
Molecular Discrimination for Two Anadenanthera Species of Seasonally Dry Tropical Forest Remnants in Brazil

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
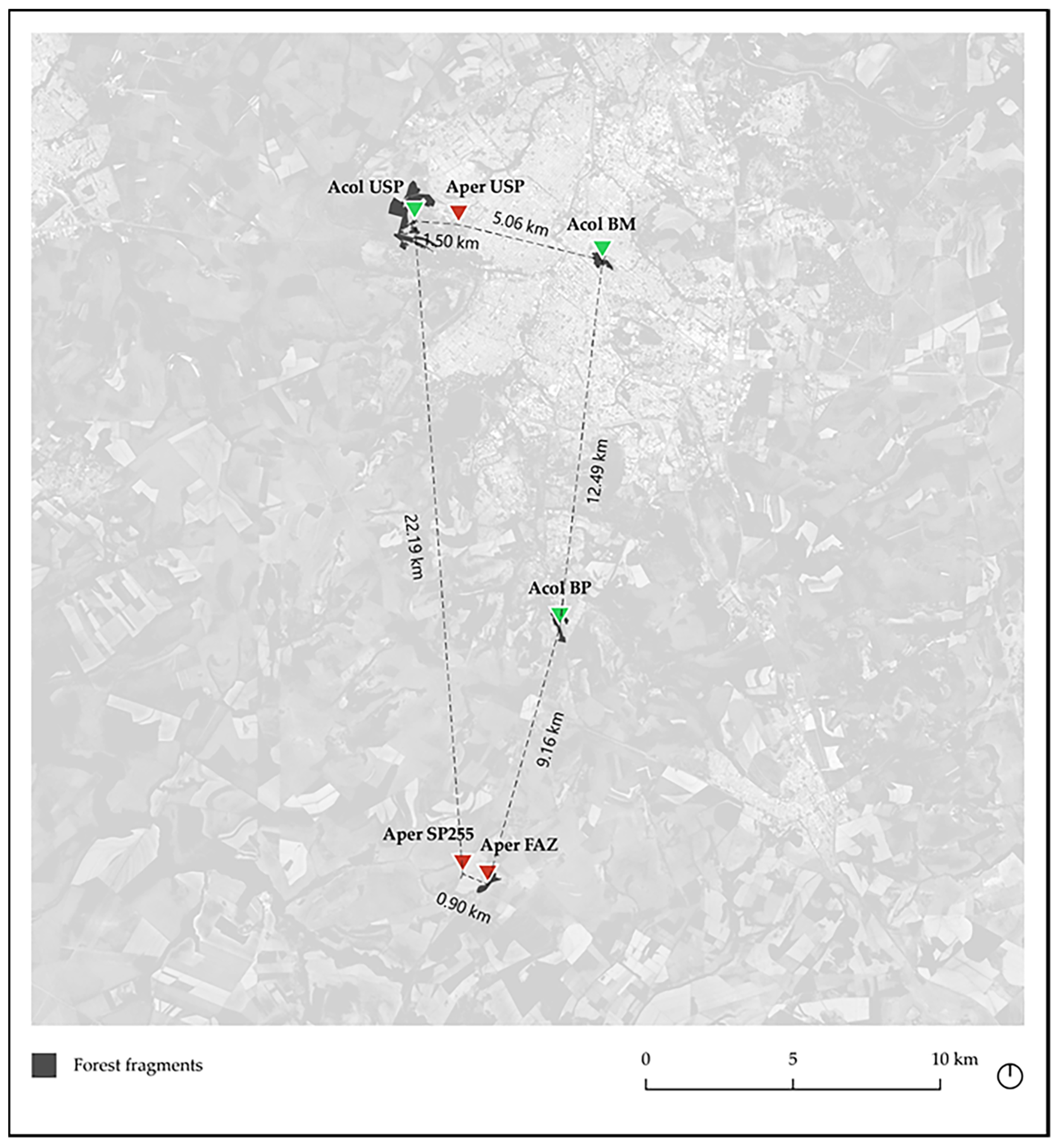
2.1. Identification and Sampling of Anadenanthera Populations
2.2. DNA Extraction, Amplification, and Electrophoresis
2.3. Potential ISSR Species-Specific Markers
2.4. Statistical Analyses of Genetic Diversity
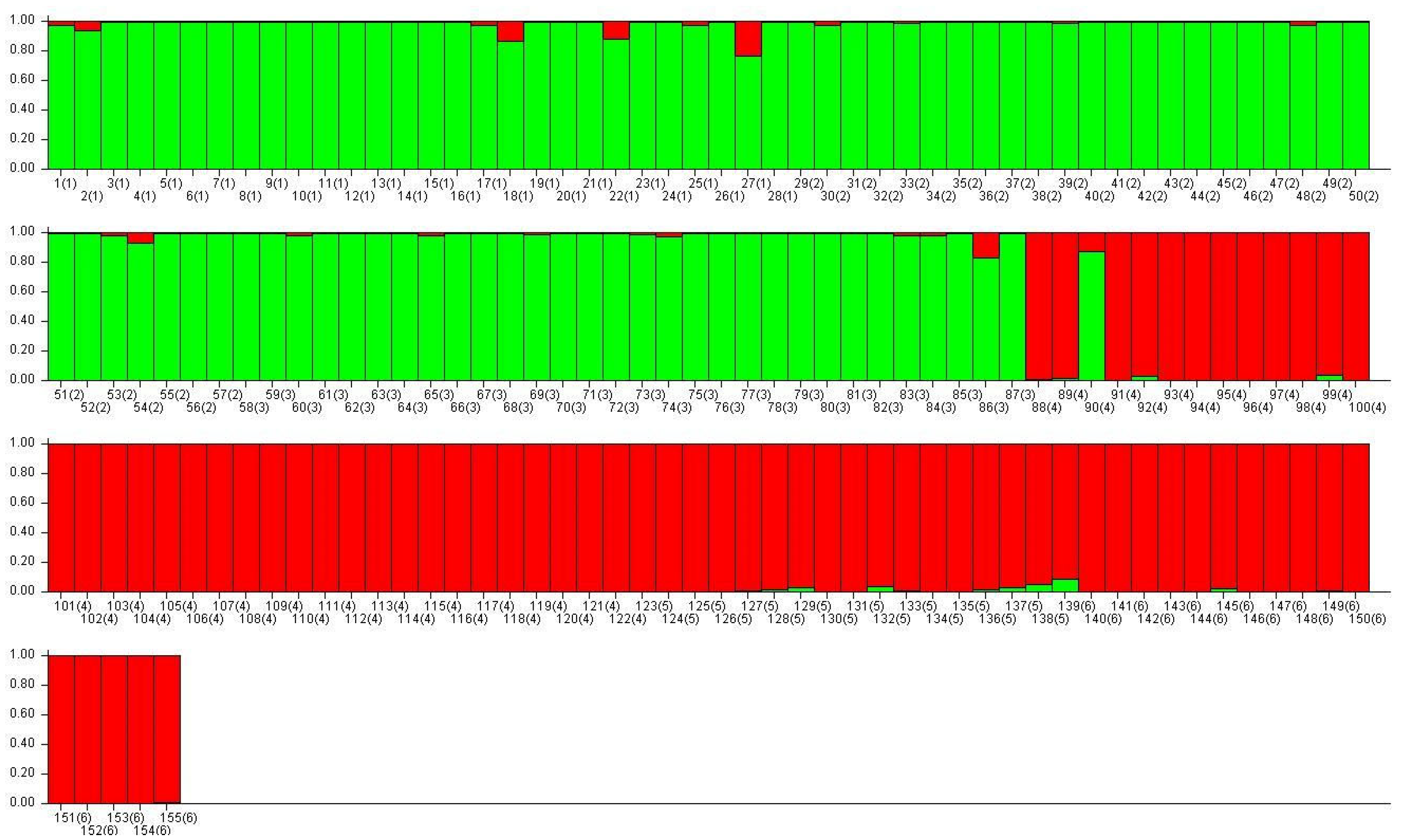
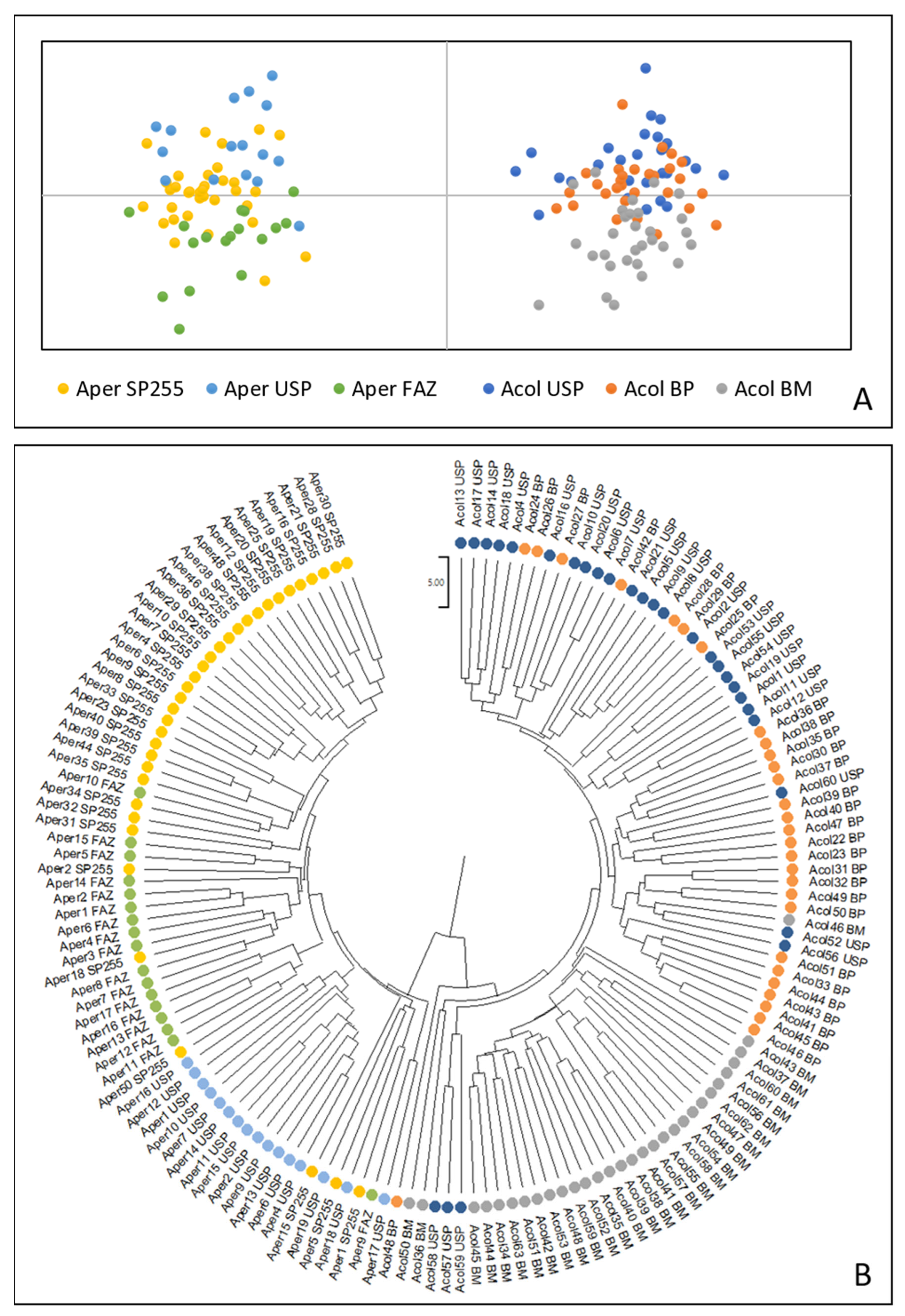
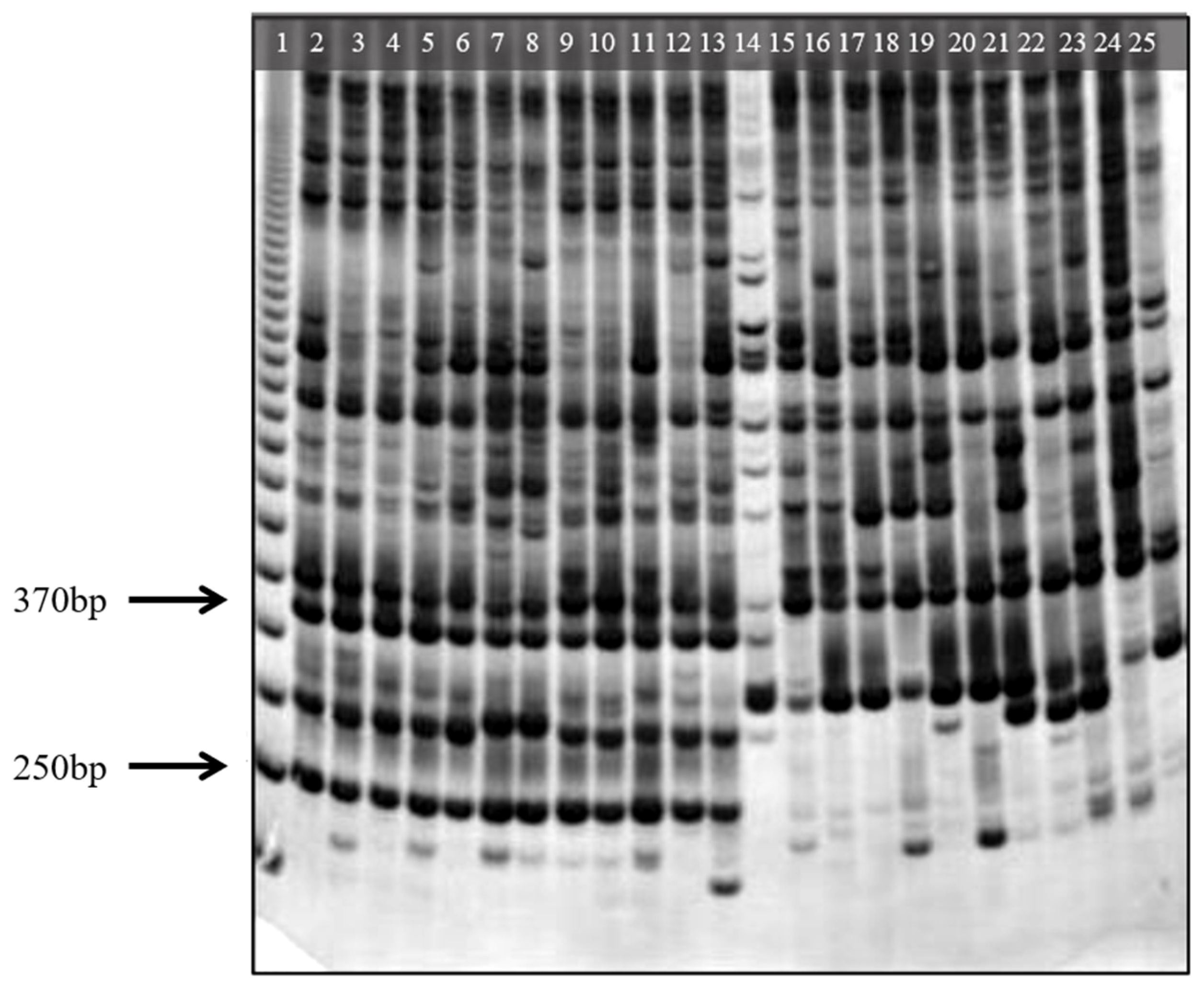
3. Results
3.1. Analysis of Ancestry and Clusters
3.2. Gst Analysis and Genetic Distances
3.3. Genetic Diversity among Populations of Anadenanthera colubrina and A. peregrina
3.4. Species-Specific Molecular Marker Analysis
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vieira, D.L.M.; Scariot, A. Principles of Natural Regeneration of Tropical Dry Forests for Restoration. Restaur. Ecol. 2006, 14, 11–20. [Google Scholar] [CrossRef]
- Pennington, R.T.; Lewis, G.P.; Ratter, J.A. An Overview of the Plant Diversity, Biogeography and Conservation of Neotropical Savannas and Seasonally Dry Forests. In Neotropical Savannas and Seasonally Dry Forests: Plant Diversity, Biogeography, and Conservation; Pennington, R.T., Lewis, G.P., Ratter, J.A., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 1–29. [Google Scholar] [CrossRef]
- Altschul, S.R. A taxonomic study of the genus Anadenanthera. Contrib. Gray Herb. Harv. Univ. 1964, 193, 3–65. Available online: http://www.jstor.org/stable/41764816 (accessed on 8 August 2023).
- Kotchetkoff-Henriques, O.; Joly, C.A.; Bernacci, L.C. Soil and floristic composition of native vegetation remnants relationship in the municipality of Ribeirão preto, sp. Braz. J. Bot. 2005, 28, 541–562. [Google Scholar] [CrossRef]
- Carvalho, P.E.R. Angico-Branco. Embrapa—Circular Técnica 56. Colombo. PR Novembro. 2002. Available online: http://www.infoteca.cnptia.embrapa.br/bitstream/doc/306306/1/CT0056.pdf (accessed on 8 August 2023).
- Feres, J.M.; Nazareno, A.G.; Borges, L.M.; Guidugli, M.C.; Bonifacio-Anacleto, F.; Alzate-Marin, A.L. Depicting the mating system and patterns of contemporary pollen flow in trees of the genus Anadenanthera (Fabaceae). PeerJ 2021, 9, e10579. [Google Scholar] [CrossRef] [PubMed]
- Lorenzi, H. Árvores Brasileiras—Manual de Identificação e Cultivo de Plantas Arbóreas Nativas no Brasil; Instituto Plantarum: Nova Odessa, Brazil, 2002; Volumes 2–3, p. 368. [Google Scholar]
- Weber, C.R.; Soares, C.M.L.; Lopes, A.B.D.; Silva, T.S.; Nascimento, M.S.; Ximenes, E. Anadenanthera colubrina: A therapeutic potential study. Ver. Bras. Farm. 2011, 92, 235–244. [Google Scholar]
- Chaves, T.L.; Ricardo, L.; Paula-Souza, J.; Brandão, M.G.L. Useful Brazilian plants under the view of the writer-naturalist João Guimarães Rosa. Rev. Bras. Farmacogn. 2015, 25, 437–444. [Google Scholar] [CrossRef][Green Version]
- Durigan, G.; Silveira, E.R. Riparian forest restoration in cerrado, Assis, SP, Brazil. Sci. For. 1999, 56, 135–144. Available online: http://www.ipef.br/publicacoes/scientia/nr56/cap10.pdf (accessed on 29 March 2023).
- Barrandeguy, M.E.; García, M.V.; Prinz, K.; Pomar, R.; Finkeldey, R. Genetic structure of disjunct Argentinean populations of the subtropical tree Anadenanthera colubrina var. cebil (Fabaceae). Plant Syst. Evol. 2014, 300, 1693–1705. [Google Scholar] [CrossRef]
- Barrandeguy, M.E.; Mogni, V.; Zerda, H.; Savino, C.; Dusset, F.; Prado, D.E.; García, M.V. Understanding the spatio-temporal dynamics of Anadenanthera colubrina var. cebil in the biogeographical context of Neotropical seasonally dry forest. Flora 2022, 295, 152149. [Google Scholar] [CrossRef]
- Cortelete, M.A.; Silva Júnior, A.L.; Pereira, M.L.S.; Miranda, F.D.; Caldeira, M.V.W. Molecular characterization as strategy for ex situ conservation of Anadenanthera peregrina (L.) Speg. Sci. For. 2021, 49, e3443. [Google Scholar] [CrossRef]
- García, M.V.; Barrandeguy, M.E.; Prinz, K. Contemporary climate influence on variability patterns of Anadenanthera colubrina var. cebil, a key species in seasonally dry tropical forests. J. For. Res. 2022, 33, 89–101. [Google Scholar] [CrossRef]
- Mangaravite, E.; Silveira, T.C.; Vinson, C.C.; Bueno, M.L.; Silva, R.S.; Carniello, M.A.; Veldman, J.W.; Garcia, M.G.; Oliveira, L.O. Unlocking the secret diversity of Anadenanthera: Insights from molecular genetics of four evolving species. Bot. J. Linn. Soc. 2023, 204, boad037. [Google Scholar] [CrossRef]
- Viana, M.L.; Giamminola, E.; Russo, R.; Ciaccio, M. Morphology and genetics of Anadenanthera colubrina var. cebil (Fabaceae) tree from Salta (Northwestern Argentina). Rev. Biol. Trop. 2014, 62, 757–767. Available online: https://www.redalyc.org/articulo.oa?id=44931383029 (accessed on 8 August 2023). [CrossRef][Green Version]
- Zietkiewicz, E.; Rafalski, A.; Labuda, D. Genome fingerprinting by simple se-quence repeat (SSR)-anchored polymerase chain reaction amplification. Genomics 1994, 20, 176–183. [Google Scholar] [CrossRef]
- Reddy, M.P.; Sarla, M.; Siddiq, E.A. Inter simple sequence repeat (ISSR) poly-morphism and its application in plant breeding. Euphytica 2002, 128, 9–17. [Google Scholar] [CrossRef]
- Tahir, N.A.-R.; Lateef, D.; Rasul, K.S.; Aziz, R. Assessment of genetic variation and population structure in Iraqi barley accessions using ISSR, CDDP, and SCoT markers. Czech, J. Genet. Plant Breed. 2023, 59, 148–159. [Google Scholar] [CrossRef]
- Uddin, M.S.; Cheng, Q. Recent Application of Biotechniques for the Improvement of Mango Research; Poltronieri, P., Hong, Y., Eds.; Applied Plant Genomics and Biotechnology, Woodhead Publishing: Cambridge, UK, 2015; pp. 195–212. [Google Scholar] [CrossRef]
- Bornet, B.; Branchard, M. Nonanchored Inter Simple Sequence Repeat (ISSR) Markers: Reproducible and Specific Tools for Genome Fingerprinting. Plant Mol. Biol. Report 2001, 19, 209–215. Available online: https://link.springer.com/content/pdf/10.1007/BF02772892.pdf (accessed on 8 August 2023). [CrossRef]
- Alzate-Marin, A.L.; Costa-Silva, C.; Rivas, P.M.S.; Bonifacio-Anacleto, F.; Santos, L.G.; Moraes Filho, R.M.; Martinez, C.A. Diagnostic fingerprints ISSR/SSR for tropical leguminous species Stylosanthes capitata and Stylosanthes macrocephala. Sci. Agric. 2020, 77, e20180252. [Google Scholar] [CrossRef]
- Araújo, F.D.S.; Felix, F.C.; Ferrari, C.D.S.; Vieira, F.D.A.; Pacheco, M.V. Seed quality and genetic diversity of a cultivated population of Mimosa caesalpiniifolia BENTH. Rev. Caatinga 2020, 33, 1000–1006. [Google Scholar] [CrossRef]
- Silva, A.V.C.D.; Amorim, J.A.E.; Vitória, M.F.D.; Ledo, A.D.S.; Rabbani, A.R.C. Characterization of trees, fruits and genetic diversity in natural populations of mangaba. Agric. Sci. 2017, 41, 255–262. [Google Scholar] [CrossRef]
- Fajardo, C.G.; Silva, R.A.R.; Chagas, K.P.T.; Vieira, F.A. Genetic and phenotypic association of the carnauba palm tree evaluated by inter-simple sequence repeat and biometric traits. Genet. Mol. Res. 2018, 17, gmr18018. [Google Scholar] [CrossRef]
- Lopes, J.S.; Costa, M.R.J.; Arriel, D.A.A. Genetic diversity of potential mother trees of Myracrodruon urundeuva Allemão in a remnant population from Brazilian Cerrado using ISSR. Adv. For. Sci. 2020, 7, 1017–1024. [Google Scholar] [CrossRef]
- Santos, A.R.; Gonçalves, E.O.; Silva Júnior, A.L.; Gibson, E.L.; Araújo, E.F.; Mi-randa, F.D.; Caldeira, M.V.W. Diversity and genetic structure in a mini-garden of Dalbergia nigra: A tree threatened with extinction in the Atlantic Forest. Plant Gene 2021, 27, 100304. [Google Scholar] [CrossRef]
- Moraes Filho, R.M.; Bonifacio-Anacleto, F.; Alzate-Marin, A.L. Fragmentation effects and genetic diversity of the key semidecidual forest species Metrodorea nigra in Southwestern Brazil. Genet. Mol. Res. 2015, 14, 3509–3524. [Google Scholar] [CrossRef] [PubMed]
- Alzate-Marin, A.L.; Guidugli, M.C.; Soriani, H.H.; Martinez, C.A.; Mestriner, M.A. An efficient and rapid DNA minipreparation procedure suitable for PCR/SSR and RAPD analyses in tropical forest tree species. Braz. Arch. Biol. Technol. 2009, 52, 1217–1224. [Google Scholar] [CrossRef]
- Sanguinetti, C.J.; Dias, E.N.; Simpson, A.J.G. Rapid silver staining and recovery of PCR products separated on polyacrylamide gels. Biotechniques 1994, 17, 914–921. [Google Scholar] [PubMed]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. Available online: http://www.genetics.org/content/genetics/155/2/945.full.pdf (accessed on 15 November 2022). [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 586–590. Available online: http://www.genetics.org/content/genetics/89/3/583.full.pdf (accessed on 17 November 2022). [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Nei, M. Analysis of gene diversity in subdivided populations. Proc. Natl. Acad. Sci. USA 1973, 70, 3321–3323. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC427228/ (accessed on 17 November 2022). [CrossRef] [PubMed]
- Yeh, F.C.; Yang, R.C.; Boyle, T. POPGENE, Version 1.31. Microsoft Windows-Based Freeware for Population Genetic Analysis. University of Alberta/CIFOR: Edmonton, Alberta, 1999. Available online: https://sites.ualberta.ca/~fyeh/popgene.pdf (accessed on 17 November 2022).
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 178kb. Available online: https://palaeo-electronica.org/2001_1/past/past.pdf (accessed on 20 November 2022).
- Hamrick, J.L.; Godt, M.J.W. Sherman-Broyles, S.L. Factors influencing levels of genetic diversity in woody plant Species. New For. 1992, 6, 95–124. Available online: https://link.springer.com/content/pdf/10.1007/BF00120641.pdf (accessed on 3 August 2023). [CrossRef]
- Borges, L.A.; Machado, I.C.; Lopes, A.V. Bee pollination and evidence of substitutive nectary in Anadenanthera colubrina (Leguminosae-Mimosoideae). Arthropod-Plant Interact. 2017, 11, 263–271. [Google Scholar] [CrossRef]
- Hamrick, J.L.; Godt, M.J.W. Effects of life history traits on genetic diversity in plant species. Phil. Trans. R. Soc. Lond. 1996, 351, 1291–1298. [Google Scholar] [CrossRef]
- Culley, T.M.; Wallace, L.E.; Gengler-Nowak, K.M.; Crawford, D.J. A comparison of two methods of calculating GST, a genetic measure of population differentiation. Am. J. Bot. 2002, 89, 460–465. [Google Scholar] [CrossRef] [PubMed]
- Reis, M.S. Dinâmica da movimentação dos alelos: Subsídios para conservação e manejo de populaçõeses naturais em plantas. Braz. J. Genet. 1996, 19, 37–47. [Google Scholar]
- Prado, D.E.; Gibbs, P.E. Patterns of species distribution in the dry seasonal forests of South America. Ann. Miss. Bot. Gard. 1993, 80, 902–927. [Google Scholar] [CrossRef]
- Oliveira-Filho, A.T.; Curi, N.; Vilela, E.A.; Carvalho, D.A. Effects of Canopy Gaps, Topography, and Soils on the Distribution of Woody Species in a Central Brazilian Deciduous Dry Forest. Biotropica 1998, 30, 362–375. Available online: http://www.jstor.org/stable/2389121 (accessed on 20 September 2023). [CrossRef]
- Miranda, C.C.; Souza, D.M.S.; Manhone, P.R.; Oliveira, P.C.; Breier, T.B. Germinação de sementes de Anadenanthera peregrina (L.) Speg. com diferentes substratos em condições laboratoriais. Floresta e Ambiente 2012, 19, 26–31. Available online: http://www.floram.org/files/v19n1/v19n1a4.pdf (accessed on 20 September 2023). [CrossRef]
- Castro, L.E.; Guimarães, C.C.; Faria, J.M.R. Physiological, cellular and molecular aspects of the desiccation tolerance in Anadenanthera colubrina seeds during germination. Braz. J. Biol. 2017, 77, 4–780. [Google Scholar] [CrossRef]
- Silva, L.M.B.; Barbosa, D.C.A. Growth and survival of Anadenanthera macrocarpa (Benth.) Brenan (Leguminosae), in an area of caatinga, Alagoinha, PE. Acta Bot. Bras. 2000, 14, 251–261. [Google Scholar] [CrossRef]
- Silva, R.M.G.; Saraiva, T.S.; Silva, R.B.; Gonçalves, L.A.; Silva, L.P. Potencial alelopático de extrato etanólico de Anadenanthera macrocarpa e Astronium graveolens. Biosci. J. 2010, 26, 632–637. [Google Scholar]
- Alzate-Marin, A.L.; Bonifacio-Anacleto, F.; Moraes Filho, R.M.; Machado, G.P.; Nazareno, A.G. Genetic analysis across life stages of Metrodorea nigra (Rutaceae) in a population located in an urban landscape of Southeastern Brazil using a new set of microsatellite markers. Braz. J. Bot. 2016, 39, 795–799. [Google Scholar] [CrossRef]
- Alzate-Marin, A.L.; Ferreira-Ramos, R.; Guidugli, M.; Martinez, C.A.; Mestriner, M.A. Genetic diversity assessed in individuals of Aspidosperma polyneuron and Cariniana estrellensis used as seed donors in an forest gene bank. BMC Proc. 2011, 5 (Suppl. 7), 8. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence 5′–3′ | AT °C | Cycles Number | |
|---|---|---|---|---|
| 1 | #UBC2 | GAGAGAGAGAGAGAGAT | 50 | 35 |
| 2 | #UBC820 | GTGTGTGTGTGTGTGTC | 50 | 35 |
| 3 | #UBC851 | GTGTGTGTGTGTGTGTYG | 50 | 35 |
| 4 | #UBC858 | TGTGTGTGTGTGTGTGRT | 50 | 35 |
| 5 | #UBC862 | AGCAGCAGCAGCAGCAGC | 55 | 30 |
| 6 | #UBC864 | ATGATGATGATGATGATG | 50 | 35 |
| 7 | #UBC866 | CTCCTCCTCCTCCTCCTC | 50 | 35 |
| 8 | #UBC885 | BHBGAGAGAGAGAGAGA | 50 | 35 |
| 9 | #UBC886 | VDVCTCTCTCTCTCTCT | 55 | 30 |
| 10 | #UBC897 | CCGACTCGAGNNNNNNATGTGG | 50 | 30 |
| Populations | GST | Nm |
|---|---|---|
| Acol + Aper | 0.143 | 3.0 |
| Acol | 0.076 | 6.1 |
| Aper | 0.102 | 4.4 |
| Populations | ||||||
|---|---|---|---|---|---|---|
| Acol USP | Acol BP | Acol BM | Aper SP255 | Aper USP | Aper FAZ | |
| Acol USP | 0.000 | |||||
| Acol BP | 0.024 | 0.000 | ||||
| Acol BM | 0.057 | 0.053 | 0.000 | |||
| Aper SP255 | 0.183 | 0.190 | 0.174 | 0.000 | ||
| Aper USP | 0.189 | 0.203 | 0.186 | 0.040 | 0.000 | |
| Aper FAZ | 0.187 | 0.202 | 0.175 | 0.044 | 0.060 | 0.000 |
| Population | N | LN | PPL % | NA | h |
|---|---|---|---|---|---|
| Acol USP | 28 | 101 | 87.04 | 1.70 (0.05) | 0.30 (0.02) |
| Acol BP | 29 | 97 | 83.33 | 1.81 (0.06) | 0.29 (0.02) |
| Acol BM | 30 | 97 | 84.26 | 1.74 (0.06) | 0.30 (0.02) |
| Average | 98 | 84.87 (1.100) | 1.75 (0.030) | 0.30 (0.004) * | |
| Aper USP | 16 | 92 | 75.93 | 1.61 (0.07) | 0.26 (0.02) * |
| Aper FAZ | 17 | 92 | 77.78 | 1.63 (0.07) | 0.23 (0.02) |
| Aper SP255 | 35 | 98 | 84.26 | 1.84 (0.05) * | 0.24 (0.02) |
| Average | 94 | 79.32 (2.500) | 1.69 (0.074) | 0.25 (0.010) * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonifácio-Anacleto, F.; Moraes Filho, R.M.; Borges, L.M.; Martinez, C.A.; Alzate-Marin, A.L. Molecular Discrimination for Two Anadenanthera Species of Seasonally Dry Tropical Forest Remnants in Brazil. Taxonomy 2024, 4, 150-162. https://doi.org/10.3390/taxonomy4010008
Bonifácio-Anacleto F, Moraes Filho RM, Borges LM, Martinez CA, Alzate-Marin AL. Molecular Discrimination for Two Anadenanthera Species of Seasonally Dry Tropical Forest Remnants in Brazil. Taxonomy. 2024; 4(1):150-162. https://doi.org/10.3390/taxonomy4010008
Chicago/Turabian StyleBonifácio-Anacleto, Fernando, Rômulo Maciel Moraes Filho, Leonardo Maurici Borges, Carlos Alberto Martinez, and Ana Lilia Alzate-Marin. 2024. "Molecular Discrimination for Two Anadenanthera Species of Seasonally Dry Tropical Forest Remnants in Brazil" Taxonomy 4, no. 1: 150-162. https://doi.org/10.3390/taxonomy4010008
APA StyleBonifácio-Anacleto, F., Moraes Filho, R. M., Borges, L. M., Martinez, C. A., & Alzate-Marin, A. L. (2024). Molecular Discrimination for Two Anadenanthera Species of Seasonally Dry Tropical Forest Remnants in Brazil. Taxonomy, 4(1), 150-162. https://doi.org/10.3390/taxonomy4010008