Abstract
Marine-derived substances are known for their beneficial influences on aquatic animals’ performances and are recommended to improve intestinal health, immunity, and anti-oxidative status. The present study investigates the role of chitosan nanoparticles on the intestinal histo-morphometrical features in association with the health and immune response of Grey Mullet (Liza ramada). Chitosan nanoparticles are included in the diets at 0, 0.5, 1, and 2 g/kg and introduced to fish in a successive feeding trial for eight weeks. The final body weight (FBW), weight gain (WG), and specific growth rate (SGR) parameters are significantly increased while feed conversion ratio (FCR) decreases by chitosan nanoparticles compared to the control (p < 0.05). The morphometric analysis of the intestines reveals a significant improvement in villus height, villus width, and the number of goblet cells in chitosan-treated groups in a dose-dependent manner. Additionally, there is a positive correlation between the thickness of the enterocyte brush border and the chitosan dose, referring to an increasing absorptive activity. Histologically, the intestinal wall of Grey Mullet consists of four layers; mucosa, sub-mucosa, tunica muscularis (muscular layers), and serosa. The histological examination of the L. ramada intestine shows a normal histo-morphology. The epithelial layer of intestinal mucosa is thrown into elongated finger-like projections, the intestinal villi. The values of hemoglobin, hematocrit, red blood cells (RBCs), total protein (TP), albumin, and globulin are significantly increased in fish fed 1, and 2 g/kg of chitosan nanoparticles compared to fish fed 0 and 0.5 g/kg (p < 0.05). The highest levels of TP and albumin are observed in fish fed 1 g/kg diet (p < 0.05). The lysozyme activity and phagocytic index are significantly enhanced by feeding chitosan nanoparticles at 0.5, 1, and 2 g/kg, whereas the phagocytic activity is improved in fish fed 1 and 2 g/kg (p < 0.05). The highest lysozyme activity and phagocytic index are observed in fish fed 1 g/kg. SOD is significantly activated by feeding chitosan nanoparticles at 1 g/kg. Simultaneously, glutathione peroxidase (GPx) and catalase (CAT) activities also are enhanced by feeding chitosan at 1 and 2 g/kg, compared to fish fed 0 and 0.5 g/kg (p < 0.05). The highest GPx and CAT activities are observed in fish fed 1 g/kg (p < 0.05). Conversely, the malondialdehyde (MDA) levels are decreased by feeding chitosan at 1 and 2 g/kg, with the lowest being in fish fed 1 g/kg (p < 0.05). To summarize, the results elucidate that L. ramada fed dietary chitosan nanoparticles have a marked growth rate, immune response, and anti-oxidative response. These improvements are attributed to the potential role of chitosan nanoparticles in enhancing intestinal histo-morphometry and intestinal health. These results soundly support the possibility of using chitosan nanoparticles at 1–2 g/kg as a feasible functional supplement for aquatic animals.
1. Introduction
The Grey Mullet fish species are classified under the family of (Mugilidae) and can grow in tropical waters with a capacity to live in marine, brackish, and freshwater conditions []. These fish species are mainly spread by the countries’ northern coastal areas located on the southern Mediterranean Sea (e.g., Egypt) []. Grey Mullet (Liza ramada) can be farmed under the Egyptian conditions mostly in polyculture systems due to their high ability to benefit from the natural food cycles in the pond and the ability to utilize the formulated pellets, regardless of their protein content, as well as their high growth rate and their resistance against aquaculture stressors [,]. Moreover, fish are expected to suffer from malnutrition and stressful conditions, which induce inflammation in their intestinal health and deteriorate their intestinal immunity []. The impairment of their local intestinal immunity is correlated with their general immune response and their ability to resist fish infectious diseases []. Traditionally, antibiotics are used to relieve the inflammatory features caused by stress and malnutrition []. However, modern food security protocols suggest the necessity for reducing the application of chemotherapies and replace them with friendly alternative substances []. There are many feed additives used during the season of farming to improve the performance and wellbeing of aquatic animals []. Therefore, marine-derived substances, including chitosan, are proven to exhibit potential effects on the performance of cultured fish []. Chitosan is derived from chitin and extracted mainly from crustaceans (e.g., shrimp, crab, and crawfish) []. Chitosan has favorable effects, including bioremediation, immunostimulant, and antibacterial properties []. Further, dietary chitosan has no toxic influences on the biodegradable and biocompatibility effects []. Markedly, chitosans are used as antibacterial agents against pathogenic bacteria in aquaculture. Accordingly, chitosan increases the abundance of beneficial bacteria and decreases the pathogenic ones []. The microbial diversity in the fish’s gastrointestinal tract (GIT) strongly participates in regulating intestinal immunity depending on the ratio between the beneficial and pathogenic microorganisms and the composition of the aquafeed nutrients []. More specifically, the beneficial bacteria secrete digestive enzymes to digest the nutrients and facilitate their absorption through the intestines of fish, enhancing the feed efficiency and the growth rate []. Additionally, anti-oxidative properties are shown in aquatic animals fed diets incorporated with chitosan []. Interestingly, chitosan is known for its high resistance against hydrolysis caused by digestive enzymes in the GIT, and the ability to reduce intestinal injury and enhance the expression of tight junction proteins []. However, there seems to be no detailed information about using chitosan in aquatic animals to enhance intestinal health and wellbeing.
Nowadays, nanotechnology is widely applied in several sectors due to its functionality and positive effects. It helps minimize particle size and increase surface functionality, showing the maximum possible impact with the lowest possible amounts of a material []. Notably, the nanomaterials are applied in the biomedical, food preparation, and aquaculture sectors []. Regarding aquaculture-related studies, chitosan nanoparticles are included in the diets of Nile tilapia (Oreochromis niloticus) [,,,], rainbow trout (Oncorhynchus mykiss) [], silver carp (Hypophthalmichthys molitrix) [], and loach (Misgurnus anguillicaudatus) []. The results show an improved growth rate, feed utilization, immune response, and resistance against pathogens.
This study aims to investigate the potential role of dietary chitosan nanoparticles on the intestinal histo-morphometry, hemato-biochemical profile, immune response, and anti-oxidative status of Grey Mullet (L. ramada). Considering the growth-promoting, anti-inflammatory, anti-oxidative, and immune-enhancing effects of chitosan, it is proposed that chitosan would be a feasible and beneficial feed additive to enhance the GIT histo-morphometrical features of L. ramada.
2. Results
2.1. Growth Performance
Chitosan nanoparticles significantly increased the final body weight (FBW), weight gain (WG), and specific growth rate (SGR) parameters compared to the control (p < 0.05). The value of feed conversion ratio (FCR) was reduced in the group of L. ramada receiving 0.5, 1, and 2 g/kg of chitosan nanoparticles, with the lowest values being in fish fed 2 g/kg (p < 0.05) (Table 1).

Table 1.
Growth performance and nutrient utilization of L. ramada fed varying levels of chitosan nanoparticles *.
2.2. Intestinal Histomorphology
The results of the morphometric analysis of the intestines revealed a significant improvement in villus height, villus width, and the number of goblet cells in chitosan-treated groups in a dose-dependent manner (Table 2). Additionally, there was a positive correlation between the thickness of the enterocyte brush border and the dose of chitosan mediating the increase in absorptive activity.
Table 2.
Intestinal morphometry indices of L. ramada fed varying levels of chitosan nanoparticles *.
Histologically, the intestinal walls of Grey Mullet consist of four layers; mucosa, sub-mucosa, tunica muscularis (muscular layers), and serosa. The histological examination of the L. ramada intestine showed a normal histo-morphology. The epithelial layer of the intestinal mucosa was thrown into elongated finger-like projections, the intestinal villi. The epithelial lining of the intestinal villi was made of the absorptive columnar epithelium (enterocyte) and mucous-secreting goblet cells, as demonstrated using periodic acid-Schiff (PAS) staining. The brush borders of enterocytes were PAS-positive and appeared in the treated groups. The intestinal villi and associated crypt appeared free of any inflammatory or degenerative changes. Furthermore, the enterocytes and goblet cells were properly arranged (Figure 1, Figure 2 and Figure 3).
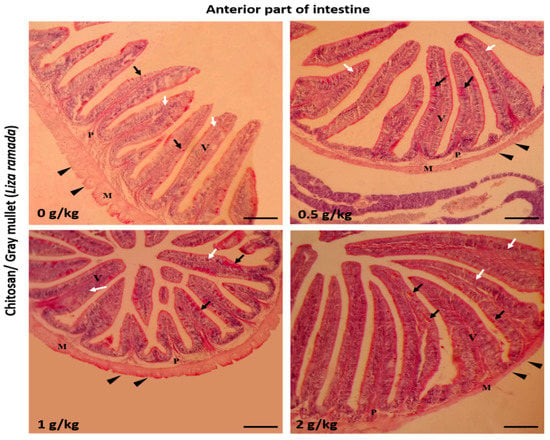
Figure 1.
Histomicrograph showing the histological structures of the anterior part of Grey Mullet intestines in the control group and chitosan-fed groups (0.5, 1, and 2 g/kg). The intestine shows normal histological structures of the intestinal wall and intestinal villi in all groups. The intestinal wall was formed of tunica mucosa of normally arranged enterocyte (white arrow) and goblet cells (black arrow), propria submucosa (P), tunica muscularis (M), and tunica serosa (black arrowhead). The villous height, width, and the number of goblet cells increased gradually in a dose-dependent manner with chitosan. Stain periodic acid-Schiff (PAS). Bar = 200 µm.
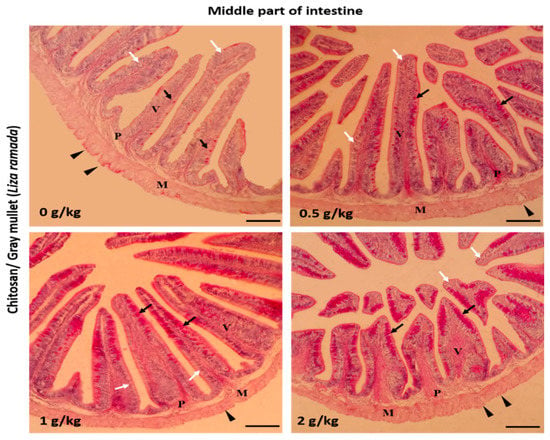
Figure 2.
Histomicrograph showing the histological structure of the middle part of Grey Mullet intestines in the control group as well as chitosan-fed groups (0.5, 1, and 2 g/kg). The intestine shows normal histological structures of the intestinal wall and intestinal villi in all groups. The intestinal wall was formed of tunica mucosa of normally arranged enterocyte (white arrow) and goblet cells (black arrow), propria submucosa (P), tunica muscularis (M), and tunica serosa (black arrowhead). The goblet cells were more prominent and gave a positive periodic acid-Schiff (PAS) reaction. The villous height, width, and the number of goblet cells increased gradually in a dose-dependent manner with chitosan. Stain PAS. Bar = 200 µm.
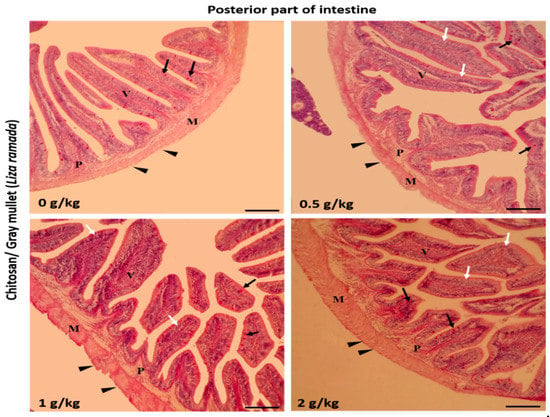
Figure 3.
Histomicrograph showing the histological structure of the posterior part of Grey Mullet intestines in the control group and chitosan-fed groups (0.5, 1, and 2 g/kg). The intestine shows normal histological structures of the intestinal wall and intestinal villi in all groups. The intestinal wall was formed of tunica mucosa of normally arranged enterocyte (white arrow) and goblet cells (black arrow), propria submucosa (P), tunica muscularis (M), and tunica serosa (black arrowhead). The villous height and width increased gradually in a dose-dependent manner with chitosan. Stain periodic acid-Schiff (PAS). Bar = 200 µm.
2.3. Haemato-Biochemical Indices
The values of hemoglobin, hematocrit, red blood cells (RBCs), total protein (TP), albumin, and globulin were significantly increased in fish fed 1 and 2 g/kg of chitosan nanoparticles compared to fish fed 0 and 0.5 g/kg (p < 0.05) (Table 3 and Table 4). The highest levels of TP and albumin were observed in fish fed 1 g/kg diet (p < 0.05).
Table 3.
Hematological indices of L. ramada fed varying levels of chitosan nanoparticles *.
Table 4.
Blood biochemical indices of L. ramada fed varying levels of chitosan nanoparticles *.
2.4. Immune Response
The lysozyme activity (Figure 4A) and phagocytic index (Figure 4B) were significantly enhanced by feeding chitosan nanoparticles at 0.5, 1, and 2 g/kg, whereas the phagocytic activity (Figure 4C) was improved in fish fed 1 and 2 g/kg (p < 0.05). The highest lysozyme activity and phagocytic index were observed in fish fed 1 g/kg.
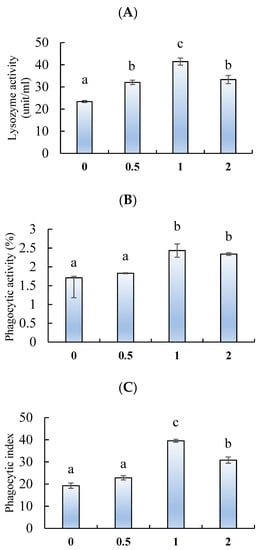
Figure 4.
(A) Lysozyme activity, (B) phagocytic activity, and (C) phagocytic index of L. ramada fed varying levels of chitosan nanoparticles. Bars with different letters indicate significant differences among the groups (as mean ± SE, n = 3).
2.5. Anti-oxidative Response
Viewing Figure 5A, the superoxide dismutase (SOD) was significantly activated by feeding chitosan nanoparticles at 1 g/kg. The glutathione peroxidase (GPx) (Figure 5B) and catalase (CAT) (Figure 5C) activities also were enhanced by feeding chitosan at 1 and 2 g/kg, when compared to fish fed 0 and 0.5 g/kg (p < 0.05). The highest GPx and CAT activities were observed in fish fed 1 g/kg (p < 0.05). Conversely, the levels of malondialdehyde (MDA) were decreased by feeding chitosan at 1 and 2 g/kg, with the lowest being in fish fed 1 g/kg (Figure 5 D) (p < 0.05).
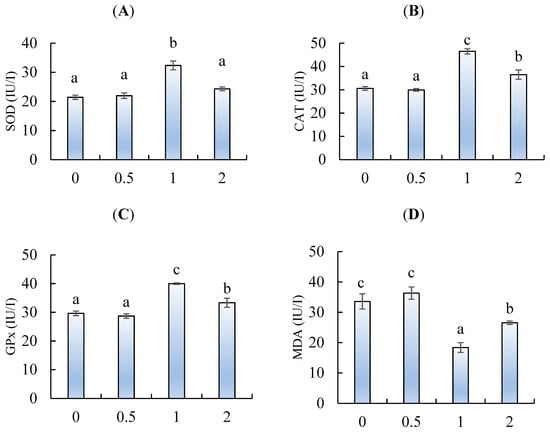
Figure 5.
(A) Superoxide dismutase (SOD), (B) catalase (CAT), (C) glutathione peroxidase, and (D) malondialdehyde (MDA) of L. ramada fed varying levels of chitosan nanoparticles. Bars with different letters indicate significant differences among the groups (as mean ± SE, n = 3).
3. Discussion
Marine-derived chitosan has diverse biological influences on the performance of aquatic animals [,]. Chitosan can be extracted from crustacean exoskeletons and structured through the deacetylation of chitin with abundant amounts of copolymers of glucosamine N-acetylglucosamine, which are stable during digestion and can tolerate the hydrolysis function of the digestive enzymes []. Correspondingly, chitosan can stay stable for a long time until it can reach the targeted organ. Further, the nanoparticle form of chitosan facilitates its functionality and absorption at low supplementary levels []. As expected, the results clearly show an enhanced growth performance and feed utilization of L. ramada fed dietary chitosan nanoparticles, which can be attributed to the potential role of chitosan in improving the intestinal-histo-morphometric features. The inclusion of dietary chitosan at the rate of 1–2 g/kg resulted in the best growth rate and histomorphometrical features for the intestine of L. ramada. Concurrently, the absorption of nutrients by the intestinal villi was enhanced and the natural metabolic functions were regulated, which resulting in improved blood health, immune response, and anti-oxidative responses. These results illustrate that the inclusion of chitosan nanoparticles is recommended in a dose-dependent manner to improve aquatic animal performances and relieve the expected inflammation that can result from stressful aquaculture practices.
The results of the current study showed enhanced growth performance of L. ramada fed diets with chitosan nanoparticles. Similarly, the growth performance of Nile tilapia [,,,], rainbow trout [], silver carp [], and loach [] was improved by dietary chitosan. The growth-promoting activity of chitosan is attributed to its role in activating the digestive enzymes by inhibiting the pathogenic bacteria and the activation of beneficial ones []. Accordingly, the beneficial microorganisms can secrete digestive enzymes to digest the feed and convert it to available nutrients for absorption through the intestinal villi []. The antibacterial capacity of chitosan reduces the abundance of harmful microorganisms in the fish intestines and, in turn, enhances the local intestinal immunity []. The obtained results also showed improved feed utilization (reduced feed conversion ratio) in L. ramada fed chitosan nanoparticles. The enhanced feed efficacy also explains the improved growth performance in the present study. Although the digestive enzyme activity was not detected in the present study, the inclusion of chitosan nanoparticles was proved to activate the intestinal digestive enzymes in other fish species (e.g., Nile tilapia [,]). The enhancement of digestive enzyme activity also could explain the potential role of chitosan nanoparticles in improving the feed utilization and the growth performance of L. ramada Zhang [] attributed the enhanced growth rate and feed utilization to chitosan’s role in activating digestive enzymes, which are responsible for the hydrolysis of nutrients (e.g., carbohydrates, lipids, and proteins) in the gastrointestinal tract (GIT). Correspondingly, the digested nutrients can be easily absorbed by the intestinal villi and be available for the cells.
The measurement of intestinal histomorphometrical features is a vital tool for evaluating the effect of dietary supplements on the intestinal absorption capacity and the neighboring intestinal immunity, as well as its relationship to the general immune system in the fish body [,]. The main findings of the current investigation are the enhancement of the intestinal morphometrical and histological features in L. ramada fed chitosan nanoparticles. The intestinal villi and width were improved, while the number of goblet cells were increased in fish treated with chitosan nanoparticles. Concurring, Abd El-Naby et al. [] reported that Nile tilapia fed chitosan nanoparticles had improved intestinal histological features. The improved intestinal morphometry refers to the increased surface of intestinal villi to absorb the digested nutrients in the intestine []. The enhanced intestinal health and morphometry indices explain the enhanced feed utilization in L. ramada fed dietary chitosan nanoparticles. Dietary chitosan nanoparticles enhanced the diversity and integrity of the intestinal epithelial cells which, correspondingly, alleviated the intestinal pathogenicity and inflammation that can be attributed to the antibacterial effect of chitosan. The results are in line with [], who stated that chitosan improved the local intestinal integrity and immunity of Pacific white shrimp (Litopenaeus vannamei). Concurrently, the enhanced local immunity led to enhanced humoral and innate immune responses due to chitosan feeding.
The suitability of feed supplements also can be assessed using hematological parameters, such as red blood cells (RBCs), white blood cells (WBCs), hemoglobin concentrations, and hematocrit levels []. Changes in the values of these parameters may occur due to the fish’s health condition and nutritional behavior []. Notably, the results show that L. ramada fed chitosan nanoparticles increased WBCs, RBCs, hemoglobin, and hematocrit values. These results confirm the safe use of chitosan nanoparticles and their beneficial influence on the health condition of L. ramada. Concurrently, Abd El-Naby et al. [] and Younus et al. [] illuminated that Nile tilapia and silver carp fed chitosan nanoparticles had increased WBCs, RBCs, hemoglobin, and hematocrit values, and attributed this influence on the potential role of chitosan in enhancing the antibacterial, immunity and anti-oxidative conditions.
Lysozyme activity is a part of the immune system that can deactivate the pathogenicity of the harmful bacteria in the fish body []. Likewise, phagocytosis is a function where phagocytic cells can attack the pathogenic bacteria and resist its pathogenic role on fish []. Marine-derived polysaccharides are known for their immunomodulation influence and anti-inflammatory and antibacterial capacities []. Relatedly, this study’s results clearly show improved lysozyme and phagocytic activities in L. ramada fed dietary chitosan nanoparticles. These results indicate that L. ramada has a resistance against the common infection attributed to pathogens in fishponds. Paralleling our study, Younus et al. [] and Khani Oushani et al. [] illustrated that silver carp and rainbow trout fed dietary chitosan nanoparticles had enhanced lysozyme and antibacterial activity. Interestingly, the enhanced phagocytosis activity seen in the present study accompanies the increased number of WBCs, which explains the improved immune response of L. ramada fed dietary chitosan nanoparticles. Victor et al. [] suggested that chitosan can activate a fish’s immunity by regulating innate immunity through Toll-like receptor-regulated signaling pathways. However, further studies are recommended to clarify the mode of action that allows chitosan to enhance aquatic animal immune responses.
Oxidative stress is caused by an imbalance in the production of reactive oxygen species (ROS) and antioxidant enzymes []. Fish have an anti-oxidative defense mechanism for ROS []. This defense mechanism includes antioxidant enzymes (e.g., superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx)) that can inhibit ROS production and protect the body from damage such as deoxyribonucleic acid (DNA) strand breaks, lipid peroxidation, protein oxidation, plus the impairment of the cell metabolism function and structure []. Lipid peroxidation occurring in the cells can be measured by the concentration of malondialdehyde (MDA) []. Considering aquaculture, chitosan is frequently used to prepare a fish feed and has been confirmed to present anti-oxidative activity when administered orally to fish []. During the present study, L. ramada fed dietary chitosan nanoparticles had enhanced SOD, CAT, and GPx and reduced MDA levels. Similarly, Nile tilapia [] and loach [] fed dietary chitosan nanoparticles showed an enhanced anti-oxidative capacity. Chitosan nanoparticles enhanced the antioxidant ability probably due its chelating ability and the scavenging role of free radicals []. The results suggest that dietary chitosan resulted in enhanced SOD, CAT, and GPx neutralizing oxidative stress-induced damage, such as lipid peroxidation (MDA) [] that may occur during the farming season of L. ramada. It has been hypothesized that dietary chitosan probably activates the anti-oxidative response by improving feeding utilization, allowing dietary antioxidants to play a role in antioxidant activity []. More specifically, chitosan’s anti-oxidative capacity overcomes oxidative stress, probably induced by harmful microorganisms, in the GIT of fish.
The results show that fish fed chitosan nanoparticles at 2 g/kg diet had relatively lower immune and anti-oxidative responses than fish fed 1 g/kg. These results are probably due to the necessity of including chitosan nanoparticles in a dose-dependent manner. The inclusion of functional feed additives is known for the species-specific doses depending on the fish species, feeding duration, and rearing conditions []. The results also confirm the proposed hypothesis that the nano form of chitosan could be included in lower levels compared to organic and other forms []. Few studies in the literature explain the role of chitosan in improving the performances of aquatic animals. However, no investigations to our knowledge were done to apply the mechanistic role (e.g., quantitative real-time polymerase chain reaction (Q-PCR) and Western Blot) of chitosan in aquatic animals. Although the study limits the application of related genetic analysis that can explain chitosan functionality in improving the histomorphometrical features, the histological study conducted in the present study opens the door for further future studies that explain the mechanistic effect of chitosan in enhancing the performance of aquatic animals.
4. Materials and Methods
4.1. Ethical Approval
The protocol of the present experiment and fish rearing techniques were approved and conducted with the guidance of the Research Ethical Committee, Faculty of Agriculture, Kafrelsheikh University, Egypt.
4.2. Preparation of Chitosan Nanoparticles
Chitosan nanoparticles were prepared by following Tang, et al. [] with some modifications. Chitosan was dissolved in 80 mL distilled water containing 1% glacial acetic acid to turn the pH acidic, then magnetically stirred for 30 min to fully dissolve. Then, 20 mL of sodium tripolyphosphate (TPP) solution was dropped into the chitosan beaker at room temperature while stirring. Subsequently, the chitosan solution was magnetically stirred for 45 mins to obtain chitosan nanoparticles (Figure 6). These chitosan nanoparticles could be stably stored at −8 °C.
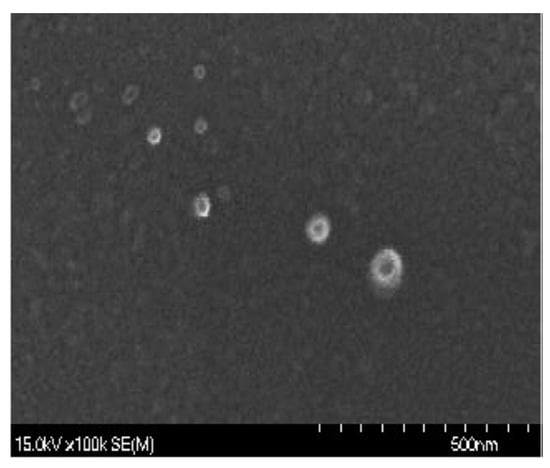
Figure 6.
The morphological feature of chitosan nanoparticles observed by a scanning electron microscope (SEM).
4.3. Fish, Design, and Diets
A stock of visually similar sizes of Grey Mullet (L. ramada) juveniles was collected from a local farm in Kafr El-Sheikh governorate and gently transported to the laboratory to avoid stressful transportation effects on the fish. Fish were then stocked in a concrete tank (5 × 10 × 1 m), which provided continuous aeration, and kept for ten days for adaptation. During the acclimatization period, fish were fed the control diet, and half of the water was replaced daily with freshly dechlorinated water. Then, fish with similar average initial weights (52.95 ± 0.81 g) were randomly selected and distributed in 12 concrete tanks (2 × 5 × 1 m) (4 groups/3 replicates), wherein each tank were placed 50 fish. The tanks were in an outdoor area under the conditions of a flow-through system. During the trial (8 weeks), the water characteristics were checked regularly and reported to be within the normal values for the optimal growth of common carp, and experienced a photoperiod (12 h light: 12 h darkness). The water temperature, pH, dissolved oxygen, and total ammonia averaged 25.34 ± 0.22 °C, 7.62 ± 0.54, 6.11 ± 0.31 mg/L, and 0.02 ± 0.001 mg/L, respectively.
The basal diet was formulated by well-mixing fish meal, soybean meal, yellow corn, wheat bran, rice bran, and a vitamin and mineral mixture (Table 5). Then, pellets were prepared with fish oil and water to prepare a dough of 1–2 mm in size using the laboratory pelletizer. Four types of diets were prepared, where the control diet was not supplemented with chitosan nanoparticles and the second, third, and fourth diets were supplemented with chitosan nanoparticles at 0.5, 1, and 2 g/kg, respectively. The prepared diets were then dried at room temperature and kept at 4 °C. Fish were fed the diets at 2–3% throughout the trial.
Table 5.
Basal diet and proximate chemical composition (%, on dry matter basis).
4.4. Final Sampling
Upon conclusion of the trial, all fish fasted 24 hours before the final sampling. Then, fish in each tank were weighed and counted to calculate the growth performance, feed efficiency, and survival indices. The following equations were used for the calculation: Weight gain (WG, %) = (FBW − IBW) × 100/IBW, specific growth rate (SGR, %g/day) = 100((LnFBW − LnIBW)/T), feed conversion ratio (FCR) = FI/WG, and survival (%) = (final NO./initial NO.) ×100, where FBW = body weight final (g), IBW = body weight initial (g), T = duration (days), FI = feed intake (g), and NO. = number of fish.
Afterward, fish were anesthetized (100 mg/L tricaine methanesulfonate), and three fish from each tank were randomly collected for blood sampling and dissection. Fish were bled from the caudal vein with a nonheparinized syringe for serum collection (3000 rpm for 15 min at 4 °C), while heparinized syringes were used for the whole blood collection. The whole blood was used for the hematological and phagocytic analysis, while the collected serum was kept at −80 °C until use. Then, fish were gently dissected and the intestines were collected and rinsed in Boin’s solution for histology sections.
4.5. Blood Analysis
Red blood cells (RBCs) and white blood cells (WBCs) were counted with a hemocytometer immediately after dilution with Natt and Herrick’s solution []. Regarding the differential leucocytic count, blood films were prepared and stained according to Lucky [], while cells were calculated by following Jain []. Blood hemoglobin concentration was determined with a spectrophotometer (Model RA 1000, Technicon Instruments Corporation Tarrytown, NY USA) at 540 nm using the Blaxhall and Daisley [] method.
According to Doumas, et al. [] and Dumas [], the serum total protein and albumin were determined. The globulin content was calculated mathematically. The activities of aspartate aminotransferase (AST) and alanine aminotransferase (ALT) were determined calorimetrically at a wavelength of 540 nm []. Serum creatinine and urea were calorimetrically determined according to Coulombe and Favreau [] and Heinegård and Tiderström [], respectively.
Blood lysozyme activity was determined using the turbidimetric assay []. The phagocytic activity and phagocytic index were determined according to Kawahara, et al. [].
Superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx) in serum were measured using diagnostic reagent kits following the manufacturer’s (Cusabio Biotech Co., Ltd.; China) instructions. The detailed instructions for SOD, CAT, and GPx measurements are written in Catalog Numbers: CSB-E15929Fh, CSB-E15928Fh, and CSB-E15930Fh, respectively. Briefly, the reagents and samples were prepared according to the instructions then the blank was set well and 50 μL of SOD, CAT, and GPx standards or samples were added to each well. Fifty microliters of Horseradish Peroxidase (HRP) conjugate (1X) was added to each well except the blank one, then incubated at 37 °C for one hour, aspirated and washed five times, then 90 μL of TMB (3,3’,5,5’-Tetramethylbenzidine) substrate was added to each well and incubated in a dark place at 37 °C for half an hour. Subsequently, 50 μL of stop solution was added to each well and read at 450 nm within 10 minutes.
Serum malonaldehyde (MDA) content was assessed, according to Ledwozyw et al. []. One mL of serum was mixed with 2 mL of trichloroacetic acid (TBA) and hydrochloric acid under acidic circumstances then the mixture was diluted to two hundred mL using water and kept for half an hour. It was then cooled and centrifuged at 300 rpm for 10 minutes, the supernatant was separated, and the absorption was measured at 535 nm.
4.6. Intestinal Morphometry
The histological investigation was carried out at the end of the experiment. Nine Grey Mullet fish were randomly selected for each treatment (3 fish from each aquarium). After deep anesthesia using 40% ethyl alcohol, the abdomen was dissected and specimens from the anterior, middle, and terminal parts of the intestine were sampled. The samples were immediately fixed for 48 h in 10% neutral buffered formalin solution. After fixation, tissue specimens were processed according to Gewaily et al. []. Briefly, the tissue samples were dehydrated in ascending grades of ethyl alcohol, cleared in xylene, embedded in paraffin wax, cut into several 5 μm thick sections using a rotary microtome (RM 20352035; Leica Microsystems, Wetzlar, Germany), and mounted on clean slides. The paraffin sections were rehydrated and stained with periodic acid-Schiff (PAS) according to the methodology described by Bancroft and Gamble [] for general morphometry and goblet cell identification. Afterward, several representative photomicrographs were captured from the stained sections with a digital camera (Leica EC3, Leica, GmbH, Wetzlar, Germany) connected to a microscope (Leica DM500).
The morphometric study was adopted by examining villus height (μm, from the tip to the base of villus), villus width (μm, at the mid-height of villi), and the number of PAS-positive goblet cells. The measurements were done using a computerized image analysis system (Image J software; Bethesda, MD, USA) []. The results were revealed as means ± standard error (SE). We used the statistical software package SPSS 22 (SPSS® Inc., Chicago, IL, USA) to perform the statistical analysis.
4.7. Statistical Analysis
Normality and homoscedasticity analyses were established using Shapiro-Wilk and Levene tests before applying a one-way ANOVA method to confirm the normal distribution of the data, then the obtained data were subjected to a one-way ANOVA using SPSS version 22 (SPSS® Inc., Chicago, IL, USA). Data were presented as means ± the standard error (SE). Differences between the means were tested at the 5% probability level using a Duncan test as a post-hoc test.
5. Conclusions
To summarize, the results elucidated that L. ramada fed dietary chitosan nanoparticles had a marked growth performance and feed utilization. These improvements were attributed to the potential role of chitosan nanoparticles in enhancing the intestinal histo-morphometry and antibacterial capacity. The immunity and anti-oxidative responses also were improved with the continuous feeding of chitosan nanoparticles. These results soundly supported the possibility of using chitosan nanoparticles as a feasible functional supplement for aquatic animals.
Author Contributions
Conceptualization, M.A.O.D., M.S.G., A.A.S., M.S., A.A.A., E.M.Y., A.-W.A.A.-W., H.V.D., A.H.S. and S.E.F.; data curation, M.A.O.D.; formal analysis, M.A.O.D., M.S.G., M.S. and M.A.; funding acquisition, M.A.O.D. and E.M.Y.; investigation, M.A.O.D., A.A.S., A.A.A., A.-W.A.A.-W., M.A. and S.E.F; methodology, M.S.G., A.A.S., M.S., A.A.A., H.V.D. and A.H.S.; project administration, M.A.O.D.; resources, A.H.S. and S.E.F.; software, M.A.; writing—original draft, M.A.O.D., E.M.Y., A.-W.A.A.-W., H.V.D., and H.M.R.A.-L.; writing—review & editing, M.A.O.D. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Researchers Supporting Project (RSP-2020/36), King Saud University, Riyadh, Saudi Arabia. This research work was partially supported by Chiang Mai University.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Whitfield, A.; Panfili, J.; Durand, J.-D. A global review of the cosmopolitan flathead mullet Mugil cephalus linnaeus 1758 (teleostei: Mugilidae), with emphasis on the biology, genetics, ecology and fisheries aspects of this apparent species complex. Rev. Fish Biol. Fish. 2012, 22, 641–681. [Google Scholar] [CrossRef]
- Bahnasawy, M.; Khidr, A.A.; Dheina, N. Seasonal variations of heavy metals concentrations in mullet, Mugil cephalus and Liza ramada (mugilidae) from lake manzala, egypt. Egypt. J. Aquat. Biol. Fish. 2009, 13, 81–100. [Google Scholar] [CrossRef]
- El-Dahhar, A.A. Effect of heat-treated feed and exogenous zymogen on survival and growth of grey mullet, liza ramada (risso), larvae in egypt. Aquac. Res. 1999, 30, 165–173. [Google Scholar] [CrossRef]
- Toutou, M.M.; Soliman, A.A.; Elokaby, M.A.; Ahmed, R.A.; Elmoneam, M.Y.A. Growth performance and biochemical blood parameters of nile tilapia, Oreochromis niloticus, and thinlip mullet, Liza ramada, fed a diet supplemented with lemon (Citrus aurantifolia) peel in a polyculture system. Egypt. J. Aquat. Biol. Fish. 2018, 22, 183–192. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S.; Esteban, M.Á. Beneficial roles of feed additives as immunostimulants in aquaculture: A review. Rev. Aquac. 2018, 10, 950–974. [Google Scholar] [CrossRef]
- Dawood, M.A.O. Nutritional immunity of fish intestines: Important insights for sustainable aquaculture. Rev. Aquac. 2020. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Abo-Al-Ela, H.G.; Hasan, M.T. Modulation of transcriptomic profile in aquatic animals: Probiotics, prebiotics and synbiotics scenarios. Fish Shellfish Immunol. 2020, 97, 268–282. [Google Scholar] [CrossRef]
- Basuini, M.F.E.; Teiba, I.I.; Zaki, M.A.A.; Alabssawy, A.N.; El-Hais, A.M.; Gabr, A.A.; Dawood, M.A.O.; Zaineldin, A.I.; Mzengereza, K.; Shadrack, R.S.; et al. Assessing the effectiveness of coq10 dietary supplementation on growth performance, digestive enzymes, blood health, immune response, and oxidative-related genes expression of nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2020, 98, 420–428. [Google Scholar] [CrossRef]
- Alishahi, A.; Aïder, M. Applications of chitosan in the seafood industry and aquaculture: A review. Food Bioprocess Technol. 2012, 5, 817–830. [Google Scholar] [CrossRef]
- Mohan, K.; Ravichandran, S.; Muralisankar, T.; Uthayakumar, V.; Chandirasekar, R.; Seedevi, P.; Abirami, R.G.; Rajan, D.K. Application of marine-derived polysaccharides as immunostimulants in aquaculture: A review of current knowledge and further perspectives. Fish Shellfish Immunol. 2019, 86, 1177–1193. [Google Scholar] [CrossRef]
- Nikapitiya, C.; Dananjaya, S.H.S.; De Silva, B.C.J.; Heo, G.-J.; Oh, C.; De Zoysa, M.; Lee, J. Chitosan nanoparticles: A positive immune response modulator as display in zebrafish larvae against Aeromonas hydrophila infection. Fish Shellfish Immunol. 2018, 76, 240–246. [Google Scholar] [CrossRef]
- Wei, X.; Liao, J.; Davoudi, Z.; Zheng, H.; Chen, J.; Li, D.; Xiong, X.; Yin, Y.; Yu, X.; Xiong, J. Folate receptor-targeted and gsh-responsive carboxymethyl chitosan nanoparticles containing covalently entrapped 6-mercaptopurine for enhanced intracellular drug delivery in leukemia. Mar. Drugs 2018, 16, 439. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.; Soliman, F.M.; Adly, M.A.; Soliman, H.A.M.; El-Matbouli, M.; Saleh, M. Recent progress in biomedical applications of chitosan and its nanocomposites in aquaculture: A review. Res. Vet. Sci. 2019, 126, 68–82. [Google Scholar] [CrossRef]
- Encarnação, P. 5—Functional feed additives in aquaculture feeds. In Aquafeed Formulation; Nates, S.F., Ed.; Academic Press: San Diego, CA, USA, 2016; pp. 217–237. [Google Scholar]
- Yan, J.; Guo, C.; Dawood, M.; Gao, J. Effects of dietary chitosan on growth, lipid metabolism, immune response and antioxidant-related gene expression in Misgurnus anguillicaudatus. Benef. Microbes 2017, 8, 439–449. [Google Scholar] [CrossRef]
- Rattigan, R.; O’Doherty, J.V.; Vigors, S.; Ryan, M.T.; Sebastiano, R.S.; Callanan, J.J.; Thornton, K.; Rajauria, G.; Margassery, L.M.; Dobson, A.D.W.; et al. The effects of the marine-derived polysaccharides laminarin and chitosan on aspects of colonic health in pigs challenged with dextran sodium sulphate. Mar. Drugs 2020, 18, 262. [Google Scholar] [CrossRef]
- Khosravi-Katuli, K.; Prato, E.; Lofrano, G.; Guida, M.; Vale, G.; Libralato, G. Effects of nanoparticles in species of aquaculture interest. Environ. Sci. Pollut. Res. 2017, 24, 17326–17346. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Zommara, M.; Eweedah, N.M.; Helal, A.I. Synergistic effects of selenium nanoparticles and vitamin e on growth, immune-related gene expression, and regulation of antioxidant status of nile tilapia (Oreochromis niloticus). Biol. Trace Elem. Res. 2020, 195, 624–635. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Naby, F.S.; Naiel, M.A.E.; Al-Sagheer, A.A.; Negm, S.S. Dietary chitosan nanoparticles enhance the growth, production performance, and immunity in Oreochromis niloticus. Aquaculture 2019, 501, 82–89. [Google Scholar] [CrossRef]
- Abdel-Tawwab, M.; Razek, N.A.; Abdel-Rahman, A.M. Immunostimulatory effect of dietary chitosan nanoparticles on the performance of nile tilapia, Oreochromis niloticus (L.). Fish Shellfish Immunol. 2019, 88, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Naiel, M.A.E.; Ismael, N.E.M.; Abd El-hameed, S.A.A.; Amer, M.S. The anti-oxidative and immunity roles of chitosan nanoparticle and vitamin c-supplemented diets against imidacloprid toxicity on Oreochromis niloticus. Aquaculture 2020, 523, 735219. [Google Scholar] [CrossRef]
- Abd El-Naby, A.S.; Al-Sagheer, A.A.; Negm, S.S.; Naiel, M.A.E. Dietary combination of chitosan nanoparticle and thymol affects feed utilization, digestive enzymes, antioxidant status, and intestinal morphology of Oreochromis niloticus. Aquaculture 2020, 515, 734577. [Google Scholar] [CrossRef]
- Khani Oushani, A.; Soltani, M.; Sheikhzadeh, N.; Shamsaie Mehrgan, M.; Rajabi Islami, H. Effects of dietary chitosan and nano-chitosan loaded clinoptilolite on growth and immune responses of rainbow trout (Oncorhynchus mykiss). Fish Shellfish Immunol. 2020, 98, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Younus, N.; Zuberi, A.; Mahmoood, T.; Akram, W.; Ahmad, M. Comparative effects of dietary micro- and nano-scale chitosan on the growth performance, non-specific immunity, and resistance of silver carp Hypophthalmichthys molitrix against Staphylococcus aureus infection. Aquac. Int. 2020, 28, 2363–2378. [Google Scholar] [CrossRef]
- Chen, J.; Chen, L. Effects of chitosan-supplemented diets on the growth performance, nonspecific immunity and health of loach fish (Misgurnus anguillicadatus). Carbohydr. Polym. 2019, 225, 115227. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Zhang, Y.; Liu, W.; Xu, L.; Yang, Y.; Zhou, Z. Effects of chito-oligosaccharides supplementation on growth performance, intestinal cytokine expression, autochthonous gut bacteria and disease resistance in hybrid tilapia Oreochromis niloticus ♀ × Oreochromis aureus ♂. Fish Shellfish Immunol. 2014, 40, 267–274. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Koshio, S. Application of fermentation strategy in aquafeed for sustainable aquaculture. Rev. Aquac. 2020, 12, 987–1002. [Google Scholar] [CrossRef]
- Zhang, B. Dietary chitosan oligosaccharides modulate the growth, intestine digestive enzymes, body composition and nonspecific immunity of loach Paramisgurnus dabryanus. Fish Shellfish Immunol. 2019, 88, 359–363. [Google Scholar] [CrossRef]
- Haygood, A.M.; Jha, R. Strategies to modulate the intestinal microbiota of tilapia (Oreochromis sp.) in aquaculture: A review. Rev. Aquac. 2018, 10, 320–333. [Google Scholar] [CrossRef]
- Zaki, M.; Shatby, E.; Shatby, E. Effect of chitosan supplemented diet on survival, growth, feed utilization, body composition & histology of sea bass (Dicentrarchus labrax). World J. Eng. Technol. 2015, 3, 38. [Google Scholar]
- Liang, F.; Li, C.; Hou, T.; Wen, C.; Kong, S.; Ma, D.; Sun, C.; Li, S. Effects of chitosan–gentamicin conjugate supplement on non-specific immunity, aquaculture water, intestinal histology and microbiota of pacific white shrimp (Litopenaeus vannamei). Mar. Drugs 2020, 18, 419. [Google Scholar] [CrossRef]
- Şahan, A.; Özütok, S.; Kurutaş, E.B. Determination of some hematological parameters and antioxidant capacity in nile tilapia (Oreochromis niloticus linnaeus, 1758) fed ginger (Zingiber officinale roscoe) to Aeromonas hydrophila. Turk. J. Fish. Aquat. Sci. 2016, 16, 197–204. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Zommara, M.; Eweedah, N.M.; Helal, A.I. The evaluation of growth performance, blood health, oxidative status and immune-related gene expression in nile tilapia (Oreochromis niloticus) fed dietary nanoselenium spheres produced by lactic acid bacteria. Aquaculture 2020, 515, 734571. [Google Scholar] [CrossRef]
- Grinde, B.; Lie, Ø.; Poppe, T.; Salte, R. Species and individual variation in lysozyme activity in fish of interest in aquaculture. Aquaculture 1988, 68, 299–304. [Google Scholar] [CrossRef]
- Seeley, K.R.; Gillespie, P.D.; Weeks, B.A. A simple technique for the rapid spectrophotometric determination of phagocytosis by fish macrophages. Mar. Environ. Res. 1990, 30, 37–41. [Google Scholar] [CrossRef]
- Victor, H.; Zhao, B.; Mu, Y.; Dai, X.; Wen, Z.; Gao, Y.; Chu, Z. Effects of se-chitosan on the growth performance and intestinal health of the loach Paramisgurnus dabryanus (sauvage). Aquaculture 2019, 498, 263–270. [Google Scholar] [CrossRef]
- Vasdev, S.; Gill, V.D.; Singal, P.K. Modulation of oxidative stress-induced changes in hypertension and atherosclerosis by antioxidants. Exp. Clin. Cardiol. 2006, 11, 206–216. [Google Scholar]
- Ngo, D.-H.; Wijesekara, I.; Vo, T.-S.; Van Ta, Q.; Kim, S.-K. Marine food-derived functional ingredients as potential antioxidants in the food industry: An overview. Food Res. Int. 2011, 44, 523–529. [Google Scholar] [CrossRef]
- Winzer, K.; Becker, W.; Van Noorden, C.J.F.; Köhler, A. Short-time induction of oxidative stress in hepatocytes of the european flounder (Platichthys flesus). Mar. Environ. Res. 2000, 50, 495–501. [Google Scholar] [CrossRef]
- Draper, H.H.; Hadley, M. [43] malondialdehyde determination as index of lipid peroxidation. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1990; Volume 186, pp. 421–431. [Google Scholar]
- Abdel-Ghany, H.M.; Salem, M.E.-S. Effects of dietary chitosan supplementation on farmed fish; A review. Rev. Aquac. 2020, 12, 438–452. [Google Scholar] [CrossRef]
- Niu, J.; Lin, H.-Z.; Jiang, S.-G.; Chen, X.; Wu, K.-C.; Liu, Y.-J.; Wang, S.; Tian, L.-X. Comparison of effect of chitin, chitosan, chitosan oligosaccharide and n-acetyl-d-glucosamine on growth performance, antioxidant defenses and oxidative stress status of Penaeus monodon. Aquaculture 2013, 372–375, 1–8. [Google Scholar] [CrossRef]
- El-Deep, M.H.; Amber, K.A.; Elgendy, S.; Dawood, M.A.O.; Zidan, A. In ovo injection of nano-selenium spheres mitigates the hatchability, histopathology image and immune response of hatched chicks. J. Anim. Physiol. Anim. Nutr. 2020. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.-X.; Qian, J.-Q.; Shi, L.-E. Preparation of chitosan nanoparticles as carrier for immobilized enzyme. Appl. Biochem. Biotechnol. 2007, 136, 77–96. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.O.; Amer, A.A.; Elbialy, Z.I.; Gouda, A.H. Effects of including triticale on growth performance, digestive enzyme activity, and growth-related genes of nile tilapia (Oreochromis niloticus). Aquaculture 2020, 528, 735568. [Google Scholar] [CrossRef]
- Houston, A. Blood and circulation/methods for fish biology. Ny. Amer. Fish. Society. Jain nc. In Schalm’s Veterinary Hematology; Lea Febiger: Philadelphia, PA, USA, 1990; pp. 21–62. [Google Scholar]
- Lucky, Z. Methods for the Diagnosis of Fish Diseases, Amerind; Publishing Co. Pvt. Ltd.: New Delhi, Bombay, India, 1977. [Google Scholar]
- Jain, N.C. Schalm’s Veterinary Hematology; Lea & Febiger: Philadelphia, PA, USA, 1986. [Google Scholar]
- Blaxhall, P.C.; Daisley, K.W. Routine haematological methods for use with fish blood. J. Fish Biol. 1973, 5, 771–781. [Google Scholar] [CrossRef]
- Doumas, B.T.; Bayse, D.D.; Carter, R.J.; Peters, T.; Schaffer, R. A candidate reference method for determination of total protein in serum. I. Development and validation. Clin. Chem. 1981, 27, 1642–1650. [Google Scholar] [CrossRef] [PubMed]
- Dumas, B.T.; Biggs, H.G. Standard Methods of Clinical Chemistry; Academic Press: New York, NY, USA, 1972. [Google Scholar]
- Reitman, S.; Frankel, S. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef]
- Coulombe, J.J.; Favreau, L. A new simple semimicro method for colorimetric determination of urea. Clin. Chem. 1963, 9, 102–108. [Google Scholar] [CrossRef]
- Heinegård, D.; Tiderström, G. Determination of serum creatinine by a direct colorimetric method. Clin. Chim. Acta 1973, 43, 305–310. [Google Scholar] [CrossRef]
- Parry, J.; Richard, M.; Chandan, R.C.; Shahani, K.M. A rapid and sensitive assay of muramidase. Proc. Soc. Exp. Biol. Med. 1965, 119, 384–386. [Google Scholar] [CrossRef]
- Kawahara, E.; Ueda, T.; Nomura, S. In vitro phagocytic activity of white-spotted char blood cells after injection with Aeromonas salmonicida extracellular products. Fish Pathol. 1991, 26, 213–214. [Google Scholar] [CrossRef]
- Ledwozyw, A.; Michalak, J.; Stepień, A.; Kadziołka, A. The relationship between plasma triglycerides, cholesterol, total lipids and lipid peroxidation products during human atherosclerosis. Clin. Chim. Acta Int. J. Clin. Chem. 1986, 155, 275–283. [Google Scholar] [CrossRef]
- Gewaily, M.S.; Kassab, M.; Farrag, F.A.; Almadaly, E.A.; Atta, M.S.; Abd-Elmaksoud, A.; Wakayama, T. Comparative expression of cell adhesion molecule1 (cadm1) in the testes of experimental mice and some farm animals. Acta Histochem. 2020, 122, 151456. [Google Scholar] [CrossRef] [PubMed]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques; Elsevier Health Sciences: New York, NY, USA, 2008. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. Nih image to imagej: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).