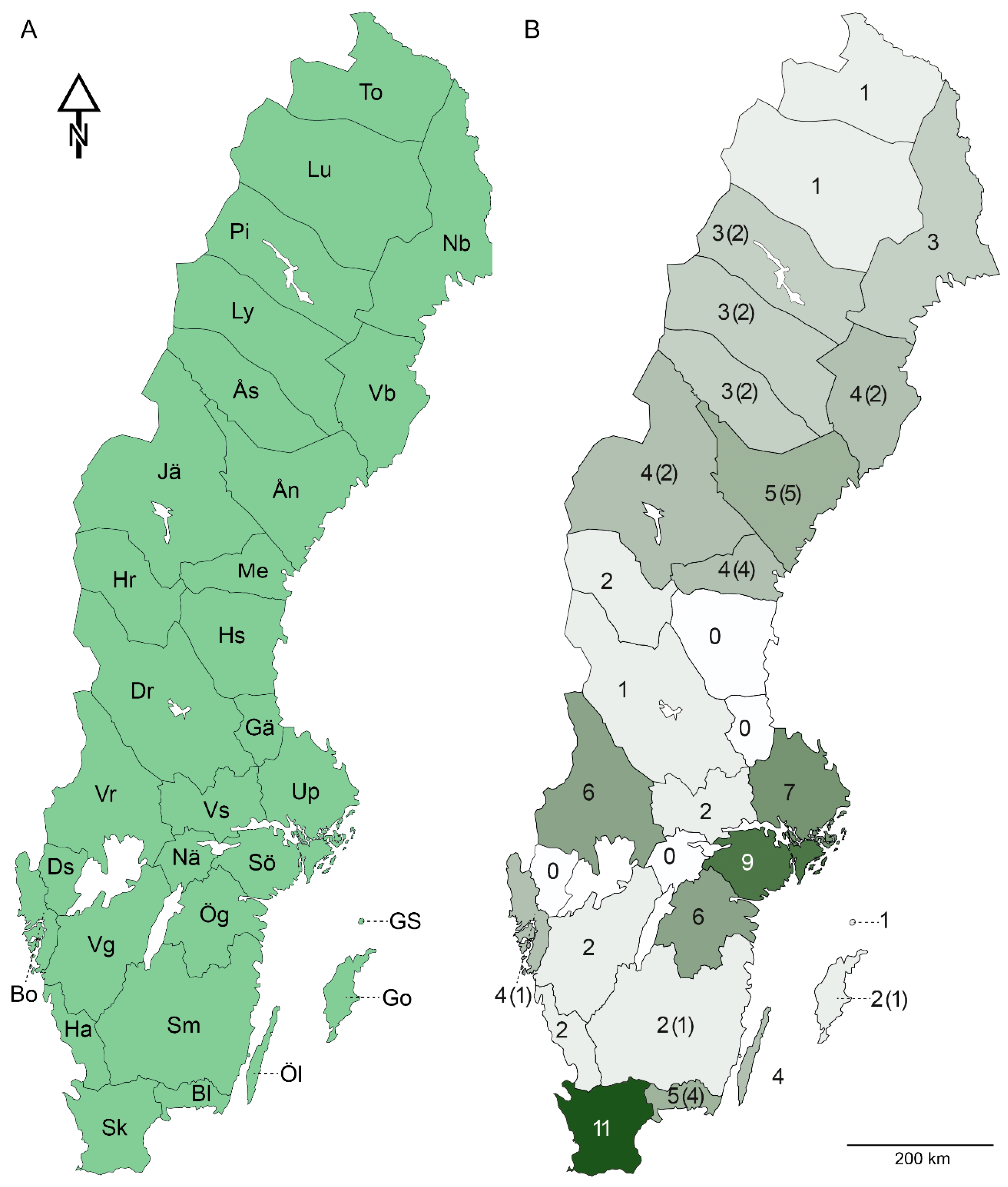
The identification key to the 13 known and 2 potentially introduced species in Sweden is followed by the current taxonomical records of Aeolothripidae and Melanthripidae with short descriptions.
3.2. Taxonomy
| Order Thysanoptera Haliday, 1836 [3] |
| Suborder Terebrantia Haliday, 1836 [3] |
| Family Aeolothripidae Uzel, 1895 [1] |
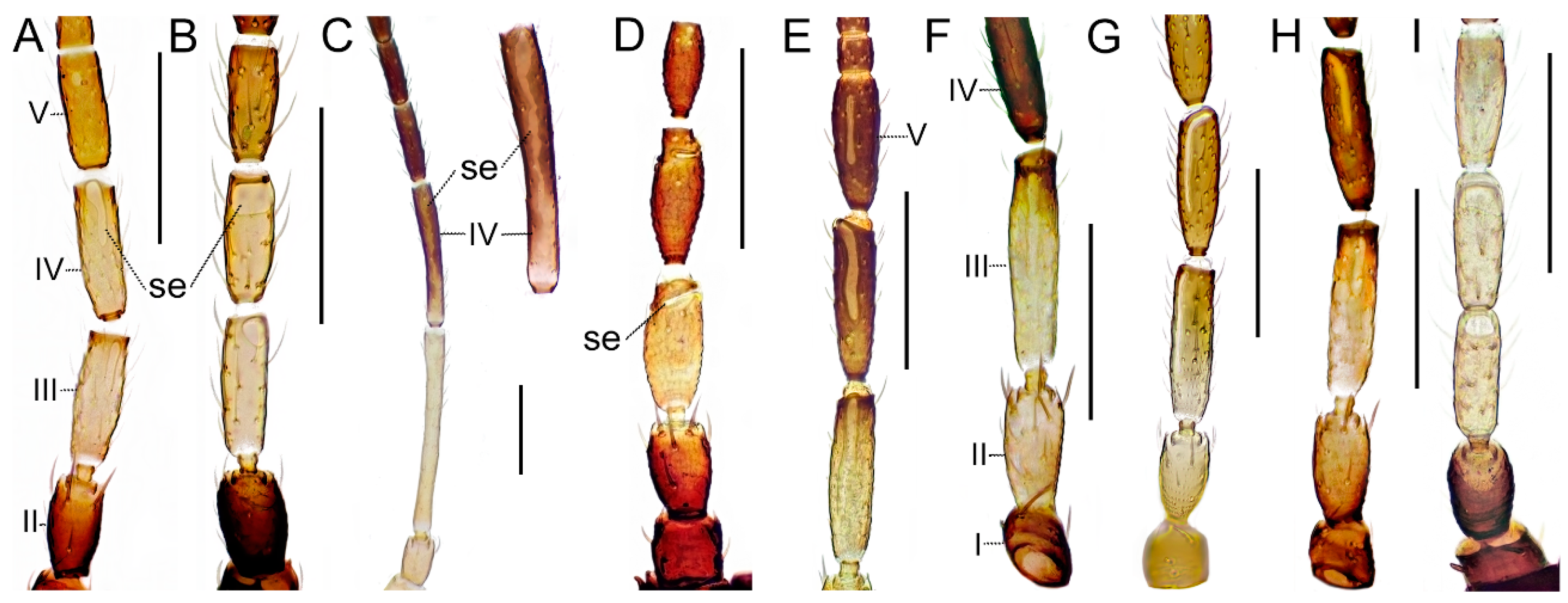
The Aeolothripidae are characterized by having nine antennal segments, but the terminal segments can be partly fused and harder to distinguish. The sensorial organs on the antennae are either elongate and longitudinal, or as regular or irregular blotches or lens-shaped structures (
Figure 3A–C). Macropterous, brachypterous and micropterous conditions exist, also within species. Males are rarely collected and, in some cases, unknown. Wings in many species are banded, but for some species they are evenly shaded or completely pale. The biology is variable, from obligate predators to omnivores to pollen feeding or feeding on plant tissue and fluids.
| Genus Aeolothrips Haliday, 1836 [3] |
| Aeolothrips albicinctus Haliday, 1836 [3] |
| Figure 1C, Figure 6E and Figure 7D |
| Aeolothrips albicinctus Haliday, 1836: 451 [3]. |
SWEDEN; 1♀; Södermanland, Tyresta national park; 2–15 July 2000; B. Viklund, L-O Wikars, H. Ahnlund leg.; Malaise trap; NHRS.; 2♂♂; Södra Värmland, Rämmens socken, Fjällrämmens beach, 5 km NNE of Liljendal; 26 May 1974; U. Qvick leg.; in Calamagrostis; MZLU 00151698, 00158168.; 1♀1♂; Uppland, Stockholm, Experimentalfältet; 10 August 1919; O. Ahlberg leg.; NHRS.; 1♀; Värmland, Gustaf Adolfs socken, Geijersholms dam; 8 June 1975; U. Qvick leg.; on Calamagrostis; MZLU 00158166.; 1♀; Blekinge, Karlskrona kommun, Torhamn; N56.092591, E15.853811; 31 May 2023; E. Wahlberg leg.; on Barbarea vulgaris; voucher ID: BK3T; BOLD ID: SETHY263-23; NHRS.; 1♀; Bohuslän, Tjörns kommun, Kebene; N57.977127, E11.605660; 11 June 2023; E. Wahlberg leg.; in mixed herbaceous vegetation; voucher ID: BD2T; BOLD ID: SETHY203-23; NHRS.; 1♀; Jämtland, Strömsunds kommun, Sävselet; N63.538795, E15.444725; 15 June 2023; E. Wahlberg leg.; mixed flowering plants; voucher ID: BQ1T; BOLD ID: SETHY310-23; NHRS.; 1♀; Lycksele lappmark, Sorsele kommun, Saxnäs; N65.455847, E17.572265; 16 June 2023; E. Wahlberg leg.; mixed flowering plants; voucher ID BP2T; BOLD ID: SETHY303-23; NHRS.; 5♀♀; Medelpad, Timrå kommun, Indalälvens delta; N62.508667, E17.448164; 14 June 2023; E. Wahlberg leg.; in mixed herbaceous vegetation; voucher IDs: BD3T, BD4T, BD9T, BE6T, BE9T; BOLD IDs: SETHY204-23, SETHY205-23, SETHY210-23, SETHY216-23, SETHY219-23; NHRS.; 1♀; Småland, Västerviks kommun, Tinderednäs; N57.977431, E16.483873; 1 June 2023; E. Wahlberg leg.; on Ranunculus; voucher ID: BL1T; BOLD ID: SETHY270-23; NHRS.; 1♀; Södermanland, Flens kommun, Malmköpings nature reserve; N59.1458, E16.7205; 7 June 2023; E. Wahlberg leg.; voucher ID BI1T; BOLD ID: SETHY245-23; NHRS.; 1♀; Södermanland, Gnesta kommun, Gåsinge; N59.0855, E17.2140; 14 May 2023; E. Wahlberg leg.; on Prunus spinosa; voucher ID: AZ2T; BOLD ID: SETHY167-23; NHRS.; 3♀♀; Västerbotten, Skellefteå kommun, Nilsliden; N64.944032, E20.367689; 18 June 2023; E. Wahlberg leg.; in mixed herbaceous vegetation; voucher IDs: BO1T, BOT2T, BOT3T; BOLD IDs: SETHY294-23, SETHY295-23, SETHY296-23; NHRS.; 2♀♀; Västerbotten, Skellefteå kommun, Yttervik; N64.680468, E21.095959; 18 June 2023; E. Wahlberg leg.; on Silene dioica; voucher IDs: BN5T, BN6T; BOLD IDs: SETHY289-23, SETHY290-23; NHRS.; 7♀♀; Ångermanland, Kramfors kommun, Västhammar; N63.030171, E17.773730; 19 June 2023; E. Wahlberg leg.; in mixed herbaceous vegetation; voucher IDs: AZ5T, AZ6T, AZ7T, AZ8T, BA6T, BA7T, BA8T; BOLD IDs: SETHY170-23, SETHY171-23, SETHY172-23, SETHY173-23, SETHY180-23, SETHY181-23, SETHY182-23; NHRS.; 1♀; Åsele lappmark, Dorotea kommun, Häggås; N64.402187, E16.434090; 16 June 2023; E. Wahlberg leg.; mixed flowering plants; voucher ID: BR7T; BOLD ID: SETHY322-23; NHRS.
Body (excl. extremities) bicolored (
Figure 6E). Antennal segments I–III yellow; segment IV brown. Sensory organs on antennal segments III and IV linear and directed longitudinally. Head mainly dark. Thorax mainly dark. Anterior and posterior margins of pronotum without long setae. Often micropterous (
Figure 6E), if wings present then forewings with two dark transverse bands; apical vein of forewing as pale as wing membrane. Abdominal segments II and III yellow, remaining segments brown (
Figure 6E). Female tergite I transversely striated (
Figure 7D). Male abdominal segment IX without lateral stout setae.
Found on various herbaceous plants and presumed to be predatory on mites [
25] and has an ant-like appearance and behavior. Holarctic distribution. In Fennoscandia, in Sweden, Norway, Denmark, and Finland; in Sweden, previously recorded in Skåne, Öland, Södermanland, Uppland, Värmland, and Lappland (not specified) [
16]. Also reported here from Blekinge, Småland, Bohuslän, Medelpad, Jämtland, Ångermanland, and Västerbotten.
Figure 4.
Head in dorsal view. (A) Aeolothrips ericae, ♂. (B) A. versicolor, ♀. (C) Rhipidothrips brunneus, ♀. (D) R. niveipennis, ♀. Scale bar: 100 µm.
Figure 4.
Head in dorsal view. (A) Aeolothrips ericae, ♂. (B) A. versicolor, ♀. (C) Rhipidothrips brunneus, ♀. (D) R. niveipennis, ♀. Scale bar: 100 µm.
Figure 5.
Forewing. (A) Aeolothrips propinquus, ♀. (B) A. vittatus, ♀. (C) A. melaleucus, ♀. (D) A. astutus, ♀. Scale bar: 100 µm.
Figure 5.
Forewing. (A) Aeolothrips propinquus, ♀. (B) A. vittatus, ♀. (C) A. melaleucus, ♀. (D) A. astutus, ♀. Scale bar: 100 µm.
| Aeolothrips astutus Priesner, 1926 [32] |
| Figure 5D. |
| Aeolothrips astutus Priesner, 1926: 109 [32]. |
SWEDEN; 2♀♀; Skåne, Maglehem, Spröjtagränd 6; N55.767558, E14.149546; 20 July 2021; C.-A. Gertsson leg.; on Anchusa officinalis; MZLU 00148155, 001583875.; 1♀; Skåne, Saxtorp, Badsjön; 2 July 2021; C.-A. Gertsson leg.; on Anchusa officinalis; MZLU 00158377.
Body (excl. extremities) wholly brown. Antennal segment I brown; II mainly brown, at most yellow apically; III bicolored, apical half brown; IV brown. Sensory organs on antennal segments III and IV linear and directed longitudinally. Anterior and posterior margins of pronotum without long setae. Forewings present with two dark transverse bands; apical ring vein as pale as wing membrane (
Figure 5D).
Found on
Anchusa officinalis [
18]. Palearctic distribution. In Fennoscandia, in Sweden and Denmark. In Sweden, found in Södermanland and Skåne [
18].
SWEDEN; 1♂; Värmland, Gustaf Adolfs s:n, Geijersholms herrråd; 10 June 1975; U. Qvick leg.; on flowering Orobus tuberosus; MZLU 00158187.; 1♀; Värmland, Gustaf Adolfs s:n, 1 km SE of Mossjön; 18 June 1975; U. Qvick leg.; on flowering Cardaminopsis arenosa; MZLU 00158202.; 2♀♀; Södermanland, Tyresta national park; 5 June–15 July 2000; B. Viklund, L.-O. Wikars, H. Ahnlund leg.; Malaise trap; NHRS.; 1♀1♂; Södermanland, Gnesta kommun, Bryggeriholmen; N59.048168, E17.318564; 23 June 2022; E. Wahlberg leg.; voucher IDs: AV6T, AV7T; BOLD IDs: SETHY140-22, SETHY141-22; NHRS.; 1♀; Jämtland, Östersunds kommun, Södergård; N63.095297, E14.820085; 30 June–13 July 2021; Swedish Insect Inventory Program (SIIP) leg.; mowing meadow next to coniferous forest; Malaise trap; locality ID: 54; collection event ID: M54-003; sample ID: 39_G6; Station Linné.; 1♀; Ångermanland, Härnösand kommun, Lockeby 375; N62.6402228, E17.770841; 15 June–1 July 2021; Swedish Insect Inventory Program (SIIP) leg.; pasture in deciduous forest; Malaise trap; locality ID: 56; collection event ID: M56-001; sample ID: 72_H9; Station Linné.; 1♀; Blekinge, Karlskrona kommun, Gamla reningsverket; N56.189540, E15.622587; 31 May 2023; E. Wahlberg leg.; on Anchusa officinalis; voucher ID: AZ4T; BOLD ID: SETHY169-23; NHRS.; 6♀♀; Blekinge, Karlskrona kommun, Torhamn; N56.092591, E15.853811; 31 May 2023; E. Wahlberg leg.; on Barbarea vulgaris; voucher IDs: BK4T, BK5T, BK6T, BK7T, BK8T, BK9T; BOLD IDs: SETHY264-23, SETHY265-23, SETHY266-23, SETHY267-23, SETHY268-23, SETHY269-23; NHRS.; 3♀♀1♂; Jämtland, Ragunda kommun, Boberg; N63.359057, E15.717947; 15 June 2023; E. Wahlberg leg.; on Astragalus alpinus; voucher IDs: BR1T, BR4T, BR5T, BQ9T; BOLD IDs: SETHY317-23, SETHY319-23, SETHY320-23, SETHY316-23; NHRS.; 3♀♀; Jämtland, Strömsunds kommun, Sävselet; N63.538795, E15.444725; 15 June 2023; E. Wahlberg leg.; mixed flowering plants; voucher IDs: BQ3T, BQ4T, BQ5T; BOLD IDs: SETHY311-23, SETHY312-23, SETHY313-23; NHRS.; 2♀♀1♂; Lycksele lappmark, Sorsele kommun, Saxnäs; N65.455847, E17.572265; 16 June 2023; E. Wahlberg leg.; mixed flowering plants; voucher IDs: BO8T, BP1T, BO9T; BOLD IDs: SETHY300-23, SETHY302-23, SETHY301-23; NHRS.; 6♀♀; Medelpad, Timrå kommun, Indalälvens delta; N62.508667, E17.448164; 14 June 2023; E. Wahlberg leg.; in mixed herbaceous vegetation; voucher IDs: BD5T, BD6T, BD7T, BE4T, BE5T, BF1T; BOLD IDs: SETHY206-23, SETHY207-23, SETHTY208-23, SETHY214-23. SETHY215-23, SETHY220-23; NHRS.; 1♀; Medelpad, Timrå kommun, Svedje; N62.519346, E17.288289; 14 June 2023; E. Wahlberg leg.; in mixed herbaceous vegetation; voucher ID: BN3T; BOLD ID: SETHY287-23; NHRS.; 2♀♀; Pite lappmark, Arjeplogs kommun, Arjeplog; N66.040783, E17.868661; 18 June 2023; E. Wahlberg leg.; on Astragalus alpinus; voucher IDs: BP7T, BP8T; BOLD IDs: SETHY307-23, SETHY308-23; NHRS.; 3♀♀1♂; Västerbotten, Skellefteå kommun, Yttervik; N64.680468, E21.095959; 18 June 2023; E. Wahlberg leg.; on Silene dioica; voucher IDs: BN7T, BN8T, BN9T, BN4T; BOLD IDs: SETHY291-23, SETHY292-23, SETHY293-23, SETHY288-23; NHRS.; 2♀♀; Ångermanland, Kramfors kommun, Västhammar; N63.030171, E17.773730; 19 June 2023; E. Wahlberg leg.; in mixed herbaceous vegetation; voucher IDs: AZ9T, BA2T; BOLD IDs: SETHY174-23, SETHY176-23; NHRS.; 14♀♀4♂♂; Ångermanland, Sollefteå kommun, Märrgård; N63.717364, E16.823417; 15 June 2023; E. Wahlberg leg.; in mixed herbaceous vegetation; voucher IDs: BB2T, BB3T, BB5T, BB7T, BB9T, BC1T, BC2T, BC3T, BC4T, BC5T, BC6T, BC7T, BC8T, BD1T, BB4T, BB6T, BB8T, BC9T; BOLD IDs: SETHY185-23, SETHY186-23, SETHY188-23. SETHY190-23, SETHY192-23, SETHY193-23, SETHY194-23, SETHY195-23, SETHY196-23, SETHY197-23, SETHY198-23, SETHY199-23, SETHY200-23, SETHY202-23, SETHY187-23, SETHY189-23, SETHY191-23, SETHY201-23; NHRS.; 15♀♀4♂♂; Åsele lappmark, Dorotea kommun, Västra Ormsjö; N64.467878, E15.951629; 16 June 2023; E. Wahlberg leg.; on Astragalus alpinus; voucher IDs: BF3T, BF5T, BF6T, BF7T, BF9T, BG1T, BG2T, BG3T, BG4T, BG7T, BG8T, BG9T, BH1T, BH3T, BH4T, BF8T, BG5T, BH2T, BH6T; BOLD IDs: SETHY222-23, SETHY224-23, SETHY225-23, SETHY226-23, SETHY228-23, SETHY229-23, SETHY230-23, SETHY231-23, SETHY232-23, SETHY235-23, SETHY236-23, SETHY237-23, SETHY238-23, SETHY240-23, SETHY241-23, SETHY227-23, SETHY233-23, SETHY239-23, SETHY243-23; NHRS.
Body (excl. extremities) mostly wholly brown (
Figure 6C). Antennal segment I yellow or pale light brown; II mainly yellow; III bicolored, apically darker; IV brown (
Figure 3F). Sensory organs on antennal segments III and IV linear and directed longitudinally (
Figure 3F). Anterior and posterior margins of pronotum without long setae. Forewings present with two dark transverse bands; apical ring vein of forewing as pale as wing membrane (
Figure 6C). Abdominal segments all brown; X often slightly lighter; male abdominal segment IX with sickle-shaped lateral stout setae (
Figure 6C).
Found in various herbaceous plants, presumed to be a facultative predator. Palaearctic distribution. In Fennoscandia in Sweden, Norway, and Finland. In Sweden broadly distributed throughout, with previous records from Skåne, Halland, Södermanland, Östergötland, Västergötland, Bohuslän, Västmanland, Dalarna, Värmland, Härjedalen and Jämtland. Also reported here from Blekinge, Medelpad, Ångermanland, Västerbotten, Åsele lappmark, Lycksele lappmark, and Pite lappmark.
Figure 6.
Habitus in dorsal view. (A) A. intermedius, ♀. (B) A. fasciatus, ♀. (C) A. ericae, ♂. (D) A. gloriosus, ♀. (E) A. albicinctus, ♀. (F) Rhipidothrips niveipennis, ♀, syntype. Scale bar: 1 mm.
Figure 6.
Habitus in dorsal view. (A) A. intermedius, ♀. (B) A. fasciatus, ♀. (C) A. ericae, ♂. (D) A. gloriosus, ♀. (E) A. albicinctus, ♀. (F) Rhipidothrips niveipennis, ♀, syntype. Scale bar: 1 mm.
| Aeolothrips fasciatus (Linnaeus, 1758) [33] |
| Figure 6B and Figure 7B. |
| Thrips fasciata Linnaeus, 1758: 457 [33]. |
SWEDEN; 1♂1♀; Uppland, Stockholm, Experimentalfältet; 10 June 1919; O. Ahlberg leg.; in grass; MZLU 00158215.; 1♂; Skåne, Lund; 18 July 1957; L. Cederholm leg.; on white filter paper; MZLU.; 1♂; Skåne, Hasslebro; N55.940809, E13.395151; 7–14 July 2020; Y. Cough et al. Leg.; blue pan trap; MZLU 00117239.; 1♂; Skåne, Rörum; N55.622867, E14.245726; 27 July–3 August 2020; white pan trap; Y. Cough et al. leg.; MZLU 00117237.; 1♀; Skåne, Kågerröd; N56.006561, E13.187555; 7–14 July 2020; Blue pant rap; Y. Cough et al. leg.; MZLU 00117238.; 1♀; Norrbotten, Övertorneå kommun, Hanhinvittikko; N66.451386, E23.483197; 7 June–22 June 2021; Swedish Insect Inventory Program (SIIP) leg.; claimed hay meadow; Malaise trap; locality ID: 50; collection event ID: M50-013; sample ID: 7_D10; Station Linné.; 1♀; Östergötland, Linköpings kommun, Västerlösa; N58.42107, E15.33843; 28 July–4 August 2019; Insect Biome Atlas (IBA) leg.; urban; Malaise trap; locality ID: TDUYXP; collection event ID: S7536S; sample ID: 231_C3; Station Linné.; 8♀♀1♂; Blekinge, Karlskrona kommun, Augerum; N56.216929, E15.676876; 1 June 2023; E. Wahlberg leg.; on Anchusa officinalis; voucher IDs: BL9T, BM1T, BM2T, BM3T, BM4T, BM6T, BM7T, BM8T, BK2T; BOLD IDs: SETHY277-23, SETHY278-23, STHY279-23, SETHY280-23, SETHY281-23, SETHY282-23, SETHY283-23, SETHY284-23, SETHY262-23; NHRS.; 1♀; Blekinge, Karlskrona kommun, Gamla reningsverket; N56.189540, E15.622587; 31 May 2023; E. Wahlberg leg.; on Anchusa officinalis; voucher ID: AZ3T; BOLD ID: SETHY168-23; NHRS.; 3♀♀; Blekinge, Karlskrona kommun, Sandhamn; N56.090980, E15.856533; 31 May 2023; E. Wahlberg leg.; in mixed herbaceous vegetation; voucher IDs: BL3T, BL6T, BL8T; BOLD IDs: SETHY272-23, SETHY275-23, SETHY276-23; NHRS.; 2♀♀1♂; Jämtland, Ragunda kommun, Boberg; N63.359057, E15.717947; 15 June 2023; E. Wahlberg leg.; on Astragalus alpinus; voucher IDs: BQ6T, BR3T, BQ8T; BOLD IDs: SETHY314-23, SETHY318-23, SETHY315-23; NHRS.; 1♀; Lycksele lappmark, Sorsele kommun, Saxnäs; N65.455847, E17.572265; 16 June 2023; E. Wahlberg leg.; mixed flowering plants; voucher ID: BO7T; BOLD ID: SETHY299-23; NHRS.; Female 3♀♀3♂♂; Medelpad, Timrå kommun, Indalälvens delta; N62.508667, E17.448164; 14 June 2023; E. Wahlberg leg.; in mixed herbaceous vegetation; voucher IDs: BE1T, BE7T, BE8T, BD8T, BE2T, BE3T; BOLD IDs: SETHY211-23, SETHY217-23, SETHY218-23, SETHY209-23, SETHY212-23, SETHY213-23; NHRS.; 1♀; Medelpad, Timrå kommun, Svedje; N62.519346, E17.288289; 14 June 2023; E. Wahlberg leg.; in mixed herbaceous vegetation; voucher ID: BM9T; BOLD ID: SETHY285-23; NHRS.; 2♀♀2♂♂; Pite lappmark, Arjeplogs kommun, Arjeplog; N66.040783, E17.868661; 18 June 2023; E. Wahlberg leg.; on Astragalus alpinus; voucher IDs: BP6T, BP9T, BP3T, BP5T; BOLD IDs: SETHY306-23, SETHY309-23, SETHY304-23, SETHY305-23; NHRS.; 1♀; Södermanland, Flens kommun, Sparreholms ekhagar; N59.086089, E16.8305657; 7 June 2023; E. Wahlberg leg.; on Ranunculus; voucher ID: BI7T; BOLD ID: SETHY251-23; NHRS.; 5♀♀; Ångermanland, Kramfors kommun, Västhammar; N63.030171, E17.773730; 19 June 2023; E. Wahlberg leg.; in mixed herbaceous vegetation; voucher IDs: BA1T, BA4T, BA5T, BA9T, BB1T; BOLD IDs: SETHY175-23, SETHY178-23, SETHY179-23, SETHY183-23, SETHY184-23; NHRS.; 1♀; Ångermanland, Nordmalings kommun, Torrböle; N63.700503, E19.604039; 19 June 2023; E. Wahlberg leg.; in mixed herbaceous vegetation; voucher ID: BI6T; BOLD ID: SETHY250-23; NHRS.; 2♀♀; Åsele lappmark, Dorotea kommun, Västra Ormsjö; N64.467878, E15.951629; 16 June 2023; E. Wahlberg leg.; on Astragalus alpinus; voucher IDs: BF4T, BH5T; BOLD IDs: SETHY223-23, SETHY242-23; NHRS.
Body (excl. extremities) wholly dark (
Figure 6B). Antennal segment I brown; II mainly brown, at most yellow apically; III yellow with apical brown band; IV brown. Sensory organs on antennal segments III and IV linear and directed longitudinally. Anterior and posterior margins of pronotum without long setae. Forewings present with two dark transverse bands; apical ring vein of forewing as pale as wing membrane (
Figure 6B). Male base of bifurcate claspers on tergite IX with setae longer than claspers (
Figure 7B).
Found in various herbaceous plants, especially flowers, and presumably a facultative predator [
34]. Difficult to separate from
A. intermedius Bagnall, 1934, and the distribution is not clearly understood. Records from Palaearctic as well as New Zealand and Tasmania [
34]. In all Fennoscandian countries, and widely distributed in Sweden, with previous records from Skåne, Halland, Öland, Gotland, Gotska Sandön, Östergötland, Västergötland, Bohuslän, Södermanland, Uppland, Västmanland, Härjedalen, and Norrbotten. Also reported here from Blekinge, Jämtland, Medelpad, Ångermanland, Åsele lappmark, Lycksele lappmark, and Pite lappmark.
Figure 7.
Tergites IX–X, dorsal view. (A) A. intermedius, ♂. (B) A. fasciatus, ♂. Tergite I, dorsal view. (C) A. ericae, ♀. (D) A. albicinctus, ♀. Pronotum, dorsal view. (E) Rhipidothrips brunneus, ♀. (F) R. niveipennis, ♀. Scale bar: 100 µm.
Figure 7.
Tergites IX–X, dorsal view. (A) A. intermedius, ♂. (B) A. fasciatus, ♂. Tergite I, dorsal view. (C) A. ericae, ♀. (D) A. albicinctus, ♀. Pronotum, dorsal view. (E) Rhipidothrips brunneus, ♀. (F) R. niveipennis, ♀. Scale bar: 100 µm.
| Aeolothrips gloriosus Bagnall, 1914 [35] |
| Figure 6D. |
| Aeolothrips gloriosus Bagnall, 1914: 375 [35]. |
SWEDEN; 2♀♀; Skåne, Sandhammaren; 17 July 1997; in Calluna; L. Vasiliu-Oromulu det.; MZLU 00109577.
Body (excl. extremities) mostly yellow, darker medially and segments VII–X, sometimes also V–VI, brown (
Figure 6D). Antennal segment I yellow or pale light brown; II mainly yellow; III bicolored, apical half brown. Sensory organs on antennal segments III and IV linear and directed longitudinally. Anterior and posterior margin of pronotum without long setae. Forewings present with two dark transverse bands; apical ring vein as pale as wing membrane (
Figure 6D). Abdominal segments IX without stout lateral setae.
Found on a variety of different flowering trees [
9], but in Sweden on
Calluna sp., and is probably a facultative predator. Remarkable in comparison with other species in the region with its brightly colored habitus. Palaearctic distribution, in south to northern Africa. In Fennoscandia, only known from a single locality in Sweden, Skåne [
16].
SWEDEN; 4♀♀; Södermanland, Tyresta national park; 15 July–26 August 2000; Malaise trap; B. Viklund, L.-O. Wikars, H. Ahnlund leg.; NHRS.; 1♂; Värmland, Gustaf Adolfs s:n, 1 km southwest of Mossjön; 18 June 1975; on flowering Cardaminopsis arenosa; U. Qvick leg.; MZLU 00109588.; 1♀; Värmland, Gustaf Adolfs s:n, 1 km southwest of Mossjön; 18 June 1975; on flowering Rumex acetosella; U. Qvick leg.; MZLU 00109590.; 1♀; Värmland, Gustaf Adolfs s:n, 1 km southwest of Mossjön; 18 June 1975; on various spp of gramines; U. Qvick leg.; MZLU 00109586.; 1♀; Ångermanland, Nordmalings kommun, Mullsjö; N63.699004, E19.741838; 19 June 2023; E. Wahlberg leg.; on Crepis and Geranium; voucher ID: BF2T; BOLD ID: SETHY221-23; NHRS.
Body (excl. extremities) wholly dark (
Figure 6A). Antennal segment I brown; II mainly brown, at most yellow apically; III yellow with apical brown band; IV brown (
Figure 3H). Sensory organs on antennal segments III and IV linear and directed longitudinally (
Figure 3H). Anterior and posterior margins of pronotum without long setae. Forewings present with two dark transverse bands; apical ring vein of forewing as pale as wing membrane (
Figure 6A). Male base of bifurcate claspers on tergite IX with setae shorter than claspers (
Figure 7A).
As
A. fasciatus is found in various herbaceous plants, especially flowers, living as a facultative predator. The difficulties in separating
A. intermedius from
A. fasciatus makes a complete view of the distribution difficult, but it is presumably mainly Palaearctic [
25]. In Fennoscandia it is present in Sweden, Norway, Denmark, and Finland. In Sweden it is limited to the southern half in contrast to the widely spread
A. fasciatus, recorded previously in Skåne, Småland, Östergötland, Södermanland, and Värmland. The new record herein is from Ångermanland.
| Aeolothrips manteli Titschack, 1962 [37] |
| Figure 3E. |
| Aeolothrips manteli Titschack, 1962: 133 [37]. |
NETHERLANDS; 1♀; Bloemendaal; 8 June 1962; on Anchusa officinalis; W. P. Mantel leg.; coll. H. Priesner; SMF.
Body (excl. extremities) wholly dark. Antennal segment I brown; II mainly brown, at most yellow apically; III yellow at base, darker apically, IV brown (
Figure 3E). Sensory organs on antennal segments III and IV linear and directed longitudinally, sensory organ on segment V also elongate and about half as long as segment (
Figure 3E). Anterior and posterior margin of pronotum without long setae. Forewings present with two dark transverse bands; apical ring vein darker than wing membrane.
Found on
Anchusa officinalis, originally described and recorded from the Netherlands, but was also recorded in Norway in 2001 [
38]. Kobro discussed the possibility of human-mediated dispersal [
38], and Gertsson raised the possibility of occurrence in Sweden due to the geographical proximity [
18]. During this study several localities with
Anchusa officinalis were sampled without finding
A. manteli, but it is included herein as its introduction to Sweden is plausible.
| Aeolothrips melaleucus Haliday, 1852 [39] |
| Figure 5C. |
| Coleothrips melaleuca Haliday, 1852: 1117 [39]. |
SWEDEN; 1♀; Öland, Mörbylånga kommun, Gamla Skogsby (Kalkstad), diversitetesängen; N56.616700, E16.507617; 25 June–13 July 2003; meadow with bushes; Malaise trap; J. Liljeblad leg.; NHRS.; 1♀; Värmland, Kristinehamn, 200 m west of Västra Klintvägen; 17 August 1974; U. Qvick leg.; MZLU 00109594.; 1♀; Skåne, Svedala kommun, Hyby; N55.56882, E13.230962; 27 July–3 August 2019; Insect Biome Atlas (IBA) leg.; grassland; Malaise trap; locality ID: TBPT1B; collection event ID: SXUEFH; sample ID: 247_F12; Station Linné.; 1♀; Blekinge, Karlskrona kommun, Sandhamn; N56.090980, E15.856533; 31 May 2023; E. Wahlberg leg.; in mixed herbaceous vegetation; voucher ID: BA3T; NHRS.; 1♀; Ångermanland, Kramfors kommun, Västhammar; N63.030171, E17.773730; 19 June 2023; E. Wahlberg leg.; in mixed herbaceous vegetation; voucher ID: BA3T; BOLD ID: SETHY177-23; NHRS.
Body (excl. extremities) wholly dark. Antennal segment I brown; II mainly brown, at most yellow apically; III mainly yellow; IV yellow at base, darker apically. Sensory organs on antennal segments III and IV linear and directed longitudinally. Anterior and posterior margin of pronotum without long setae. Forewings present with two dark transverse bands, middle area between bands also dark posteriorly; apical ring vein as pale as wing membrane (
Figure 5C). Male abdominal segment IX with lateral stout and sickle-shaped setae.
Found in various deciduous trees, mainly on leaves and flowers as a predator on e.g., mites [
9,
25]. Very widespread Holarctic distribution [
25,
40]. In Fennoscandia reported in Sweden, Norway, and Finland. In Sweden previously recorded in Skåne, Öland, Östergötland, Södermanland, Uppland, and Värmland, and herein also recorded in Blekinge and Ångermanland.
| Aeolothrips propinquus Bagnall, 1924 [41] |
| Figure 3G and Figure 5A. |
| Aeolothrips propinquus Bagnall, 1924: 269 [41]. |
SWEDEN; 7♀♀; Öland, Neptuni åkrar; 14 June 2022; on Echium vulgare; C.-A. Gertsson leg.; MZLU 00158394.
Body (excl. extremities) wholly dark. Antennal segment I brown; II mainly brown, at most yellow apically; III bicolored, apical half brown; IV brown (
Figure 3G). Sensory organs on antennal segments III and IV linear and directed longitudinally, extending basal third and basal quarter of segment, respectively (
Figure 3G). Anterior and posterior margin of pronotum without long setae. Forewings present with two dark transverse bands; apical ring vein brown (
Figure 5A). Male abdominal segment IX without lateral stout setae.
Found in flowers of Boraginaceae and Scrophulariaceae [
9,
41], and has large Palaearctic distribution extending in south to northern Africa [
9,
25]. In Fennoscandia limited to Sweden and Denmark, and in Sweden, recorded in Skåne, Södermanland, Jämtland, Öland, and Bohuslän [
16,
42].
| Aeolothrips versicolor Uzel, 1895 [1] |
| Figure 1A and Figure 4A. |
| Aeolothrips versicolor Uzel, 1895: 69 [1]. |
AUSTRIA∙ 1♀; Oberösterreich, Linz; 30 May 1919; on Quercus robur; NHRS. SWEDEN; 1♀; Öland, Jordtorpsåsen; 2 July 2014; C. Hansson leg.; MZLU 00109491.; 1♀; Skåne; Kullaberg; 10 June 1954; on oak; L. Cederholm leg.; MZLU 00109500.; 1♀; Gotland, Slite, Filehajdar; N57.706733, E18.712835; 28 June 2023; R. Vicente leg.; rich fen; voucher ID: BR8T; BOLD ID: SETHY323-23; NHRS.; 3♀♀; Södermanland, Flens kommun, Sparreholms ekhagar; N59.086089, E16.8305657; 7 June 2023; E. Wahlberg leg.; on Tilia cordata; voucher IDs: BI3T, BI4T, BI5T; BOLD IDs: SETHY247-23, SETHY248-23, SETHY249-23; NHRS.
Body (excl. extremities) wholly dark. Antennal segment I brown; II, III and basis of IV mainly yellow. Sensory organs on antennal segments III and IV linear and directed longitudinally Anterior part of head prolonged in front of eyes (
Figure 4A). Anterior and posterior margin of pronotum without long setae. Forewings present; forewings with one dark transverse band that is very broad and covering a large part of the middle portion of wing, and seldom has two dark transverse bands in Sweden; apical ring vein as pale as wing membrane. Male abdominal segment IX without lateral stout setae.
This species occurs in two variants, one with two dark bands on the forewings and one with one broad dark band covering almost the complete wing. The Swedish specimens often belong to the first variant, which is also the continental character [
25]. Often collected from
Pinus, probably a predator on small arthropods, and with a Holarctic distribution [
9]. Also widespread in Fennoscandia in Sweden, Norway, Denmark, and Finland as well as within Sweden, with previous records from Skåne, Östergötland, Södermanland, Uppland, Värmland, Västerbotten, and Norrbotten. Here, a new record from Gotland is presented.
| Aeolothrips vittatus Haliday, 1836 [3] |
| Figure 3A and Figure 5B. |
| Aeolothrips (Coleothrips) vittatus Haliday, 1836: 451 [3]. |
SWEDEN; 1♀; Skåne, Kullaberg; 10 June 1954; on oak; L. Cederholm leg.; MZLU 00109500.; 1♀; Värmland, Ekshärads s:n, 500 E of Bergängstjärnen; 3 June 1974; beaten from young Pinus sylvestris; U. Qvick leg.; MZLU 00109501.; 1♀; Värmland, Gustaf Adolfs socken, Geijersholms skola; 2 June 1974; beaten from branches of young Pica abies; U. Qvick leg.; MZLU 00109502.; 1♀; Medelpad, Sundsvalls kommun, Björköviken, Njurunda; N62.2458168, E17.5283155; 21 June–7 July 2021; Swedish Insect Inventory Program (SIIP) leg.; sandy mixed forest; Malaise trap; locality ID: 55; collection event ID: M55-001; sample ID: 104_C6; Station Linné.; 1♀; Norrbotten, Jokkmokks kommun, Kåskats; N66.47580, E20.26901; 8 June–16 June 2019; Insect Biome Atlas (IBA) leg.; wetland; Malaise trap; locality ID: TDGVY8; collection event ID: SVLS5M; sample ID: 223_D4; Station Linné.
Body (excl. extremities) wholly dark. Antennal segment I brown; II mainly brown, at most yellow apically; III–IV mostly yellow but darker apically (
Figure 3A). Sensory organs on antennal segments III and IV linear and directed longitudinally (
Figure 3A). Anterior and posterior margin of pronotum without long setae. Forewings present; forewings with one dark transverse band and posterior border dark from the middle and extending almost all the way to wing apex; apical ring vein as pale as wing membrane (
Figure 5B).
Mainly found on
Pinus and probably a predator of small arthropods such as mites [
9,
25]. The distribution is Holarctic, in Fennoscandia in Sweden, Norway, Denmark, and Finland. The Swedish distribution includes Skåne, Östergötland, Södermanland, Uppland, Värmland, Västerbotten, and Norrbotten. Here also reported from Medelpad.
| Genus Franklinothrips Back, 1912 [43] |
| Franklinothrips vespiformis (Crawford, 1909) [44] |
| Figure 3C. |
| Aeolothrips vespiformis Crawford, 1909: 109 [44]. |
RÉUNION (FRANCE); 1♀; Piteon de Grande Anse; 20 Jan. 1989; in low roadside vegetation; H. G. Müller leg.; R. zur Strassen det.; SMF T 15992.
Body (excl. extremities) sharply bicolored with yellow segments. Antennal segment I–III yellow; III eight times as long as wide; IV brown (
Figure 3C). Sensory organs on antennal segments III and IV as irregular blotches (
Figure 3C). Head and thorax mainly dark. Anterior and posterior margin of pronotum without long setae. Forewings present with two dark transverse bands; apical ring vein brown. Abdominal segments II and III yellow.
A neotropical species but has been introduced in several regions. The use of the species as a biological pest control against other thrips species increases the risk of further introductions around the world. Wild populations have not been recorded in Fennoscandia. The climatic requirements of this species make unintentional introduction and reproduction unlikely at northern latitudes, but with climatic changes the risk increases. Recently
F. vespiformis was approved for biological control in Sweden [
45] and therefore the species is included here.
| Genus Rhipidothrips Uzel, 1895 [1] |
| Rhipidothrips brunneus Williams, 1913 [29] |
| Figure 3B, Figure 4C and Figure 7E. |
| Rhipidothrips brunneus Williams, 1913: 216 [29]. |
ITALY; 1♀; Catania, 6 km N of Granieri; 10 May 1984; on fresh green grass; A. Vesmanis leg.; R. zur Strassen det.; SMF T 12861. EGYPT; 1♀; Rhipidothrips cahirensis, PARATYPE; pyramids at canal bank; 27 Mar. 1930; H. Priesner leg.; SMF T 19326.; 11♀♀; Blekinge, Karlskrona kommun, Knösö; N56.167848, E15.664502; 18 May 2023; E. Wahlberg leg.; on Poaceae; voucher IDs: BI2T, BI9T, BJ1T, BJ2T, BJ3T, BJ5T, BJ6T, BJ7T, BJ8T, BJ9T, BK1T; BOLD IDs: SETHY246-23, SETHY252-23, SETHY253-23, SETHY354-23, SETHY255-23, SETHY256-23, SETHY257-23, SETHY258-23, SETHY259-23, SETHY260-23, SETHY261-23; NHRS.; 3♀♀; Blekinge, Karlskrona kommun, Sandhamn; N56.090980, E15.856533; 31 May 2023; E. Wahlberg leg.; in mixed herbaceous vegetation; voucher IDs: BL4T, BL5T, BT9T; BOLD IDs: SETHY273-23, SETHY274-23, SETHY325-23; NHRS.
Body (excl. extremities) wholly dark. Antennal segment I–II brown; III–IV mainly yellow (
Figure 3B). Sensory organs on antennal segments III and IV lens-shaped (
Figure 3B). Interocellar setae prominent and long (at least 20 μm) and lateral setae posterior to eyes strong (
Figure 4C). Anterior margin of pronotum with one pair of stout but short angular bristles; posterior margin with two to three pairs of stout but short setae; posteroangular setae strong and dark (
Figure 7E). Forewings sometimes present but often brachypterous, pale; apical ring vein pale as wing membrane.
Mainly found on grasses and has a widespread Palaearctic distribution [
24,
25] in south to Egypt and Iran. Also found in North America and Australia, and possibly introduced there [
46]. In Fennoscandia recorded in Finland and Sweden; in Sweden, found in Blekinge in 2022 [
31].
FINLAND; 1♀; SYNTYPE (macropterous); Pargas, Lofsdal, garden; O. M. Reuter leg.; on Abies; MZF GV.33091; Spec. typ. No. 6058.; 1♀; SYNTYPE (brachypterous); Pargas, Lenholmen; on Covallaria; O. M. Reuter leg.; MZF GV.33092; Spec. typ. No. 6059.; 1♀; SYNTYPE (macropterous); Helsingin maalaiskunta, Vantaa; B. Wasastjärna leg.; MZF GX.6325; Spec. typ. No. 6060. SWEDEN; 1♀; Uppland, Stockholm, Experimentalfältet; 19 May 1919; on Phleum; O. Ahlberg leg.; MZLU 00109507.; 4♀; Uppland, Stockholm, Experimentalfältet; 19 May 1919; on Phleum; O. Ahlberg leg.; NHRS.
Similar to
R. brunneus with body dark brown with the tips of the tibia and whole tarsi yellow, brown antennal segments I, II, VI–IX, segments III and IV yellow (
Figure 3I, 6F). Antennal segment V yellow to bright brown (
Figure 3I) in contrast to the darker
R. brunneus (
Figure 3B). Head striated, with stronger striations posteriorly (
Figure 4D), but not as heavy as in
R. brunneus (
Figure 4C). Laterodorsal bristles behind eyes present, but weak and bright (
Figure 4D). Interocellar bristles present, at most 20 μm long (
Figure 4D). Posteroangular bristle present on pronotum, stout but pale in colour (
Figure 7F). Most often macropterous (
Figure 6F), and brachypterous specimens are rare.
Body (excl. extremities) wholly dark (
Figure 6F). Antennal segment I–II brown; III–IV mainly yellow, V yellow to bright brown (
Figure 3I). Sensory organs on antennal segments III and IV lens-shaped (
Figure 3I). Head transversely striated with stronger striations posteriorly, forming a weakly defined collar, lateral sides of the head straight (
Figure 4D). Eyes elongate and extend posteriorly on the ventral side. Dorsal setae on head all weak except for the interocellar setae that are stout but short, lateral setae posterior to eyes weak (
Figure 4D). Pronotum transversely striated, anterior margin with one pair of stout but short angular bristles; posterior margin with two to three pairs of stout but short setae; posteroangular setae present, stout and yellow (
Figure 7F). Forewings often present, but sometimes brachypterous, wings very pale to white; apical ring vein pale as wing membrane (
Figure 6F). Mesonotum transversely striated, metanotum longitudinally striated. Abdominal tergites transversely striated; I with 1 pair, II–VII with three pairs of weak median bristles; one to two pairs of lateral bristles on II–VII; sternites II–VII similarly striated but with three pairs of weak setae posteriomarginally. Abdominal segment IX striated with two pairs of strong posteromarginal dorsal bristles; the lateral pair long and median short; three pairs of lateral strong setae, the posterior pair shorter. Terminal segment X with three pairs of strong bristles situated slightly posterior to medially, with the lateral setae the longest.
The characteristics separating this species from other species in
Rhipidothrips was, according to Reuter, the lack of pronotal posteroangular setae [
28], but these are present on the type specimens but are yellow. This was also noted by Bailey, who added the interocellar setae as a defining character separating
R. niveipennis and
R. brunneus [
24]. In
R. niveipennis these are weak and short. Rarely collected, described from Finland but recorded in Norway, Sweden, and France. Type material collected on
Convallaria majalis, but also found on grasses and different flowering plants [
24]. The only Swedish record is from Uppland.
| Family Melanthripidae Bagnall, 1913 [2] |
The Melanthripidae family are, like the Aeolothripidae, characterized by having nine antennal segments but which are never fused terminally and the sensorial organs on the antennae are elongate and transverse (
Figure 3D).
Melanthrips fuscus, the only species in Sweden, is macropterous as are all European species. Characterized by having long bristles on the head and pronotum, as well as strong bristles on wings. Biology little studied, but probably feeds on plant tissue.
| Genus Melanthrips Haliday, 1836 [3] |
| Melanthrips fuscus (Sulzer, 1776) [47] |
| Figure 3D. |
| Thrips fuscus Sulzer, 1776: 113 [47]. |
SWEDEN; 1♀; Uppland, Rydbo; 30 June 1919; on Triticum aestivum; O. Ahlberg, leg.; MZLU 00109506.; 2♀♀; Skåne, Alnarp; May 1899; on Euphorbia palustris; F. Trybom leg; NHRS.
Body (excl. extremities) wholly dark. Antennal segments all more or less brown (
Figure 3D). Sensory organs on antennal segments III and IV linear and directed transversely (
Figure 3D). Anterior margin of pronotum with two pairs and posterior with at least four pairs of well-developed and long bristles; 1 pair of well-developed and strong posteroangular bristles. Forewings present; mostly shaded, paler at base, never with pale round patterns; apical ring vein of brown.
Found mainly in different species of Brassicaceae [
6], but not exclusively, and is Palearctic in its distribution with records in south towards northern Africa. In Fennoscandia recorded in Sweden and Denmark, and in Sweden in Skåne, Östergötland, Södermanland, and Uppland.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}