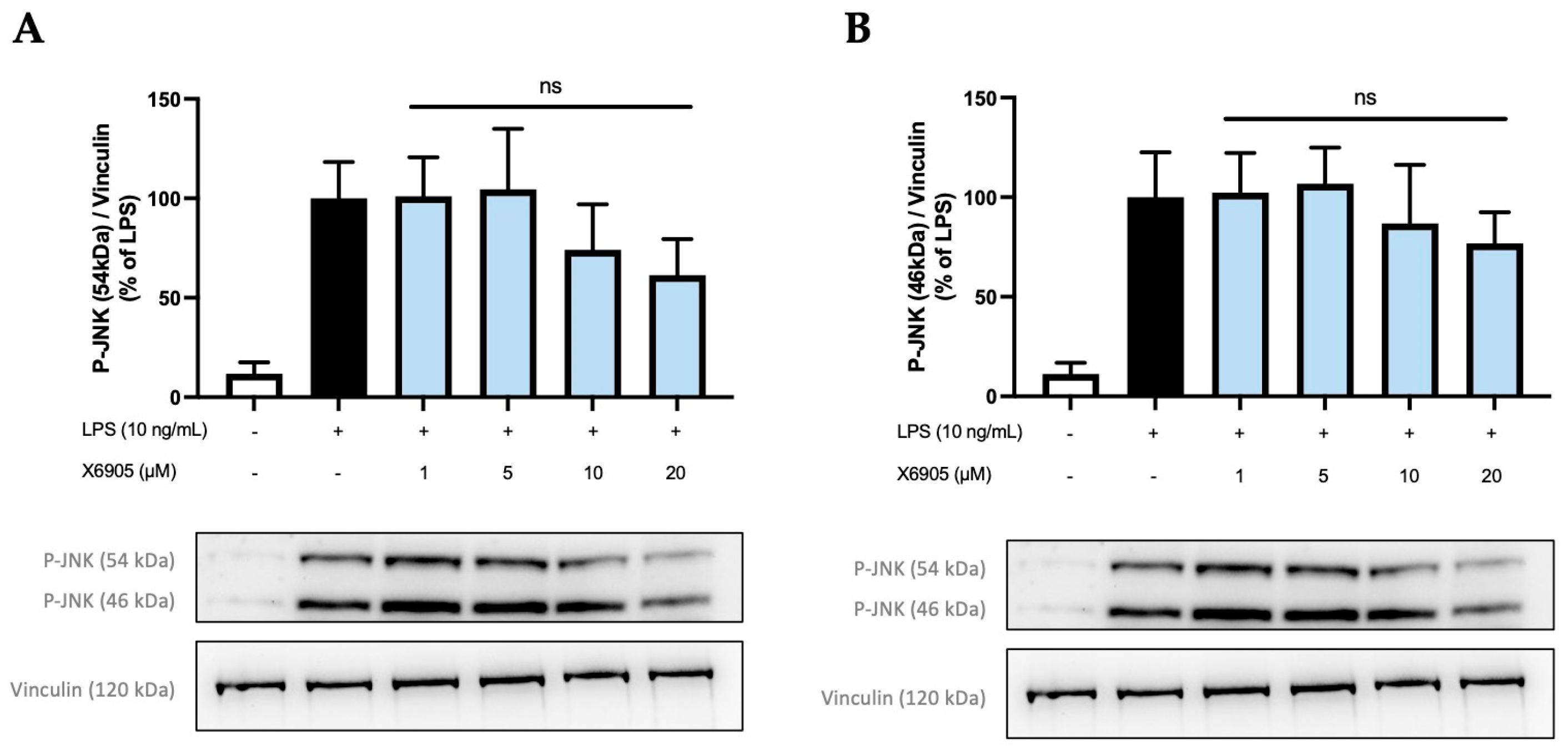
2.5. Effects of X6905 on Other Kinases and Signaling Proteins
3. Discussion
4. Materials and Methods
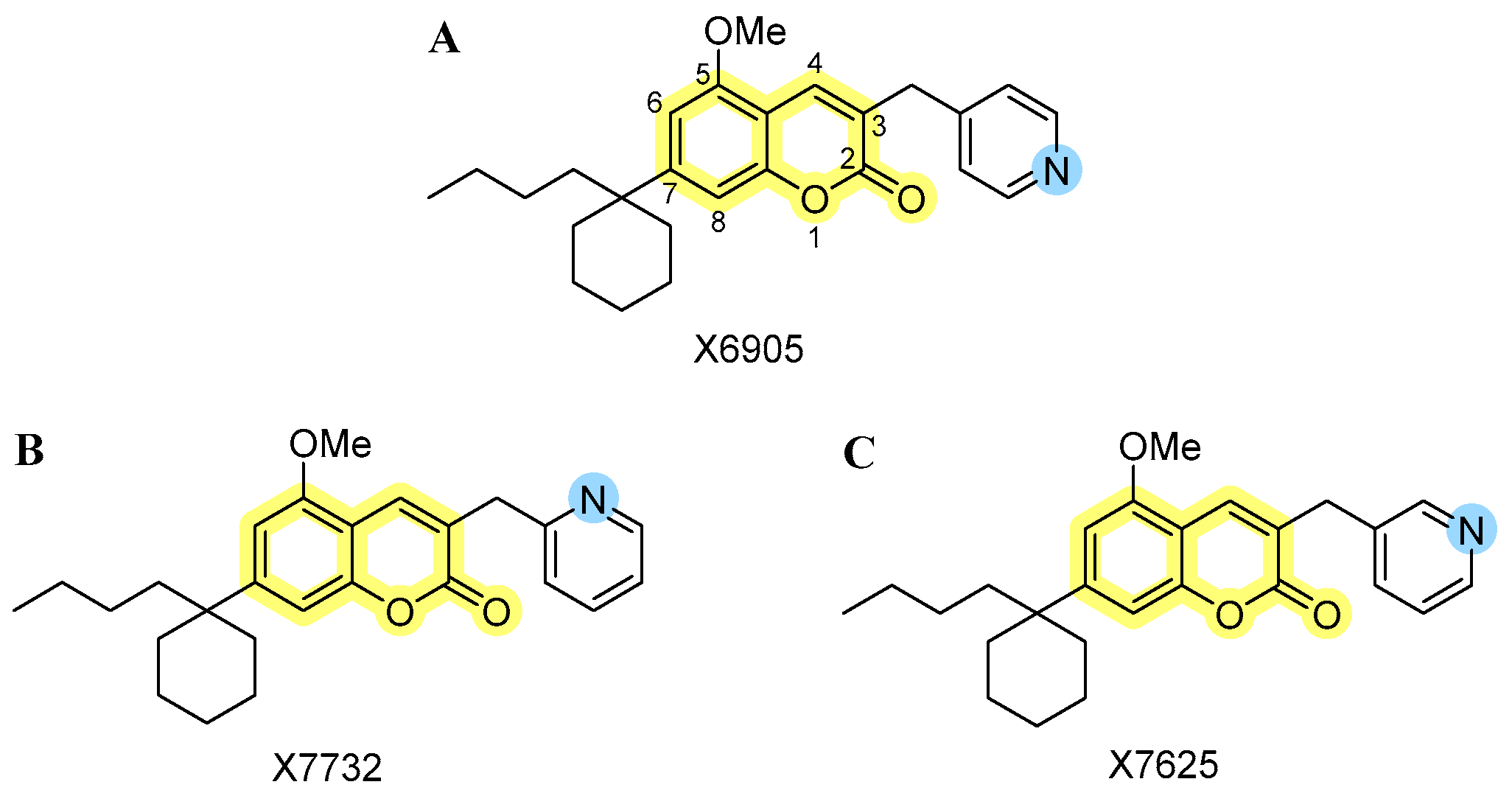
4.1. Novel Coumarin-Derivatives X6905, X7732, and X7625
4.2. BV2 Microglial Cell Culture
4.3. Cell Viability ATP Assay
4.4. Cell Treatment and Determination of Cytokine Release with ELISA
4.5. RNA Isolation and Real-Time PCR
4.6. Electrophoresis and Immunoblotting
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hanley, J.P. Warfarin Reversal. J. Clin. Pathol. 2004, 57, 1132–1139. [Google Scholar] [CrossRef] [PubMed]
- Alshibl, H.M.; Al-Abdullah, E.S.; Alkahtani, H.M. Coumarin: A Promising Scaffold for Design and Development of Bioactive Agents. CBC 2020, 16, 837–852. [Google Scholar] [CrossRef]
- Rempel, V.; Volz, N.; Gläser, F.; Nieger, M.; Bräse, S.; Müller, C.E. Antagonists for the Orphan G-Protein-Coupled Receptor GPR55 Based on a Coumarin Scaffold. J. Med. Chem. 2013, 56, 4798–4810. [Google Scholar] [CrossRef] [PubMed]
- Saliba, S.W.; Jauch, H.; Gargouri, B.; Keil, A.; Hurrle, T.; Volz, N.; Mohr, F.; van der Stelt, M.; Bräse, S.; Fiebich, B.L. Anti-Neuroinflammatory Effects of GPR55 Antagonists in LPS-Activated Primary Microglial Cells. J. Neuroinflammation 2018, 15, 322. [Google Scholar] [CrossRef]
- Apweiler, M.; Saliba, S.W.; Streyczek, J.; Hurrle, T.; Gräßle, S.; Bräse, S.; Fiebich, B.L. Targeting Oxidative Stress: Novel Coumarin-Based Inverse Agonists of GPR55. Int. J. Mol. Sci. 2021, 22, 11665. [Google Scholar] [CrossRef]
- Apweiler, M.; Streyczek, J.; Saliba, S.W.; Collado, J.A.; Hurrle, T.; Gräßle, S.; Muñoz, E.; Normann, C.; Hellwig, S.; Bräse, S.; et al. Functional Selectivity of Coumarin Derivates Acting via GPR55 in Neuroinflammation. Int. J. Mol. Sci. 2022, 23, 959. [Google Scholar] [CrossRef]
- Saliba, S.W.; Gläser, F.; Deckers, A.; Keil, A.; Hurrle, T.; Apweiler, M.; Ferver, F.; Volz, N.; Endres, D.; Bräse, S.; et al. Effects of a Novel GPR55 Antagonist on the Arachidonic Acid Cascade in LPS-Activated Primary Microglial Cells. Int. J. Mol. Sci. 2021, 22, 2503. [Google Scholar] [CrossRef]
- Apweiler, M.; Saliba, S.W.; Streyczek, J.; Normann, C.; Hellwig, S.; Bräse, S.; Fiebich, B.L. Modulation of Neuroinflammation and Oxidative Stress by Targeting GPR55—New Approaches in the Treatment of Psychiatric Disorders. Mol. Psychiatry 2024, 29, 3779–3788. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Liu, J.; Lan, J.-S.; Ding, J.; Sun, Y.; Fang, Y.; Jiang, N.; Yang, Z.; Sun, L.; Jin, Y.; et al. Coumarin-Dithiocarbamate Hybrids as Novel Multitarget AChE and MAO-B Inhibitors against Alzheimer’s Disease: Design, Synthesis and Biological Evaluation. Bioorganic Chem. 2018, 81, 512–528. [Google Scholar] [CrossRef]
- Xiang, X.; Wang, X.; Jin, S.; Hu, J.; Wu, Y.; Li, Y.; Wu, X. Activation of GPR55 Attenuates Cognitive Impairment and Neurotoxicity in a Mouse Model of Alzheimer’s Disease Induced by Aβ1–42 through Inhibiting RhoA/ROCK2 Pathway. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2022, 112, 110423. [Google Scholar] [CrossRef]
- Xiang, X.; Wang, X.; Wu, Y.; Hu, J.; Li, Y.; Jin, S.; Wu, X. Activation of GPR55 Attenuates Cognitive Impairment, Oxidative Stress, Neuroinflammation, and Synaptic Dysfunction in a Streptozotocin-Induced Alzheimer’s Mouse Model. Pharmacol. Biochem. Behav. 2022, 214, 173340. [Google Scholar] [CrossRef]
- He, Y.; Shen, H.; Bi, G.-H.; Zhang, H.-Y.; Soler-Cedeño, O.; Alton, H.; Yang, Y.; Xi, Z.-X. GPR55 Is Expressed in Glutamate Neurons and Functionally Modulates Drug Taking and Seeking in Rats and Mice. Transl. Psychiatry 2024, 14, 101. [Google Scholar] [CrossRef] [PubMed]
- Wróbel, A.; Serefko, A.; Szopa, A.; Ulrich, D.; Poleszak, E.; Rechberger, T. O-1602, an Agonist of Atypical Cannabinoid Receptors GPR55, Reverses the Symptoms of Depression and Detrusor Overactivity in Rats Subjected to Corticosterone Treatment. Front. Pharmacol. 2020, 11, 1002. [Google Scholar] [CrossRef] [PubMed]
- García-Gutiérrez, M.S.; Navarrete, F.; Navarro, G.; Reyes-Resina, I.; Franco, R.; Lanciego, J.L.; Giner, S.; Manzanares, J. Alterations in Gene and Protein Expression of Cannabinoid CB2 and GPR55 Receptors in the Dorsolateral Prefrontal Cortex of Suicide Victims. Neurotherapeutics 2018, 15, 796–806. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, A.; Hajizadeh Moghaddam, A.; Roohbakhsh, A. Central Administration of GPR55 Receptor Agonist and Antagonist Modulates Anxiety-Related Behaviors in Rats. Fundam. Clin. Pharmacol. 2015, 29, 185–190. [Google Scholar] [CrossRef]
- Dimopoulos, N.; Piperi, C.; Psarra, V.; Lea, R.W.; Kalofoutis, A. Increased Plasma Levels of 8-Iso-PGF2α and IL-6 in an Elderly Population with Depression. Psychiatry Res. 2008, 161, 59–66. [Google Scholar] [CrossRef]
- Basterzi, A.D.; Aydemir, Ç.; Kisa, C.; Aksaray, S.; Tuzer, V.; Yazici, K.; Göka, E. IL-6 Levels Decrease with SSRI Treatment in Patients with Major Depression. Hum. Psychopharmacol. Clin. Exp. 2005, 20, 473–476. [Google Scholar] [CrossRef]
- Yoshimura, R.; Hori, H.; Ikenouchi-Sugita, A.; Umene-Nakano, W.; Ueda, N.; Nakamura, J. Higher Plasma Interleukin-6 (IL-6) Level Is Associated with SSRI- or SNRI-Refractory Depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2009, 33, 722–726. [Google Scholar] [CrossRef]
- Lyra e Silva, N.M.; Gonçalves, R.A.; Pascoal, T.A.; Lima-Filho, R.A.S.; Resende, E.d.P.F.; Vieira, E.L.M.; Teixeira, A.L.; de Souza, L.C.; Peny, J.A.; Fortuna, J.T.S.; et al. Pro-Inflammatory Interleukin-6 Signaling Links Cognitive Impairments and Peripheral Metabolic Alterations in Alzheimer’s Disease. Transl. Psychiatry 2021, 11, 251. [Google Scholar] [CrossRef]
- Di Vincenzo, M.; Pellegrino, P.; Schiappa, G.; Campanati, A.; Del Vescovo, V.; Piccirillo, S.; Ambrogini, P.; Arnaldi, G.; Orciani, M. Role of 11β-Hydroxysteroid Dehydrogenase and Mineralocorticoid Receptor on Alzheimer’s Disease Onset: A Systematic Review. Int. J. Mol. Sci. 2025, 26, 1357. [Google Scholar] [CrossRef]
- Knezevic, E.; Nenic, K.; Milanovic, V.; Knezevic, N.N. The Role of Cortisol in Chronic Stress, Neurodegenerative Diseases, and Psychological Disorders. Cells 2023, 12, 2726. [Google Scholar] [CrossRef] [PubMed]
- Raven, P.W.; Taylor, N.F. 11β-HSD and 17β-HSD as Biological Markers of Depression: Sex Differences and Correlation with Symptom Severity. Endocr. Res. 1998, 24, 659–662. [Google Scholar] [CrossRef]
- Seckl, J. 11β-Hydroxysteroid Dehydrogenase and the Brain: Not (yet) Lost in Translation. J. Intern. Med. 2024, 295, 20–37. [Google Scholar] [CrossRef]
- Dodd, S.; Skvarc, D.R.; Dean, O.M.; Anderson, A.; Kotowicz, M.; Berk, M. Effect of Glucocorticoid and 11β-Hydroxysteroid-Dehydrogenase Type 1 (11β-HSD1) in Neurological and Psychiatric Disorders. Int. J. Neuropsychopharmacol. 2022, 25, 387–398. [Google Scholar] [CrossRef]
- Cherry, J.D.; Olschowka, J.A.; O’Banion, M.K. Neuroinflammation and M2 Microglia: The Good, the Bad, and the Inflamed. J. Neuroinflammation 2014, 11, 98. [Google Scholar] [CrossRef] [PubMed]
- Romero-Sanchiz, P.; Nogueira-Arjona, R.; Araos, P.; Serrano, A.; Barrios, V.; Argente, J.; Garcia-Marchena, N.; Lopez-Tellez, A.; Rodriguez-Moreno, S.; Mayoral, F.; et al. Variation in Chemokines Plasma Concentrations in Primary Care Depressed Patients Associated with Internet-Based Cognitive-Behavioral Therapy. Sci. Rep. 2020, 10, 1078. [Google Scholar] [CrossRef] [PubMed]
- Rocha, N.P.; Scalzo, P.L.; Barbosa, I.G.; Souza, M.S.; Morato, I.B.; Vieira, E.L.M.; Christo, P.P.; Teixeira, A.L.; Reis, H.J. Cognitive Status Correlates with CXCL10/IP-10 Levels in Parkinson’s Disease. Park. Dis. 2014, 2014, 903796. [Google Scholar] [CrossRef] [PubMed]
- de la Peña, F.R.; Cruz-Fuentes, C.; Palacios, L.; Girón-Pérez, M.I.; Medina-Rivero, E.; Ponce-Regalado, M.D.; Alvarez-Herrera, S.; Pérez-Sánchez, G.; Becerril-Villanueva, E.; Maldonado-García, J.L.; et al. Serum Levels of Chemokines in Adolescents with Major Depression Treated with Fluoxetine. World J. Psychiatry 2020, 10, 175–186. [Google Scholar] [CrossRef]
- Wong, M.-L.; Dong, C.; Maestre-Mesa, J.; Licinio, J. Polymorphisms in Inflammation-Related Genes Are Associated with Susceptibility to Major Depression and Antidepressant Response. Mol. Psychiatry 2008, 13, 800–812. [Google Scholar] [CrossRef]
- Steiner, J.; Prüss, H.; Köhler, S.; Frodl, T.; Hasan, A.; Falkai, P. Autoimmune Encephalitis with Psychosis: Warning Signs, Step-by-Step Diagnostics and Treatment. World J. Biol. Psychiatry 2020, 21, 241–254. [Google Scholar] [CrossRef]
- Yuan, N.; Chen, Y.; Xia, Y.; Dai, J.; Liu, C. Inflammation-Related Biomarkers in Major Psychiatric Disorders: A Cross-Disorder Assessment of Reproducibility and Specificity in 43 Meta-Analyses. Transl. Psychiatry 2019, 9, 233. [Google Scholar] [CrossRef] [PubMed]
- Köhler-Forsberg, O.N.; Lydholm, C.; Hjorthøj, C.; Nordentoft, M.; Mors, O.; Benros, M.E. Efficacy of Anti-inflammatory Treatment on Major Depressive Disorder or Depressive Symptoms: Meta-analysis of Clinical Trials. Acta Psychiatr. Scand. 2019, 139, 404–419. [Google Scholar] [CrossRef] [PubMed]
- Kopschina Feltes, P.; Doorduin, J.; Klein, H.C.; Juárez-Orozco, L.E.; Dierckx, R.A.; Moriguchi-Jeckel, C.M.; de Vries, E.F. Anti-Inflammatory Treatment for Major Depressive Disorder: Implications for Patients with an Elevated Immune Profile and Non-Responders to Standard Antidepressant Therapy. J. Psychopharmacol. 2017, 31, 1149–1165. [Google Scholar] [CrossRef]
- Aslantürk, Ö.S. In Vitro Cytotoxicity and Cell Viability Assays: Principles, Advantages, and Disadvantages. In Genotoxicity—A Predictable Risk to Our Actual World; Larramendy, M.L., Soloneski, S., Eds.; InTech: London, UK, 2018; ISBN 978-1-78923-418-3. [Google Scholar]
- Nomura, J.; So, A.; Tamura, M.; Busso, N. Intracellular ATP Decrease Mediates NLRP3 Inflammasome Activation upon Nigericin and Crystal Stimulation. J. Immunol. 2015, 195, 5718–5724. [Google Scholar] [CrossRef]
- Shiratori, R.; Furuichi, K.; Yamaguchi, M.; Miyazaki, N.; Aoki, H.; Chibana, H.; Ito, K.; Aoki, S. Glycolytic Suppression Dramatically Changes the Intracellular Metabolic Profile of Multiple Cancer Cell Lines in a Mitochondrial Metabolism-Dependent Manner. Sci. Rep. 2019, 9, 18699. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, K.; Komori, T.; Ishibashi, K.; Uto, T.; Kobayashi, I.; Kadokawa, R.; Kato, Y.; Ninomiya, K.; Takahashi, K.; Hirata, E. Non-Aqueous, Zwitterionic Solvent as an Alternative for Dimethyl Sulfoxide in the Life Sciences. Commun. Chem. 2020, 3, 163. [Google Scholar] [CrossRef]
- Shore, D.M.; Reggio, P.H. The Therapeutic Potential of Orphan GPCRs, GPR35 and GPR55. Front. Pharmacol. 2015, 6, 69. [Google Scholar] [CrossRef]
- Mohr, F.; Hurrle, T.; Burggraaff, L.; Langer, L.; Bemelmans, M.P.; Knab, M.; Nieger, M.; van Westen, G.J.P.; Heitman, L.H.; Bräse, S. Synthesis and SAR Evaluation of Coumarin Derivatives as Potent Cannabinoid Receptor Agonists. Eur. J. Med. Chem. 2021, 220, 113354. [Google Scholar] [CrossRef]
- Henstridge, C.M.; Balenga, N.A.; Schröder, R.; Kargl, J.K.; Platzer, W.; Martini, L.; Arthur, S.; Penman, J.; Whistler, J.L.; Kostenis, E.; et al. GPR55 Ligands Promote Receptor Coupling to Multiple Signalling Pathways. Br. J. Pharmacol. 2010, 160, 604–614. [Google Scholar] [CrossRef]
- Lucas, R.M.; Luo, L.; Stow, J.L. ERK1/2 in Immune Signalling. Biochem. Soc. Trans. 2022, 50, 1341–1352. [Google Scholar] [CrossRef]
- Leonard, M.; Ryan, M.P.; Watson, A.J.; Schramek, H.; Healy, E. Role of MAP Kinase Pathways in Mediating IL-6 Production in Human Primary Mesangial and Proximal Tubular Cells. Kidney Int. 1999, 56, 1366–1377. [Google Scholar] [CrossRef] [PubMed]
- Buscà, R.; Pouysségur, J.; Lenormand, P. ERK1 and ERK2 Map Kinases: Specific Roles or Functional Redundancy? Front. Cell Dev. Biol. 2016, 4, 53. [Google Scholar] [CrossRef] [PubMed]
- Kulesza, A.; Zielniok, K.; Hawryluk, J.; Paczek, L.; Burdzinska, A. Ibuprofen in Therapeutic Concentrations Affects the Secretion of Human Bone Marrow Mesenchymal Stromal Cells, but Not Their Proliferative and Migratory Capacity. Biomolecules 2022, 12, 287. [Google Scholar] [CrossRef] [PubMed]
- Drevets, W.C.; Wittenberg, G.M.; Bullmore, E.T.; Manji, H.K. Immune Targets for Therapeutic Development in Depression: Towards Precision Medicine. Nat. Rev. Drug Discov. 2022, 21, 224–244. [Google Scholar] [CrossRef]
- Treutlein, E.-M.; Kern, K.; Weigert, A.; Tarighi, N.; Schuh, C.-D.; Nüsing, R.M.; Schreiber, Y.; Ferreirós, N.; Brüne, B.; Geisslinger, G.; et al. The Prostaglandin E2 Receptor EP3 Controls CC-Chemokine Ligand 2–Mediated Neuropathic Pain Induced by Mechanical Nerve Damage. J. Biol. Chem. 2018, 293, 9685–9695. [Google Scholar] [CrossRef]
- Knight, J.M.; Costanzo, E.S.; Singh, S.; Yin, Z.; Szabo, A.; Pawar, D.S.; Hillard, C.J.; Rizzo, J.D.; D’Souza, A.; Pasquini, M.; et al. The IL-6 Antagonist Tocilizumab Is Associated with Worse Depression and Related Symptoms in the Medically Ill. Transl. Psychiatry 2021, 11, 58. [Google Scholar] [CrossRef]
- Bavaresco, D.V.; Uggioni, M.L.R.; Ferraz, S.D.; Marques, R.M.M.; Simon, C.S.; Dagostin, V.S.; Grande, A.J.; Da Rosa, M.I. Efficacy of Infliximab in Treatment-Resistant Depression: A Systematic Review and Meta-Analysis. Pharmacol. Biochem. Behav. 2020, 188, 172838. [Google Scholar] [CrossRef]
- Baghdadi, L. Tocilizumab Reduces Depression Risk in Rheumatoid Arthritis Patients: A Systematic Review and Meta-Analysis. Psychol. Res. Behav. Manag. 2024, 17, 3419–3441. [Google Scholar] [CrossRef]
- Kohler, O.; Krogh, J.; Mors, O.; Benros, M.E. Inflammation in Depression and the Potential for Anti-Inflammatory Treatment. Curr. Neuropharmacol. 2016, 14, 732–742. [Google Scholar] [CrossRef]
- Stuart, M.J.; Singhal, G.; Baune, B.T. Systematic Review of the Neurobiological Relevance of Chemokines to Psychiatric Disorders. Front. Cell Neurosci. 2015, 9, 357. [Google Scholar] [CrossRef]
- Bräse, S.; Gläser, F.; Hurrle, T. Chapter 11. Coumarins. In Drug Discovery; Bräse, S., Ed.; Royal Society of Chemistry: Cambridge, UK, 2015; pp. 287–311. ISBN 978-1-78262-030-3. [Google Scholar]
- Bräse, S. (Ed.) Privileged Scaffolds in Medicinal Chemistry: Design, Synthesis, Evaluation; Drug Discovery; Royal Society of Chemistry: Cambridge, UK, 2015; ISBN 978-1-78262-030-3. [Google Scholar]
- Röttger, C.; Jung, N.; Bräse, S. Chemotion Repository Homepage. Available online: https://www.chemotion-repository.net/home/publications/collections/9374 (accessed on 2 June 2025). [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schulzki, R.; Apweiler, M.; Röttger, C.; Grathwol, C.W.; Struchtrup, N.; Abou El Mirate, S.; Normann, C.; Bräse, S.; Fiebich, B.L. Isomeric 3-Pyridinylmethylcoumarins Differ in Erk1/2-Inhibition and Modulation of BV2 Microglia-Mediated Neuroinflammation. Molecules 2025, 30, 2452. https://doi.org/10.3390/molecules30112452
Schulzki R, Apweiler M, Röttger C, Grathwol CW, Struchtrup N, Abou El Mirate S, Normann C, Bräse S, Fiebich BL. Isomeric 3-Pyridinylmethylcoumarins Differ in Erk1/2-Inhibition and Modulation of BV2 Microglia-Mediated Neuroinflammation. Molecules. 2025; 30(11):2452. https://doi.org/10.3390/molecules30112452
Chicago/Turabian StyleSchulzki, Rami, Matthias Apweiler, Caroline Röttger, Christoph W. Grathwol, Nora Struchtrup, Sophia Abou El Mirate, Claus Normann, Stefan Bräse, and Bernd L. Fiebich. 2025. "Isomeric 3-Pyridinylmethylcoumarins Differ in Erk1/2-Inhibition and Modulation of BV2 Microglia-Mediated Neuroinflammation" Molecules 30, no. 11: 2452. https://doi.org/10.3390/molecules30112452
APA StyleSchulzki, R., Apweiler, M., Röttger, C., Grathwol, C. W., Struchtrup, N., Abou El Mirate, S., Normann, C., Bräse, S., & Fiebich, B. L. (2025). Isomeric 3-Pyridinylmethylcoumarins Differ in Erk1/2-Inhibition and Modulation of BV2 Microglia-Mediated Neuroinflammation. Molecules, 30(11), 2452. https://doi.org/10.3390/molecules30112452