Effect of Nitrogen Sources on Omega-3 Polyunsaturated Fatty Acid Biosynthesis and Gene Expression in Thraustochytriidae sp.




Abstract
1. Introduction
2. Results
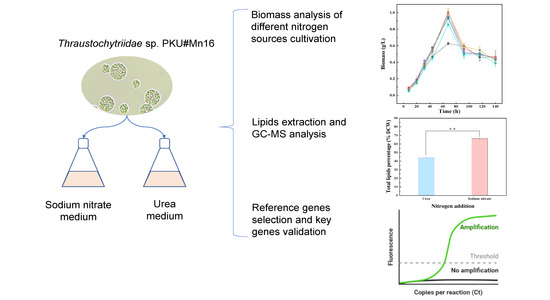
2.1. Effects of Sodium Nitrate and Urea on the Growth of Thraustochytriidae sp. PKU#Mn16
2.2. Effects of Sodium Nitrate and Urea on Fatty Acid Contents and Composition in Thraustochytriidae sp. PKU#Mn16
2.3. Screen of the Reference Genes for RT-qPCR
2.4. Effect of Various Nitrogen Sources on the Expression of the Key FA Synthesis Genes of Thraustochytriidae sp. PKU#Mn16
3. Discussion
3.1. Essential Genes Involved in FAS and PKS Pathways of Fatty Acid Production
3.2. Effects of Nitrate Reductase on Nitrogen Assimilation and Lipids Production
4. Materials and Methods
4.1. Microorganism Cultivation
4.2. Lipids Extraction and Fatty Acid Composition Analysis
4.3. RNA Extraction and cDNA Synthesis
4.4. Selection of Candidate Reference Genes from RNA-Seq Data
4.5. Real-Time Quantitative Polymerase Chain Reaction (RT-qPCR)
4.6. Assessment of Gene Expression Stability
4.7. Validation of Reference Genes
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- De Carvalho, C.; Caramujo, M.J. The Various Roles of Fatty Acids. Molecules 2018, 23. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Keum, Y.-S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.S.; Sampson, E.L. Fatty acid and fatty aldehyde composition of the major brain lipids in normal human gray matter, white matter, and myelin. J. Lipid Res. 1965, 6, 545–551. [Google Scholar] [PubMed]
- Guesnet, P.; Alessandri, J.-M. Docosahexaenoic acid (DHA) and the developing central nervous system (CNS) —Implications for dietary recommendations. Biochimie 2011, 93, 7–12. [Google Scholar] [CrossRef]
- Bernasconi, A.A.; Wiest, M.M.; Lavie, C.J.; Milani, R.V.; Laukkanen, J.A. Effect of Omega-3 Dosage on Cardiovascular Outcomes: An Updated Meta-Analysis and Meta-Regression of Interventional Trials. Mayo Clin. Proc. 2020. [Google Scholar] [CrossRef]
- Abdelhamid, A.S.; Brown, T.J.; Brainard, J.S.; Biswas, P.; Thorpe, G.C.; Moore, H.J.; Deane, K.H.; AlAbdulghafoor, F.K.; Summerbell, C.D.; Worthington, H.V.; et al. Omega-3 fatty acids for the primary and secondary prevention of cardiovascular disease. Cochrane Database Syst. Rev. 2018, 11, Cd003177. [Google Scholar] [CrossRef]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes: From molecules to man. Biochem. Soc. Trans. 2017, 45, 1105–1115. [Google Scholar] [CrossRef]
- Gillet, L.; Roger, S.; Bougnoux, P.; Le Guennec, J.-Y.; Besson, P. Beneficial effects of omega-3 long-chain fatty acids in breast cancer and cardiovascular diseases: Voltage-gated sodium channels as a common feature? Biochimie 2011, 93, 4–6. [Google Scholar] [CrossRef]
- Ward, O.P.; Singh, A. Omega-3/6 fatty acids: Alternative sources of production. Process Biochem. 2005, 40, 3627–3652. [Google Scholar] [CrossRef]
- Adarme-Vega, T.C.; Thomas-Hall, S.R.; Schenk, P.M. Towards sustainable sources for omega-3 fatty acids production. Curr. Opin. Biotechnol. 2014, 26, 14–18. [Google Scholar] [CrossRef]
- Yokochi, T.; Honda, D.; Higashihara, T.; Nakahara, T. Optimization of docosahexaenoic acid production by Schizochytrium limacinum SR21. Appl. Microbiol. Biotechnol. 1998, 49, 72–76. [Google Scholar] [CrossRef]
- Byreddy, A.R.; Barrow, C.J.; Puri, M. Bead milling for lipid recovery from thraustochytrid cells and selective hydrolysis of Schizochytrium DT3 oil using lipase. Bioresour. Technol. 2016, 200, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Li, S.; Liu, L.; Li, S.; Luo, Y.; Lv, C.; Wang, B.; Cheng, C.H.; Chen, H.; Yang, X.J.M.D. Genome Sequencing and Analysis of Thraustochytriidae sp. SZU445 Provides Novel Insights into the Polyunsaturated Fatty Acid Biosynthesis Pathway. Mar. Drugs 2020, 18, 118. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.-J.; Sun, L.-N.; Zhuang, X.-Y.; Qu, L.; Ji, X.-J.; Huang, H. Regulation of docosahexaenoic acid production by Schizochytrium sp.: Effect of nitrogen addition. Bioprocess Biosyst. Eng. 2014, 37, 865–872. [Google Scholar] [CrossRef]
- Yang, X.; Li, S.; Li, S.; Liu, L.; Hu, Z. De Novo Transcriptome Analysis of Polyunsaturated Fatty Acid Metabolism in Marine Protist Thraustochytriidae sp. PKU#Mn16. J. Am. Oil Chem. Soc. 2019, 97, 35–48. [Google Scholar] [CrossRef]
- Shen, X.F.; Chu, F.F.; Lam, P.K.; Zeng, R.J. Biosynthesis of high yield fatty acids from Chlorella vulgaris NIES-227 under nitrogen starvation stress during heterotrophic cultivation. Water Res 2015, 81, 294–300. [Google Scholar] [CrossRef]
- Chokshi, K.; Pancha, I.; Ghosh, A.; Mishra, S. Nitrogen starvation-induced cellular crosstalk of ROS-scavenging antioxidants and phytohormone enhanced the biofuel potential of green microalga Acutodesmus dimorphus. Biotechnol. Biofuels 2017, 10, 60. [Google Scholar] [CrossRef]
- Chen, B.; Wan, C.; Mehmood, M.A.; Chang, J.-S.; Bai, F.; Zhao, X. Manipulating environmental stresses and stress tolerance of microalgae for enhanced production of lipids and value-added products–A review. Bioresour. Technol. 2017, 244, 1198–1206. [Google Scholar] [CrossRef]
- Jakobsen, A.N.; Aasen, I.M.; Josefsen, K.D.; Strom, A.R. Accumulation of docosahexaenoic acid-rich lipid in thraustochytrid Aurantiochytrium sp. strain T66: Effects of N and P starvation and O2 limitation. Appl. Microbiol. Biotechnol. 2008, 80, 297–306. [Google Scholar] [CrossRef]
- Park, W.-K.; Moon, M.; Shin, S.-E.; Cho, J.M.; Suh, W.I.; Chang, Y.K.; Lee, B. Economical DHA (Docosahexaenoic acid) production from Aurantiochytrium sp. KRS101 using orange peel extract and low cost nitrogen sources. Algal Res. 2018, 29, 71–79. [Google Scholar] [CrossRef]
- Gao, M.; Song, X.; Feng, Y.; Li, W.; Cui, Q. Isolation and characterization of Aurantiochytrium species: High docosahexaenoic acid (DHA) production by the newly isolated microalga, Aurantiochytrium sp. SD116. J. Oleo Sci. 2013, 62, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Paiva-Cavalcanti, M.; Regis-da-Silva, C.G.; Gomes, Y.M. Comparison of real-time PCR and conventional PCR for detection of Leishmania (Leishmania) infantum infection: A mini-review. J. Venom. Anim. Toxins Incl. Trop. Dis. 2010, 16, 537–542. [Google Scholar] [CrossRef]
- Bin, S.; Pu, X.; Shu, B.; Kang, C.; Luo, S.; Tang, Y.; Wu, Z.; Lin, J. Selection of Reference Genes for Optimal Normalization of Quantitative Real-Time Polymerase Chain Reaction Results for Diaphorina citri Adults. J. Econ. Entomol. 2018, 355. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Tan, Y.; Cui, G.; Feng, Y.; Cui, Q.; Song, X. Transcriptome and gene expression analysis of DHA producer Aurantiochytrium under low temperature conditions. Sci. Rep. 2015, 5, 14446. [Google Scholar] [CrossRef]
- Chen, W.; Zhou, P.-p.; Zhang, M.; Zhu, Y.-m.; Wang, X.-p.; Luo, X.-a.; Bao, Z.-d.; Yu, L.-j. Transcriptome analysis reveals that up-regulation of the fatty acid synthase gene promotes the accumulation of docosahexaenoic acid in Schizochytrium sp. S056 when glycerol is used. Algal Res. 2016, 15, 83–92. [Google Scholar] [CrossRef]
- Ye, J.; Liu, M.; He, M.; Ye, Y.; Huang, J. Illustrating and Enhancing the Biosynthesis of Astaxanthin and Docosahexaenoic Acid in Aurantiochytrium sp. SK4. Mar Drugs 2019, 17. [Google Scholar] [CrossRef]
- Sagri, E.; Koskinioti, P.; Gregoriou, M.E.; Tsoumani, K.T.; Bassiakos, Y.C.; Mathiopoulos, K.D. Housekeeping in Tephritid insects: The best gene choice for expression analyses in the medfly and the olive fly. Sci. Rep. 2017, 7, 45634. [Google Scholar] [CrossRef]
- Martins, P.K.; Mafra, V.; de Souza, W.R.; Ribeiro, A.P.; Vinecky, F.; Basso, M.F.; da Cunha, B.A.; Kobayashi, A.K.; Molinari, H.B. Selection of reliable reference genes for RT-qPCR analysis during developmental stages and abiotic stress in Setaria viridis. Sci. Rep. 2016, 6, 28348. [Google Scholar] [CrossRef]
- Pinto, F.; Pacheco, C.C.; Ferreira, D.; Moradas-Ferreira, P.; Tamagnini, P. Selection of suitable reference genes for RT-qPCR analyses in cyanobacteria. PLoS ONE 2012, 7, e34983. [Google Scholar] [CrossRef]
- Johnson, T.J.; Jahandideh, A.; Isaac, I.C.; Baldwin, E.L.; Muthukumarappan, K.; Zhou, R.; Gibbons, W.R. Determining the optimal nitrogen source for large-scale cultivation of filamentous cyanobacteria. J. Appl. Phycol. 2016, 29, 1–13. [Google Scholar] [CrossRef]
- Arora, V.; Ghosh, M.K.; Singh, P.; Gangopadhyay, G. Light regulation of Nitrate reductase gene expression and enzyme activity in the leaves of mulberry. Indian J. Biochem. Biophys. 2018, 55, 62–66. [Google Scholar]
- Murphy, M.G.; Spence, M.W. Long-chain fatty acid:CoA ligase in rat brain in vitro: A comparison of activities with oleic and cis-vaccenic acids. J. Neurochem. 1980, 34, 367–373. [Google Scholar] [CrossRef]
- Inagaki, K.; Aki, T.; Fukuda, Y.; Kawamoto, S.; Shigeta, S.; Ono, K.; Suzuki, O. Identification and expression of a rat fatty acid elongase involved in the biosynthesis of C18 fatty acids. Biosci. Biotechnol. Biochem. 2002, 66, 613–621. [Google Scholar] [CrossRef] [PubMed]
- Agnihotri, G.; Liu, H.-w. Enoyl-CoA hydratase. Bioorg. Med. Chem. 2003, 11, 9–20. [Google Scholar] [CrossRef]
- Maier, T.; Leibundgut, M.; Boehringer, D.; Ban, N. Structure and function of eukaryotic fatty acid synthases. Q. Rev. Biophys. 2010, 43, 373–422. [Google Scholar] [CrossRef] [PubMed]
- Schonauer, M.S.; Kastaniotis, A.J.; Hiltunen, J.K.; Dieckmann, C.L. Intersection of RNA Processing and the Type II Fatty Acid Synthesis Pathway in Yeast Mitochondria. Mol. Cell. Biol. 2008, 28, 6646. [Google Scholar] [CrossRef]
- Ohara, J.; Sakaguchi, K.; Okita, Y.; Okino, N.; Ito, M.J.M.b. Two fatty acid elongases possessing C18-Δ6/C18-Δ9/C20-Δ5 or C16-Δ9 elongase activity in Thraustochytrium sp. ATCC 26185. Mar. Biotechnol. 2013, 15, 476–486. [Google Scholar] [CrossRef] [PubMed]
- Heggeset, T.M.B.; Ertesvag, H.; Liu, B.; Ellingsen, T.E.; Vadstein, O.; Aasen, I.M. Lipid and DHA-production in Aurantiochytrium sp.—Responses to nitrogen starvation and oxygen limitation revealed by analyses of production kinetics and global transcriptomes. Sci. Rep. 2019, 9, 19470. [Google Scholar] [CrossRef]
- Cook, O.; Hildebrand, M. Enhancing LC-PUFA production in Thalassiosira pseudonana by overexpressing the endogenous fatty acid elongase genes. J. Appl. Phycol. 2015, 28, 897–905. [Google Scholar] [CrossRef]
- Hamilton, M.L.; Haslam, R.P.; Napier, J.A.; Sayanova, O. Metabolic engineering of Phaeodactylum tricornutum for the enhanced accumulation of omega-3 long chain polyunsaturated fatty acids. Metab. Eng. 2014, 22, 3–9. [Google Scholar] [CrossRef]
- Khozin-Goldberg, I.; Cohen, Z. Unraveling algal lipid metabolism: Recent advances in gene identification. Biochimie 2011, 93, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Hunkeler, M.; Hagmann, A.; Stuttfeld, E.; Chami, M.; Guri, Y.; Stahlberg, H.; Maier, T. Structural basis for regulation of human acetyl-CoA carboxylase. Nature 2018, 558, 470–474. [Google Scholar] [CrossRef]
- Hasan, H.; Abd Rahim, M.H.; Campbell, L.; Carter, D.; Abbas, A.; Montoya, A. Overexpression of acetyl-CoA carboxylase in Aspergillus terreus to increase lovastatin production. New Biotechnol. 2018, 44, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.W.; Da Silva, N.A. Improving polyketide and fatty acid synthesis by engineering of the yeast acetyl-CoA carboxylase. J. Biotechnol. 2014, 187, 56–59. [Google Scholar] [CrossRef] [PubMed]
- Engel, C.K.; Mathieu, M.; Zeelen, J.P.; Hiltunen, J.K.; Wierenga, R.K. Crystal structure of enoyl-coenzyme A (CoA) hydratase at 2.5 angstroms resolution: A spiral fold defines the CoA-binding pocket. EMBO J. 1996, 15, 5135–5145. [Google Scholar] [CrossRef] [PubMed]
- Goepfert, S.; Hiltunen, J.K.; Poirier, Y. Identification and functional characterization of a monofunctional peroxisomal enoyl-CoA hydratase 2 that participates in the degradation of even cis-unsaturated fatty acids in Arabidopsis thaliana. J. Biol. Chem. 2006, 281, 35894–35903. [Google Scholar] [CrossRef]
- Cao, Y.; Traer, E.; Zimmerman, G.A.; McIntyre, T.M.; Prescott, S.M. Cloning, expression, and chromosomal localization of human long-chain fatty acid-CoA ligase 4 (FACL4). Genomics 1998, 49, 327–330. [Google Scholar] [CrossRef]
- Ratledge, C. Fatty acid biosynthesis in microorganisms being used for Single Cell Oil production. Biochimie 2004, 86, 807–815. [Google Scholar] [CrossRef]
- Morabito, C.; Bournaud, C.; Maes, C.; Schuler, M.; Aiese Cigliano, R.; Dellero, Y.; Marechal, E.; Amato, A.; Rebeille, F. The lipid metabolism in thraustochytrids. Prog. Lipid Res. 2019, 76, 101007. [Google Scholar] [CrossRef]
- Rai, V.; Sarkar, S.; Satpati, S.; Dey, N. Overexpression of human peroxisomal enoyl-CoA delta isomerase2 HsPECI2, an ortholog of bamboo expressed during gregarious flowering alters salinity stress responses and polar lipid content in tobacco. Funct. Plant Biol. 2016, 43, 232–243. [Google Scholar] [CrossRef]
- González, P.J.; Correia, C.; Moura, I.; Brondino, C.D.; Moura, J.J.G. Bacterial nitrate reductases: Molecular and biological aspects of nitrate reduction. J. Inorg. Biochem. 2006, 100, 1015–1023. [Google Scholar] [CrossRef] [PubMed]
- Hong, D.D.; Anh, H.T.; Thu, N.T. Study on biological characteristics of heterotrophic marine microalga-Schizochytrium Mangrove PQ6 isolated from Phu Quoc island, Kien Giang Province, Vietnam. J. Phycol. 2011, 47, 944–954. [Google Scholar] [CrossRef] [PubMed]
- Auma, K.; Hamid, A.A.; Yusoff, W.M.W. Effect of nitrogen sources on biomass, lipid and docosahexanoic acid production by Aurantiochytrum sp. SW1. In Proceedings of AIP Conference Proceedings; AIP Publishing: Melville, NY, USA, 1940; p. 020065. [Google Scholar]
- Crawford, N.M.; Arst, H.N., Jr. The molecular genetics of nitrate assimilation in fungi and plants. Annu. Rev. Genet. 1993, 27, 115–146. [Google Scholar] [CrossRef]
- Sanz-Luque, E.; Chamizo-Ampudia, A.; Llamas, A.; Galvan, A.; Fernandez, E. Understanding nitrate assimilation and its regulation in microalgae. Front. Plant Sci. 2015, 6, 899. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, J.K.; Smith, S.R.; McCrow, J.P.; Tan, M.; Zheng, H.; Beeri, K.; Roth, R.; Lichtle, C.; Goodenough, U.; Bowler, C.P.; et al. Nitrate Reductase Knockout Uncouples Nitrate Transport from Nitrate Assimilation and Drives Repartitioning of Carbon Flux in a Model Pennate Diatom. Plant Cell 2017, 29, 2047–2070. [Google Scholar] [CrossRef] [PubMed]
- Syrett, P.; Leftley, J. Nitrate and urea assimilation by algae. Perspect. Exp. Biol. 2016, 2, 221–234. [Google Scholar]
- Taoka, Y.; Nagano, N.; Okita, Y.; Izumida, H.; Sugimoto, S.; Hayashi, M. Extracellular enzymes produced by marine eukaryotes, thraustochytrids. Biosci. Biotechnol. Biochem. 2009, 73, 180–182. [Google Scholar] [CrossRef]
- Benhima, R.; El Arroussi, H.; Kadmiri, I.M.; El Mernissi, N.; Wahby, I.; Bennis, I.; Smouni, A.; Bendaou, N. Nitrate Reductase Inhibition Induces Lipid Enhancement of Dunaliella Tertiolecta for Biodiesel Production. Sci. World J. 2018, 2018, 6834725. [Google Scholar] [CrossRef]
- Sun, L.; Ren, L.; Zhuang, X.; Ji, X.; Yan, J.; Huang, H. Differential effects of nutrient limitations on biochemical constituents and docosahexaenoic acid production of Schizochytrium sp. Bioresour. Technol. 2014, 159, 199–206. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Liu, Y.; Singh, P.; Sun, Y.; Luan, S.; Wang, G. Culturable diversity and biochemical features of thraustochytrids from coastal waters of Southern China. Appl. Microbiol. Biotechnol. 2014, 98, 3241–3255. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Zhang, J.; Zhao, J.; Gao, Z.; Zhang, C.; Chen, M. Regulation of lipid accumulation in Schizochytrium sp. ATCC 20888 in response to different nitrogen sources. Eur. J. Lipid Sci. Technol. 2017, 119. [Google Scholar] [CrossRef]
- Carvalho, D.M.; de Sa, P.H.; Castro, T.L.; Carvalho, R.D.; Pinto, A.; Gil, D.J.; Bagano, P.; Bastos, B.; Costa, L.F.; Meyer, R.; et al. Reference genes for RT-qPCR studies in Corynebacterium pseudotuberculosis identified through analysis of RNA-seq data. Antonie Van Leeuwenhoek 2014, 106, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Irvine, N.A.; Ruyter, B.; Ostbye, T.K.; Sonesson, A.K.; Lillycrop, K.A.; Berge, G.; Burdge, G.C. Dietary Fish Oil Alters DNA Methylation of Genes Involved in Polyunsaturated Fatty Acid Biosynthesis in Muscle and Liver of Atlantic Salmon (Salmo salar). Lipids 2019, 54, 725–739. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Symbol | Primer Sequence (5′–3′) (Forward/Reverse) | Product Size (bp) | RT-qPCR Efficiency (%) | R2 |
|---|---|---|---|---|---|
| Chitinase | CHIA | GACCCGCTCACCTACTTCAA | 118 | 101.616 | 0.998 |
| TCTGTCATCTGCTGCTCCAC | |||||
| Ribose-phosphate pyrophosh-okinase | RPPK | GAGGCCGTGGGTAGTAAAGG | 172 | 98.02 | 0.996 |
| GGATTGGACTGAGGAGGAGC | |||||
| Nucleolin | NUC | GACGAGCGTGAACTTGAGCG | 106 | 109.201 | 0.994 |
| AACCAGACGAAGAGGACGAG | |||||
| Mitochondrial folate transporter | MFT | ACACTACCGCAGCCTATCAC | 113 | 97.816 | 0.995 |
| ATCCATCTGTCAAGCCATCC | |||||
| Calcium dependent protein kinase I | CAMK1 | CACCAGAACGGCATCATCCA | 154 | 95.226 | 0.997 |
| GACCAAAGTCACAAAGCACC | |||||
| Heat shock transcription factor | HSF | TCCCTTCAGTTTCACCACAT | 192 | 102.598 | 0.998 |
| AAGCACCCACTATTCCAACG | |||||
| Vacuolar protein sorting-associated protein | VPS | CCCCAGAAGGAAACCATCAC | 164 | 102.201 | 0.996 |
| TTCATCGCACAGCAGTAGGC | |||||
| Saccharolysin | SAC | TAAGGGTCCAAGAAGAATGA | 100 | 99.577 | 0.996 |
| CTGACGGCGAAGTTCCTGTG | |||||
| Ribosome-binding ATPase | RBATP | CCTTGGGCATGTCTACTTCT | 126 | 97.341 | 0.998 |
| GTCTGAAACGAGCGAACACC | |||||
| H+-transporting ATPase | HTATP | TATCCAACCGTAGCCACAGA AGGGTTTCGAGTAGGAGTGC | 123 | 102.301 | 0.998 |
| Method | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 |
|---|---|---|---|---|---|---|---|---|---|---|
| Delta Ct | MFT | NUC | HSF | CHIA | CAMK | RPPK | VPS | SAC | RBATP | HTATP |
| BestKeeper | HTATP | SAC | RPPK | CHIA | RBATP | MFT | NUC | HSF | CAMK | VPS |
| NormFinder | MFT | CHIA | NUC | HSF | CAMK | RPPK | VPS | RBATP | SAC | HTATP |
| geNorm | VPS | HSF | CAMK | NUC | MFT | CHIA | RPPK | SAC | RBATP | HTATP |
| Comprehensive ranking | MFT | NUC | CHIA | HSF | VPS | RPPK | CAMK | HTATP | SAC | RBATP |
| Nitrogen Source | Species | Culture Time | Culture Mode | Lipid Content (%) | References | |
|---|---|---|---|---|---|---|
| TFAs (% DCW) | DHA (%TFAs) | |||||
| Orange peel extract & sodium nitrate | Aurantiochytrium sp. KRS101 | 108 h | baffled flasks | 50.23% | 28.51% | [20] |
| Yeast extract | Schizochytrium sp. PQ6 | 96 h | 10 L fermenter | 38.67% | 43.58% | [52] |
| Monosodium glutamate | Aurantiochytrium sp. SW1 | 96 h | Shake flasks | 79.60% | 47.90% | [53] |
| Ammonium nitrate | Aurantiochytrium sp. SW1 | 96 h | Shake flasks | 46.76% | 31.25% | [53] |
| Sodium nitrate | Thraustochytriidae sp. PKU#Mn16 | 68 h | Shake flasks | 66.27% | 44.51% | This study |
| Urea | Thraustochytriidae sp. PKU#Mn16 | 68 h | Shake flasks | 44.34% | 49.49% | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Hu, Z.; Yang, X.; Li, Y. Effect of Nitrogen Sources on Omega-3 Polyunsaturated Fatty Acid Biosynthesis and Gene Expression in Thraustochytriidae sp. Mar. Drugs 2020, 18, 612. https://doi.org/10.3390/md18120612
Li S, Hu Z, Yang X, Li Y. Effect of Nitrogen Sources on Omega-3 Polyunsaturated Fatty Acid Biosynthesis and Gene Expression in Thraustochytriidae sp. Marine Drugs. 2020; 18(12):612. https://doi.org/10.3390/md18120612
Chicago/Turabian StyleLi, Siting, Zhangli Hu, Xuewei Yang, and Yan Li. 2020. "Effect of Nitrogen Sources on Omega-3 Polyunsaturated Fatty Acid Biosynthesis and Gene Expression in Thraustochytriidae sp." Marine Drugs 18, no. 12: 612. https://doi.org/10.3390/md18120612
APA StyleLi, S., Hu, Z., Yang, X., & Li, Y. (2020). Effect of Nitrogen Sources on Omega-3 Polyunsaturated Fatty Acid Biosynthesis and Gene Expression in Thraustochytriidae sp. Marine Drugs, 18(12), 612. https://doi.org/10.3390/md18120612