
Efficiency of Resistant Starch and Dextrins as Prebiotics: A Review of the Existing Evidence and Clinical Trials



Abstract
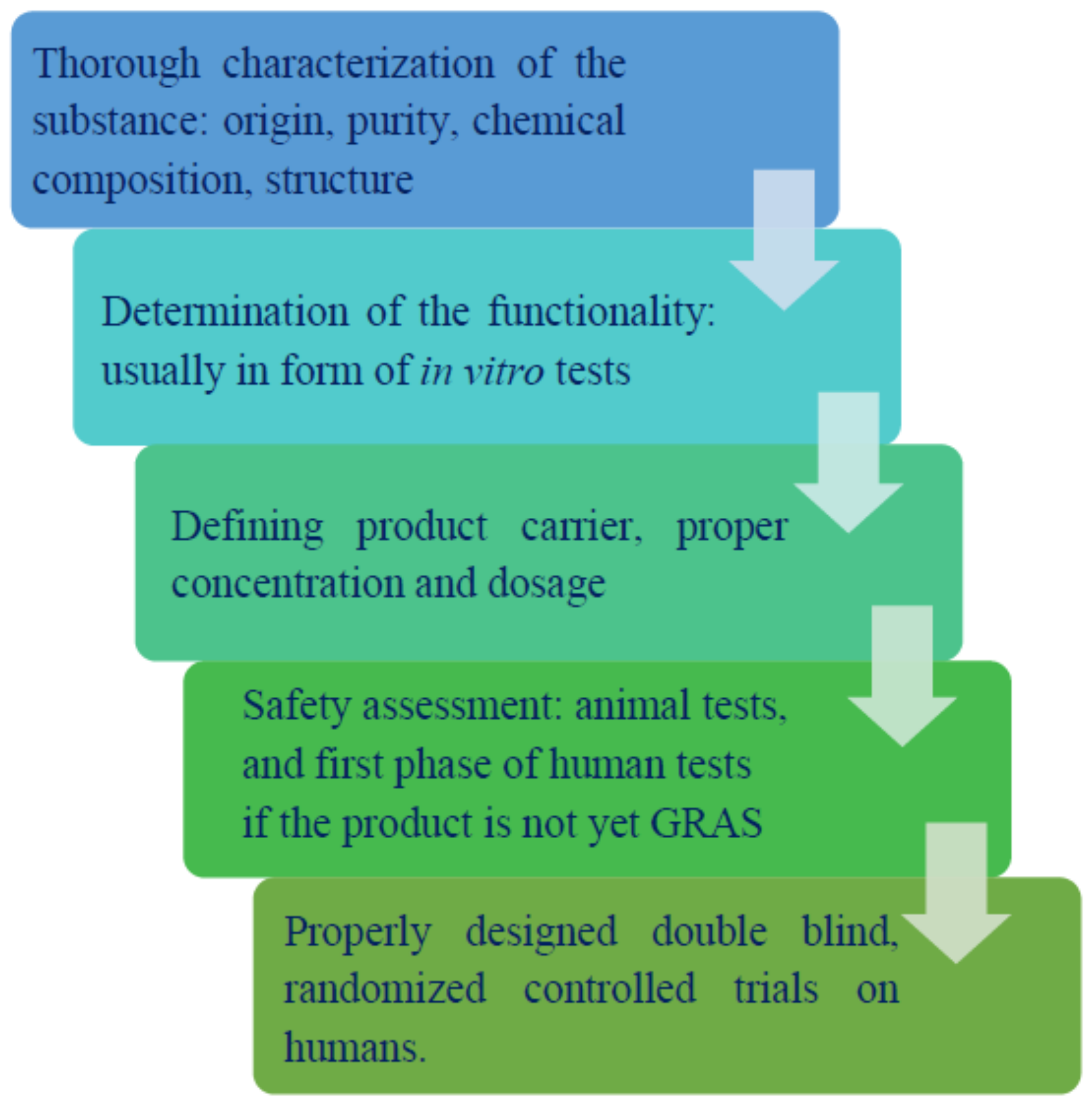
1. Prebiotics
2. Dietary Fibers
- ➢
- reduce the stool transit time, while increasing its bulk [33];
- ➢
- be used as a carbon source by colonic microbiota (as fermentation substrate) [34];
- ➢
- reduce the levels of cholesterol (either total or LDL cholesterol) [35]; and
- ➢
- support glucose and insulin metabolism (reduce the levels of blood glucose and insulin) [36].
- ➢
- non-starch polysaccharides including cellulose, hemicelluloses, mannans, pectins, and other hydrocolloids (i.e., b-glucans, gums, and mucilages), inulin, and fructans;
- ➢
- resistant oligosaccharides including fructo-oligosaccharides (FOS), galactooligosaccharides (GOS);
- ➢
- resistant starch and dextrins [30].
3. Short-Chain Fatty Acids
3.1. Effects of SCFA Absorption in the Human Colon
3.2. SCFA Receptors
3.3. Influence of SCFA on Gut-Brain Axis
3.4. Functions of SCFA in the Liver
3.4.1. Resistant Dextrins in Clinical Trials
3.4.2. Overview of Resistant Starch
RS1 = TS − (RDS + SDS) − RS2 − RS3
RS2 = TS − (RDS + SDS) − RS1 − RS3
RS3 = TS − (RDS + SDS) − RS1 − RS2
3.4.3. Resistant Starch Type I
3.4.4. Resistant Starch Type II
3.4.5. Resistant Starch Type III
3.4.6. Resistant Starch Type IV
3.4.7. Resistant Starch Type 5
3.4.8. Resistant Starch in Clinical Trials
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Hutkins, R.W.; Krumbeck, J.A.; Bindels, L.B.; Cani, P.D.; Fahey, G., Jr.; Goh, Y.J.; Hamaker, B.; Martens, E.C.; Mills, D.A.; Rastal, R.A.; et al. Prebiotics: Why definitions matter. Curr. Opin. Biotechnol. 2016, 37, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Markowiak, P.; Śliżewska, K. Effects of probiotics, prebiotics, and synbiotics on human health. Nutrients 2017, 9, 1021. [Google Scholar] [CrossRef]
- Gibson, G.R.; Hutkins, R.; Sanders, M.E.; Prescott, S.L.; Reimer, R.A.; Salminen, S.J.; Scott, K.; Stanton, C.; Swanson, K.S.; Cani, P.D.; et al. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 491–502. [Google Scholar] [CrossRef]
- Slavin, J. Fiber and Prebiotics: Mechanisms and Health Benefits. Nutrients 2013, 5, 1417–1435. [Google Scholar] [CrossRef]
- Rajendran, S.R.C.K.; Okolie, C.L.; Udenigwe, C.C.; Mason, B. Structural features underlying prebiotic activity of conventional and potential prebiotic oligosaccharides in food and health. J. Food Biochem. 2017, 41, e12389. [Google Scholar] [CrossRef]
- Zaman, S.A.; Sarbini, S.R. The potential of resistant starch as a prebiotic. Crit. Rev. Biotechnol. 2016, 36, 578–584. [Google Scholar] [CrossRef] [PubMed]
- Gómez, B.; Gullón, B.; Remoroza, C.; Schols, H.A.; Parajó, J.C.; Alonso, J.L. Purification, Characterization, and Prebiotic Properties of Pectic Oligosaccharides from Orange Peel Wastes. J. Agric. Food Chem. 2014, 62, 9769–9782. [Google Scholar] [CrossRef] [PubMed]
- Schrezenmeir, J.; de Vrese, M. Probiotics, prebiotics, and synbiotics—Approaching a definition. Am. J. Clin. Nutr. 2001, 73, 361S–364S. [Google Scholar] [CrossRef] [PubMed]
- Carlson, J.L.; Erickson, J.M.; Lloyd, B.B.; Slavin, J.L. Health Effects and Sources of Prebiotic Dietary Fiber. Curr. Dev. Nutr. 2018, 2, nzy005. [Google Scholar] [CrossRef]
- Delcour, J.A.; Aman, P.; Courtin, C.M.; Hamaker, B.R.; Verbeke, K. Prebiotics, fermentable dietary fiber, and health claims. Adv. Nutr. 2016, 7, 1–4. [Google Scholar] [CrossRef]
- Torres-Fuentes, C.; Golubeva, A.V.; Zhdanov, A.V.; Wallace, S.; Arboleya, S.; Papkovsky, D.B.; Aidy, S.; El Ross, P.; Roy, B.L.; Stanton, C.; et al. Short-chain fatty acids and microbiota metabolites attenuate ghrelin receptor signaling. FASEB J. 2019, 33, 13546–13559. [Google Scholar] [CrossRef] [PubMed]
- Lamsal, B.P. Production, health aspects and potential food uses of dairy prebiotic galactooligosaccharides. J. Sci. Food Agric. 2012, 92, 2020–2028. [Google Scholar] [CrossRef]
- Byrne, C.; Chambers, E.; Morrison, D.; Frost, G. The role of short chain fatty acids in appetite regulation and energy homeostasis. Int. J. Obes. 2015, 39, 1331–1338. [Google Scholar] [CrossRef] [PubMed]
- Cai, T.; Gallelli, L.; Cione, E.; Perletti, G.; Ciarleglio, F.; Malossini, G.; De Pretis, G.; Palmieri, A.; Mirone, V.; Bartoletti, R.; et al. The use of Lactobacillus casei DG® prevents symptomatic episodes and reduces the antibiotic use in patients affected by chronic bacterial prostatitis: Results from a phase IV study. World J. Urol. 2021, 39, 3433–3440. [Google Scholar] [CrossRef] [PubMed]
- Costa, G.; Vasconcelos, Q.; Abreu, G.; Albuquerque, A.; Vilarejo, J.; Aragão, G. Changes in nutrient absorption in children and adolescents caused by fructans, especially fructooligosaccharides and inulin. Arch. Pédiatrie 2020, 27, 166–169. [Google Scholar] [CrossRef]
- Peredo-Lovillo, A.; Romero-Luna, H.E.; Jiménez-Fernández, M. Health promoting microbial metabolites produced by gut microbiota after prebiotics metabolism. Food Res. Int. 2020, 136, 109473. [Google Scholar] [CrossRef]
- Skrypnik, K.; Suliburska, J. Association between the gut microbiota and mineral metabolism. J. Sci. Food Agric. 2018, 98, 2449–2460. [Google Scholar] [CrossRef]
- Bergandi, L.; Apprato, G.; Silvagno, F. Vitamin D and Beta-Glucans Synergically Stimulate Human Macrophage Activity. Int. J. Mol. Sci. 2021, 22, 4869. [Google Scholar] [CrossRef] [PubMed]
- Maldonado Galdeano, C.; Cazorla, S.I.; Lemme Dumit, J.M.; Vélez, E.; Perdigón, G. Beneficial effects of probiotic consumption on the immune system. Ann. Nutr. Metab. 2019, 74, 115–124. [Google Scholar] [CrossRef]
- Steimbach, L.; Borgmann, A.V.; Gomar, G.G.; Hoffmann, L.V.; Rutckeviski, R.; de Andrade, D.P.; Smiderle, F.R. Fungal beta-glucans as adjuvants for treating cancer patients—A systematic review of clinical trials. Clin. Nutr. 2021, 40, 3104–3113. [Google Scholar] [CrossRef]
- Lomax, A.R.; Calder, P.C. Prebiotics, immune function, infection and inflammation: A review of the evidence. Br. J. Nutr. 2009, 101, 633–658. [Google Scholar] [CrossRef] [PubMed]
- Van Zyl, W.F.; Deane, S.M.; Dicks, L.M.T. Molecular insights into probiotic mechanisms of action employed against intestinal pathogenic bacteria. Gut Microbes 2020, 12, 1831339. [Google Scholar] [CrossRef]
- An, J.; Ha, E.M. Combination therapy of Lactobacillus plantarum supernatant and 5-fluouracil increases chemosensitivity in colorectal cancer cells. J. Microbiol. Biotechnol. 2016, 26, 1490–1503. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, H.S.; Quigley, E.M.M. Bifidobacteria and Mucosal-Associated Invariant T (MAIT) Cells: A New Approach to Colorectal Cancer Prevention? Gastrointest. Disord. 2019, 1, 266–272. [Google Scholar] [CrossRef]
- Geier, M.S.; Butler, R.N.; Howarth, G.S. Probiotics, prebiotics and synbiotics: A role in chemoprevention for colorectal cancer? Cancer Biol. Ther. 2006, 5, 1265–1269. [Google Scholar] [CrossRef] [PubMed]
- Roberfroid, M.; Gibson, G.R.; Hoyles, L.; McCartney, A.L.; Rastall, R.; Rowland, I.; Wolvers, D.; Watzl, B.; Szajewska, H.; Stahl, B.; et al. Prebiotic effects: Metabolic and health benefits. Br. J. Nutr. 2010, 104 (Suppl. 2), S1–S63. [Google Scholar] [CrossRef] [PubMed]
- Valdés-Varela, L.; Hernández-Barranco, A.M.; Ruas-Madiedo, P.; Gueimonde, M. Effect of Bifidobacterium upon Clostridium difficile Growth and Toxicity When Co-cultured in Different Prebiotic Substrates. Front. Microbiol. 2016, 7, 738. [Google Scholar] [CrossRef]
- Wollowski, I.; Rechkemmer, G.; Pool-Zobel, B.L. Protective role of probiotics and prebiotics in colon cancer. Am. J. Clin. Nutr. 2001, 73, 451s–455s. [Google Scholar] [CrossRef]
- Mundi, M.; Mikal, K.M.; Ahmed, O.H.; Sarbini, S.R. A review on the effects of prebiotics on cell toxicity and integrity. Int. J. Food Prop. 2017, 20 (Suppl. 1), S1045–S1052. [Google Scholar] [CrossRef]
- Stephen, A.M.; Champ, M.M.-J.; Cloran, S.J.; Fleith, M.; van Lieshout, L.; Mejborn, H.; Burley, V.J. Dietary fibre in Europe: Current state of knowledge on definitions, sources, recommendations, intakes and relationships to health. Nutr. Res. Rev. 2017, 30, 149–190. [Google Scholar] [CrossRef]
- Verspreet, J.; Damen, B.; Broekaert, W.F.; Verbeke, K.; Delcour, J.A.; Courtin, C.M. A Critical Look at Prebiotics within the Dietary Fiber Concept. Annu. Rev. Food Sci. Technol. 2016, 7, 167–190. [Google Scholar] [CrossRef] [PubMed]
- FDA. Review of the Scientific Evidence on the Physiological Effects of Certain Non-Digestible Carbohydrates; FDA: Silver Spring, MD, USA, 2018. [Google Scholar]
- Abellán Ruiz, M.S.; Barnuevo Espinosa, M.D.; Contreras Fernández, C.J.; Luque Rubia, A.J.; Sánchez Ayllón, F.; Aldeguer García, M.; García Santamaría, C.; López Román, F.J. Digestion-resistant maltodextrin effects on colonic transit time and stool weight: A randomized controlled clinical study. Eur. J. Nutr. 2016, 55, 2389–2397. [Google Scholar] [CrossRef] [PubMed]
- Burns, A.M.; Solch, R.J.; Dennis-Wall, J.C.; Ukhanova, M.; Nieves, C., Jr.; Mai, V.; Christman, M.C.; Gordon, D.T.; Langkamp-Henken, B. In healthy adults, resistant maltodextrin produces a greater change in fecal bifidobacteria counts and increases stool wet weight: A double-blind, randomized, controlled crossover study. Nutr. Res. 2018, 60, 33–42. [Google Scholar] [CrossRef]
- Halajzadeh, J.; Milajerdi, A.; Reiner, Ž.; Amirani, E.; Kolahdooz, F.; Barekat, M.; Mirzaei, H.; Mirhashemi, S.M.; Asemi, Z. Effects of resistant starch on glycemic control, serum lipoproteins and systemic inflammation in patients with metabolic syndrome and related disorders: A systematic review and meta-analysis of randomized controlled clinical trials. Crit. Rev. Food Sci. Nutr. 2020, 60, 3172–3184. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Yu, H.; Liu, L.; Lu, T.; Li, J.; Ji, Y.; Le, Z.; Bao, L.; Ma, W.; Xiao, R.; et al. Milk powder co-supplemented with inulin and resistant dextrin improves glycemic control and insulin resistance in elderly type 2 diabetes mellitus: A 12-Week randomized, double-blind, placebo-controlled trial. Mol. Nutr. Food Res. 2018, 62, 1800865. [Google Scholar] [CrossRef]
- Health Canada List of Dietary Fibres Reviewed and Accepted by Health Canada’s Food Directorate—Canada.ca. Available online: https://www.canada.ca/en/health-canada/services/publications/food-nutrition/list-reviewed-accepted-dietary-fibres.html (accessed on 12 August 2021).
- McCleary, B.; Sloane, N.; Draga, A.; Lazewska, I. Measurement of Total Dietary Fiber Using AOAC Method 2009.01 (AACC International Approved Method 32-45.01): Evaluation and Updates. Cereal Chem. 2013, 90, 396–414. [Google Scholar] [CrossRef]
- McCleary, B. Importance of Enzyme Purity and Activity in the Measurement of Total Dietary Fiber and Dietary Fiber Components. J. AOAC Int. 2000, 83, 997–1005. [Google Scholar] [CrossRef] [PubMed]
- Jonathan, M.C.; van den Borne, J.J.G.C.; van Wiechen, P.; Souza da Silva, C.; Schols, H.A.; Gruppen, H. In vitro fermentation of 12 dietary fibres by faecal inoculum from pigs and humans. Food Chem. 2012, 133, 889–897. [Google Scholar] [CrossRef]
- Tuncil, Y.E.; Nakatsu, C.H.; Kazem, A.E.; Arioglu-Tuncil, S.; Reuhs, B.; Martens, E.C.; Hamaker, B.R. Delayed utilization of some fast-fermenting soluble dietary fibers by human gut microbiota when presented in a mixture. J. Funct. Foods 2017, 32, 347–357. [Google Scholar] [CrossRef]
- Poutanen, K.S.; Dussort, P.; Erkner, A.; Fiszman, S.; Karnik, K.; Kristensen, M.; Marsaux, C.F.M.; Miquel-Kergoat, S.; Pentikäinen, S.P.; Putz, P.; et al. A review of the characteristics of dietary fibers relevant to appetite and energy intake outcomes in human intervention trials. Am. J. Clin. Nutr. 2017, 106, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Canfora, E.E.; van der Beek, C.M.; Hermes, G.D.A.; Goossens, G.H.; Jocken, J.W.; Holst, J.J.; van Eijk, H.M.; Venema, K.; Smidt, H.; Zoetendal, E.G.; et al. Supplementation of Diet With Galacto-oligosaccharides Increases Bifidobacteria, but Not Insulin Sensitivity, in Obese Prediabetic Individuals. Gastroenterology 2017, 153, 87–97.e3. [Google Scholar] [CrossRef]
- Nicolucci, A.C.; Hume, M.P.; Martínez, I.; Mayengbam, S.; Walter, J.; Reimer, R.A. Prebiotics Reduce Body Fat and Alter Intestinal Microbiota in Children Who Are Overweight or With Obesity. Gastroenterology 2017, 153, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Gulfi, M.; Arrigoni, E.; Amadò, R. The chemical characteristics of apple pectin influence its fermentability in vitro. LWT Food Sci. Technol. 2006, 39, 1001–1004. [Google Scholar] [CrossRef]
- Phillips, K.M.; McGinty, R.C.; Couture, G.; Pehrsson, P.R.; McKillop, K.; Fukagawa, N.K. Dietary fiber, starch, and sugars in bananas at different stages of ripeness in the retail market. PLoS ONE 2021, 16, e0253366. [Google Scholar] [CrossRef] [PubMed]
- Dhingra, D.; Michael, M.; Rajput, H.; Patil, R.T. Dietary fibre in foods: A review. J. Food Sci. Technol. 2011, 49, 255–266. [Google Scholar] [CrossRef]
- Hobden, M.R.; Commane, D.M.; Guérin-Deremaux, L.; Wils, D.; Thabuis, C.; Martin-Morales, A.; Wolfram, S.; Dìaz, A.; Collins, S.; Morais, I.; et al. Impact of dietary supplementation with resistant dextrin (NUTRIOSE®) on satiety, glycaemia, and related endpoints, in healthy adults. Eur. J. Nutr. 2021, 1–9. [Google Scholar] [CrossRef]
- Lefranc-Millot, C.; Guérin-Deremaux, L.; Wils, D.; Neut, C.; Miller, L.; Saniez-Degrave, M. Impact of a Resistant Dextrin on Intestinal Ecology: How Altering the Digestive Ecosystem with NUTRIOSE®, a Soluble Fibre with Prebiotic Properties, May Be Beneficial for Health. J. Int. Med. Res. 2012, 40, 211–224. [Google Scholar] [CrossRef]
- Vulevic, J.; Juric, A.; Tzortzis, G.; Gibson, G.R. A mixture of trans-galactooligosaccharides reduces markers of metabolic syndrome and modulates the fecal microbiota and immune function of overweight adults. J. Nutr. 2013, 143, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Brum, J.; Ramsey, D.; McRorie, J.; Bauer, B.; Kopecky, S.L. Meta-Analysis of Usefulness of Psyllium Fiber as Adjuvant Antilipid Therapy to Enhance Cholesterol Lowering Efficacy of Statins. Am. J. Cardiol. 2018, 122, 1169–1174. [Google Scholar] [CrossRef]
- Jovanovski, E.; Yashpal, S.; Komishon, A.; Zurbau, A.; Mejia, S.B.; Ho, H.V.T.; Li, D.; Sievenpiper, J.; Duvnjak, L.; Vuksan, V. Effect of psyllium (Plantago ovata) fiber on LDL cholesterol and alternative lipid targets, non-HDL cholesterol and apolipoprotein B: A systematic review and meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 2018, 108, 922–932. [Google Scholar] [CrossRef] [PubMed]
- Jane, M.; McKay, J.; Pal, S. Effects of daily consumption of psyllium, oat bran and polyGlycopleX on obesity-related disease risk factors: A critical review. Nutrition 2019, 57, 84–91. [Google Scholar] [CrossRef]
- Marcotuli, I.; Hsieh, Y.S.-Y.; Lahnstein, J.; Yap, K.; Burton, R.A.; Blanco, A.; Fincher, G.B.; Gadaleta, A. Structural Variation and Content of Arabinoxylans in Endosperm and Bran of Durum Wheat (Triticum turgidum L.). J. Agric. Food Chem. 2016, 64, 2883–2892. [Google Scholar] [CrossRef] [PubMed]
- Wilson, B.; Whelan, K. Prebiotic inulin-type fructans and galacto-oligosaccharides: Definition, specificity, function, and application in gastrointestinal disorders. J. Gastroenterol. Hepatol. 2017, 32 (Suppl. 1), 64–68. [Google Scholar] [CrossRef] [PubMed]
- Grootaert, C.; Van den Abbeele, P.; Marzorati, M.; Broekaert, W.F.; Courtin, C.M.; Delcour, J.A.; Verstraete, W.; Van de Wiele, T. Comparison of prebiotic effects of arabinoxylan oligosaccharides and inulin in a simulator of the human intestinal microbial ecosystem. FEMS Microbiol. Ecol. 2009, 69, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Van den Abbeele, P.; Venema, K.; Van de Wiele, T.; Verstraete, W.; Possemiers, S. Different human gut models reveal the distinct fermentation patterns of Arabinoxylan versus inulin. J. Agric. Food Chem. 2013, 61, 9819–9827. [Google Scholar] [CrossRef]
- Van den Abbeele, P.; Taminiau, B.; Pinheiro, I.; Duysburgh, C.; Jacobs, H.; Pijls, L.; Marzorati, M. Arabinoxylo-Oligosaccharides and Inulin Impact Inter-Individual Variation on Microbial Metabolism and Composition, Which Immunomodulates Human Cells. J. Agric. Food Chem. 2018, 66, 1121–1130. [Google Scholar] [CrossRef] [PubMed]
- Geraylou, Z.; Souffreau, C.; Rurangwa, E.; D’Hondt, S.; Callewaert, L.; Courtin, C.M.; Delcour, J.A.; Buyse, J.; Ollevier, F. Effects of arabinoxylan-oligosaccharides (AXOS) on juvenile Siberian sturgeon (Acipenser baerii) performance, immune responses and gastrointestinal microbial community. Fish Shellfish Immunol. 2012, 33, 718–724. [Google Scholar] [CrossRef]
- Lynch, K.M.; Strain, C.R.; Johnson, C.; Patangia, D.; Stanton, C.; Koc, F.; Gil-Martinez, J.; O’Riordan, P.; Sahin, A.W.; Ross, R.P.; et al. Extraction and characterisation of arabinoxylan from brewers spent grain and investigation of microbiome modulation potential. Eur. J. Nutr. 2021. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef]
- Mueller, N.T.; Zhang, M.; Juraschek, S.P.; Miller, E.R., 3rd; Appel, L.J. Effects of high-fiber diets enriched with carbohydrate, protein, or unsaturated fat on circulating short chain fatty acids: Results from the OmniHeart randomized trial. Am. J. Clin. Nutr. 2020, 111, 545–554. [Google Scholar] [CrossRef]
- Wolfram, T.; Ismail-Beigi, F. Efficacy of High-Fiber Diets in the Management of Type 2 Diabetes Mellitus. Endocr. Pract. 2011, 17, 132–142. [Google Scholar] [CrossRef]
- Zhang, M.; Juraschek, S.P.; Appel, L.J.; Pasricha, P.J.; Miller, E.R., 3rd; Mueller, N.T. Effects of High-Fiber Diets and Macronutrient Substitution on Bloating: Findings From the OmniHeart Trial. Clin. Transl. Gastroenterol. 2020, 11, e00122. [Google Scholar] [CrossRef]
- Tarini, J.; Wolever, T.M. The fermentable fibre inulin increases postprandial serum short-chain fatty acids and reduces free-fatty acids and ghrelin in healthy subjects. Appl. Physiol. Nutr. Metab. 2010, 35, 9–16. [Google Scholar] [CrossRef]
- Dewulf, E.M.; Cani, P.D.; Claus, S.P.; Fuentes, S.; Puylaert, P.G.; Neyrinck, A.M.; Bindels, L.B.; de Vos, W.M.; Gibson, G.B.; Thissen, J.P.; et al. Insight into the prebiotic concept: Lessons from an exploratory, double blind intervention study with inulin-type fructans in obese women. Gut 2013, 62, 1112–1121. [Google Scholar] [CrossRef]
- Nilsson, A.C.; Johansson-Boll, E.V.; Bjorck, I.M. Increased gut hormones and insulin sensitivity index following a 3-d intervention with a barley kernel-based product: A randomised cross-over study in healthy middle-aged subjects. Br. J. Nutr. 2015, 114, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Visekruna, A.; Luu, M. The Role of Short-Chain Fatty Acids and Bile Acids in Intestinal and Liver Function, Inflammation, and Carcinogenesis. Front. Cell Dev. Biol. 2021, 9, 703218. [Google Scholar] [CrossRef]
- Kasubuchi, M.; Hasegawa, S.; Hiramatsu, T.; Ichimura, A.; Kimura, I. Dietary gut microbial metabolites, short-chain fatty acids, and host metabolic regulation. Nutrients 2015, 7, 2839–2849. [Google Scholar] [CrossRef]
- LeBlanc, J.G.; Chain, F.; Martín, R.; Bermúdez-Humarán, L.G.; Courau, S.; Langella, P. Beneficial effects on host energy metabolism of short-chain fatty acids and vitamins produced by commensal and probiotic bacteria. Microb. Cell Factories 2017, 16, 79. [Google Scholar] [CrossRef] [PubMed]
- Luu, M.; Monning, H.; Visekruna, A. Exploring the Molecular Mechanisms Underlying the Protective Effects of Microbial SCFAs on Intestinal Tolerance and Food Allergy. Front. Immunol. 2020, 11, 1225. [Google Scholar] [CrossRef]
- Ferrer-Picón, E.; Dotti, I.; Corraliza, A.M.; Mayorgas, A.; Esteller, M.; Perales, J.C.; Ricart, E.; Masamunt, M.C.; Carrasco, A.; Tristán, E.; et al. Intestinal inflammation modulates the epithelial response to butyrate in patients with inflammatory bowel disease. Inflamm. Bowel Dis. 2020, 26, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Gasaly, N.; Hermoso, M.A.; Gotteland, M. Butyrate and the fine-tuning of colonic homeostasis: Implication for inflammatory bowel diseases. Int. J. Mol. Sci. 2021, 22, 3061. [Google Scholar] [CrossRef]
- van der Beek, C.M.; Canfora, E.E.; Lenaerts, K.; Troost, F.J.; Olde Damink, S.W.; Holst, J.J.; Masclee, A.A.; Dejong, C.H.; Blaak, E.E. Distal, not proximal, colonic acetate infusions promote fat oxidation and improve metabolic markers in overweight/obese men. Clin. Sci. 2016, 130, 2073–2082. [Google Scholar] [CrossRef]
- Martin-Gallausiaux, C.; Marinelli, L.; Blottière, H.M.; Larraufie, P.; Lapaque, N. SCFA: Mechanisms and functional importance in the gut. Proc. Nutr. Soc. 2021, 80, 37–49. [Google Scholar] [CrossRef]
- Boets, E.; Deroover, L.; Houben, E.; Vermeulen, K.; Gomand, S.V.; Delcour, J.A.; Verbeke, K. Quantification of in Vivo Colonic Short Chain Fatty Acid Production from Inulin. Nutrients 2015, 7, 8916–8929. [Google Scholar] [CrossRef]
- Sivaprakasam, S.; Bhutia, Y.D.; Yang, S.; Ganapathy, V. Short-Chain Fatty Acid Transporters: Role in Colonic Homeostasis. Compr. Physiol. 2017, 8, 299–314. [Google Scholar] [CrossRef]
- Straß, S.; Schwamborn, A.; Keppler, M.; Cloos, N.; Guezguez, J.; Guse, J.-H.; Burnet, M.; Laufer, S. Synthesis, Characterization, and in vivo Distribution of Intracellular Delivered Macrolide Short-Chain Fatty Acid Derivatives. ChemMedChem 2021, 16, 2254–2269. [Google Scholar] [CrossRef]
- Nedjadi, T.; Moran, A.W.; Al-Rammahi, M.A.; Shirazi-Beechey, S.P. Characterization of butyrate transport across the luminal membranes of equine large intestine. Exp. Physiol. 2014, 99, 1335–1347. [Google Scholar] [CrossRef] [PubMed]
- Teramae, H.; Yoshikawa, T.; Inoue, R.; Ushida, K.; Takebe, K.; Nio-Kobayashi, J.; Iwanaga, T. The cellular expression of SMCT2 and its comparison with other transporters for monocarboxylates in the mouse digestive tract. Biomed. Res. 2010, 31, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Le Poul, E.; Loison, C.; Struyf, S.; Springael, J.-Y.; Lannoy, V.; Decobecq, M.-E.; Brezillon, S.; Dupriez, V.; Vassart, G.; Van Damme, J.; et al. Functional characterization of human receptors for short chain fatty acids and their role in polymorphonuclear cell activation. J. Biol. Chem. 2003, 278, 25481–25489. [Google Scholar] [CrossRef] [PubMed]
- Binder, H.J. Role of colonic short-chain fatty acid transport in diarrhea. Annu. Rev. Physiol. 2010, 72, 297–313. [Google Scholar] [CrossRef]
- Ghosh, S.; Pramanik, S. Structural diversity, functional aspects and future therapeutic applications of human gut microbiome. Arch. Microbiol. 2021, 203, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.J.; Goldsworthy, S.M.; Barnes, A.A.; Eilert, M.M.; Tcheang, L.; Daniels, D.; Muir, A.I.; Wigglesworth, M.J.; Kinghorn, I.; Fraser, N.J.; et al. The orphan G protein-coupled receptors GPR41 and GPR43 are activated by propionate and other short chain carboxylic acids. J. Biol. Chem. 2003, 278, 11312–11319. [Google Scholar] [CrossRef] [PubMed]
- Thangaraju, M.; Cresci, G.A.; Liu, K.; Ananth, S.; Gnanaprakasam, J.P.; Browning, D.D.; Mellinger, J.D.; Smith, S.B.; Digby, G.J.; Lambert, N.A.; et al. GPR109A is a G-protein-coupled receptor for the bacterial fermentation product butyrate and functions as a tumor suppressor in colon. Cancer Res. 2009, 69, 2826–2832. [Google Scholar] [CrossRef]
- Kobayashi, M.; Mikami, D.; Kimura, H.; Kamiyama, K.; Morikawa, Y.; Yokoi, S.; Kasuno, K.; Takahashi, N.; Taniguchi, T.; Iwano, M. Short-chain fatty acids, GPR41 and GPR43 ligands, inhibit TNF-α-induced MCP-1 expression by modulating p38 and JNK signaling pathways in human renal cortical epithelial cells. Biochem. Biophys. Res. Commun. 2017, 486, 499–505. [Google Scholar] [CrossRef]
- Hu, J.; Lin, S.; Zheng, B.; Cheung, P.C.K. Short-chain fatty acids in control of energy metabolism. Crit. Rev. Food Sci. Nutr. 2018, 58, 1243–1249. [Google Scholar] [CrossRef]
- Park, J.; Kim, M.; Kang, S.G.; Jannasch, A.H.; Cooper, B.; Patterson, J.; Kim, C.H. Short-chain fatty acids induce both effector and regulatory T cells by suppression of histone deacetylases and regulation of the mTOR–S6K pathway. Mucosal Immunol. 2015, 8, 80–93. [Google Scholar] [CrossRef] [PubMed]
- Vijay, N.; Morris, M.E. Role of monocarboxylate transporters in drug delivery to the brain. Curr. Pharm. Des. 2014, 20, 1487–1498. [Google Scholar] [CrossRef]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.; Anastasovska, J.; Ghourab, S.; Hankir, M.; Zhang, S.; et al. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat. Commun. 2014, 5, 3611. [Google Scholar] [CrossRef]
- Maric, G.; Gazibara, T.; Zaletel, I.; Labudovic Borovic, M.; Tomanovic, N.; Ciric, M.; Puskas, N. The role of gut hormones in appetite regulation (review). Acta Physiol. Hung. 2014, 101, 395–407. [Google Scholar] [CrossRef]
- Chambers, E.S.; Viardot, A.; Psichas, A.; Morrison, D.J.; Murphy, K.G.; Zac-Varghese, S.E.; MacDougall, K.; Preston, T.; Tedford, C.; Finlayson, G.S.; et al. Effects of targeted delivery of propionate to the human colon on appetite regulation, body weight maintenance and adiposity in overweight adults. Gut 2015, 64, 1744–1754. [Google Scholar] [CrossRef]
- Hume, M.P.; Nicolucci, A.C.; Reimer, R.A. Prebiotic supplementation improves appetite control in children with overweight and obesity: A randomized controlled trial. Am. J. Clin. Nutr. 2017, 105, 790–799. [Google Scholar] [CrossRef]
- Polyviou, T.; MacDougall, K.; Chambers, E.S.; Viardot, A.; Psichas, A.; Jawaid, S.; Harris, H.C.; Edwards, C.A.; Simpson, L.; Murphy, K.G.; et al. Randomised clinical study: Inulin short-chain fatty acid esters for targeted delivery of short-chain fatty acids to the human colon. Aliment. Pharmacol. Ther. 2016, 44, 662–672. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Wu, W.; Liu, Z.; Cong, Y. Microbiota metabolite short chain fatty acids, GPCR, and inflammatory bowel diseases. J. Gastroenterol. 2017, 52, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chambers, E.; Morrison, D.; Frost, G. Control of appetite and energy intake by SCFA: What are the potential underlying mechanisms? Proc. Nutr. Soc. 2015, 74, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Boets, E.; Gomand, S.V.; Deroover, L.; Preston, T.; Vermeulen, K.; De Preter, V.; Hamer, H.M.; Mooter, G.V.D.; De Vuyst, L.; Courtin, C.M.; et al. Systemic availability and metabolism of colonic-derived short-chain fatty acids in healthy subjects: A stable isotope study. J. Physiol. 2016, 595, 541–555. [Google Scholar] [CrossRef]
- Shen, M.; Shi, H. Sex hormones and their receptors regulate liver energy homeostasis. Int. J. Endocrinol. 2015, 2015, 294278. [Google Scholar] [CrossRef]
- Den Besten, G.; Lange, K.; Havinga, R.; van Dijk, T.H.; Gerding, A.; van Eunen, K.; Muller, M.; Groen, A.K.; Hooiveld, G.J.; Bakker, B.M.; et al. Gut-derived short-chain fatty acids are vividly assimilated into host carbohydrates and lipids. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 305, G900–G910. [Google Scholar] [CrossRef] [PubMed]
- den Besten, G.; Bleeker, A.; Gerding, A.; van Eunen, K.; Havinga, R.; van Dijk, T.H.; Oosterveer, M.H.; Jonker, J.W.; Groen, A.K.; Reijngoud, D.J.; et al. Short-chain fatty acids protect against high-fat diet-induced obesity via a PPAR gamma-dependent switch fom lipogenesis to fat oxidation. Diabetes 2015, 64, 2398–2408. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef]
- Bakshi, I.; Suryana, E.; Small, L.; Quek, L.-E.; Brandon, A.E.; Turner, N.; Cooney, G.J. Fructose bisphosphatase 2 overexpression increases glucose uptake in skeletal muscle. J. Endocrinol. 2018, 237, 101–111. [Google Scholar] [CrossRef]
- Alvaro, A.; Solà, R.; Rosales, R.; Ribalta, J.; Anguera, A.; Masana, L.; Vallvé, J.C. Gene expression analysis of a human enterocyte cell line reveals downregulation of cholesterol biosynthesis in response to short-chain fatty acids. IUBMB Life 2008, 60, 757–764. [Google Scholar] [CrossRef]
- Jakobsdottir, G.; Xu, J.; Molin, G.; Ahrné, S.; Nyman, M. High-fat diet reduces the formation of butyrate, but increases succinate, inflammation, liver fat and cholesterol in rats, while dietary fibre counteracts these effects. PLoS ONE 2013, 8, e80476. [Google Scholar] [CrossRef]
- Sahuri-Arisoylu, M.; Brody, L.P.; Parkinson, J.R.; Parkes, H.; Navaratnam, N.; Miller, A.D.; Thomas, E.L.; Frost, G.; Bell, J.D. Reprogramming of hepatic fat accumulation and “browning” of adipose tissue by the short-chain fatty acid acetate. Int. J. Obes. 2016, 40, 955–963. [Google Scholar] [CrossRef]
- Zhang, S.; Zhao, J.; Xie, F.; He, H.; Johnston, L.J.; Dai, X.; Wu, C.; Ma, X. Dietary fiber-derived short-chain fatty acids: A potential therapeutic target to alleviate obesity-related nonalcoholic fatty liver disease. Obes. Rev. 2021, 22, e13316. [Google Scholar] [CrossRef]
- Śliżewska, K. Resistant Dextrins as Prebiotic; Kapuśniak, J., Ed.; IntechOpen: Rijeka, Croatia, 2012. [Google Scholar] [CrossRef]
- Jochym, K.K.; Nebesny, E. Enzyme-resistant dextrins from potato starch for potential application in the beverage industry. Carbohydr. Polym. 2017, 172, 152–158. [Google Scholar] [CrossRef] [PubMed]
- Barczynska, R.; Jochym, K.; Slizewska, K.; Kapusniak, J.; Libudzisz, Z. The effect of citric acid-modified enzyme-resistant dextrin on growth and metabolism of selected strains of probiotic and other intestinal bacteria. J. Funct. Foods 2010, 2, 126–133. [Google Scholar] [CrossRef]
- Fouache, C.; Duflot, P. U.S. Patent 2003/6630586, 7 October 2003.
- van den Heuvel, E.G.; Wils, S.D.; Pasman, W.J.; Bakker, M.; Saniez, M.H.; Kardinaal, A.F. Short-term digestive tolerance of different doses of NUTRIOSE®FB, a food dextrin, in adult men. Eur. J. Clin. Nutr. 2004, 58, 1046–1055. [Google Scholar] [CrossRef] [PubMed]
- Van den Heuvel, E.G.H.M.; Wils, D.; Pasman, W.J.; Saniez, M.-H.; Kardinaal, A.F.M. Dietary supplementation of different doses of NUTRIOSE®FB, a fermentable dextrin, alters the activity of faecal enzymes in healthy men. Eur. J. Nutr. 2005, 44, 445–451. [Google Scholar] [CrossRef] [PubMed]
- Guérin-Deremaux, L.; Ringard, F.; Desailly, F.; Wils, D. Effects of a soluble dietary fibre NUTRIOSE® on colonic fermentation and excretion rates in rats. Nutr. Res. Pr. 2010, 4, 470–476. [Google Scholar] [CrossRef][Green Version]
- Serpelloni, M. U.S. Patent 2003/0077368 A1, 2003.
- Serpelloni, M. U.S. Patent 2006/0112956 A1, 2006.
- Brendel, R.; Boursier, B.; Leroux, P. U.S. Patent 2002/0192344 A1, 2002.
- Martirosyan, D.; Chen, S. FOSHU-approved Fibersol®-2 product review. Bioact. Compd. Health Dis. 2021, 4, 79. [Google Scholar] [CrossRef]
- Ohkuma, K.; Wakabayashi, S. Fibersol-2: A soluble, non-digestible, starch-derived dietary fibre. In Advanced Dietary Fibre Technology; McCleary, B.V., Prosky, L., Eds.; Blackwell Science Ltd.: Oxford, UK, 2001; pp. 509–523. [Google Scholar]
- Flickinger, E.A.; Wolf, B.W.; Garleb, K.A.; Chow, J.; Leyer, G.J.; Johns, P.W.; Fahey, G.C. Glucose-based oligosaccharides exhibit different in vitro fermentation patterns and affect in vivo apparent nutrient digestibility and microbial populations in dogs. J. Nutr. 2000, 130, 1267–1273. [Google Scholar] [CrossRef] [PubMed]
- Unno, T.; Nagata, K.; Horiguchi, T. Effects of green tea supplemented with indigestible dextrin on postprandial levels of blood glucose and insulin in human subjects. J. Nutr. Food 2002, 5, 31–39. [Google Scholar]
- Yamamoto, T. Effect of indigestible dextrin on visceral fat accumulation. J. Jpn. Soc. Study. Obes. 2007, 13, 34–41. [Google Scholar]
- Takagak, K.; Ikeguchi, M.; Artura, Y.; Fujinaga, N.; Ishibashi, Y.; Sugawa-Katayama, Y. The effect of AOJIRU drink powder containing indigestible dextrin on defecation frequency and faecal characteristics. J. Nutr. Food 2001, 4, 29–35. [Google Scholar]
- Hopkins, M.J.; Cummings, J.H.; Macfarlane, G.T. Inter-species differences in maximum specific growth rates and cell yields of bifidobacteria cultured on oligosaccharides and other simple carbohydrate sources. J. Appl. Microbiol. 2002, 85, 381–386. [Google Scholar] [CrossRef]
- Matsuda, I.; Satouchi, M. Agent for Promoting the Proliferation of Bifidobacterium. U.S. Patent 5698437, 1997. [Google Scholar]
- Aliasgharzadeh, A.P.; Dehghan, B.; Gargari, P.; Asghari-Jafarabadi, M. Resistant dextrin, as a prebiotic, improves insulin resistance and inflammation in women with type 2 diabetes: A randomised controlled clinical trial. Br. J. Nutr. 2015, 113, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Gholizadeh Shamasbi, S.; Dehgan, P.; Mohammad-Alizadeh Charandabi, S.; Aliasgarzadeh, A.; Mirghafourvand, M. The effect of resistant dextrin as a prebiotic on metabolic parameters and androgen level in women with polycystic ovarian syndrome: A randomized, triple-blind, controlled, clinical trial. Eur. J. Nutr. 2018, 58, 629–640. [Google Scholar] [CrossRef]
- Shamasbi, S.; Dehghan, P.; Mohammad-Alizadeh, S.; Aliasgarzadeh, A.; Mirghafourvand, M. Effect of Prebiotic on Anthropometric Indices in Women with Polycystic Ovarian Syndrome: A Triple-Blind, Randomized, Controlled Clinical Trial. Iran. Red Crescent Med. J. 2018, 20. [Google Scholar] [CrossRef]
- Farhangi, M.A.; Dehghan, P.; Namazi, N. Prebiotic supplementation modulates advanced glycation end-products (AGEs), soluble receptor for AGEs (sRAGE), and cardiometabolic risk factors through improving metabolic endotoxemia: A randomized-controlled clinical trial. Eur. J. Nutr. 2019, 59, 3009–3021. [Google Scholar] [CrossRef]
- Farhangi, M.A.; Javid, A.Z.; Sarmadi, B.; Karimi, P.; Dehghan, P. A randomized controlled trial on the efficacy of resistant dextrin, as functional food, in women with type 2 diabetes: Targeting the hypothalamic–pituitary–adrenal axis and immune system. Clin. Nutr. 2018, 37, 1216–1223. [Google Scholar] [CrossRef]
- Guérin-Deremaux, L.; Pochat, M.; Reifer, C.; Wils, D.; Cho, S.; Miller, L.E. The soluble fiber NUTRIOSE induces a dose-dependent beneficial impact on satiety over time in humans. Nutr. Res. 2011, 31, 665–672. [Google Scholar] [CrossRef]
- Guérin-Deremaux, L.; Pochat, M.; Reifer, C.; Wils, D.; Cho, S.; Miller, L.E. Dose–response impact of a soluble fiber, NUTRIOSE®, on energy intake, body weight and body fat in humans. Global Epidemic Obes. 2013, 1, 2. [Google Scholar] [CrossRef]
- Mateo-Gallego, R.; Pérez-Calahorra, S.; Lamiquiz-Moneo, I.; Marco-Benedí, V.; Bea, A.M.; Fumanal, A.J.; Prieto-Martín, A.; Laclaustra, M.; Cenarro, A.; Civeira, F. Effect of an alcohol-free beer enriched with isomaltulose and a resistant dextrin on insulin resistance in diabetic patients with overweight or obesity. Clin. Nutr. 2020, 39, 475–483. [Google Scholar] [CrossRef]
- Mateo-Gallego, R.; Moreno-Indias, I.; Bea, A.M.; Sánchez-Alcoholado, L.; Fumanal, A.J.; Quesada-Molina, M.; Prieto-Martín, A.; Gutiérrez-Repiso, C.; Civeira, F.; Tinahones, F.J. An alcohol-free beer enriched with isomaltulose and a resistant dextrin modulates gut microbiome in subjects with type 2 diabetes mellitus and overweight or obesity: A pilot study. Food Funct. 2021, 12, 3635–3646. [Google Scholar] [CrossRef]
- Mukai, J.; Tsuge, Y.; Yamada, M.; Otori, K.; Atsuda, K. Effects of resistant dextrin for weight loss in overweight adults: A systematic review with a meta-analysis of randomized controlled trials. J. Pharm. Health Care Sci. 2017, 3, 15. [Google Scholar] [CrossRef]
- Eliana, F.; Pranoto, B.A. A randomized controlled clinical trial of carbohydrate mix-fortified nutrition in type 2 diabetes mellitus patients. Med. J. Indones. 2020, 29. [Google Scholar] [CrossRef]
- Xie, X.; He, Y.; Li, H.; Yu, D.; Na, L.; Sun, T.; Zhang, D.; Shi, X.; Xia, Y.; Jiang, T.; et al. Effects of prebiotics on immunologic indicators and intestinal microbiota structure in perioperative colorectal cancer patients. Nutrition 2019, 61, 132–142. [Google Scholar] [CrossRef]
- Li, S.; Guerin-Deremaux, L.; Pochat, M.; Wils, D.; Reifer, C.; Miller, L.E. NUTRIOSE dietary fiber supplementation improves insulin resistance and determinants of metabolic syndrome in overweight men: A double-blind, randomized, placebo-controlled study. Appl. Physiol. Nutr. Metab. 2010, 35, 773–782. [Google Scholar] [CrossRef] [PubMed]
- Nazare, J.-A.; Sauvinet, V.; Normand, S.; Guérin-Deremaux, L.; Gabert, L.; Désage, M.; Wils, D.; Laville, M. Impact of a resistant dextrin with a prolonged oxidation pattern on day-long ghrelin profile. J. Am. Coll. Nutr. 2011, 30, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Arumugam, V.; Haugabrooks, E.; Williamson, P.; Hendrich, S. Soluble dietary fiber (Fibersol-2) decreased hunger and increased satiety hormones in humans when ingested with a meal. Nutr. Res. 2015, 35, 393–400. [Google Scholar] [CrossRef] [PubMed]
- So, E.Y.; Ouchi, M.; Cuesta-Sancho, S.; Olson, S.L.; Reif, D.; Shimomura, K.; Ouchi, T. Tumor suppression by resistant maltodextrin, Fibersol-2. Cancer Biol. Ther. 2012, 16, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Hendrich, S.; Ye, Z.; Arumugam, V.; Haugabrooks, E.; Williamson-Hughes, P. Fibersol-2 increases subjective and biochemical measures of satiety when ingested with a meal. FASEB J. 2010, 24, 230–238. [Google Scholar] [CrossRef]
- López Román, J.; Martínez Gonzálvez, A.B.; Luque, A.; Pons Miñano, J.A.; Vargas Acosta, A.; Iglesias, J.R.; Hernández, M.; Villegas, J.A. The effect of a fibre enriched dietary milk product in chronic primary idiopatic constipation. Nutr. Hosp. 2008, 23, 12–19. [Google Scholar]
- Hashizume, C.; Kishimoto, Y.; Kanahori, S.; Yamamoto, T.; Okuma, K.; Yamamoto, K. Improvement effect of resistant maltodextrin in humans with metabolic syndrome by continuous administration. J. Nutr. Sci. Vitaminol. 2012, 58, 423–430. [Google Scholar] [CrossRef]
- Fernández-Raudales, D.; Yor-Aguilar, M.; Andino-Segura, J.; Hernández, A.; Egbert, R.; López-Cintrón, J. Effects of high plant protein and high soluble fiber beverages on satiety, appetite control and subsequent food intake in healthy men. Food Nutr. Sci. 2018, 9, 751–762. [Google Scholar] [CrossRef]
- Dreher, M.L.; Dreher, C.J.; Berry, J.W.; Fleming, S.E. Starch digestibility of foods: A nutritional perspective. CRC Crit. Rev. Food Sci. Nutr. 1984, 20, 47–71. [Google Scholar] [CrossRef]
- Englyst, H.N.; Kingman, S.M.; Cummings, J.H. Classification and measurement of nutritionally important starch fractions. Eur. J. Clin. Nutr. 1992, 46 (Suppl. 2), S33–S50. [Google Scholar]
- Sajilata, M.G.; Singhal, R.S.; Kulkarni, P.R. Resistant Starch—A Review. Compr. Rev. Food Sci. Food Saf. 2006, 5, 1–17. [Google Scholar] [CrossRef]
- Homayouni, A.; Amini, A.; Keshtiban, A.; Amir, M.; Mortazavian, A.; Esazadeh, K.; Pourmoradian, S. Resistant starch in food industry: A changing outlook for consumer and producer. Starch Starke 2014, 6, 102–114. [Google Scholar] [CrossRef]
- Ma, Z.; Boye, J.I. Research advances on structural characterization of resistant starch and its structure-physiological function relationship: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1059–1083. [Google Scholar] [CrossRef]
- Hoover, R.; Vasanthan, T. The effect of annealing on the physicochemical properties of wheat, oat, potato and lentil starches. J. Food Biochem. 2007, 17, 303–325. [Google Scholar] [CrossRef]
- Zheng, Y.; Wang, Q.; Li, B.; Lin, L.; Tundis, R.; Loizzo, M.R.; Zheng, B.; Xiao, J. Characterization and Prebiotic Effect of the Resistant Starch from Purple Sweet Potato. Molecules 2016, 21, 932. [Google Scholar] [CrossRef]
- Ariyantoro, A.; Katsuno, N.; Nishizu, T. Effect of annealing process on physicochemical, morphological and gelatinization properties of cereal starches. Rev. Agric. Sci. 2018, 6, 81–92. [Google Scholar] [CrossRef]
- Genkina, N.; Wasserman, L.; Yuryev, V. Annealing of starches from Potato tubers grown at different environmental temperatures. Effect of heating duration. Carbohydr. Polym. 2004, 56, 367–370. [Google Scholar] [CrossRef]
- Gomand, S.; Lamberts, L.; Gommes, C.; Visser, R.; Delcour, J.; Goderis, B. Molecular and morphological aspects of annealing-induced stabilization of starch crystallites. Biomacromolecules 2012, 13, 1361–1370. [Google Scholar] [CrossRef]
- Ma, Z.; Hu, X.; Boye, J.I. Research advances on the formation mechanism of resistant starch type III: A review. Crit. Rev. Food Sci. Nutr. 2020, 60, 276–297. [Google Scholar] [CrossRef]
- Leszczyñski, W. Resistant starch—Classification, structure, production. Pol. J. Food Nutr. Sci. 2004, 13, 37–50. [Google Scholar]
- Lu, L.; Baik, B.-K. Starch characteristics influencing resistant starch content of cooked buckwheat groats. Cereal Chem. J. 2015, 92, 65–72. [Google Scholar] [CrossRef]
- Shin, M.; Woo, K.; Seib, P. Hot-Water Solubilities and Water Sorptions of Resistant Starches at 25 °C. Cereal Chem. 2003, 80, 564–566. [Google Scholar] [CrossRef]
- Wang, R.; Li, M.; Strappe, P.; Zhou, Z. Preparation, structural characteristics and physiological property of resistant starch. Adv. Food Nutr. Res. 2021, 95, 1–40. [Google Scholar] [CrossRef]
- Hoover, R.; Zhou, Y. In vitro and in vivo hydrolysis of legume starches by α-amylase and resistant starch formation in legumes—A review. Carbohydr. Polym. 2003, 54, 401–417. [Google Scholar] [CrossRef]
- Li, G.; Zeng, J.; Gao, H.; Li, X. Characterization of phosphate monoester resistant starch. Int. J. Food Prop. 2011, 14, 978–987. [Google Scholar] [CrossRef]
- Cornejo-Ramírez, Y.I.; Martínez-Cruz, O.; Del Toro-Sánchez, C.L.; Wong-Corral, F.J.; Borboa-Flores, J.; Cinco-Moroyoqui, F.J. The structural characteristics of starches and their functional properties. CyTA J. Food 2018, 16, 1003–1017. [Google Scholar] [CrossRef]
- Ahmadi-Abhari, S.; Woortman, A.J.J.; Hamer, R.J.; Loos, K. Assessment of the influence of amylose-LPC complexation on the extent of wheat starch digestibility by size-exclusion chromatography. Food Chem. 2013, 141, 4318–4323. [Google Scholar] [CrossRef]
- Bui, A.T.; Williams, B.A.; Hoedt, E.C.; Morrison, M.; Mikkelsen, D.; Gidley, M.J. High amylose wheat starch structures display unique fermentability characteristics, microbial community shifts and enzyme degradation profiles. Food Funct. 2020, 11, 5635–5646. [Google Scholar] [CrossRef]
- Yang, X.; Darko, K.O.; Huang, Y.; He, C.; Yang, H.; He, S.; Li, J.; Li, J.; Hocher, B.; Yin, Y. Resistant Starch Regulates Gut Microbiota: Structure, Biochemistry and Cell Signalling. Cell. Physiol. Biochem. 2017, 42, 306–318. [Google Scholar] [CrossRef]
- Alfa, M.J.; Strang, D.; Tappia, P.S.; Olson, N.; DeGagne, P.; Bray, D.; Murray, B.-L.; Hiebert, B. A randomized placebo controlled clinical trial to determine the impact of digestion resistant starch MSPrebiotic (®) on glucose, insulin, and insulin resistance in elderly and mid-age adults. Front. Med. 2018, 4, 260. [Google Scholar] [CrossRef]
- Karimi, P.; Farhangi, M.A.; Sarmadi, B.; Gargari, B.P.; Javid, A.Z.; Pouraghaei, M.; Dehghan, P. The Therapeutic Potential of Resistant Starch in Modulation of Insulin Resistance, Endotoxemia, Oxidative Stress and Antioxidant Biomarkers in Women with Type 2 Diabetes: A Randomized Controlled Clinical Trial. Ann. Nutr. Metab. 2015, 68, 85–93. [Google Scholar] [CrossRef]
- Al-Tamimi, E.K.; Seib, P.A.; Snyder, B.S.; Haub, M.D. Consumption of cross-linked resistant starch (RS4XL) on glucose and insulin responses in humans. J. Nutr. Metab. 2010, 2010, 651063. [Google Scholar] [CrossRef]
- Stewart, M.L.; Zimmer, J.P. Postprandial glucose and insulin response to a high-fiber muffin top containing resistant starch type 4 in healthy adults: A double-blind, randomized, controlled trial. Nutrition 2018, 53, 59–63. [Google Scholar] [CrossRef]
- Maki, K.C.; Pelkman, C.L.; Finocchiaro, E.T.; Kelley, K.M.; Lawless, A.L.; Schild, A.L.; Rains, T.M. Resistant Starch from High-Amylose Maize Increases Insulin Sensitivity in Overweight and Obese Men. J. Nutr. 2012, 142, 717–723. [Google Scholar] [CrossRef]
- Peterson, C.M.; Beyl, R.A.; Marlatt, K.L.; Martin, C.K.; Aryana, K.J.; Marco, M.L.; Martin, R.J.; Keenan, M.J.; Ravussin, E. Effect of 12 wk of resistant starch supplementation on cardiometabolic risk factors in adults with prediabetes: A randomized controlled trial. Am. J. Clin. Nutr. 2018, 108, 492–501. [Google Scholar] [CrossRef]
- Nichenametla, S.N.; Weidauer, L.A.; Wey, H.E.; Beare, T.M.; Specker, B.L.; Dey, M. Resistant starch type 4-enriched diet lowered blood cholesterols and improved body composition in a double blind controlled cross-over intervention. Mol. Nutr. Food Res. 2014, 58, 1365–1369. [Google Scholar] [CrossRef]
- Vahdat, M.; Hosseini, S.A.; Khalatbari Mohseni, G.; Heshmati, J.; Rahimlou, M. Effects of resistant starch interventions on circulating inflammatory biomarkers: A systematic review and meta-analysis of randomized controlled trials. Nutr. J. 2020, 19, 33. [Google Scholar] [CrossRef]
- Dainty, S.A.; Klingel, S.L.; Pilkey, S.E.; McDonald, E.; McKeown, B.; Emes, M.J.; Duncan, A.M. Resistant starch bagels reduce fasting and postprandial insulin in adults at risk of type 2 diabetes. J. Nutr. 2016, 146, 2252–2259. [Google Scholar] [CrossRef]
- Dodevska, M.S.; Sobajic, S.S.; Djordjevic, P.B.; Dimitrijevic-Sreckovic, V.S.; Spasojevic-Kalimanovska, V.V.; Djordjevic, B.I. Effects of total fibre or resistant starch-rich diets within lifestyle intervention in obese prediabetic adults. Eur. J. Nutr. 2016, 55, 127–137. [Google Scholar] [CrossRef]
- Vieira, C.R.; Lomeu, F.L.R.D.O.; Moreira, M.E.D.C.; Martino, H.S.D.; Silva, R.R. Clinical application of a cocoa and unripe banana flour beverage for overweight women with abdominal obesity: Prospective, double-blinded and randomized clinical trial. J. Food Biochem. 2017, 41, e12372. [Google Scholar] [CrossRef]
- Schioldan, A.G.; Gregersen, S.; Hald, S.; Bjørnshave, A.; Bohl, M.; Hartmann, B.; Holst, J.J.; Stødkilde-Jørgensen, H.; Hermansen, K. Effects of a diet rich in arabinoxylan and resistant starch compared with a diet rich in refined carbohydrates on postprandial metabolism and features of the metabolic syndrome. Eur. J. Nutr. 2017, 57, 795–807. [Google Scholar] [CrossRef]
- Esgalhado, M.; Kemp, J.A.; Azevedo, R.; Paiva, B.R.; Stockler-Pinto, M.B.; Dolenga, C.J.; Borges, N.A.; Nakao, L.S.; Mafra, D. Could resistant starch supplementation improve inflammatory and oxidative stress biomarkers and uremic toxins levels in hemodialysis patients? A pilot randomized controlled trial. Food Funct. 2018, 9, 6508–6516. [Google Scholar] [CrossRef]
- Tayebi Khosroshahi, H.; Vaziri, N.D.; Abedi, B.; Asl, B.H.; Ghojazadeh, M.; Jing, W.; Vatankhah, A.M. Effect of high amylose resistant starch (HAM-RS2) supplementation on biomarkers of inflammation and oxidative stress in hemodialysis patients: A randomized clinical trial. Hemodial. Int. 2018, 22, 492–500. [Google Scholar] [CrossRef]
- Khosroshahi, H.T.; Abedi, B.; Ghojazadeh, M.; Samadi, A.; Jouyban, A. Effects of fermentable high fiber diet supplementation on gut derived and conventional nitrogenous product in patients on maintenance hemodialysis: A randomized controlled trial. Nutr. Metab. 2019, 16, 1–8. [Google Scholar] [CrossRef]
- Meng, Y.; Bai, H.; Yu, Q.; Yan, J.; Zhao, L.; Wang, S.; Li, Z.; Wang, Q.; Chen, L. High–Resistant Starch, Low-Protein Flour Intervention on Patients With Early Type 2 Diabetic Nephropathy: A Randomized Trial. J. Ren. Nutr. 2019, 29, 386–393. [Google Scholar] [CrossRef]
- Costa, E.S.; França, C.N.; Fonseca, F.A.H.; Kato, J.T.; Bianco, H.T.; Freitas, T.T.; Fonseca, H.A.R.; Figueiredo Neto, A.M.; Izar, M.C. Beneficial effects of green banana biomass consumption in patients with pre-diabetes and type 2 diabetes: A randomised controlled trial. Br. J. Nutr. 2019, 121, 1365–1375. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AOAC Method | Measurement Type | Enzymes Used | Other Properties |
|---|---|---|---|
| 985.29 | Total HMWDF | Bacterial α-amylase, protease, and amyloglucosidase | 1 g sample, phosphate buffer |
| 991.42 | IDF in food products | Bacterial α-amylase, protease, and amyloglucosidase | 1 g sample, phosphate buffer |
| 991.43 | IDF and SDFP | Bacterial α-amylase, protease, and amyloglucosidase | 1 g sample, Tris or MES buffer, pH = 8.2 |
| 993.19 | SDFP in food products | Bacterial α-amylase, protease, and amyloglucosidase | 1 g sample, phosphate buffer |
| 993.21 | HMWDF (samples with more than 10% fiber and less than 2% starch) | Carried out without enzymes | 0.1 g sample, without buffer, |
| 994.13 | HMWDF | Bacterial α-amylase and amyloglucosidase | 0.25–0.5 g sample, acetate buffer, also provides composition of sugars and Klason lignin |
| 2001.03 | HMWDF and SDFS | Bacterial α-amylase, protease, and amyloglucosidase | 1 g sample, phosphate buffer, only for samples free of resistant starch |
| 2009.01 | HMWDF and SDFS | Pancreatic α-amylase, protease, and amyloglucosidase | 1 g sample, meleate buffer, pH = 6.0, available for any kind of food |
| 2011.25 | IDF, SDFP, and SDFS | Pancreatic α-amylase, protease, and amyloglucosidase | 1 g sample, meleate buffer, pH = 6.0, available for any kind of food |
| Dietary Fibers | Supplements Available on the Market | Dosage of Fiber Per Serving |
|---|---|---|
| Wheat dextrin | Benefiber (Novartis Consumer Health, USA) | 3 g |
| Wheat dextrin | Optifiber (Kirkland Signature, USA) | 3 g |
| Psyllium | 3-in-1 FIBER (Metamucil, AUS) | 2.4 g |
| Resistant tapioca dextrin | Completely Clear Organic Prebiotic Fiber (RenewLife, USA) | 6 g |
| Chia seed, organic flaxseed | Flax & Chia (Whole Foods Market, USA) | 8 g |
| Dextrin | Fiber Powder (Equate, KW) | 3.5 g |
| Flax seed, chia seed, sunflower seed and others | Raw Organic Fiber (Garden of Life, USA) | 9 g |
| Soluble stabilized rice bran, rice germ, chicory root | Zeal for Life (Zurvita, USA) | 4 g |
| Polydextrose | Fiber Well Fit (Vitafusion, USA) | 4 g |
| Chicory, inulin | Daily Fiber Gummies (Konsyl Pharmaceuticals, USA) | 3 g |
| Flax seed, chia seed | Super Seed Beyond Fiber (Garden of Life, USA) | 6 g |
| Chicory root, tapioca starch | Prebiotic Fiber Gummies (Lifeable, USA) | 4 g |
| Psyllium | Psyllium Husk (Sunergetic, USA) | 1.5 g |
| Soluble corn fiber | Prebiotic Soluble Fiber (Just Better, USA) | 5 g |
| Reference | Dextrin | Patients | Dosage [g/Day] | Time of Study | Outcome |
|---|---|---|---|---|---|
| [125] | Nutriose® | 55 women with type-2 diabetes (age 30–65) | 10 g | 8 weeks | Significant decrease in fasting insulin, malondialdehyde (MDA), and endotoxin. |
| [126] | Nutriose® | 62 women with polycystic ovary syndrome (age 18–45) | 20 g | 3 months | Positive influence on metabolic parameters, androgen levels, hirsutism, and menstrual cycle regularity |
| [127] | Nutriose® | 62 females (age 18–45) | 20 g | 12 weeks | Confirmed the positive and significant effects in reducing anthropometric indices |
| [128] | Nutriose® | 65 females with type-2 diabetes (age 30–65) | 10 g | 8 weeks | Improved end products of advanced glycation and other risk factors of cardiometabolic diseases |
| [129] | Nutriose® | 55 females with type-2 diabetes (age 30–65) | 10 g | 8 weeks | Supplementation yielded significant decrease in levels of cortisol, LPS. Increased levels of CD8 lymphocytes. Improved mental health and immune response. |
| [130] | Nutriose® | 50 males, 50 females (age 35–55) | 8 g, 14 g, 18 g, 24 g | 3 weeks | Decreased hunger over longer supplementation. Significant increase of short-term satiety |
| [131] | Nutriose® | 50 males, 50 females (age 35–55) | 8 g, 14 g, 18 g, 24 g | 9 weeks | Significant reduction of energy intake, BMI, and body fat in groups with intake of 14–24 g Nutriose per day |
| [132] | Resistant maltodextrin and isomaltose | 27 males, 14 females (age 18–80) | 5.28 g and 16.5 g | 20 weeks | Improvement of insulin resistance in patients with type-2 diabetes, overweight, and obesity |
| [36] | Resistant dextrin (MPCIR) | 38 males, 61 females (age 45–70) | 8–34 g | 12 weeks | Improvement of glycemic control, insulin resistance, and blood pressure. |
| [133] | Resistant maltodextrin and isomaltose | 11 males, 3 females (age 18–80) | 5.28 g and 16.5 g | 20 weeks | Enhancement of pathways related to metabolism, including terpenoid-quinone, lipopolysaccharides, and N-glycan biosynthesis. Significant impact on gut microbiota in diabetic subjects. |
| [134] | Resistant dextrin | 275 subjects (meta-analysis) | 10–34 g | 8–12 weeks | Beneficial effects on BMI and weight loss in overweight adults. |
| [49] | Nutriose® | 22 males, 64 females (age 18–59) | 10–20 g | 2 weeks | Increased counts of Bacteroides spp. and inhibition of Clostridium perfringens. Increased β-glucosidase activity and decreased colonic pH. No indications of gastrointestinal intolerance were found. |
| [48] | Nutriose® | 17 males, 19 females (age 22–55) | 14 g | 4 weeks | Supplementation was associated with higher fasted satiety scores and attenuation of the glycemic response |
| [135] | Isomaltulose, resistant dextrin, and inulin | 8 males, 22 females (age 18–60) | 45 g in total | 4 days | Reduction of glycemic response and longer term of satiety without causing any serious side effects |
| [136] | Fructooligosaccharides, xylooligosaccharides, polydextrose, and resistant dextrin | 90 males, 50 females (age 40–75) | 30 g total (7.5 g each component) | 1 week | Improved serum immunologic indicators |
| [137] | Nutriose® | 120 overweight males (age 26–35) | 34 g | 12 weeks | Reduction of energy intake, BMI, body fat percentage, and waist circumference. Improved glucose metabolism markers. Improved lipid metabolism. No adverse effects. |
| [138] | Nutriose® | 12 males (age 20–65) | 50 g | 10 h | Decreased ghrelin concentrations in response to the lunch, prolonged energy release. Reduced glycemic and insulinemic responses to breakfast. |
| [34] | Fibersol-2 | 23 males, 28 females (age 19–33) | 25 g | 3 weeks | Increased fecal Bifidobacteria counts and stool wet weight |
| [139] | Fibersol-2 | 10 males, 9 females (age 20–65) | 5 or 10 g | 4 h | A total of 10 g of Fibersol-2 stimulates production of satiety hormones (peptide-YY). |
| [140] | Fibersol-2 | HTC116 cell line | - | - | Significant inhibition of tumor growth of HCT116 cells by induction of apoptosis without visible signs of toxicity in vivo. |
| [141] | Fibersol-2 | 19 subjects | 5 or 10 g | 1 day | FS-2 administration stimulated production of specific satiety peptides such as PYY, decreased the hunger peptide ghrelin, and enhanced satiety after a meal. |
| [33] | Fibersol-2 | 32 males, 34 females (age 18–30) | 15 g | 3 weeks | Improved colonic functions, transit time, stool volume, and consistency. |
| [142] | Fibersol-2 | 4 males, 28 females (age 32–63) | 20 g | 20 days | Improved symptoms of the idiopathic primary chronic constipation. |
| [143] | Fibersol-2 | 20 males, 10 females (age 50–72) | 27 g | 12 weeks | Improved state of the risk factors of metabolic syndrome through the reduction of visceral fat and improvement of glucose and lipid metabolism. |
| [144] | Fibersol-2 | 24 males (age 20–24) | 11 g | 4 days | Increased satiety |
| Type of Resistant Starch | Description | Production | Source |
|---|---|---|---|
| RS1 | Protected physically, trapped in matrix resistant to enzymatic digestion | Milling or partial grinding | Grains, seeds, legumes, pastas |
| RS2 | Raw starch granules with crystalline structure of type B. | Gelatinization by thermal treatment | Green bananas, raw potatoes, corn with high amylose content, specific legumes |
| RS3 | Starch that underwent retrogradation process | Retrogradation by repeated thermal treatment (cooking and cooling) | Starch products that underwent specific thermal treatment (breads, cakes, cornflakes) |
| RS4 | Chemically or physically modified starches, cross-linked with chemical reagents | Mainly chemical, enzymatic, or thermal treatment often resulting in substitution with phosphates | Hardly available for human consumption. Present in specially designed starch products and food additives. |
| RS5 | Amylose complexes with lipids or fatty acids | Formed during food processing or naturally occurring within foods of high amylose content | Products with high amylose content |
| Reference | Resistant Starch | Patients | Dosage [g/Day] | Time and Type of Study | Outcome |
|---|---|---|---|---|---|
| [166] | MSPrebiotic® | 24 females, 18 males (age ≥ 70) and 25 females, 17 males (age 30–50) | 30 g/day | 3 months/RCT study | Significant reduction of insulin resistance, which is an important risk factor for developing type-2 diabetes. |
| [167] | RS2: Hi-maize 260, National Starch LLC | 56 females with type 2 diabetes (age 32–65) | 10 g/day | 8 weeks/RCT study | Significantly decreased levels of MDA, glycosylated hemoglobin, insulin, improved homeostasis model of insulin resistance and lowered endotoxins levels, a significant increase in TAC and glutathione peroxidase |
| [168] | Cross-linked RS type 4 | 7 females, 6 males (age 22–32, BMI 22–28) | 27 g/day | 1 day/ RCT study | Peak glucose and insulin concentrations in subjects were decreased |
| [169] | VERSAFIBE™ 2470 | 14 males, 14 females (age 24–58) | 11.6 g/day | 1 day/ RCT study | Significant reduction in postprandial serum glucose and decrease in maximum glucose concentration. Reduced postprandial serum insulin. |
| [170] | High-amylose maize type 2 resistant starch | 11 males, 22 females (age 18–69, BMI < 35) | 15, 30 g/day | 4 weeks/ RCT study | Improved insulin sensitivity in male subjects. |
| [171] | High-amylose maize (RS2) | 20 males, 39 females (BMI ≥ 27, age 35–75) | 45 g/day | 12 weeks/ RCT study | Reduced the inflammatory marker TNF-α and heart rate, but no significant improvement of glycemic control and other cardiovascular disease risk factors |
| [172] | RS4-enriched flour (30% v/v) | 86 adults (gender not specified) | 25.7 g/day | 12 weeks/RCT study | No significant effect for glycemic variables and blood pressures. Improved dyslipidemia (lowered cholesterol levels) and body composition. |
| [173] (Meta-analysis) | Resistant starch | 13 studies, 15–75 subjects per study | - | 4–14 weeks | Improved inflammatory biomarkers |
| [35] (Meta-analysis) | Resistant starch | 19 studies, 1014 subjects in total | - | - | Significant reduction in fasting plasma glucose, insulin, total cholesterol, and tumor necrosis factor alpha. |
| [174] | HAM-(RS2) | 16 males, 8 females (BMI = 30, average age 55) | 25 g | 57 days | Improved glycemic efficiency and fasting insulin sensitivity in adults at increased risk of T2D |
| [175] | Resistant starch | 19 males, 31 females (age > 50, overweight) | 25 g | 12 months | Glycemic control in prediabetic patients was unaffected by RS-rich diet in contrast to the regular fibre rich diet. |
| [176] | Resistant starch in form of cocoa and unripe banana flour beverage | 60 females (age 20–50) | 30 g | 6 weeks | Decreased the symptoms of dyspepsia, improved gastrointestinal symptoms, and increased production of propionic acid. The cocoa beverage showed an anti-inflammatory effect. |
| [177] | Arabinoxylan and resistant starch | 14 males, 5 females (age 39–75) | 21 g | 4 weeks | Improved fasting LDL and total cholesterol. No diet related impact on postprandial lipaemia. |
| [36] | Milk powder co-supplemented with inulin and resistant dextrin | 38 males, 61 females (age 45–70) | 45 g | 12 weeks | Supplementation improved glycemic control, insulin resistance, and blood pressure. |
| [178] | Resistant starch Hi-Maize® | 18 males, 13 females (age 42–65) | 16 g | 4 weeks | Supplementation improved inflammation and oxidative stress and reduced indoxyl sulfate plasma levels |
| [179] | HAM-resistant starch type 2 | 28 males, 16 females (age 41–74) | 25 g | 8 weeks | Significant reduction of levels of inflammatory and oxidative markers in hemodialysis patients |
| [180] | HAM-resistant starch type 2 | 29 males, 21 females (age 43–71) | 25 g | 8 weeks | Decreased serum levels of serum creatinine and p-cresol |
| [181] | Resistant starch (potato starch and high-amylase starch) | 39 males, 31 females (age 18–80) | 50 g | 12 weeks | Improvement of the blood glucose and blood lipid levels, decrease in the serum uric acid (UA) and urine β2-MG, and reduced antioxidative stress |
| [182] | Green banana biomass | 26 males, 87 females (age 18–85) | 40 g (approx. 5 g of resistant starch) | 24 weeks | Consumption of bioactive starches can improve metabolic control and body composition |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Włodarczyk, M.; Śliżewska, K. Efficiency of Resistant Starch and Dextrins as Prebiotics: A Review of the Existing Evidence and Clinical Trials. Nutrients 2021, 13, 3808. https://doi.org/10.3390/nu13113808
Włodarczyk M, Śliżewska K. Efficiency of Resistant Starch and Dextrins as Prebiotics: A Review of the Existing Evidence and Clinical Trials. Nutrients. 2021; 13(11):3808. https://doi.org/10.3390/nu13113808
Chicago/Turabian StyleWłodarczyk, Michał, and Katarzyna Śliżewska. 2021. "Efficiency of Resistant Starch and Dextrins as Prebiotics: A Review of the Existing Evidence and Clinical Trials" Nutrients 13, no. 11: 3808. https://doi.org/10.3390/nu13113808
APA StyleWłodarczyk, M., & Śliżewska, K. (2021). Efficiency of Resistant Starch and Dextrins as Prebiotics: A Review of the Existing Evidence and Clinical Trials. Nutrients, 13(11), 3808. https://doi.org/10.3390/nu13113808