Abstract
Salinity is a major abiotic stress limiting rice production worldwide. This study aims to elucidate the effects of heading date on salt tolerance in rice. Five near-isogenic lines (NILs) developed from the SL2038/Koshihikari backcross population were grown with or without salt stress. SL2038 is a salt-tolerant line with delayed heading (~18 days) compared to the salt-sensitive background Koshihikari. The results showed that late-heading NILs produced significantly higher plant dry weight, panicle weight, percentage of filled grains, and grain weight (p < 0.05) under long-term salt stress. In Koshihikari, which exhibited delayed heading due to long-day treatment, the percentage of white heads was low, and panicle and grain weights were significantly higher under salt stress. Experiments with different sowing times indicated that late heading, such as sowing in June, resulted in higher grain weights. This is the first report to assess the impact of heading date on agronomic and yield-related traits under salt stress. In conclusion, even with a prolonged salt treatment period, heading during periods of low temperature and solar radiation results in higher grain weight under salt stress. This is proposed as one of the strategies for salt escape. These findings can be used to improve rice yield and implement crop management in salt-affected regions.
1. Introduction
Rice is one of the most important cereal crops and serves as a primary dietary source and staple food for half of the world’s population. Global crop production must double by 2050 to meet the growing food demand [1]. Salinity has become one of the major abiotic stresses affecting rice production worldwide, particularly in coastal and arid regions where there is increasing salinization of agricultural soils [2,3]. Recent studies have confirmed that salinization is an escalating threat to rice yields due to ionic imbalance and oxidative damage [4,5]. Further, the stable growth of rice production has been a matter of concern in achieving food security, especially in developing countries [6]. Over the past few decades, salinization has intensified due to human activities and natural factors, such as tidal flooding and a decline in freshwater availability. Currently, soil salinity affects approximately 1 billion hectares of land worldwide [7]. Therefore, there is an urgent need to develop more salt-tolerant rice varieties and effective crop management practices to ensure stable rice production in salt-affected regions worldwide.
Salt tolerance is vital during the seedling and reproductive stages, with the latter being particularly crucial, as it greatly affects grain yield [8]. Salinity stress can cause yield losses exceeding 50% at the reproductive stage [9]. This sensitivity is commonly attributed to reductions in plant dry weight, panicle number, spikelets per panicle, grain-filling percentage, and grain yield [10]. This vulnerability is exacerbated by combined high-temperature salt interactions, which disrupt pollen viability and spikelet fertility [11]. Hence, understanding how rice responds to salt stress is crucial for developing rice cultivars that can withstand such conditions [12]. Changes in climatic conditions, such as daily average temperature and sunshine hours during the reproductive stages, can significantly impact both rice yield and grain quality traits [13,14]. This situation highlights the need for the development of new salt-tolerant rice varieties and the implementation of effective crop management practices to ensure sustainable rice production [15]. Severe stress should be avoided during the reproductive growth stages of plants. In this context, the significant impact of climatic variables on agronomic and yield-related traits under salt stress remains largely unknown.
The combined effect of high temperature and salt stress is often more damaging than their individual impacts. Heading date or flowering time is a vital agronomic trait that plays a critical role in the seasonal and regional adaptation of rice varieties [16,17]. The heading stage, a critical reproductive phase, is severely affected by this combination, as both temperature and salt stresses disrupt essential physiological processes [18]. Recent research indicates that changes in heading date can affect both salt tolerance and rice yield [19]. One study showed that among the tested phenology groups related to crop maturity, short-duration rice varieties are predicted to exhibit better tolerance due to the escape period from salinity stress [20]. Another study suggested that adjusting rice transplanting schedules could help avoid salinity stress, particularly during crucial growth stages, such as the booting and flowering stages [21]. While several studies have described drought escape [22], salt escape has rarely been reported in rice. To date, no study has determined whether early or late heading improves salt tolerance and yield-related traits under salt stress. We hypothesized that avoiding high temperature and solar radiation during the heading period can alleviate salt stress by affecting agronomic and yield-related traits in rice. Therefore, this study aims to investigate the effects of differences in heading dates and yield-related traits under long-term salt stress by using NILs, long-day treatments, and different sowing periods. From these results, salt escape is discussed.
2. Materials and Methods
2.1. Plant Materials
SL2038 and Koshihikari seeds were obtained from the Rice Genome Resource Center in Japan. In Experiment 1, five NILs derived from the SL2038/Koshihikari backcross progeny and their parents were evaluated under control and salt stress conditions in 2019 to investigate the effects of differences in heading dates on growth and yield under long-term salt conditions. SL2038 showed salt tolerance and delayed heading by approximately two weeks compared to the salt-sensitive Koshihikari [23]. SL2038 contains IR64 chromosomal segments on chromosomes 3, 10, and 11 in the Koshihikari background (Figure 1).
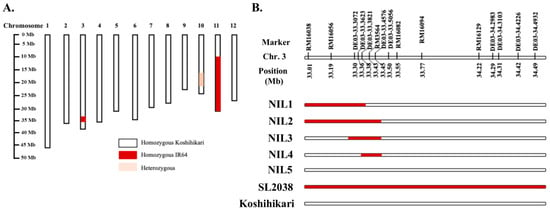
Figure 1.
Graphical illustration of SL2038 and five Koshihikari-NILs with two parents. SL2038 carries chromosomal regions derived from IR64 on chromosomes 3, 10, and 11 in the genetic background of Koshihikari (A). Development of NILs from the SL2038/Koshihikari backcross population with or without the IR64 allele in the genomic regions between 33.01 and 34.49 Mb on chromosome 3 (B). Red and white bars indicate chromosome segments for homozygous IR64 and Koshihikari, respectively.
Five NILs were developed with or without the IR64 allele in the 33.01–34.49 Mb region of chromosome 3. A major heading date gene, Hd16, located on chromosome 3, exhibits functional differences between the IR64 and Koshihikari alleles [24]. Consequently, these NILs have the potential for studies focusing on heading date. In Experiment 2, the same rice variety, Koshihikari, was subjected to one month of long-day treatment to alter the heading date in 2024 and 2025. In Experiment 3, Koshihikari and two late-heading lines in the Koshihikari background (Figure 2) were sown at three different times to examine growth and yield when the growth period and date were changed. L102 and L74 were developed from the SL2038/Koshihikari backcross population with or without the IR64 allele in the physical distance between 32.89 and 34.62 Mb on chromosome 3. Both L102 and L74 are late-heading lines attributed to the presence of the heading date gene Hd16 in this genomic region, making them appropriate for studying heading date in rice.
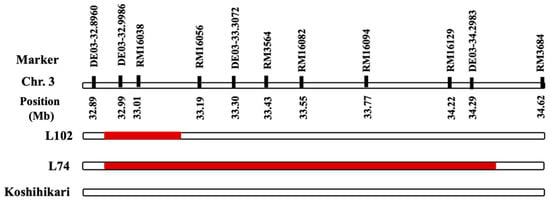
Figure 2.
Development of L102 and L74 from the SL2038/Koshihikari backcross population with or without the IR64 allele in the genomic regions between 32.89 and 34.62 Mb on chromosome 3. Red and white bars indicate chromosome segments for homozygous IR64 and Koshihikari, respectively.
2.2. Growing Conditions
The experiments were conducted in a vinyl house at the Faculty of Agriculture, Okayama University, Japan (34°41′05.4″ N 133°54′58.1″ E). Briefly, rice seeds were surface-sterilized with a mixture of Sumithion (1:200, v/v) and Sportak (1:1000, v/v) for one day. They were then soaked in tap water for two days. Uniformly germinated rice seeds were sown in nursery boxes containing 220 holes (Minoru pot 220, Minoru Industrial Co. Ltd., Okayama, Japan), one seed in each hole. All holes were filled with nursery soil designated for rice seedlings (Kumiai Ube Ryu-joh Baido, Ube Industries Ltd., Tokyo, Japan). The sowing dates were 21 May 2019, 20 May 2024, and 18 May 2025. In 2025, Koshihikari and two late-heading lines in the Koshihikari background (L102 and L74) were sown on 18 April, 18 May, and 17 June. A single seedling, 20 days old, was transplanted into a 2 L pot and arranged in a randomized complete block design. Each pot was filled with paddy soil supplemented with 10 g of NPK (14-14-14) as a basal fertilizer. The experimental pool was irrigated with fresh water before transplanting. For control conditions, no NaCl was added, and normal growth conditions were provided. For salt stress, NaCl was applied to the pool 7 and 10 days after transplanting, from which the days from salt application to heading were calculated. The electrical conductivity (EC) was maintained at approximately 5.5 dS m−1 throughout the growing period until maturity, which was regarded as long-term salt stress. The EC was measured daily using an EC meter (2265FS, Spectrum Technologies, Inc., Aurora, IL, USA). Water was supplied through two irrigation tubes (Evaflow Type-A, Mitsui Chemicals, Tokyo, Japan) on both sides.
2.3. Long-Day Treatment
In Experiment 2, 42 Koshihikari plants were grown with or without salt stress following the growing conditions described above in 2024 and 2025. In 2024, plants were grown in a pool measuring 14.5 m × 1.15 m, arranged with two pots at 30 cm intervals. In 2025, the plants were grown in a larger pool, 14.5 m × 1.70 m, arranged with three pots at 50 cm intervals. Each pool was vertically divided into two sections, one half assigned to control plants and the other to salt stress treatment. All plants were systematically arranged according to their distance from the light source, ranging from 0 to 600 cm in 2024 and 50 to 600 cm in 2025 (Table 1). Long-day treatment was induced with an LED light (Mitsubishi Electric, Model: ELLU47033N, Tokyo, Japan), irradiated for 2 h (8 PM–10 PM) daily for one month, from 28 June to 28 July in both years, to alter the heading date. Supplementary Figure S1 shows details of the recorded light intensity.

Table 1.
Distance from the light source for long-day treatment for Experiment 2 in 2024 and 2025.
2.4. Trait Measurements
The heading date was recorded when the first panicle emerged in each plant. The days to heading were obtained from the difference between the date of sowing and the date of heading. Plants were harvested 40–45 days after the recorded heading date. Stems and leaves were oven-dried at 75 °C for two days, and the plant dry weight was measured. All white heads and normal panicles were manually separated and kept in paper bags. These panicles were dried in the oven at 35 °C for two days, after which the panicle number and weight were measured using normal panicles. The percentage of white heads was calculated using the following equation:
Percentage of white heads = (Number of white heads/Number of total panicles) × 100
The filled spikelets were separated from unfilled grains by sinking in salt water with a specific gravity of 1.06. Then, spikelets per panicle, percentage of filled grains, 1000-grain weight, and grain weight were recorded. In Experiment 2, conducted in 2025, as most of the panicles turned into white heads, yield components were not examined.
2.5. Climatic Variables During the Heading Period
Daily average temperature of each experimental year measured at Okayama University were obtained from the Japan Meteorological Agency (https://www.jma.go.jp/jma/index.html, accessed on 6 January 2026) and daily solar radiation were obtained from the Agro-Meteorological Grid Square Data System operated by the National Agriculture and Food Research Organization (https://amu.rd.naro.go.jp/wiki_open/doku.php?id=start, accessed on 6 January 2026), and analyzed per experimental objectives.
2.6. Statistical Analysis
Statistical analyses were performed using Microsoft Excel 2022, R (version 4.3.0). Differences among means were determined by one-way analysis of variance (ANOVA), and statistical significance was defined at p < 0.05. Data are presented as mean ± standard error (SE), and each observation consists of at least four independent replicates. A correlation heatmap was produced using the ggcorrplot package in R software (version 4.3.0) [25].
3. Results
3.1. Trend of Temperature and Solar Radiation During the Growing Periods
Figure 3 shows the daily average temperature during the growing period from April to October in 2019, 2024, and 2025. Over all three years, the daily average temperature increased steadily from April, exceeding 30 °C by June, reaching a maximum in early August, and gradually declining toward October. Monthly solar radiation was summarized across three periods: early, mid, and late periods of each month (Figure 4). Briefly, in 2024 and 2025, solar radiation was the highest in late July and then gradually decreased toward October. In 2019, it was the highest in May and early August. Comparing the years of the experiment, the average temperature and solar radiation in July and August were higher in 2024 and 2025 than in 2019.
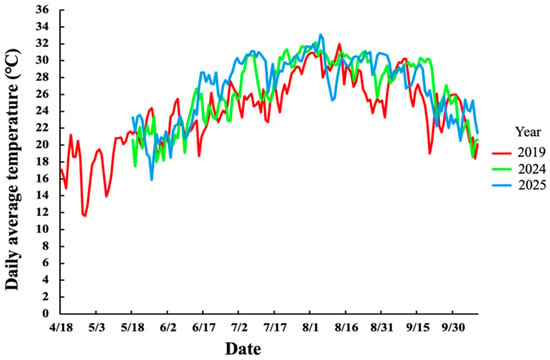
Figure 3.
Daily average temperature (°C) recorded at the experimental site during the growing periods from April to October in 2019, 2024, and 2025.
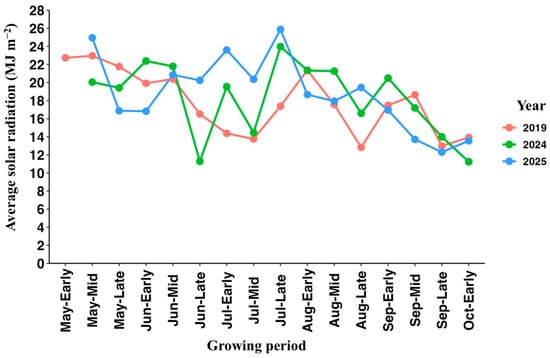
Figure 4.
Average solar radiation (MJ m−2) recorded at the experimental site during three growing periods of each month (early, mid, and late) in 2019, 2024, and 2025. Early = 1–10, Mid = 11–20, and Late = 21–end date; Growing period: April–October.
3.2. Heading Date Among Newly Developed NILs (Experiment 1)
Clear differences were found in heading dates between Koshihikari and SL2038 under both control and salt stress conditions. Koshihikari started heading on 3rd August in control conditions and on 5th August under salt conditions, while SL2038 exhibited delayed heading of approximately two weeks. Under both conditions, NIL3, NIL4, and NIL5 showed early heading similar to Koshihikari, while NIL1 and NIL2 displayed delayed heading comparable to SL2038 (Table 2). This heading pattern was consistent across control and salt stress, allowing classification of NIL1 and NIL2 as late-heading lines and NIL3, NIL4, and NIL5 as early-heading lines. Further, significant differences in days to heading and days from salt application to heading were observed between late-heading NILs and early-heading NILs under both salt and control conditions. These results indicated that late-heading lines experienced a longer period of salt exposure before heading, whereas early-heading lines transitioned to heading more rapidly after salt application.
Table 2.
Heading dates, days to heading, and days from salt application to heading of two parents and five Koshihikari-NILs under control and salt stress conditions in 2019.
3.2.1. Agronomic Parameters and Yield-Related Traits of Early-Heading and Late-Heading NILs (Experiment 1)
The agronomic parameters of five Koshihikari-NILs and two parents showed significant differences under our experimental conditions, except for panicle number under salt stress (Table 3).
Table 3.
Agronomic parameters of five Koshihikari-NILs and two parental lines under control and salt stress conditions in 2019.
The salt-tolerant line SL2038 showed significantly higher plant dry weight and panicle weight than the salt-sensitive Koshihikari under long-term salt stress. Among NILs, the late-heading NILs (NIL1 and NIL2) showed significantly higher plant dry weight than the early-heading NILs (NIL3, NIL4, and NIL5) under control conditions. Similarly, late-heading NILs exhibited significantly higher plant dry weight than early-heading NILs under salt stress. In terms of panicle number, NIL2 showed significantly higher panicle numbers than NIL3 and NIL5 under control conditions. However, there were no significant differences in the panicle number under salt stress. Regarding panicle weight, there were no significant differences among the NILs under control conditions. In contrast, the panicle weight of the late-heading NILs was significantly higher than that of the early-heading NILs under salt stress. Taken together, these findings demonstrated that late-heading NILs exhibited salt tolerance with significant improvements in plant dry weight and panicle weight under salt stress conditions. The yield-related traits of five Koshihikari-NILs and two parental lines exhibited significant differences in all phenotypic traits except for the percentage of filled grains and grain weight under control conditions (Table 4).
Table 4.
Yield components of five Koshihikari-NILs and two parental lines under control and salt stress conditions in 2019.
The salt-tolerant line SL2038 showed significantly higher spikelets per panicle, percentage of filled grains, and grain weight than the salt-sensitive Koshihikari under long-term salt stress. Regarding spikelets per panicle, the late-heading NILs (NIL1 and NIL2) showed significantly higher spikelets per panicle compared to NIL3 under control conditions. However, NIL1 showed significantly higher spikelets per panicle than early-heading NILs under salt stress. Concerning 1000-grain weight, the early-heading NILs (NIL3, NIL4, and NIL5) showed significantly higher 1000-grain weight than late-heading NILs. For the percentage of filled grains and grain weight, significant differences were observed only under salt stress. The late-heading NILs exhibited significantly higher percentages of filled grains than early-heading NILs. Similarly, late-heading NILs produced significantly higher grain weight than early-heading NILs under salt stress. Overall, these results demonstrated that late-heading NILs had significant improvements in yield-related traits under salt stress, especially for the percentage of filled grains and grain weight.
3.2.2. Relationship Between Heading Date and Plant Dry Weight, Grain Weight, and Percentage of White Head (Experiment 1)
We examined the relationship between heading date and plant dry weight, grain weight, and percentage of white heads among five Koshihikari-NILs and their parents (Figure 5). In Experiment 1, the parental line SL2038 and late-heading NILs consistently exhibited higher plant dry weight than Koshihikari and early-heading NILs under both control and salt stress. Grain weight did not differ markedly between early-heading and late-heading NILs under control conditions. However, SL2038 and late-heading NILs tended to maintain higher grain weight compared to Koshihikari and early-heading NILs under salt stress. In contrast, Koshihikari and early-heading NILs showed a higher percentage of white heads than late-heading NILs and SL2038. Overall, these findings revealed a positive association between delayed heading and improved grain weight under saline conditions, suggesting that delayed heading in August contributed to enhanced salt tolerance.
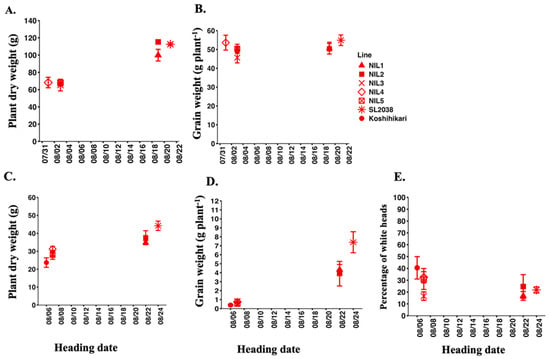
Figure 5.
Relationship between heading date and plant dry weight, grain weight, and percentage of white heads of five Koshihikari-NILs and two parents in 2019. The upper panels represent control conditions (A,B) and the lower ones salt stress conditions (C–E). Data points indicate mean ± SE (n = 4).
3.2.3. Correlation Between Agronomic Parameters and Yield-Related Traits (Experiment 1)
In Experiment 1, the correlation analyses revealed distinct trait relationships under control and salt stress conditions (Figure 6). Under control conditions, plant dry weight was strongly correlated with days to heading and spikelets per panicle (p < 0.001). However, 1000-grain weight showed a negative correlation with plant dry weight. Panicle weight was positively correlated with grain weight (p < 0.001) but showed no significant association with plant dry weight, days to heading, and percentage of filled grains (p > 0.05). Under salt stress, plant dry weight and panicle weight exhibited strong positive correlations with days to heading, spikelets per panicle, percentage of filled grains, and grain weight (p < 0.001). Grain weight was positively correlated with plant dry weight, panicle weight, days to heading, spikelets per panicle, and percentage of filled grains (p < 0.001) but not associated with panicle number (p > 0.05). Overall, these results indicate that differences in heading date are closely associated with improved plant dry weight and grain weight under salt stress.
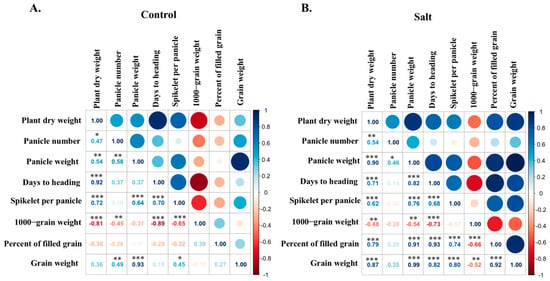
Figure 6.
Correlation heatmap of agronomic parameters and yield-related traits of five Koshihikari-NILs and two parents under control (A) and salt (B) in 2019. *, **, and *** indicate significant differences at the 5%, 1%, and 0.1% levels, respectively.
3.3. Heading Dates and Plant Dry Weight in the Rice Variety Koshihikari After Long-Day Treatment (Experiment 2)
Salt-sensitive Koshihikari plants were subjected to one month of long-day treatment to examine its effect on heading dates under control and salt stress conditions in 2024 and 2025. Six treatment groups were established based on plant distance from the light source. In both years, plant dry weight measured immediately after long-day treatment did not differ between plants under light and those 6 m from the light source, indicating no effect of long-day treatment on plant dry weight (Figure 7A,B).
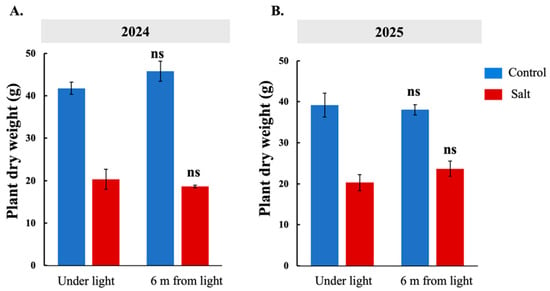
Figure 7.
Effects of one month of long-day treatment in salt-sensitive Koshihikari plants in 2024 and 2025. Comparison of plant dry weight between plants under light and those 6 m from light after one-month long-day treatment (A,B). ns = not significant.
In contrast, heading dates were markedly influenced by the long-day treatment. Plants positioned closer to the light consistently exhibited delayed heading, with days to heading and days from salt application to heading significantly higher under both control and salt stress conditions (Table 5 and Table 6). Overall, these two-year results demonstrated that one month of long-day treatment influenced the heading dates in Koshihikari plants based on their distance from the light source.
Table 5.
Heading dates, days to heading, and days from salt application to heading of salt-sensitive Koshihikari plants corresponding to six long-day treatment groups under control and salt stress conditions in 2024.
Table 6.
Heading dates, days to heading, and days from salt application to heading of salt-sensitive Koshihikari plants corresponding to six long-day treatment groups under control and salt stress conditions in 2025.
3.3.1. Agronomic Parameters and Yield-Related Traits of Six Long-Day Treatment Groups (Experiment 2)
In 2024, significant differences were observed in plant dry weight and panicle weight among the six long-day treatment groups under both control and salt stress (Table 7). Under salt stress, plants positioned closer to the light (T1 and T2) exhibited significantly higher panicle weight than those in distant treatments (T4, T5, and T6).
Table 7.
Agronomic parameters of salt-sensitive Koshihikari plants corresponding to six long-day treatment groups under control and salt stress conditions in 2024.
Yield-related traits also differed significantly among treatments (Table 8). Under control conditions, spikelets per panicle varied mainly in the most distant group (T6). Under salt stress, plants closer to the light (T1 and T2) showed significantly higher percentages of filled grains and grain weight than distant groups (T4, T5, and T6). Taken together, one month of long-day treatment influenced heading date in Koshihikari positioned closer to light by significantly improving the percentage of filled grains and grain weight.
Table 8.
Yield-related traits of salt-sensitive Koshihikari plants corresponding to six long-day treatment groups under control and salt stress conditions in 2024.
3.3.2. Relationship Between Heading Date and Plant Dry Weight, Grain Weight, and Percentage of White Heads in Koshihikari (Experiment 2)
In 2024, after evaluating agronomic parameters and yield-related traits among six long-day treatment groups in salt-sensitive Koshihikari plants, we investigated the relationship between heading date and plant dry weight, panicle weight, and percentage of white heads. Our findings indicated a tendency for plant dry weight and grain weight to increase as heading date became later under control conditions (Figure 8). However, there was no difference in plant dry weight under salt stress. The late-heading group, including T1, T2, and T3, demonstrated higher grain weight than the early-heading group (T4, T5, and T6). Additionally, the early-heading group exhibited a higher percentage of white heads than the late-heading groups. Overall, these results indicate that late-heading plants tended to increase both plant dry weight and grain weight under salt stress, primarily due to their delayed heading toward the end of the month.
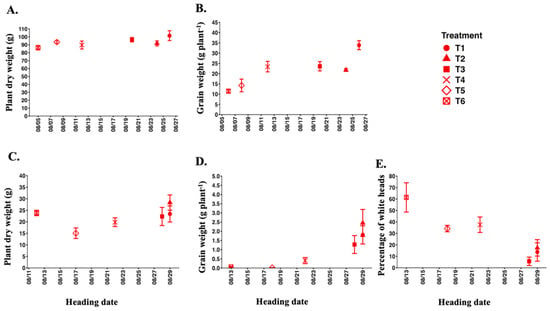
Figure 8.
Relationship between heading date and plant dry weight, grain weight, and percentage of white heads of salt-sensitive Koshihikari plants corresponding to six long-day treatment groups in 2024. The upper panels represent control conditions (A,B) and the lower ones salt stress conditions (C–E). Data points indicate mean ± SE (n = 6).
In 2025, we investigated the relationship between late heading and salt tolerance in Koshihikari plants by examining the relationship between heading date and plant dry weight, grain weight, and percentage of white heads across six long-day treatment groups. In Experiment 2, no association was observed between heading date and plant dry weight among the six long-day treatment groups under control conditions (Figure 9). There was a tendency for grain weight to increase when heading started later in the month under control conditions. In contrast, there was a tendency for plant dry weight to increase when heading dates were later under salt stress. Similarly, grain weight increased due to later heading dates. Further, the percentage of white heads tended to decrease when the heading dates were late. Overall, these results demonstrated that heading later in the month contributed to higher plant dry weight and grain weight in Koshihikari plants under salt stress.
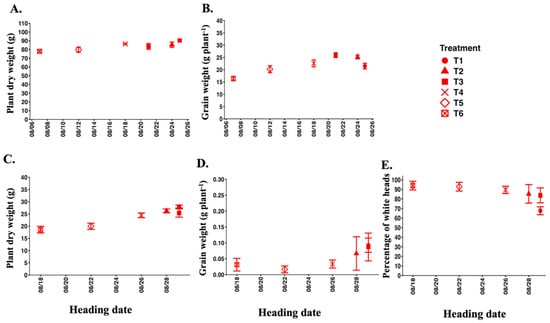
Figure 9.
Relationship between heading date and plant dry weight, grain weight, and percentage of white heads of salt-sensitive Koshihikari plants corresponding to six long-day treatment groups in 2025. The upper panels represent control conditions (A,B) and the lower ones salt stress conditions (C–E). Data points indicate mean ± SE (n = 6).
3.4. Heading Dates Across Different Sowing Periods (Experiment 3)
Koshihikari and two late-heading lines in the Koshihikari background (L102 and L74) were evaluated under long-term salt stress conditions (Table 9). All genotypes showed marked differences in heading date depending on sowing time under both control and salt stress conditions. Koshihikari consistently initiated heading earlier than L102 and L74 across all sowing periods, whereas the late-heading lines predominantly started heading in late August, particularly when sown in May and June. Similar heading patterns were observed under salt stress, with slight delays in June-sown plants. Significant differences were observed in days to heading and days from salt application to heading across three sowing periods. This period was the longest in April-sown plants, the shortest in June-sown plants, and intermediate in May-sown plants across all lines.
Table 9.
Heading dates, days to heading, and days from salt application to heading of Koshihikari, L102, and L74 across different sowing periods, including April, May, and June in 2025.
Relationship Between Heading Date and Plant Dry Weight, Grain Weight, and Percentage of White Head in Three Different Sowing Times (Experiment 3)
In Experiment 3, we compared the relationship between heading dates and plant dry weight, grain weight, and percentage of white heads across three different sowing times (Figure 10). Under control conditions, late-heading lines (L102 and L74) demonstrated higher plant dry weights than Koshihikari when sown in April. There was a tendency for plant dry weights to increase in the late-heading lines (L102 and L74) when sown in May. However, the differences in plant dry weight among Koshihikari, L102, and L74 decreased when their heading dates were altered to mid to late August for sowing in June. Under control conditions, there were no differences in grain weight, except for Koshihikari sown in May. Conversely, Koshihikari, L102, and L74 tended to maintain higher plant dry weight under salt stress due to later heading for those sown in May and June. Overall, these results demonstrated that later heading dates were consistently associated with higher grain weight under salt stress, regardless of the line or sowing time.
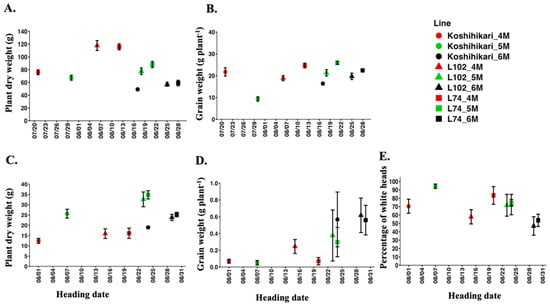
Figure 10.
Relationship between heading date with plant dry weight, grain weight, and percentage of white heads of Koshihikari, L102, and L74 in 2025. The 4M, 5M, and 6M denote sowing in April, May, and June, respectively. The upper panels represent control conditions (A,B) and the lower ones salt stress conditions (C–E). Data points indicate mean ± SE (n = 6).
4. Discussion
Salinity is one of the widespread abiotic stresses limiting grain productivity in rice. Salt tolerance at the reproductive stage is crucial, as grain yield is determined during this stage. Understanding how plants respond to salt stress at heading stages, alongside climatic variables such as air temperature and solar radiation, is essential to ensure sustainable rice production across salt-affected areas. We used five Koshihikari-NILs to investigate whether different heading dates under salt stress affect agronomic parameters and yield-related traits in rice. Our results demonstrated that late-heading NILs were more salt tolerant, with significant improvements in agronomic and yield-related traits. In another study, we also examined whether different heading dates induced by one month of long-day treatment affect salt tolerance in Koshihikari plants. We found that even if the salt treatment period is longer, lower temperatures and reduced solar radiation during the heading period resulted in higher plant dry weight and grain weight in late-heading plants. Further, experiments with different sowing times demonstrated that late-heading and shorter salt treatment periods, such as sowing in June, resulted in higher grain weight. To date, this is the first report to assess the relationship between heading dates for salt tolerance and improvement in agronomic and yield-related traits in rice in late-heading plants. Such strategies complement cultivar-specific priming and reactive oxygen species management for enhanced tolerance [26].
Understanding the pleiotropic effects of heading date genes on major agronomic traits is crucial for developing rice varieties with optimal maturity and productivity for target environments. Several key genes controlling heading date in rice significantly influence yield-related traits [27,28]. Hd16 encodes a casein kinase-I protein. A non-synonymous substitution in Hd16 changes the photoperiod sensitivity in rice. Hd16 is reportedly involved in the photoperiodic flowering pathway through its phosphorylation of Ghd7 [29]. The late-heading NILs (NIL1 and NIL2) maintained significantly higher plant dry weight and panicle weight under salt stress than early-heading NILs (Table 3). The yield-related traits, including 1000-grain weight, percentage of filled grains, and grain weight, were significantly higher in the late-heading NILs under salt stress (Table 4). Previous studies have shown that delayed heading might provide an advantage by enabling plants to sustain vegetative growth and accumulate greater biomass before the onset of reproductive development [30]. In contrast, DTH8 (days to heading) transgenic lines exhibited early flowering, increased yield, and higher salinity tolerance [19]. In Experiments 1 and 3, differences in heading date did not affect grain weight under control conditions (Figure 5 and Figure 10). However, under salt stress conditions, the later the heading date, the higher the grain weight. Therefore, the high grain weight at later heading dates under salt stress conditions was attributed to factors other than heading date-related genes.
Temperature and sunshine hours are two important climatic variables for rice growth and cultivation under changing climatic conditions [31]. Elevated temperatures may drastically affect both rice yield and grain quality [32]. Similarly, increased sunshine hours, along with elevated temperatures, can affect plant growth, shorten the grain-filling period, and reduce assimilate accumulation, thereby compromising yield potential [33,34]. In the present study, rice plants tended to maintain higher grain weight when heading was initiated in late August, when the temperature and solar radiation started to decrease (Figure 3 and Figure 4). Under saline conditions, grain weight exhibited strong positive correlations with panicle number, panicle weight, total weight, days to heading, and percentage of filled grains, whereas no significant correlation was observed with panicle number (Figure 6B). The present study suggested that lower average temperatures and reduced solar radiation during the heading stage might be involved in alleviating salt stress in rice, supporting previous findings that dry matter per unit area before the heading stage is important for mitigating rice yield loss [35].
In the experiments conducted in 2024 and 2025, the percentage of white heads was higher than in the experiment conducted in 2019, and the increase in yield due to the later heading date was related to the lower percentage of white heads. White heads are induced by hot, dry conditions and strong winds [36,37,38], drought conditions [39,40], and salt stress conditions [41]. Excessive water loss from spikelets, resulting from low silicon deposition, could induce white heads under salt stress [41]. In 2024 and 2025, the average temperature in July–August was over 2.1 °C higher, and solar radiation was over 3.3 MJ m−2, both of which were higher than those in 2019. Particularly, solar radiation in July increased by approximately 4.2 MJ m−2 in 2024 and 8.1 MJ m−2 in 2025, relative to that in 2019. Leaf Na+ accumulation is affected by light [42], and in 2024 and 2025, high solar radiation may have increased Na+ accumulation, which could have contributed to the increased percentage of white heads. In 2019, higher grain weight at later heading was more closely associated with the percentage of filled grains than with the percentage of white heads. As increased leaf sodium is associated with decreased photosynthesis [43], high solar radiation during the heading stage may decrease starch accumulation through increased leaf sodium accumulation, thereby reducing the percentage of filled grains. There is also a correlation between flag leaf Na+ content and pollen Na+ content, and between pollen Na+ content and pollen germination [44,45], suggesting that sodium accumulation may be directly associated with reduced fertility. Collectively, our findings demonstrate that heading date is an important trait that may improve salinity tolerance at the reproductive stage in rice.
In another study, we examined the effects of one month of long-day treatment in Koshihikari plants over two consecutive years. Plant dry weight measured immediately after long-day treatment did not differ among light positions under either control or salt stress conditions, indicating no direct effect of light and heat from the light source on plant dry weight. Previous studies have demonstrated that Koshihikari exhibits high sensitivity to photoperiod and temperature during the reproductive transition period [46]. In this study, late-heading Koshihikari plants exhibited greater salt tolerance at the reproductive stage, as reflected by higher percentages of filled grains and grain weight (Table 8). Grain weight increased with delayed heading, with higher grain weight observed in plants heading in late August. In experiments with different sowing times, plants sown in June, which initiated heading in late August, maintained higher grain weight despite shorter salt exposure (Figure 10). Therefore, delayed heading might be associated with improved grain weight under salt stress, possibly by avoiding unfavorable higher temperature and high solar radiation periods.
Despite different experimental settings in three independent experiments, rice plants that started heading in late August showed improved salt tolerance. However, our experiments with Koshihikari-NILs and Koshihikari plants under long-day treatment revealed that they might share a common mechanism for affecting salt tolerance at the reproductive stage due to lower temperature and solar radiation during the heading period. Rice is a facultative short-day plant, which flowers early in short-day and late in long-day conditions. Heading date is a crucial agronomic trait that not only determines the regional adaptation of elite rice cultivars but also serves as a key determinant for yield [16]. Therefore, altering heading dates might greatly benefit the development of salt-tolerant rice varieties under changing climatic conditions. Our findings on salt tolerance at the reproductive stage have strong implications for developing rice production technology and management practices suitable for salt-affected areas. In some areas of South Asian countries like Bangladesh, farmers must take measures to alter transplanting times to ensure secure rice production in salt-affected soils [21]. A recent study concluded that high tolerance to heat-induced spikelet sterility and high yield potential under elevated CO2 are two important characteristics required for rice genotypes to adapt to a global warming environment [47]. To the best of our knowledge, this is the first evidence of alleviating salt stress in rice due to lower temperature and solar radiation during the heading period. These findings might be further deployed in rice breeding programs to develop salt-tolerant varieties for future climatic conditions.
5. Conclusions
Using Koshihikari-NILs with varying heading dates, we confirmed that late-heading NILs significantly produced higher plant dry weight and grain weight under long-term salt stress conditions. In another experiment, the heading dates of the same variety, Koshihikari, were altered through long-day treatment, revealing a tendency for increased grain weight as heading dates were delayed under long-term salt stress. Additionally, experiments with different sowing times indicated that later heading dates were associated with higher grain weight under salt stress, regardless of line or sowing time. This finding informs breeding and sowing strategies that consider heading dates in affected areas. In conclusion, our results demonstrate that heading during periods of low temperature and solar radiation contributes to higher grain weight under long-term salt stress, even if the salt treatment period is long. This phenomenon is proposed as one of the strategies for salt escape. These findings provide valuable insights for using heading dates in the development of salt-tolerant rice varieties, thereby ensuring stable rice production in salt-affected regions.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/crops6020028/s1, Figure S1: Light intensity recorded at various distances from the light source during the long-day treatment in Experiment 2 conducted in 2024 and 2025.
Author Contributions
Conceptualization, Y.H.; methodology, S.A. and Y.H.; software, S.A. and M.M.E.A.; validation, S.A., A.T. and Y.H.; formal analysis, S.A. and S.I.; investigation, S.A., S.I. and S.O.; resources, Y.H.; data curation, S.A., S.I. and S.O.; writing—original draft preparation, S.A.; writing—review and editing, M.M.E.A., A.T. and Y.H.; visualization, S.A. and M.M.E.A.; supervision, Y.H.; project administration, Y.H.; funding acquisition, Y.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by JSPS KAKENHI (grant number JP20H02969 and JP25K01997) and OU-SPRING.
Data Availability Statement
The original contributions presented in this study are included in the article and Supplementary Material. Further inquiries can be directed to the corresponding author.
Acknowledgments
The authors also would like to express their appreciation to all students and staff for their contributions to the execution of this research.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
The following abbreviation is used in this manuscript:
| NIL | near-isogenic line |
References
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef]
- Reddy, I.N.B.L.; Kim, B.K.; Yoon, I.S.; Kim, K.H.; Kwon, T.R. Salt tolerance in rice: Focus on mechanisms and approaches. Rice Sci. 2017, 24, 123–144. [Google Scholar] [CrossRef]
- Sackey, O.K.; Feng, N.; Mohammed, Y.Z.; Dzou, C.F.; Zheng, D.; Zhao, L.; Shen, X. A comprehensive review on rice responses and tolerance to salt stress. Front. Plant Sci. 2025, 16, 1561280. [Google Scholar] [CrossRef]
- Peng, Y.; Zhu, H.; Wang, Y.; Kang, J.; Hu, L.; Li, L.; Zhu, K.; Yan, J.; Bu, X.; Wang, X.; et al. Revisiting the role of light signaling in plant responses to salt stress. Hortic. Res. 2024, 12, uhae262. [Google Scholar] [CrossRef]
- Chanthini, K.M.P.; Pavithra, G.S.; Senthil-Nathan, S.; Malafaia, G. An in-depth review on the mechanistic insights of marine macroalgal compounds in enhancing plant tolerance to stress induced by saline soil conditions. Toxin Rev. 2024, 43, 651–669. [Google Scholar] [CrossRef]
- Bandumula, N. Rice production in Asia: Key to global food security. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2018, 88, 1323–1328. [Google Scholar] [CrossRef]
- Hopmans, J.W.; Qureshi, A.S.; Kisekka, I.; Munns, R.; Grattan, S.R.; Rengasamy, P.; Ben-Gal, A.; Assouline, S.; Javaux, M.; Minhas, P.S.; et al. Critical knowledge gaps and research priorities in global soil salinity. Adv. Agron. 2021, 169, 1–191. [Google Scholar] [CrossRef]
- Singh, R.K.; Kota, S.; Flowers, T.J. Salt tolerance in rice: Seedling and reproductive stage QTL mapping come of age. Theor. Appl. Genet. 2021, 134, 3495–3533. [Google Scholar] [CrossRef] [PubMed]
- Eynerd, A.; Lal, R.; Wiebe, K. Crop response in salt-affected soils. J. Sustain. Agric. 2005, 27, 5–50. [Google Scholar] [CrossRef]
- Xu, Y.; Bu, W.; Xu, Y.; Fei, H.; Zhu, Y.; Ahmad, I.; Nimir, N.E.A.; Zhou, G.; Zhu, G. Effects of salt stress on physiological and agronomic traits of rice genotypes with contrasting salt tolerance. Plants 2024, 13, 1157. [Google Scholar] [CrossRef]
- Tibesigwa, D.G.; Zhuang, W.; Matola, S.H.; Zhao, H.; Li, W.; Yang, L.; Ren, J.; Liu, Q.; Yang, J. Molecular insights into salt stress adaptation in plants. Plant Cell Environ. 2025, 48, 5604–5615. [Google Scholar] [CrossRef]
- Ponce, K.S.; Meng, L.; Guo, L.; Leng, Y.; Ye, G. Advances in sensing, response and regulation mechanism of salt tolerance in rice. Int. J. Mol. Sci. 2021, 22, 2254. [Google Scholar] [CrossRef] [PubMed]
- Takimoto, T.; Masutomi, Y.; Tamura, M.; Nitta, Y.; Tanaka, K. The effect of air temperature and solar radiation on the occurrence of chalky rice grains in rice cultivars “Koshihikari” and “Akitakomachi.”. J. Agric. Meteorol. 2019, 75, 203–210. [Google Scholar] [CrossRef]
- Senguttuvel, P.; Jaldhani, V.; Raju, N.S.; Balakrishnan, D.; Beulah, P.; Bhadana, V.P.; Mangrauthia, S.K.; Neeraja, C.N.; Subrahmanyam, D.; Rao, P.R.; et al. Breeding rice for heat tolerance and climate change scenario; possibilities and way forward. A review. Agron. Soil Sci. 2022, 68, 115–132. [Google Scholar] [CrossRef]
- Cabusora, C.C. Developing climate-resilient crops: Adaptation to abiotic stress-affected areas. Technol. Agron. 2024, 4, e005. [Google Scholar] [CrossRef]
- Wei, H.; Wang, X.; Xu, H.; Wang, L. Molecular basis of heading date control in rice. aBIOTECH 2020, 1, 219–232. [Google Scholar] [CrossRef]
- Zhou, S.; Cai, L.; Wu, H.; Wang, B.; Gu, B.; Cui, S.; Huang, X.; Xu, Z.; Hao, B.; Hou, H.; et al. Fine-tuning rice heading date through multiplex editing of the regulatory regions of key genes by CRISPR-Cas9. Plant Biotechnol. J. 2024, 22, 751–758. [Google Scholar] [CrossRef]
- Radha, B.; Sunitha, N.C.; Sah, R.P.; Md, M.A.; Krishna, G.K.; Umesh, D.K.; Thomas, S.; Anilkumar, C.; Upadhyay, S.; Kumar, A.; et al. Physiological and molecular implications of multiple abiotic stresses on yield and quality of rice. Front. Plant Sci. 2023, 13, 996514. [Google Scholar] [CrossRef]
- Mishra, M.; Rathore, R.S.; Joshi, R.; Pareek, A.; Singla-Pareek, S.L. DTH8 overexpression induces early flowering, boosts yield, and improves stress recovery in rice cv. IR64. Physiol. Plant 2022, 174, e13691. [Google Scholar] [CrossRef] [PubMed]
- Radanielson, A.M.; Gaydon, D.S.; Rahman Khan, M.M.; Chaki, A.K.; Rahman, M.A.; Angeles, O.; Li, T.; Ismail, A. Varietal improvement options for higher rice productivity in salt affected areas using crop modelling. Field Crops Res. 2018, 229, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.; Landis, D.A. Adapting to salinity in coastal rice farming: Integrating farmer perceptions with empirical field evidence. Clim. Action 2025, 4, 90. [Google Scholar] [CrossRef]
- Shavrukov, Y.; Kurishbayev, A.; Jatayev, S.; Shvidchenko, V.; Zotova, L.; Koekemoer, F.; De Groot, S.; Soole, K.; Langridge, P. Early flowering as a drought escape mechanism in plants: How can it aid wheat production? Front. Plant Sci. 2017, 8, 1950. [Google Scholar] [CrossRef] [PubMed]
- Mai, N.S.; Hanh, D.D.; Nakashima, M.; Kumamoto, K.; Thuy, N.T.T.; Kobata, T.; Saitoh, K.; Hirai, Y. Identification and validation of QTLs for yield and yield components under long-term salt stress using IR64 CSSLs in the genetic background of koshihikari and their backcross progenies. Agriculture 2021, 11, 777. [Google Scholar] [CrossRef]
- Nonoue, Y.; Hori, K.; Ono, N.; Shibaya, T.; Ogiso-Tanaka, E.; Mizobuchi, R.; Fukuoka, S.; Yano, M. Detection of heading date QTLs in advanced-backcross populations of an elite indica rice cultivar, IR64. Breed. Sci. 2019, 69, 352–358. [Google Scholar] [CrossRef]
- Kassambara, A. ggcorrplot: Visualization of a Correlation Matrix Using ggplot2. 2022. Available online: https://cran.r-project.org/web/packages/ggcorrplot/readme/README.html (accessed on 5 December 2025).
- Xu, P.; Hao, S.; Wen, X.; Ma, G.; Yang, Q.; Liu, L.; Anis, G.B.; Zhang, Y.; Sun, L.; Shen, X.; et al. A series of novel alleles of Ehd2 modulating heading and salt tolerance in rice. Plants 2025, 14, 297. [Google Scholar] [CrossRef]
- Lee, S.Y.; Jeung, J.U.; Mo, Y. Allelic combinations of Hd1, Hd16, and Ghd7 exhibit pleiotropic effects on agronomic traits in rice. Genes Genomes Genet. 2024, 14, jkad300. [Google Scholar] [CrossRef] [PubMed]
- Fujino, K. Days to heading, controlled by the heading date genes, Hd1 and DTH8, limits rice yield-related traits in Hokkaido, Japan. Breed. Sci. 2020, 70, 277–282. [Google Scholar] [CrossRef]
- Hori, K.; Ogiso-Tanaka, E.; Matsubara, K.; Yamanouchi, U.; Ebana, K.; Yano, M. Hd16, a gene for casein kinase I, is involved in the control of rice flowering time by modulating the day-length response. Plant J. 2013, 76, 36–46. [Google Scholar] [CrossRef]
- Liu, J.; Yi, Q.; Dong, G.; Chen, Y.; Guo, L.; Gao, Z.; Zhu, L.; Ren, D.; Zhang, Q.; Li, Q.; et al. Improving rice quality by regulating the heading dates of rice varieties without yield penalties. Plants 2024, 13, 2221. [Google Scholar] [CrossRef] [PubMed]
- Deng, N.; Ling, X.; Sun, Y.; Zhang, C.; Fahad, S.; Peng, S.; Cui, K.; Nie, L.; Huang, J. Influence of temperature and solar radiation on grain yield and quality in irrigated rice system. Eur. J. Agron. 2015, 64, 37–46. [Google Scholar] [CrossRef]
- Krishnan, P.; Ramakrishnan, B.; Reddy, K.R.; Reddy, V.R. High-temperature effects on rice growth, yield, and grain quality. Adv. Agron. 2011, 111, 87–206. [Google Scholar] [CrossRef]
- Tu, D.; Wu, W.; Xi, M.; Zhou, Y.; Xu, Y.; Chen, J.; Shao, C.; Zhang, Y.; Zhao, Q. Effect of temperature and radiation on indica rice yield and quality in middle rice cropping system. Plants 2022, 11, 2697. [Google Scholar] [CrossRef] [PubMed]
- Oh, D.; Ryu, J.H.; Jeong, H.; Moon, H.D.; Kim, H.; Jo, E.; Kim, B.K.; Choi, S.; Cho, J. Effect of elevated air temperature on the growth and yield of paddy rice. Agronomy 2023, 13, 2887. [Google Scholar] [CrossRef]
- Wang, W.; Cai, C.; Lam, S.K.; Liu, G.; Zhu, J. Elevated CO2 cannot compensate for japonica grain yield losses under increasing air temperature because of the decrease in spikelet density. Eur. J. Agron. 2018, 99, 21–29. [Google Scholar] [CrossRef]
- Asano, Y.; Kusaka, H. Numerical simulation study of the effects of foehn winds on white head incidences in Yamagata Prefecture, Japan. Meteorol. Appl. 2021, 28, e2042. [Google Scholar] [CrossRef]
- Muramatsu, K.; Kamota, F. Damage of paddy rice plant caused by foehn. Boll Hokurku Natl. Agric. Res. Stn. 1981, 23, 19–56, (In Japanese with English Abstract). [Google Scholar]
- Tsuboi, Y. Ecological studies on rice plants with regard to damages caused by wind. J. Artic. Bull. Nat. Inst. Agric. Sci. 1961, 8, 81–156. [Google Scholar]
- O’Toole, J.C.; Hsiao, T.C.; Namuco, O.S. Panicle water relations during water stress. Plant Sci. Lett. 1984, 33, 137–143. [Google Scholar] [CrossRef]
- Ekanayake, I.J.; De Datta, S.K.; Steponkus, P.L. Effect of water deficit stress on diffusive resistance, transpiration, and spikelet desiccation of rice; (Oryza sativa L.). Ann. Bot. 1993, 72, 73–80. [Google Scholar] [CrossRef]
- Makoto, T.; Miho, M.; Daigo, M.; Yoshihiko, H. The involvement of silicon deposition in salinity-induced white head in rice (Oryza sativa L.). Plant. Prod. Sci. 2000, 3, 328–334. [Google Scholar] [CrossRef]
- Mitsuya, S.; Kawasaki, M.; Taniguchi, M.; Miyake, H. Light dependency of salinity-induced chloroplast degradation. Plant. Prod. Sci. 2003, 6, 219–223. [Google Scholar] [CrossRef]
- Yeo, A.R.; Capron, S.J.M.; Flowers, T.J. The Effect of salinity upon photosynthesis in rice (Oryza sativa L.): Gas exchange by individual leaves in relation to their salt content. J. Exp. Bot. 1985, 36, 1240–1248. [Google Scholar] [CrossRef]
- Khatun, S.; Rizzo, C.A.; Flowers, T.J. Genotypic variation in the effect of salinity on fertility in rice. Plant Soil 1995, 173, 239–250. [Google Scholar] [CrossRef]
- Khatun, S.; Flowers, T.J. Effects of salinity on seed set in rice. Plant Cell Env. 1995, 18, 61–67. [Google Scholar] [CrossRef]
- Hori, K.; Saisho, D.; Nagata, K.; Nonoue, Y.; Uehara-Yamaguchi, Y.; Kanatani, A.; Shu, K.; Hirayama, T.; Yonemaru, J.I.; Fukuoka, S.; et al. Genetic elucidation for response of flowering time to ambient temperatures in Asian rice cultivars. Int. J. Mol. Sci. 2021, 22, 1024. [Google Scholar] [CrossRef]
- Horie, T. Global warming and rice production in Asia: Modeling, impact prediction and adaptation. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2019, 95, 211–245. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2026 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license.